








-
Juices have gradually expanded their market share in the food industry due to their natural health benefits, fresh nutrition, and unique flavors. The annual growth rate is predicted to reach 5.27% by 2028, playing a significant role in people's diets[1,2]. Since the acidity of most juices ranges from pH 2.4 to 4.6, pasteurization is generally sufficient to eliminate most non-sporulating bacteria, and the product is often considered 'commercially sterile'[3]. However, over the past three decades, several cases of spoilage have occurred in commercially pasteurized juices, leading to significant contamination issues and substantial economic losses within the juice industry[4−6]. Subsequent investigations revealed that spoilage occurred before the end of the shelf life, with Alicyclobacillus acidoterrestris spores isolated from the spoiled juice[7]. Since then, A. acidoterrestris has become one of the most serious threats and primary quality control target organisms in the global juice industry[8−10].
A. acidoterrestris grows within 25–65 °C, with an optimal growth temperature between 42–50 °C and an optimal pH of 3.5–4.5[11,12]. Unlike other neutral and acid-tolerant Bacillus species, A. acidoterrestris is particularly acidophilic and heat-resistant[13]. The high heat and acid resistance of these spores stems from their unique cell membrane composition and architecture. The membrane is rich in ω-cyclohexane fatty acids and hopanoids, whose tightly packed lipids form a robust hydrophobic barrier[14,15]. This structure enhances membrane stability and reduces permeability, allowing the spores to withstand acidic and high-temperature environments[16]. These spores are capable of surviving pasteurization in acidic juices, and they can germinate and return to a vegetative state once conditions become favorable[17]. During this process, they produce off-flavors, such as guaiacol, leading to spoilage, and deterioration of the juice[18]. To effectively inactivate A. acidoterrestris spores, extremely high temperatures (5 log reduction at 115 °C/8 s) are required[12,19]. However, such extreme heat can severely damage the functional nutrients, color, and flavor of the juice, leading to significant quality degradation[16]. Therefore, to ensure both the high quality and the stable shelf-life of acidic juices, developing a more efficient and gentler sterilization method, with A. acidoterrestris spores as an indicator, is crucial[20].
The 'germination-inactivation' strategy aims to artificially induce spore germination, thereby reducing their inherent resistance, by applying application of milder inactivation treatments such as pasteurization[21,22]. A comprehensive understanding of spore germination behavior is therefore essential for the successful application of this approach. For Bacillus spores, germination can be triggered by nutrients like glucose, amino acids, or the mixture AGFK, as well as non-nutrient substances like dodecylamine (DDA) and CaDPA[23,24]. Additionally, high hydrostatic pressure (HHP), an emerging non-thermal food sterilization technology[25], can effectively induce spore germination[26]. It has been proposed that HHP can induce spore germination either by activating germinant receptors at moderate pressure (50–300 MPa) or opening CaDPA channels at very high pressure (400–600 MPa), thereby rendering the 'germination-inactivation' strategy potentially applicable[27]. As for A. acidoterrestris spores, although several studies have examined A. acidoterrestris spore germination[28−30], systematic investigations into its germination behavior remain limited, and the underlying mechanisms are not fully elucidated. This knowledge gap constrains the effective implementation of the 'germination-inactivation' strategy for controlling A. acidoterrestris spores in acidic food systems. Moreover, superdormant (SD) spores pose a major challenge to the effective implementation of the 'germination-inactivation' strategy. These spores, defined by their markedly delayed or incomplete germination in response to germinants, constitute a subpopulation with enhanced resilience[31]. Notably, emerging evidence indicates that SD spores isolated following HHP treatment can undergo spontaneous germination[32], which can promote the better application of the 'germination-inactivation' strategy. However, it remains unclear whether SD spores of A. acidoterrestris under HHP demonstrate a similar capacity for spontaneous germination.
In this study, a comprehensive analysis of the germination behavior of A. acidoterrestris spores across a wide pH range (pH 2–9) with various germination stimuli, including L-alanine, AGFK, and HHP was performed. Key parameters influencing spore germination were systematically optimized, including pH conditions, nutrient germinant concentrations, heat activation temperature and duration, incubation temperature, and the pH of the heat activation medium. The study also reveals the instability of A. acidoterrestris SD spores following HHP treatment, characterized by the spontaneous germination of them. Building on these findings, an effective inactivation strategy was developed using HHP treatment at 30 °C followed by incubation and mild pasteurization, achieving 5.0 ± 0.40 to 5.3 ± 0.51 log reductions in spore viability in acidic systems.
-
The following chemical reagents were used in this study: 53 mM K-Hepes buffer, 5 mM TbCl3, 100 mM L-ala, 10 x AGFK (containing 2.5 mM L-asparagine, 5 mg/mL D-glucose, 5 mg/mL D-fructose, and 50 mM KCl), 0.01M PBS (pH 7.2–7.4), and 2.5 M H2SO4. All these chemicals and reagents were purchased from Sigma.
Bacteria and spore production
-
The bacteria used in this research were the wild-type A. acidoterrestris ATCC 49025, purchased from China Center of Industrial Culture Collection, and the wild-type B. subtilis PY79. All standard methods for A. acidoterrestris were conducted as previously described[33]. Spores were induced by plating bacterial cultures on BAT solid medium at 45 °C. After 24 h of incubation at 45 °C, 200 μL of the culture was spread on BAT agar and incubated again at 45 °C. Every 24 h, spore growth was observed under a phase contrast microscope. After 48 h, when 90% of the field appeared as phase-bright spores, the spores were harvested from the surface of the solid medium using a cell scraper and transferred to sterile double-distilled water (SDDW). Pellets were then washed three times by centrifugation at 4 °C. The harvested spores were resuspended in SDDW and continuously stirred. After 7 d of washing, spores were resolubilized in 20% Nycodenz. Two hundred μL of the suspension was placed onto 900 µL 44% Nycodenz. The spore suspension was centrifuged at 12,000 rpm/20 min/4 °C. This purification step was repeated at least three times. After clearing the supercolumn, the spore was washed at least five times with SDDW. Finally, the purity of the spores was confirmed by phase contrast microscopy, ensuring > 99% purity, and the spores were stored in SDDW at 4 °C for subsequent experiments. If purity was not achieved, the purification process was repeated.
Additionally, B. subtilis spores utilized in this research were produced in accordance with a previously established method with some modifications[34]. Spores were induced by spreading overnight bacterial cultures on Difco Sporulation Medium solid medium at 37 °C. After 36 h of incubation, spores were washed three times with SDDW at 4 °C. Following a 7-d washing period, 200 μL 20% Nycodenz of the spore suspension was placed atop 900 μL of 50% Nycodenz. With the purification steps performed as described previously.
DPA release measurement to monitor spore germination under different pH conditions
-
DPA release assay was performed as previously described, with relevant modifications[35]. Unless otherwise specified, spores were germinated in 53 mM K-Hepes (pH adjusted with 1 M HCl to pH 2, 3, 4, 5, 6, 7, or 9) at 37 °C, and an optical density (OD600 = 0.5), with 2 μL of 5 mM TbCl3 added to the buffer. For nutrient-induced germination, spores were first subjected to a 30-min heat shock treatment at 75 °C, followed by immediate transfer to ice for 10 min of cooling. The nutrient germinants used included: (i) L-alanine (L-Ala) at concentrations of 0, 0.5, 1, 2, 5, 10, and 20 mM; and (ii) AGFK at concentrations of 0 x, 0.5 x, 1 x, 4 x, 8 x, 12 x, and 15 x (10 x AGFK, containing 2.5 mM L-asparagine, 5 mg/mL D-glucose, 5 mg/mL D-fructose, 50 mM KCl). The spore germination dynamics were monitored by measuring the release of DPA, as previously described. The Tb3+-DPA fluorescence intensity was measured using a Spark 10M (Tecan, Switzerland) microplate reader with an emission wavelength of 545 nm, and an excitation wavelength of 270 nm.
To optimize the heat shock and incubation conditions for spore germination, the heat shock temperatures were adjusted to 75, 70, 65, and 60 °C for 15 min. For heat shock time optimization, spores were treated at the optimal heat shock temperature of 70 °C for 60, 45, 30, 15, 5, and 1 min. The incubation temperature was adjusted to 42, 37, 30, 25, and 18 °C, with heat shock at 70 °C for 15 min. For optimizing the pH during the heat shock treatment and germination measurement, spores were suspended in K-Hepes buffer at pH 4 or pH 7 and subjected to 70 °C/15 min. Then, DPA measurement was conducted at 37 °C in K-Hepes at pH 4 or pH 7. Afterward, the spores were subjected to sterile PBS gradient dilution to determine their germination or viability.
Determination of spore heat resistance
-
The method for determining the heat resistance of A. acidoterrestris spores involves treating spores with OD600 = 0.5 in SDDW at 95, 90, 85, 80, 75, 70, 65 °C for 1 h. After processing, the spores are immediately cooled on ice, followed by serial 10-fold dilutions in sterile PBS buffer. A 100 μL aliquot from an appropriate dilution is spread on BAT agar, then incubated overnight at 45 °C. Each dilution gradient experiment is performed in triplicate.
Determination of spore germination and viability under HHP at different pH levels
-
Samples of A. acidoterrestris and B. subtilis spores were prepared to an OD600 = 0.5 in sterile K-Hepes buffer with pH values of 2, 3, 4, 5, 6, 7, and 9, and sealed in flexible packaging bags, then placed on ice for later use. Unless otherwise specified, all spore samples were treated using a HHP apparatus (SHHP-5L, Shanxi Leadflow Technology Co., Ltd) at 30 °C, 200 or 500 MPa, and treatment durations of 1, 3, 5, 7, and 10 min. After HHP treatment, the spores were immediately transferred to centrifuge tubes and placed on ice. A 420 μL aliquot of the spore suspension was centrifuged at 12,000 rpm for 8 min at 4 °C to collect the supernatant. A 198 μL aliquot of the supernatant (at pH 2, 3, 4, 5, 6, 7, and 9) was mixed with 2 μL of TbCl3. DPA release was measured for each pressure treatment time point. The remaining spore suspension in the centrifuge tube (100 μL) was used for plate counting to test viability as previously described[36].
Spontaneous germination induced by HHP and viability test of the HHP-treated spores
-
Spores of A. acidoterrestris were adjusted to an OD600 of 2 in K-Hepes buffer with pH 4 and pH 7. The spores were treated at 200 MPa or 500 MPa for 3 min at 30 °C. After the HHP treatment, the spores were transferred to ice, and 30 μL of the treated spores were taken into new sterile centrifuge tubes for colony counting to test viability. The remaining spore suspension was immediately centrifuged at 12,000 rpm for 2 min at 4 °C. The spore was reinstated in K-Hepes buffer at pH 4 to an OD600 of 0.5. A 198 μL aliquot of the resuspended spore suspension was mixed with 2 μL of TbCl3, and DPA release was monitored at 37 °C for 30 min. The increase in DPA release over time was used to assess the spontaneous germination of spores post-HHP treatment. After DPA measurement, 50 μL of spore supernatant was added to a to a mixture of 148 μL of K-Hepes followed by 2 μL of TbCl3.
Inactivation of A. acidoterrestris spores by 'germination-inactivation' strategy under pH 4 conditions
-
Spores with an OD600 of 0.5 at pH 4 were first subjected to heat shock at 70 °C for 15 min or HHP treatment at 30 °C for 10 min. Some of the spores were then transferred to 37 °C for incubation to induce germination. After 1 h, colony counting was performed. The remaining spores were subjected to additional treatments: either heat at 80 °C for 30 min or HHP at 500 MPa for 10 min, followed by colony counting to assess spore inactivation. Control groups included untreated spores, spores treated only with 80 °C for 30 min, spores treated only with 200 MPa for 10 min, and spores treated only with 500 MPa for 10 min.
Statistical analysis
-
Graphs were created using GraphPad Prism 10.1.2 software, and data analysis was performed using SPSS software. Significant differences were compared using one-way ANOVA combined with Bonferroni multiple comparison tests (* p < 0.05). Each experiment in the figure included three replicate groups.
-
The pH value is a key factor influencing spore germination[37]. To investigate the optimal pH conditions for spore germination, typical germinants L-Ala and AGFK were used to induce germination of both A. acidoterrestris and B. subtilis spores suspended in K-Hepes buffer at pH 2, 3, 4, 5, 6, 7, and 9. DPA release was monitored to examine the spore germination. In the L-Ala-induced system, A. acidoterrestris spores exhibited the highest DPA release and germination efficiency under acidic conditions (pH 2, 3, 4, 5, and 6), with lower germination efficiency in neutral (pH 7) and alkaline (pH 9) conditions (Fig. 1a). In contrast, B. subtilis spores showed the highest DPA release and germination efficiency at pH 4, 5, 6, and 7, with lower germination efficiency at strongly acidic (pH 2 and 3) and alkaline (pH 9) conditions (Fig. 1b). In the AGFK-induced system, A. acidoterrestris spores showed similar optimal pH condition as with L-Ala, with the highest DPA release and germination efficiency under acidic conditions (pH 2, 3, 4, 5, and 6), and lower efficiency in pH 7 and pH 9 conditions (Fig. 1c). For B. subtilis spores, the highest DPA release and germination efficiency were observed at pH 7 and 9, with lower efficiency in weakly acidic (pH 5 and 6) conditions and almost no germination in strongly acidic conditions (pH 2, 3, 4) (Fig. 1d). The differences in germination rates mentioned above are statistically significant (p < 0.05). These results suggest that acidic conditions promote the germination of A. acidoterrestris spores, while neutral conditions favor the germination of B. subtilis spores.
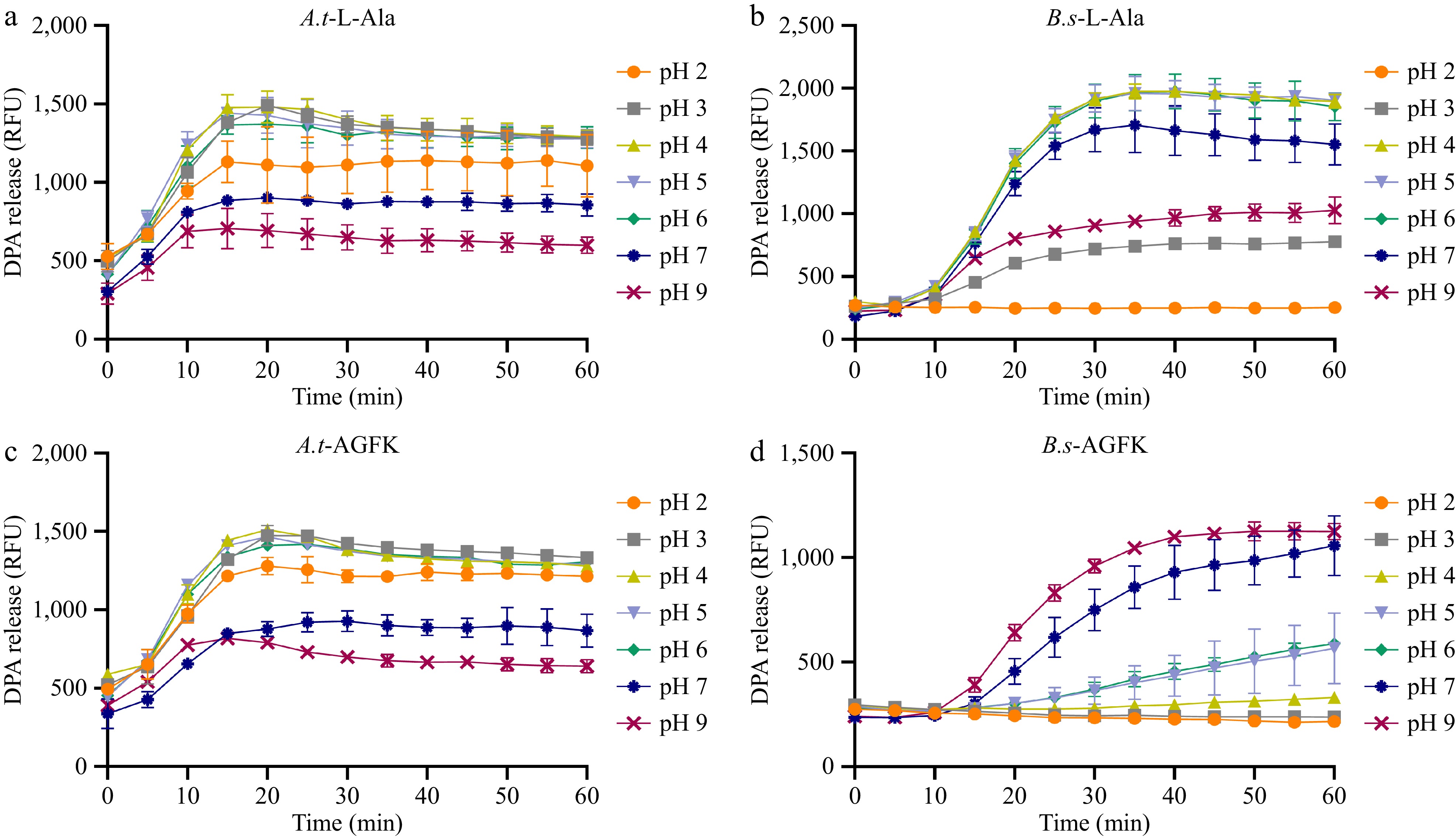
Figure 1.
Germination of A. acidoterrestris (A.t) and B. subtilis (B.s) spores induced by nutrient germinants under pH conditions. (a) DPA release from A. acidoterrestris spores induced by 10 mM L-alanine. (b) DPA release from A. acidoterrestris spores induced by 10 x AGFK; DPA release from B. subtilis spores induced by (c) 10 mM L-alanine, and (d) 10 x AGFK. Each experiment in the figure included three replicate groups.
An acidic condition is crucial to induce A. acidoterrestris spore germination
-
The efficiency of spore germination is highly reliant on the concentration of nutrient germinants, and the application of heat shock[38]. Therefore, we further optimized the concentrations of the two nutrient germinants, L-Ala and AGFK, for inducing germination of A. acidoterrestris spores at pH 4 (Fig. 2a, b). Interestingly, following heat shock at 75 °C for 30 min, spores of A. acidoterrestris exhibited significant DPA release regardless of whether L-Ala or AGFK was present (Fig. 2a, b). Moreover, the addition of extra nutrient germinants did not further enhance spore germination (Fig. 2a, b). These results suggest that A. acidoterrestris spores are capable of germinating in the absence of nutrient germinants, which is sharply distinct from the germination behavior of B. subtilis spores that require nutrient germinant signaling or external stimuli to initiate germination[39]. For A. acidoterrestris, nutrient germinants appear to have limited influence, whereas acidic conditions and heat activation seem to play important roles in triggering germination.
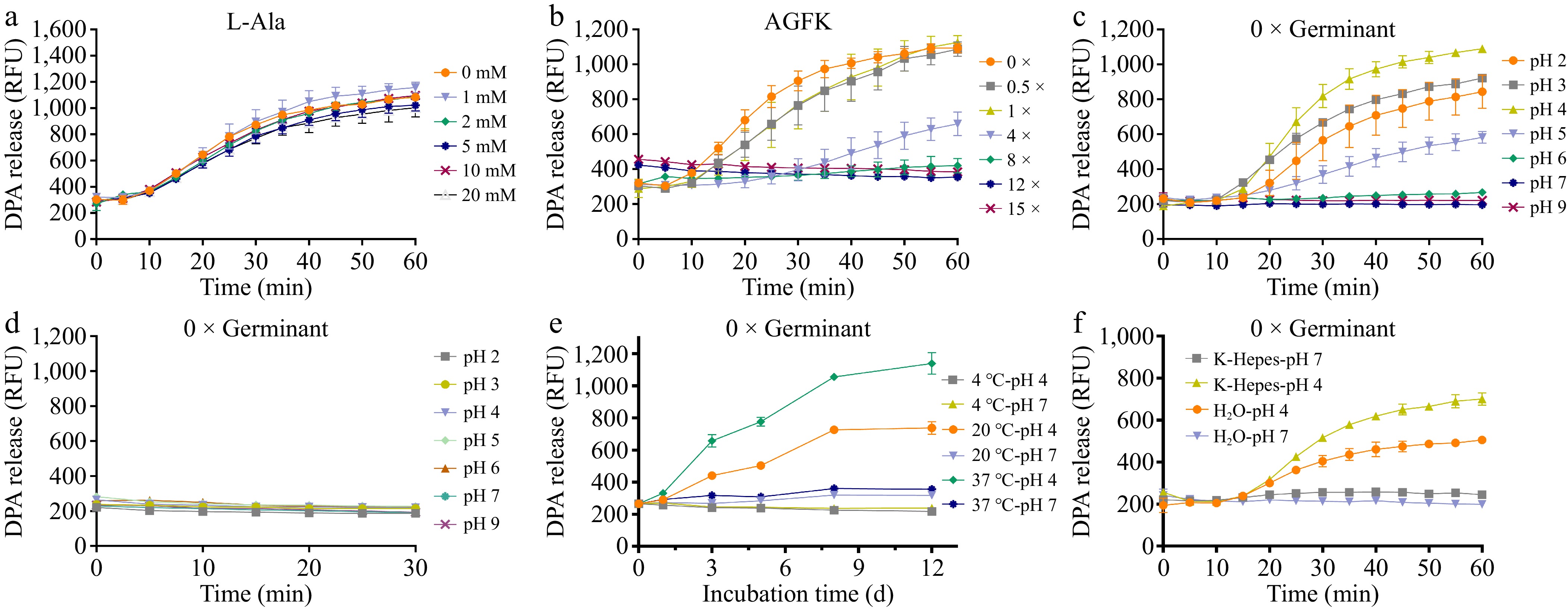
Figure 2.
Germination of A. acidoterrestris spores under varying nutrient germinant concentrations and pH conditions. DPA release from heat-activated spores induced by different concentrations of (a) L-alanine, and (b) AGFK in pH 4 K-Hepes; DPA release from (c) heat-activated spores, and (d) non-heat-activated spores under pH 2–9 conditions in the absence of nutrient germinants; (e) DPA release of spores incubated in pH 4 and 7 systems at 4, 20, and 37 °C for 1, 3, 5, 8, and 12 d. (f) DPA release from spores was monitored in germinant-free sterile water and K-Hepes buffers at pH 4 and 7. Each experiment in the figure included three replicate groups.
Based on this observation, nutrient germinants were removed from the germination system and DPA release evaluated under different pH conditions. The results showed that A. acidoterrestris spores could still germinate without nutrient germinants, with the highest germination efficiency observed at pH 4 (Fig. 2c). To investigate the role of heat shock in germination, non-heat-shock spores incubated at 37 °C for 30 min were monitored. These spores did not germinate within this short incubation period (Fig. 2d). To further examine whether extended incubation could induce germination without heat activation, non-heat-shock spores were suspended in pH 4 and 7 K-Hepes buffer, and incubated at 4, 20, and 37 °C. Germination was assessed periodically over 12 d (Fig. 2e). Notably, non-heat-shock spores in pH 4 buffer showed significant germination after 5 d at 37 °C (Fig. 2e), albeit at a much slower rate than heat-activated spores (Fig. 2c). Meanwhile, the germination efficiency at pH 4 was slowed at 20 °C and strongly suppressed at 4 °C (Fig. 2e). In contrast, non-heat-shock spores in pH 7 buffer showed no germination at all. To determine whether K-Hepes acts as a germinant for A. acidoterrestris spores, germination in both sterile water and K-Hepes systems at acidic and neutral pH were compared (Fig. 2f). Notably, substantial DPA release was observed in sterile water at pH 4, whereas minimal release occurred at pH 7 (Fig. 2f). These results confirm that K-Hepes does not function as a germinant for A. acidoterrestris spores. Collectively, these findings indicate that an acidic environment is essential for initiating germination of A. acidoterrestris spores, while heat activation primarily serves to accelerate the germination process.
Optimization of heat shock and incubation parameters for A. acidoterrestris spore germination
-
Heat shock is a standard procedure in spore germination assays, serving to activate spores and enhance synchronization and uniformity within the spore population, thereby ensuring experimental reproducibility and consistency[40]. Heat shock treatments are essential for optimal spore germination analysis[40]. To achieve optimal heat activation effect without compromising spore viability, the heat shock temperature was first optimized for the germination process. A. acidoterrestris spores were subjected to heat shock at temperatures ranging from 65 to 95 °C for 1 h, followed by plate counting to assess spore viability. The results showed that the number of spores began to significantly decrease at 80 °C, with a 3.9 ± 0.27 log reduction at 95 °C. At 75, 70, and 65 °C, there was minimal spore death (Fig. 3a). Given that spores did not die at lower temperatures, it was then investigated whether these spores could still germinate properly after these activation treatments. At pH 4 (optimal for germination as shown in Fig. 2c), spore germination was measured at heat activation temperatures of 75, 70, 65, and 60 °C. The highest DPA release and germination efficiency were observed at 70 °C, and as the heat shock temperature decreased, the germination rate progressively decreased (Fig. 3b). Next, the heat shock time was optimized at 70 °C for spore germination. As shown in Fig. 3c, spores that were not subjected to heat shock or were heat shocked for 1 min showed minimal DPA release. When heat shock was applied at 70 °C for at least 5 min, the germination rate increased significantly. As heat shock time increased, the DPA release rate also increased, reaching its peak at 15 min Fig. 3c). Since no significant difference was observed in DPA release rate among 15–60 min of heat shock, it was further examined if the heat shock time could affect the germination ratio at the end of the germination. To this end, spores heat activated at 70 °C for 15, 30, 45, and 60 min were cultured at 37 °C for 60 min. Spore germination ratios were measured after treatment at 80 °C for 30 min. As shown in Fig. 3d, no significant difference was observed in germination ratio among 15–60 min of heat shock. Since heat activation time from 15–60 min showed no effect on both germination rate and ratio, the optimal heat shock condition for A. acidoterrestris spores was selected as 70 °C for 15 min. To further investigate whether different incubation temperatures during measurement affected spore germination, A. acidoterrestris spores heat-activated at 70 °C/15 min were incubated at 18–42 °C to measure germination. As shown in Fig. 3e, at the incubation temperature of 18 °C, the spore germination rate was the lowest, and germination was inhibited. As the temperature increased, the germination rate significantly improved, and the total DPA release gradually increased, with the highest DPA release and rate observed at 37 °C (Fig. 3e).
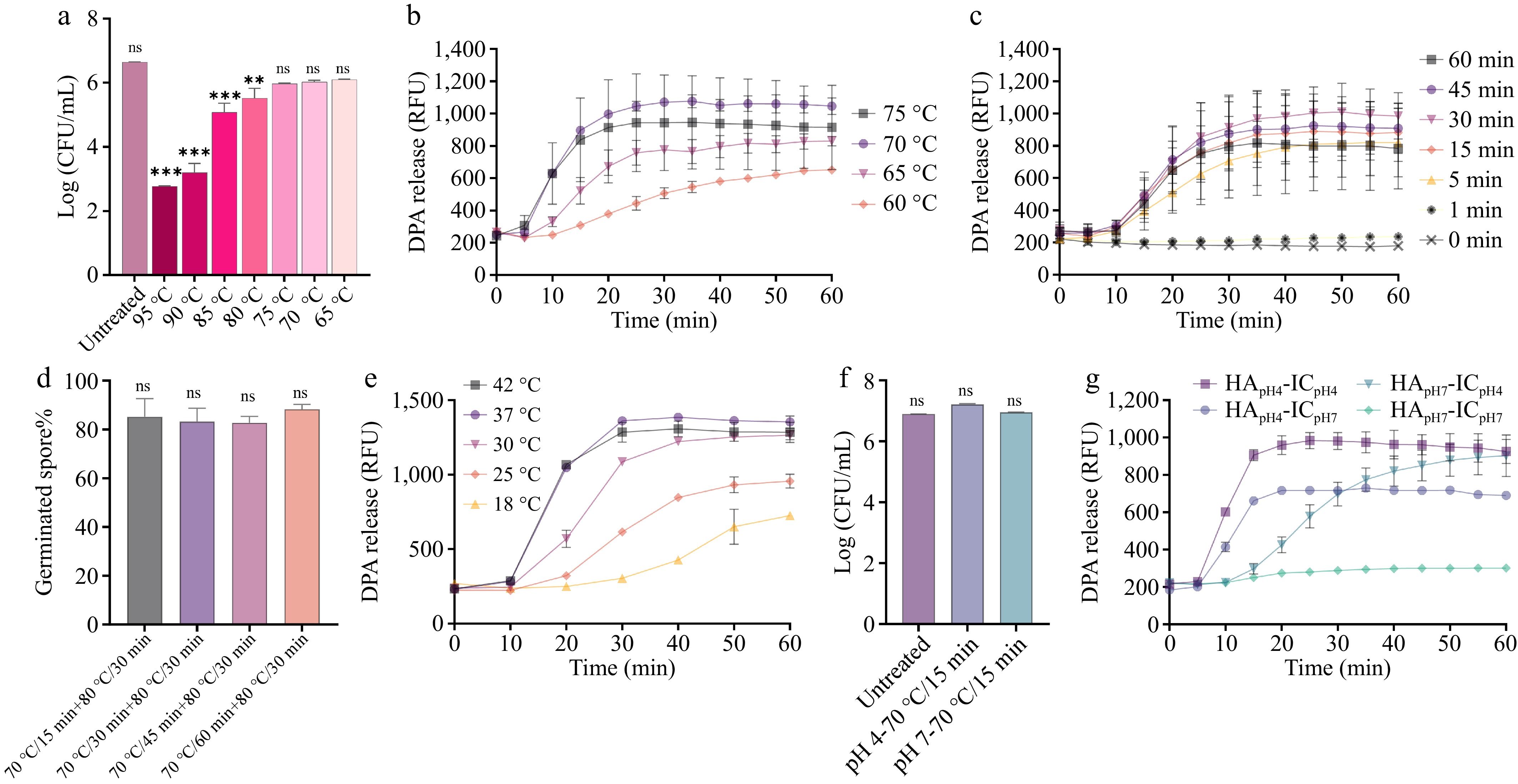
Figure 3.
Optimization of heat activation conditions for A. acidoterrestris spore germination. (a) Heat resistance of A. acidoterrestris spores at 95, 90, 85, 80, 75, 70, and 65 °C for 60 min; (b) DPA release from spores after heat activation at 75, 70, 65, and 60 °C for 30 min in pH 4; (c) heat activation at 70 °C for 1, 5, 15, 30, 45, and 60 min; (d) spores were heat activated at 70 °C for 15, 30, 45, or 60 min, then divided into two groups: one was directly plated for colony enumeration, and the other underwent an additional 80 °C/30 min treatment prior to plating. Percentage of germinated spores was calculated as: (CFU after heat activation-CFU after 80 °C treatment)/CFU after heat activation × 100%; (e) DPA release from spores heat-activated at 70 °C for 15 min and incubated at 18, 25, 30, 37, and 42 °C; (f) viable counts of untreated spores and spores heat activated at 70 °C for 15 min in pH 4 and 7; (g) DPA release from spores heat activated at pH 4 or 7, then immediately incubated (IC) at 37 °C and pH 4 or 7. Each experiment in the figure included three replicate groups.
Based on the above findings, we know that acidic pH conditions significantly influence spore germination efficiency. It was further examined whether the pH level during heat shock treatment would also affect germination efficiency. Notably, heat shock treatments were conducted at pH 4 and 7 without inactivation of spores (Fig. 3f), thus excluding the possibility that DPA release resulted from spore inactivation. As shown in Fig. 3g, heat activation and subsequent incubation at pH 4 yielded the highest germination rate and ratio. By contrast, when incubation was performed at pH 7, the germination ratio decreased significantly, although the germination rate remained largely unaffected. When heat activation was conducted at pH 7 followed by incubation at pH 4, a pronounced reduction in germination rate was observed, yet the overall germination ratio was comparable to that under optimal conditions. In neutral conditions (i.e., both heat activation and incubation at pH 7), spore germination was strongly suppressed, with nearly no germination detected. These findings suggest that while both steps influence germination, the pH during incubation for the germination assay is the most critical factor determining the germination outcome of A. acidoterrestris spores.
Optimizing pH conditions for A. acidoterrestris spore germination induced by HHP
-
The germination of bacterial spores induced by HHP is well-established, with pH recognized as a critical factor regulating germination efficiency[41]. The germination response of A. acidoterrestris spores to HHP treatment across a range of pH conditions were characterized. A. acidoterrestris and B. subtilis spores were treated at 30 °C, 200 or 500 MPa HHP for 1–10 min. The release of DPA was measured to examine germination, while the number of surviving spores was determined to examine their viability after 10 min of HHP treatment. As shown in Fig. 4a, A. acidoterrestris spores treated at 200 MPa exhibited substantial DPA release within the pH range of 5–7, whereas the DPA release was reduced under pH 2–4 conditions. Notably, spore viability remained consistent across pH 3–9 (Fig. 4b), confirming that DPA release resulted from germination rather than loss of viability. A similar pH-dependent germination profile was observed for B. subtilis spores, with pronounced DPA release occurring between pH 5 and 7 and stronger suppression at pH 2–4 compared with A. acidoterrestris spores (Fig. 4c). Consistent with this, no significant loss in viability was detected between pH 2 and 9 (Fig. 4d), indicating that the HHP treatment at 200 MPa did not cause spore inactivation. Together, these data demonstrate that at 200 MPa, both A. acidoterrestris and B. subtilis spores exhibited optimal germination rates and ratios within pH 5–7, whereas A. acidoterrestris spores remained capable of germinating under acidic conditions (pH 2–5), confirming their acidophilic nature. At 500 MPa, A. acidoterrestris spores showed elevated germination rates and ratios at pH 6–9, whereas germination was considerably slower between pH 3 and 5 (Fig. 4e). The concomitant decrease in viable counts at pH 2–3 (Fig. 4f) suggests that part of the DPA release under these conditions may be attributable to spore rupture or inactivation. In contrast, B. subtilis spores displayed substantial DPA release across pH 2–9 under 500 MPa (Fig. 4g). However, a sharp decline in viability was observed from pH 2 to 6 (Fig. 4h), implying that the synergistic interaction of high pressure and acidic pH increases spore sensitivity, ultimately leading to death-associated DPA release. Taken together, A. acidoterrestris spores can also exhibited extensive germination efficiency under acidic pH following exposure to 500 MPa.
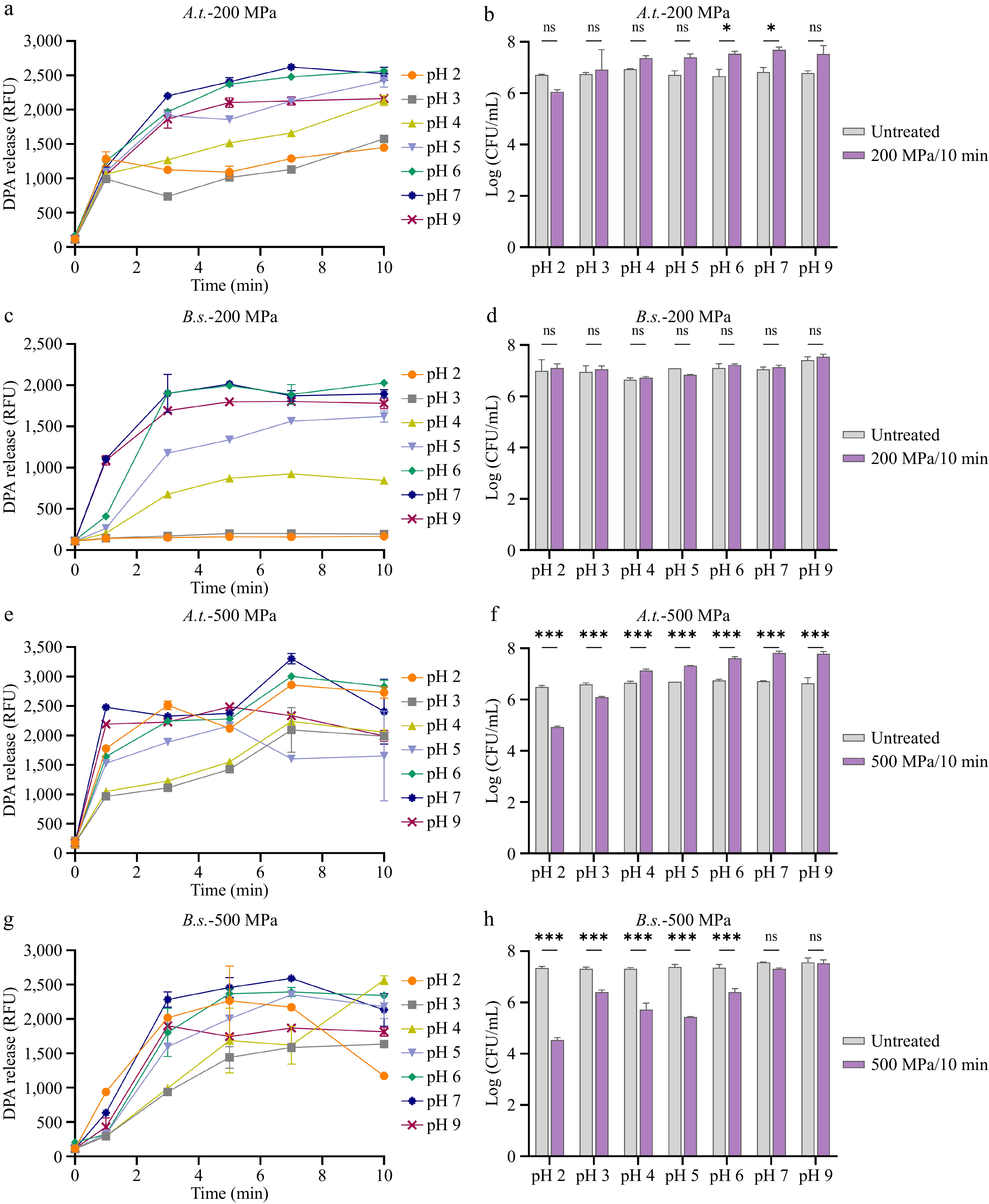
Figure 4.
Germination and viability of A. acidoterrestris (A.t) and B. subtilis (B.s) spores under HHP at various pH conditions. At 200 MPa/30 °C: (a) DPA release, and (b) colony counts of A. acidoterrestris spores; (c) DPA release, and (d) colony counts of B. subtilis spores. At 500 MPa/30 °C: (e) DPA release, and (f) colony counts of A. acidoterrestris spores; (g) DPA release, and (h) colony counts of B. subtilis spores. Each experiment in the figure included three replicate groups.
Spontaneous germination of HHP-treated A. acidoterrestris spores
-
Building on previous findings regarding B. subtilis SD spores undergo spontaneous germination at 500 MPa[32], the aim was to investigate whether A. acidoterrestris spores also exhibit spontaneous germination after HHP treatment. A. acidoterrestris spores were treated at 200 or 500 MPa for 3 min at 30 °C under both pH 4 and 7 conditions. After the HHP treatment, the spores were immediately placed on ice, followed by centrifugation. The spore pellet was resuspended in pH 4 and 7 K-Hepes buffer, and incubated at 37 °C. Spontaneous germination was detected by monitoring DPA release. As shown in Fig. 5a, untreated spores did not germinate, whereas spores treated at both 200 and 500 MPa showed significant DPA release during incubation at 37 °C, signifying extensive spontaneous germination occurred. Notably, spontaneous germination efficiency was elevated under acidic conditions, even though the final germination ratio remained largely unchanged. Colony counting results confirmed that the germination observed was not due to spore death but true spontaneous germination, as the spore viability remained unchanged after HHP treatment (Fig. 5b).
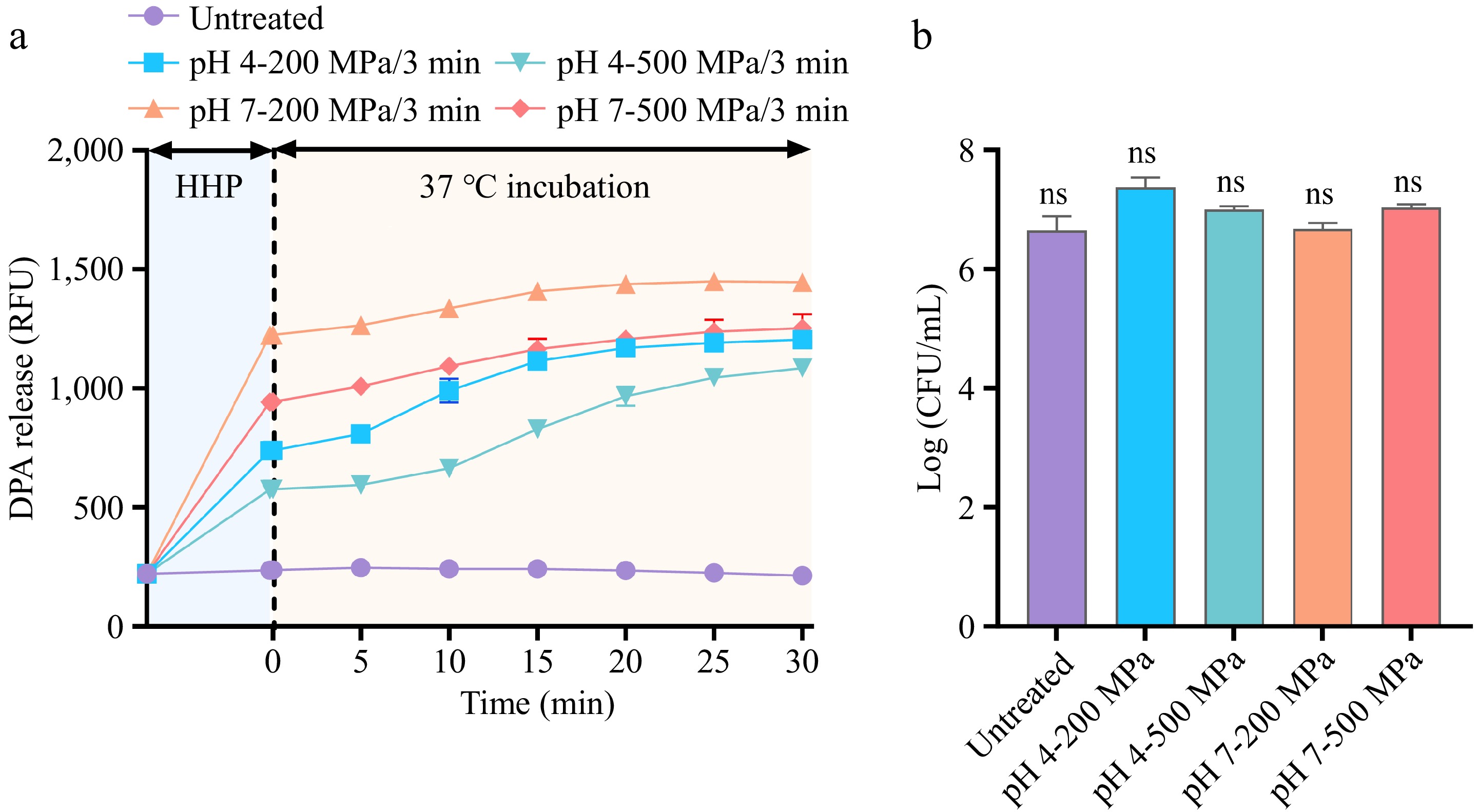
Figure 5.
Spontaneous germination and viability of A. acidoterrestris spores under acidic and neutral conditions. (a) After HHP (200 or 500 MPa/3 min/30 °C) , DPA release in the supernatant was measured immediately, the pellet was subsequently incubated at 37 °C to monitor DPA release over a 30 min period. (b) Colony counts of spores at pH 4 and 7 immediately after the HHP treatment. Each experiment in the figure included three replicate groups.
Efficient inactivation of A. acidoterrestris spores by 'germination-inactivation' strategy
-
Given that A. acidoterrestris spores are predominantly associated with acidic food products, pH 4 was selected as the representative condition for subsequent experiments[42]. Building on the above findings of A. acidoterrestris spore germination triggered by heat activation, acidic stimulation, and HHP, the next aim was to develop an efficient 'germination-inactivation' strategy for eliminating A. acidoterrestris spores. Two sequential approaches were designed. The first involved heat activation of spores at 70 °C for 15 min, followed by incubation at 37 °C for 1 h to promote germination, and then application of either thermal treatment (80 °C/30 min) or HHP (500 MPa/10 min). The second approach began with HHP treatment (200/10 min or 500 MPa/10 min), followed by incubation at 37 °C for 1 h to promote spontaneous germination, and then a secondary treatment of either heat (80 °C/30 min) or HHP (500 MPa/10 min). As shown in Fig. 6, neither 80 °C/30 min nor 500 MPa/10 min alone sufficiently inactivated A. acidoterrestris spores with none germinant stimuli activation. However, when spores were heat-activated (70 °C, 15 min) and incubated (37 °C, 1 h) prior to treatment with 80 °C/30 min or 500 MPa/10 min, 0.92 ± 0.04 and 0.57 ± 0.23 log reductions were achieved, respectively, with no significant difference observed between these two treatments. In contrast, spores pretreated with 200 MPa/10 min exhibited minimal reduction after subsequent incubation alone, but additional treatment with either 80 °C for 30 min or 500 MPa HHP led to substantial log reductions of 5.01 ± 0.40 and 3.88± 0.26, respectively. Similarly, pretreatment with 500 MPa/10 min resulted in limited inactivation after incubation, but when followed by 80 °C/30 min or 500 MPa/10 min, 5.31 ± 0.51 and 4.34 ± 0.44 log reductions were reached, respectively. These findings demonstrate that a combined approach comprising HHP pretreatment, incubation at 37 °C, and subsequent mild pasteurization effectively eliminates A. acidoterrestris spores. Notably, the enhanced inactivation observed with high-pressure pretreatment, as compared to heat shock treatment (70 °C, 15 min), suggests that HHP (200 or 500 MPa) promotes a greater extent of spore germination.
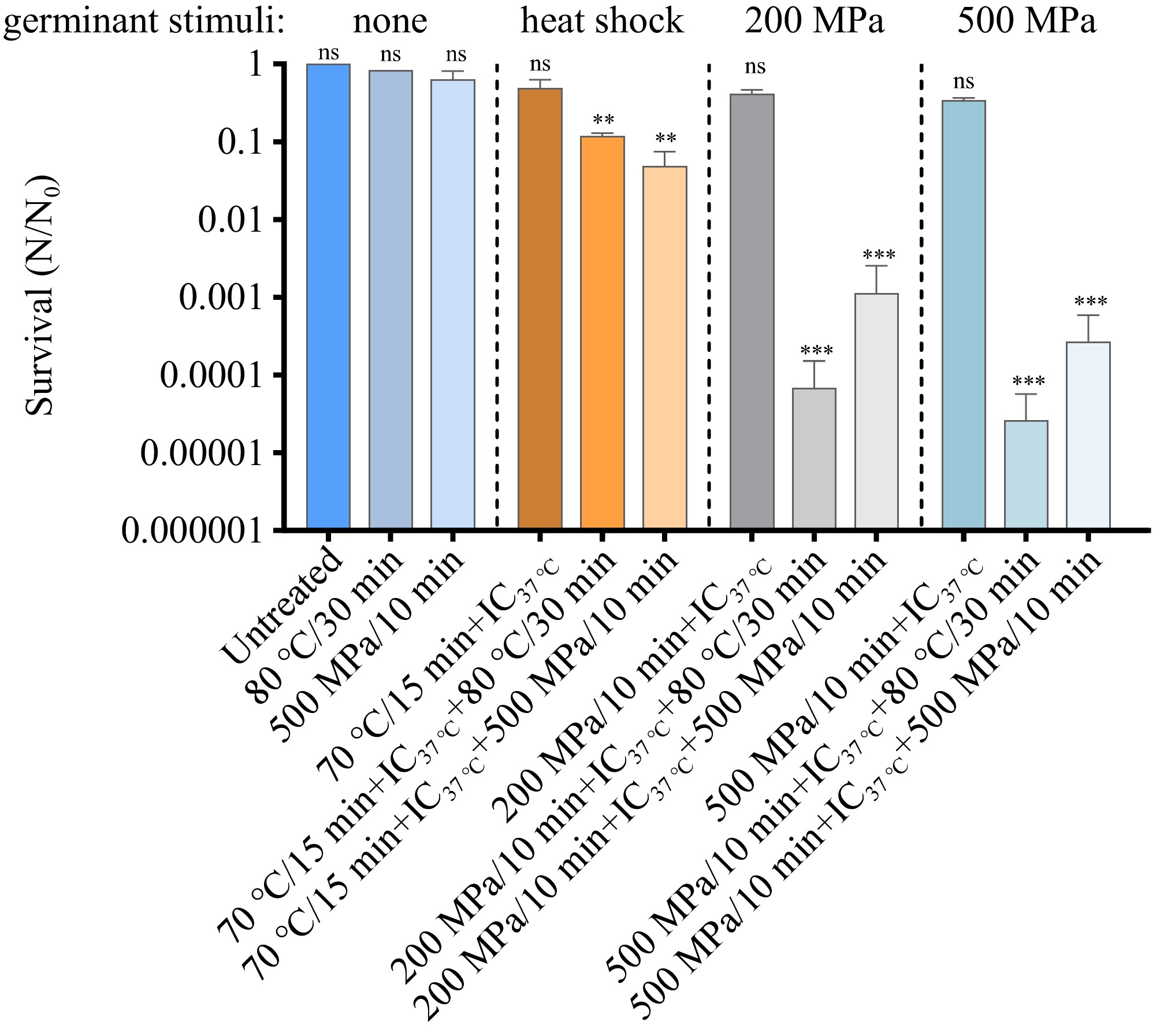
Figure 6.
Inactivation of A. acidoterrestris spores using germination-inactivation strategy. Spores were first subjected to heat activation at 70 °C for 15 min, followed by incubation at 37 °C for 1 h, then application of 80 °C/30 min or HHP (500 MPa/10 min). The second approach began with HHP treatment (200 Mpa/10 min or 500 MPa/10 min), followed by incubation at 37 °C for 1 h, then application of 80 °C/30 min or HHP (500 MPa/10 min). Control groups included untreated spores, spores treated only at 80 °C for 30 min, only at 200 MPa for 10 min, or only at 500 MPa for 10 min. Each experiment in the figure included three replicate groups.
-
In this study, it is reported for the first time, that A. acidoterrestris spores are able to germinate without nutrient germinant (Fig. 2). Acidic conditions (such as pH 4) was identified as a critical trigger for this process, while heat activation further enhanced germination efficiency (Figs 2 and 3). However, B. subtilis spores need germinants for triggering germination, and the germination process would be inhibited when exposed to the acidic conditions (Fig. 1)[43]. This phenotypic divergence may stem from structural and physiological differences between A. acidoterrestris and B. subtilis spores. As for B. subtilis spores, their germination can be triggered by the typical germinant L-Ala, which binds to and activates the germinant receptor GerA, initiating the sequential germination events, including ions release (H+, Na+, K+), DPA release, and cortex hydrolysis[44]. It has been proposed that acidic conditions inhibit B. subtilis spore germination by preventing L-Ala binding to the binding site of GerA, a process likely involved protonation of ionizable groups associated with the binding site[45,46]. This can prevent L-Ala-GerA binding by altering the configuration of the binding site or by blocking access of the germinant to the site[46]. As for A. acidoterrestris spores, BLASTP analysis (
https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome ) strongly suggests that they have GerA-type GRs and SpoVA proteins similar to those of B. subtilis spores. However, their germination in response to acidic stimuli, rather than nutrient germinants, is quite unexpected and interesting. Since the vegetative cells of A. acidoterrestris need the acidic condition for growth[12], the necessity of the acidic conditions for their spores to germinate seems reasonable, as the acidic condition indicates a favorable condition for their proliferation. Then, the obvious question is: How does the acidic stimuli trigger A. acidoterrestris spore germination? The answer to this question is currently unclear. By referring to acidic-induced L-Ala germination inhibition in B. subtilis spores, we can speculate that the acidic-induced protonation of ionizable regions of GerA-type GRs of A. acidoterrestris spores may somehow activate the GRs, and then induce subsequent germination events. Alternatively, it is also possible that acidic conditions may directly open the SpoVA channel by the protonation effect, and thus induce germination. Nonetheless, both of these possibilities need to be further investigated.Another key finding is that when exposed to HHP, A. acidoterrestris spores exhibited a germination response similar to that of B. subtilis, with markedly higher efficiency at neutral pH and significant inhibition under acidic conditions (Fig. 4). This aligns with the report by Vercammen et al., who observed suppressed germination of A. acidoterrestris spores in citrate buffer at pH 4.2 compared to pH 7.0 under HHP at 40 °C[30]. These results suggest that the mechanism governing HHP-induced A. acidoterrestris spore germination is distinct from that activated by acidic stimuli, warranting further investigation into the underlying molecular pathways. The potential mechanism may involve the release of H+, monovalent cations, and Zn2+ during the initial stages of spore germination. H+ release causes an increase in the pH of the spore core[47]. HHP treatment under acidic conditions promotes massive proton influx into the spore core, displacing minerals and forming demineralized H-type spores[48]. This results in decreased pH within the germ nucleus, inhibiting the initiation of key germination events. Consequently, pressure-induced germination is significantly suppressed under acidic conditions. It was also observed that A. acidoterrestris spores treated at 200 or 500 MPa underwent spontaneous germination during subsequent incubation at 37 °C. This demonstrated the presence of a subpopulation of SD spores with inherent instability. Although not isolated here, these SD spores resemble the HPSD spores reported in B. subtilis[32,49]. Similar instability has been described in B. subtilis spores separated after 150 MPa[50], and Heydenreich et al. observed that HPSD spores germinate more effectively upon re-exposure to very high pressure[51]. The existence of such unstable SD spores necessitates refinement of the 'germination-inactivation' strategy. In particular, combining HHP with immediate heat treatment may fail to inactivate SD spores fully. Instead, a multi-step approach-incorporating sufficient incubation to commit SD spores to germination, followed by mild inactivation treatments-is required to achieve maximal reduction.
Accordingly, an optimized 'HHP-incubation-pasteurization' for A. acidoterrestris spore inactivation under acidic conditions was developed: treatment at 200 or 500 MPa for 10 min at 30 °C, incubation at 37 °C for 1 h, and finally pasteurization at 80 °C for 30 min, which achieved 5.0 ± 0.40 and 5.3 ± 0.51 log reductions of A. acidoterrestris spores, respectively (Fig. 6). In comparison, heat activation followed by incubation and pasteurization only achieved 0.92 ± 0.04 log reduction (Fig. 6). These results indicate that HHP triggers more effective germination than heat activation alone, and that subsequent incubation further depletes the resistance of the HHP-treated spores. Then, two questions are consequently raised: (1) Why is HHP more effective than heat activation in inducing germination? and (2) Why does incubation further reduce spore resistance after HHP treatment? For the first question, one possible explanation is that HHP can act simultaneously on both membranes and GR proteins[23,50], exerting multiple activating effects on germinant receptors, thereby inducing germination more effectively than heat alone. For the second question, two mechanisms may account for this phenomenon: (1) In a subset of HHP-treated spores, partial DPA release occurs without cortex degradation. These spores retain strong resistance, particularly to pressure, and incubation promotes cortex degradation, thereby lowering their resistance[52]. (2) Deeply dormant spores subjected to HHP become unstable, with some entering a commitment state. During incubation, these committed spores complete germination, which reduces the overall resistance of the spore population[32]. Based on these findings, it is hypothesized that HHP efficiently initiates germination, and incubation facilitates cortex hydrolysis, core hydration, and spontaneous germination, collectively sensitizing spores to mild inactivation treatments. The residual surviving population likely represents true SD spores, which will require further tailored strategies for complete eradication. In summary, A. acidoterrestris is a major spoilage organism responsible for the deterioration of acidic fruit juices after pasteurization, leading to substantial quality loss and significant economic damage. Therefore, in this study, A. acidoterrestris was selected as a model organism to investigate in depth the germination phenotypes and behaviors of this group of acidophilic and thermophilic spoilage bacteria. Moreover, an efficient inactivation strategy and an acid-compatible 'HHP–incubation–pasteurization' treatment was developed that effectively eliminates spores under acidic conditions. This approach holds considerable practical relevance for the fruit juice processing industry.
-
In this research, it was demonstrated that A. acidoterrestris spores can germinate in buffer systems in the absence of exogenous germinants, with acidic conditions acting as a key trigger, while heat activation further enhanced germination efficiency. Meawhile, HHP-induced germination of A. acidoterrestris spores also occurred under acidic conditions, demonstrating their acidophilic nature. Furthermore, spores treated with HHP (200 or 500 MPa, 3 min, 30 °C) underwent spontaneous germination during subsequent incubation at 37 °C, independent of the pH conditions. A gentle, yet effective inactivation strategy, using HHP at moderate temperature, followed by pasteurization, was also established, tailored for these acidophilic and thermotolerant spores. These findings advance our understanding of the germination physiology of A. acidoterrestris spores, including superdormant subpopulations, and propose practical strategies to control spoilage in acidic foods. This integrated approach, combining HHP and post-incubation, represents a promising, minimally severe method for managing A. acidoterrestris spores in high-acid juice and beverage processing.
-
The authors confirm their contributions to this paper as follows: study conception and design, funding acquisition: Rao L; writing − review and editing: Ding Y, Rao L; data curation, investigation: Ding Y, Lyu F, Gong Z; formal analysis: Yang D, Wang Y; data collection: Ding Y; project administration and supervision: Rao L, Yang D, Wang Y. All authors reviewed the results and approved the final version of the manuscript.
-
All statistics generated or analyzed during this study are included in this published article.
-
This work was supported by National Key R&D Program of China (Grant No. 2023YFD2100303), Key Research and Development Program of Ningxia Hui Autonomous Region (Grant No. 2025BEE02003), National Natural Science Foundation of China (NSFC) (Grant Nos 32522084 and 32372470), Agricultural Research Outstanding Talents of China (Grant No. 13210317). And 2115 Talent Development Program of China Agricultural University.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of China Agricultural University, Zhejiang University and Shenyang Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ding Y, Lyu F, Gong Z, Yang D, Wang Y, et al. 2026. Optimizing germination conditions for Alicyclobacillus acidoterrestris spores, and achieving efficient inactivation via high hydrostatic pressure using a germination-inactivation strategy. Food Innovation and Advances 5(1): 102−111 doi: 10.48130/fia-0026-0006 

Optimizing germination conditions for Alicyclobacillus acidoterrestris spores, and achieving efficient inactivation via high hydrostatic pressure using a germination-inactivation strategy
- Received: 18 October 2025
- Revised: 28 December 2025
- Accepted: 15 January 2026
- Published online: 13 March 2026
Abstract: The 'germination-inactivation' strategy can potentially eliminate Alicyclobacillus acidoterrestris spores by exploiting the loss of their resistance that occurs during germination. However, the germination behavior of A. acidoterrestris spores remains incompletely characterized. In this study, the germination responses of A. acidoterrestris spores to various stimuli, including L-alanine, AGFK (L-asparagine, D-glucose, D-fructose, and KCl), and high hydrostatic pressure (HHP) were comprehensively examined. Surprisingly, it was observed that A. acidoterrestris spores germinate in acidic buffer, even in the absence of L-alanine or AGFK. Optimal acidic-induced germination efficiency was achieved at pH 4.0 following heat shock (70 °C, 15 min), and incubation at 37 °C. While heat treatment (65–75 °C) accelerated germination, acidic conditions (pH 3.0–4.0) were strictly necessary to initiate the process. Meanwhile, HHP-induced germination can also occur under acidic conditions (pH 3.0–5.0), demonstrating the acidophilic nature of A. acidoterrestris spores. Notably, spores subjected to HHP (200 or 500 MPa, 3 min, 30 °C) underwent spontaneous germination during subsequent incubation at 37 °C, irrespective of the medium's pH. Building on these findings, a combined 'HHP-incubation-pasteurization' approach was developed that achieved up to a 5.3 ± 0.51 log reduction in A. acidoterrestris spores within acidic systems. The results of this study provide a valuable foundation for the development of efficient sterilization techniques in acidic juice processing.