










-
Soil salinization is a major environmental constraint that severely limits plant growth, and reduces agricultural and forestry productivity worldwide[1−3]. It is estimated that approximately 800 million hectares of global land area, accounting for about 6% of total terrestrial land, are affected by excessive salt concentrations, with salinity impacting nearly 20% of irrigated arable land. These saline-affected areas continue to expand due to climate change and unsustainable irrigation practices[2,4]. Under salt stress, plants experience secondary stresses including osmotic imbalance, ion toxicity, and oxidative damage, which collectively disrupt metabolic homeostasis, photosynthetic efficiency, and developmental processes[5]. Therefore, elucidating the genetic mechanisms and signaling networks underlying salt tolerance in woody species is critical for developing stress-resilient cultivars and mitigating productivity losses in marginal environments.
Salt stress induces ionic toxicity in plant cells by disrupting the Na+/K+ ratio, which impairs physiological functions, compromises metabolic homeostasis, and inhibits growth[6,7]. Concurrently, high salinity triggers the excessive accumulation of reactive oxygen species (ROS) such as hydroxyl radicals (·OH), hydrogen peroxide (H2O2), and superoxide anions (O2·−) within plant cells[8−11]. This oxidative burst promotes lipid peroxidation, damaging cellular components such as nucleic acids, proteins, and lipids, and can ultimately lead to cell death[12−14]. To cope with prolonged stress, plants have evolved sophisticated adaptive mechanisms involving coordinated physiological, biochemical, and signaling pathways[2,15]. Among these mechanisms, enzymatic antioxidants play a critical role in scavenging excess ROS and maintaining normal growth under stressful conditions[16−18].
Abscisic acid (ABA) is a key phytohormone that mediates stress signaling and promotes sodium ion efflux and water uptake under salt stress conditions[19−23]. AREB/ABF transcription factors are members of the bZIP family that function in the ABA signaling pathway. They bind to ABA-responsive elements (ABRE: PyACGTGG/TC) in the promoters of stress-responsive genes, thereby activating their expression to enhance abiotic stress tolerance. The role of AREB transcription factors in regulating drought and salt stress responses is well established. For instance, transgenic Arabidopsis plants overexpressing AREB1/ABF2, AREB2/ABF4, or ABF3 exhibit enhanced drought tolerance[23]. In poplar, PtrAREB1-2 binds to the ABRE motifs in drought-responsive genes such as PtrNAC and PtrHox, playing a critical role in their transcriptional activation under drought conditions in Populus[24−26]. Furthermore, PagAREB1-3 has been identified as a positive regulator of salt tolerance, and the PagAFP2a-PagAREB1s module forms a negative feedback loop in ABA signaling that fine-tunes salt stress responses in Populus[27].
At the transcriptional level, stress adaptation is coordinated by an intricate regulatory network involving multiple transcription factor families, including HD-ZIP, bZIP, MYB, NAC, WRKY, and AP2/ERF[26,28−33], which collectively modulate stress-responsive gene expression and signal transduction pathways. Among these, the homeodomain-leucine zipper (HD-ZIP) family represents a plant-specific group of transcription factors characterized by a homeodomain (HD) responsible for DNA binding and an adjacent leucine zipper (LZ) motif that facilitates protein dimerization[34]. Phylogenetic analyses classify HD-ZIP proteins into four distinct subfamilies (HD-ZIP I-IV) based on sequence homology, gene structure, and functional specialization[35,36]. As one of the largest subfamilies, HD-ZIP I members orchestrate crucial regulatory functions in abiotic stress responses, hormonal signaling cascades, and developmental processes[37−39]. Notably, several HD-ZIP I members have been closely linked to stress adaptation. In Arabidopsis, ATHB7 and ATHB12 are upregulated by ABA, water deficit, and osmotic stress[40,41], while ATHB6 interacts with the protein phosphatase ABI1 to modulate ABA signaling[42]. In Medicago truncatula, MtHB1 is induced by salt stress and expressed in root meristems, suggesting its potential role in stress adaptation and root development[36]. Overexpression of MdHB7, as in apple, enhances salt tolerance by improving photosynthetic performance, reducing ROS and Na+ accumulation, and promoting osmolyte synthesis[43]. Similarly, in poplar, PagHB7 interacts with PagABF4 to negatively regulate PagEPFL9 expression, thereby reducing stomatal density and enhancing drought tolerance[28]. These findings underscore the pivotal and evolutionarily conserved role of HD-ZIP I transcription factors, particularly HB7 homologs, in coordinating adaptive responses to abiotic stresses.
Populus is both an ecologically important genus and a valuable economic resource, serving as a model system for studying stress adaptation mechanisms in woody plants. However, soil salinity has become a critical factor that severely limits poplar growth and productivity[44]. In this study, we characterized the function of PagHB7a in poplar was characterized, and its essential role demonstrated in salt stress response. PagHB7a expression was significantly induced by salt stress and ABA treatment, and PagAREB1s were identified as upstream transcriptional activators of PagHB7a under salt stress. Genetic analysis with PagHB7 overexpression or knockout plants demonstrated that it is a positive regulator of salt tolerance. Transcriptome profiling further corroborated these findings and identified the differentially expressed genes in PagHB7 overexpression plants. Collectively, our results demonstrate that PagHB7a enhances salt tolerance in poplar by maintaining redox homeostasis, providing new insights into the molecular mechanisms of stress adaptation in woody plants.
-
The hybrid poplar (Populus alba × Populus glandulosa) clone 84K was used in this study. Poplar cuttings were propagated through tissue culture on Murashige and Skoog (MS) medium. All plants were cultivated in a controlled growth chamber under a 16 h light/8 h dark cycle at 25 °C.
To examine the tissue-specific expression of PagHB7a in poplar, stem, leaf, root, and petiole samples were collected from 2-month-old wild-type (WT) plants grown in a growth chamber. The expression pattern of PagHB7a in response to NaCl and ABA was analyzed in 45-day-old 84K poplar seedlings using previously described methods with minor modifications[45,46]. For NaCl treatment, 20-day-old WT plants cultivated in 1/4 Hoagland’s solution were transferred to fresh 1/4 Hoagland’s solution supplemented with 100 mM NaCl. Root tissues were harvested after 0, 1, 3, 6, 12, 24, and 48 h of treatment for RT-qPCR analysis. For ABA treatment, 20-day-old poplar plants maintained in 1/4 Hoagland’s solution were sprayed with 200 μM ABA solution containing 0.1% (v/v) TWEEN 20[45]. Leaf samples were collected at 0, 1, 3, 6, 12, and 24 h post-treatment for subsequent RT-qPCR. The primers used are listed in Supplementary Table S1.
Salt stress treatments were applied as previously described[27]. For short-term NaCl treatment, 1-month-old plants were pre-cultured in 1/4 Hoagland’s solution for 20 d, and then exposed to 150 mM NaCl solution for 2 d. At the end of the treatment, two leaves from the same nodal position on three individual plants were collected, immediately frozen in liquid nitrogen, and used for subsequent physiological assays. For long-term salt stress evaluation, 1-month-old plants were transplanted into soil and grown for 30 d, followed by irrigation with 100 mM NaCl solution for 20 d. Growth and biomass parameters were measured at the end of the treatment period, with 6–12 plants analyzed per genotype in each experiment.
To assess ABA sensitivity, seven-day-old poplar seedlings were transplanted onto MS medium supplemented with 5 μM ABA for 23 d[46]. After the treatment, growth phenotypes were documented, root fresh weight was measured, and root length was quantified using ImageJ software. Leaf tissues were harvested after ABA treatment for RT-qPCR analysis. Four plants of each genotype were used in the experiment. The primers used are listed in Supplementary Table S1.
Gene cloning and plant transformation
-
To construct the Pro35S::PagHB7a vector, the full-length coding sequence (CDS) of PagHB7a was amplified from 84K poplar cDNA using gene-specific primers, and cloned into the PzP211-35S-PolyA binary vector for expression under the control of the CaMV 35S promoter. For the ProPagHB7a::GUS reporter construct, a 2,000 bp fragment upstream of the PagHB7a start codon was amplified from 84K poplar genomic DNA, and inserted into the pBI121 vector to drive β-glucuronidase (GUS) expression. The Pro35S::PagAREB1-3‐GFP overexpression construct was generated by fusing the full-length CDS of PagAREB1-3 (without the stop codon) into the pROKII-GFP vector, also under the control of the CaMV 35S promoter. For CRISPR/Cas9-mediated mutagenesis, two sgRNAs targeting PagHB7a were designed using the web tool (
http://crispr.dbcls.jp ) and assembled into the binary pYLCRISPR/Cas9 vector, according to established protocols[47]. All constructs were introduced into 84K poplar via Agrobacterium-mediated transformation as described previously[48]. The primers used are listed in Supplementary Table S1.RNA extraction and real-time quantitative PCR (RT-qPCR)
-
Total RNA was extracted from 84K poplar plants using the FastPure Universal Plant Total RNA Isolation Kit (Vazyme, RC411-01). Subsequently, 1 μg of total RNA was reverse-transcribed into cDNA using the HiScript II Q RT SuperMix for qPCR Kit (Vazyme, R233-01). Quantitative real-time PCR (RT-qPCR) was performed with ChamQ SYBR Color qPCR Master Mix (Vazyme, Q312-02), using PagActin (Potri.006G192700) as an internal reference gene. Gene expression levels were normalized and calculated using the 2−ΔΔCq method[49]. At least three biological replicates were performed. The primers used for RT-qPCR are listed in Supplementary Table S1.
Subcellular localization analysis
-
To examine the subcellular localization of PagHB7a, the full-length CDS of PagHB7a (without the stop codon) was cloned into the pROKII-GFP vector to generate a C-terminal GFP fusion. The resulting Pro35S::PagHB7a-GFP construct, and an empty pROKII-GFP control vector, were separately transfected into 84K poplar protoplasts. Nuclei were counterstained with DAPI. Transfected protoplasts were observed under an LSM880 confocal microscope, with GFP fluorescence detected at 488 nm. Transient transfection of protoplasts was performed according to a previously established method[50]. The primers used are listed in Supplementary Table S1.
GUS staining
-
The positive GUS reporter lines driven by the promoter of PagHB7a (ProPagHB7a::GUS) were cultivated on MS solid medium for 4 weeks. To evaluate salt-induced expression, primary roots of 4-week-old ProPagHB7a::GUS plants were treated with 100 mM NaCl solution, and harvested at 0 and 6 h for GUS staining, as previously described[51]. Histochemical staining was performed using a β-Glucuronidase Reporter Gene Staining Kit (Coolaber, SL7160, China), according to the manufacturer's protocol. To analyze the expression of the GUS gene driven by the PagHB7a promoter under salt stress, 4-week-old ProPagHB7a::GUS transgenic plants were grown in 1/4 Hoagland’s solution for 10 d before being transferred to 100 mM NaCl solution. Roots were harvested at 0 and 6 h after treatment for RT-qPCR analysis. Three independent ProPagHB7a::GUS plants were used for staining and RT-qPCR analysis.
Physiological analysis
-
Salt-treated poplar leaf tissues were harvested and ground in liquid nitrogen. Catalase (CAT) and peroxidase (POD) activities, as well as malondialdehyde (MDA) content, were measured using commercial assay kits (Suzhou Keming Biotechnology, China) following the manufacturer's instructions. Hydrogen peroxide (H2O2), and superoxide anion (O2·−) levels were quantified with corresponding detection kits (Solarbio, China). For histochemical staining, fresh leaf discs (0.6 mm in diameter) were harvested and stained following the kit manufacturer's protocols (Solarbio, China). Staining with 3,3′-diaminobenzidine (DAB) was performed for 12 h, and with nitroblue tetrazolium (NBT) for 6 h, both at 30 °C in the dark. The presented images are representative of results obtained from at least three independent biological replicates. Relative conductivity (REC), relative water content (RWC), fresh weight (FW), dry weight (DW), and chlorophyll content were determined using previously described methods[27].
Yeast one-hybrid (Y1H) assay
-
The ABRE cis-element-containing sequences derived from the PagHB7a promoter were cloned and inserted into the pLacZi2u vector[52]. The full-length CDS of PagAREB1-2, PagAREB1-3, PagAREB1-4, and PagABF3 were cloned into the pJG4-5 vector to generate AD-PagAREB1-2/PagAREB1-3/PagAREB1-4/PagABF3 construct, respectively. Various combinations of these recombinant constructs were co-transformed into yeast EGY48 cells. Transformants were selected on synthetic dropout (SD) medium lacking tryptophan and uracil (SD/–Trp/–Ura) and incubated at 30 °C for 2–3 d. Positive colonies were cultured overnight in liquid medium and then transferred to SD/–Trp/–Ura solid medium containing raffinose, galactose, and X-gal (40 mg/L) for chromogenic detection of interactions over 48 to 72 h. Each experiment included at least three biological replicates. The primers used are listed in Supplementary Table S1.
Dual-luciferase assay
-
The full-length coding sequences of PagAREB1-2, PagAREB1-3, PagAREB1-4, and PagABF3 were cloned into the PzP211-35S-PolyA vector to generate effector constructs[53]. The promoter region of PagHB7 containing ABRE cis-elements was inserted into the pGreenII0800−LUC vector to create the reporter construct[53]. Each construct was introduced into Agrobacterium tumefaciens strain GV3101 (pSoup-p19). Bacterial cultures were resuspended in infiltration buffer (10 mM MES, pH 5.6, 1 mM MgCl2, 100 μM acetosyringone), adjusted to an OD600 of 1.0, and mixed at a 1:1 ratio (effector:reporter) before co-infiltration into leaves of Nicotiana benthamiana[54]. After 72 h of transient expression, firefly and Renilla luciferase activities were measured using a dual-luciferase assay kit (Vazyme, DL101-01, China). Three independent biological replicates were performed for each experiment. The primers used are listed in Supplementary Table S1.
Chromatin immunoprecipitation (ChIP)-qPCR
-
Leaf and stem samples from 1-month-old PagAREB1-3-GFP overexpression poplar plants were collected and cross-linked in 1% formaldehyde under vacuum before being ground into a fine powder in liquid nitrogen. Nuclei were isolated using the CelLytic™ PN extraction kit (Sigma-Aldrich). Chromatin was fragmented into 300–500 bp pieces via sonication in lysis buffer (50 mM Tris-HCl pH 8.0, 10 mM EDTA, 0.3% SDS, 0.5% sodium deoxycholate, supplemented with protease inhibitors and 1 mM PMSF). Chromatin immunoprecipitation was carried out using an Anti-GFP antibody (HT801, TransGen Biotech, China) with the ChIP-IT® Express kit (Active Motif; CA, USA; 53008) according to the manufacturer's protocol. Gene-specific primers used for ChIP–qPCR are provided in Supplementary Table S1.
RNA-seq and data analysis
-
One-month-old PagHB7a-OE, PagHB7a-KO, and WT plants were first cultured in 1/4 Hoagland’s solution for 18 d and subsequently treated with 100 mM NaCl for 0 and 6 h, and then roots were collected for total RNA extraction. mRNA sequencing was performed using the Illumina X Ten platform (Anoroad, Beijing, China). Three biological replicates were included per treatment, with each replicate consisting of three pooled plants. Paired-end sequencing (150 bp read length) was conducted. Clean reads were aligned to the Populus trichocarpa v3.0 reference genome using HISAT2 with default parameters[55]. Read counts per gene were obtained using HTSeq-count[56]. Differential gene expression analysis was performed with the edgeR package, considering genes with a p-value < 0.05 as significantly differentially expressed[57]. Gene Ontology (GO) enrichment analysis was carried out using the GOstats package in Bioconductor, with an adjusted p-value < 0.01 as the significance threshold[58].
Accession number
-
Genes from this article can be found with the accession numbers: PagHB7a (Potri.014G103000), PagHB7b (Potri.002G176300), PagAREB1-2 (Potri.002G125400), PagAREB1-3 (Potri.009G101200), PagAREB1-4 (Potri.014G028200), PagABF3 (Potri.004G140600), PagRD26 (Potri.001G404100), PagNAC072 (Potri.011G123300), PagSTZ (Potri.001G295500), PagSTZ1-h4 (Potri.009G089400), PagLEA (Potri.002G165000), PagPUB19 (Potri.015G074200), PagPUB79 (Potri.012G078900), PagRBOH (Potri.003G159800), PagNCED3 (Potri.001G393800), PagNCED5 (Potri.011G112400), PagCYP707A1 (Potri.004G235400).
-
Based on transcriptome data from salt-stress experiments, two genes were identified, designated PagHB7a and PagHB7b, which were rapidly and significantly induced in poplar stems under salt treatment (Supplementary Fig. S1a)[59], suggesting their potential role in salt stress response. PagHB7a and PagHB7b are homologs in Populus with 98% sequence similarity and belong to the HD-ZIP I subfamily. Similar to the Arabidopsis ATHB7 gene, both genes contain a highly conserved DNA-binding homeodomain (HD), and an adjacent leucine zipper domain (LZ) (Supplementary Fig. S1b). Since PagHB7a exhibited more pronounced induction under salt stress, this gene was focused on to further investigate its function in salt stress tolerance.
To further characterize the expression pattern of PagHB7a, RT-qPCR was performed to examine the expression of PagHB7a in various tissues, and under different treatments. Results showed that PagHB7a was expressed in all tissues examined, with relatively lower expression in roots (Fig. 1a). Under NaCl treatment, PagHB7a expression was significantly induced in roots, increasing nearly 10-fold within 6 h (Fig. 1b). Given the key role of abscisic acid (ABA) in plant stress responses, the response of PagHB7a to ABA was also examined. The results showed that ABA treatment strongly induced PagHB7a expression in roots, with a 26-fold increase observed after 12 h (Fig. 1c). β-Glucuronidase (GUS) histochemical staining of ProPagHB7a::GUS transgenic poplar plants further confirmed that the expression of PagHB7a was significantly induced in roots after NaCl treatment (Fig. 1d). Consistent with this, RT-qPCR analysis revealed that the expression of the GUS reporter gene increased more than 15-fold after 6 h of NaCl treatment (Fig. 1e). In agreement with its predicted function as a transcription factor, the PagHB7a-GFP fusion protein was localized exclusively in the nucleus (Fig. 1f).
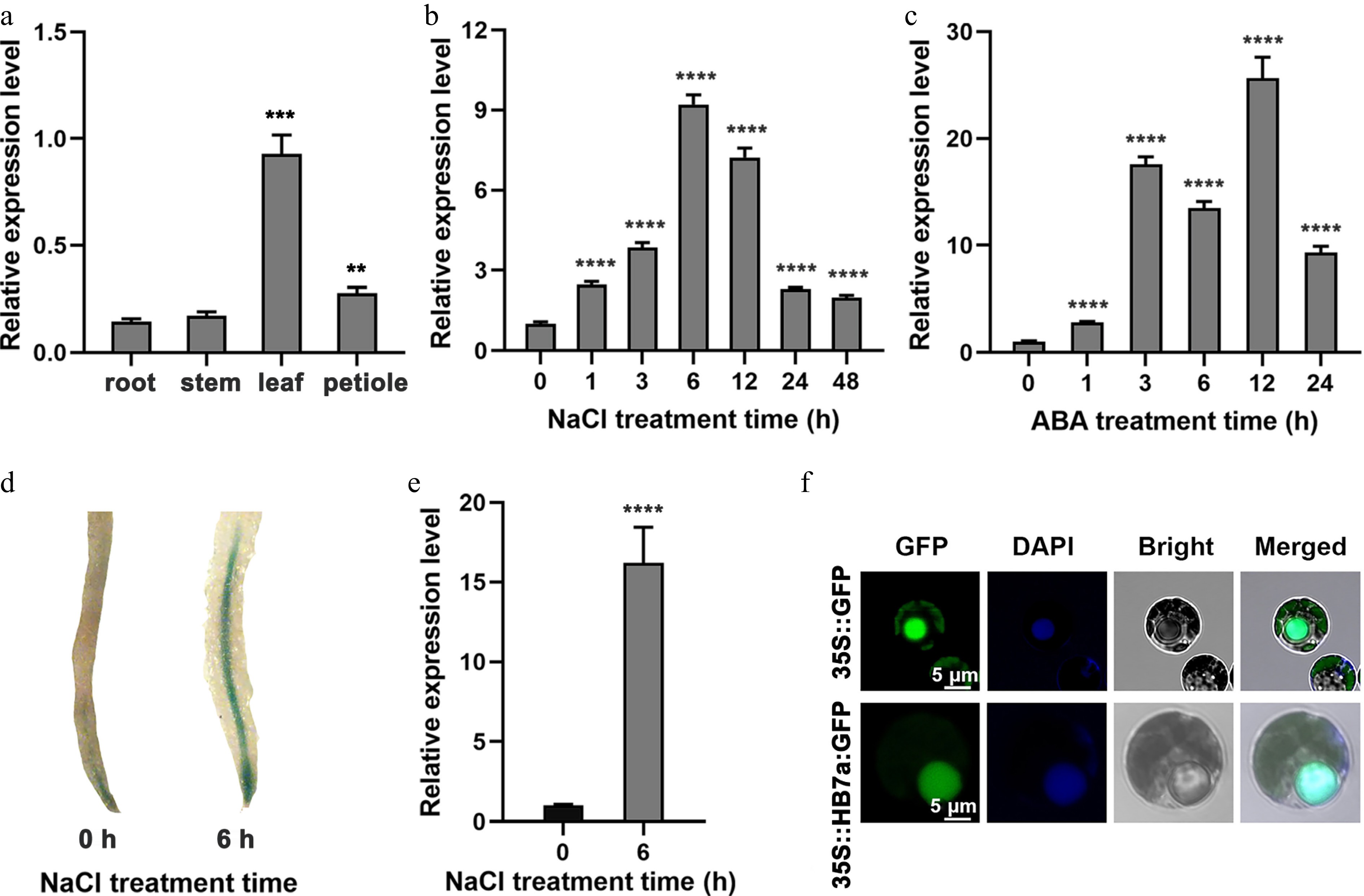
Figure 1.
Expression profiles of PagHB7a in Populus. (a) Relative expression levels of PagHB7a in different tissues analyzed by RT-qPCR. Relative expression of PagHB7a in total roots of 84K poplar under (b) 100 mM NaCl, and (c) 200 μM ABA (c) treatments for the indicated times. (d) Histochemical GUS staining of roots from ProPagHB7a::GUS transgenic plants under 100 mM NaCl treatment. (e) RT-qPCR analysis of GUS expression level in total roots of ProPagHB7a::GUS transgenic plants under 100 mM NaCl treatment. (f) Subcellular localization of PagHB7a-GFP in 84K poplar protoplasts; nuclei were stained with DAPI (4′,6-diamidino-2-phenylindole). In (a)–(c) and (e), error bars represent ± SD (n = 3). Asterisks indicate significant differences compared to the 0 h control (Student's t-test): * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001. Error bars represent SD values (n = 3).
PagAREB1s directly activate the expression of PagHB7a
-
The AREB/ABF gene family plays a critical role in ABA-mediated responses to salt and drought stresses[60]. Analysis of the PagHB7a promoter identified two ABRE motifs (P1 and P2) within the 2,000 bp region upstream of the start codon (Fig. 2a). We previously identified four AREB genes (PagAREB1-2, PagAREB1-3, PagAREB1-4, and PagABF3) that were significantly upregulated under salt stress and co-expressed with PagHB7a within the same co-expression module[59]. To determine whether PagHB7a is a direct target of these transcription factors, yeast one-hybrid (Y1H) assays were performed. The results showed that PagAREB1-2/PagAREB1-3/PagAREB1-4 bound directly to the PagHB7a promoter region P1, while PagABF3 exhibited no binding (Fig. 2b). Chromatin immunoprecipitation (ChIP)-qPCR assays further confirmed the binding of PagAREB1-3 to the P1 region of the PagHB7a promoter (Fig. 2c). Subsequently, transient expression experiments were performed to test whether PagAREB1s could activate the expression of PagHB7a transcription (Fig. 2d). The results revealed that the relative luciferase activity driven by the PagHB7a-P1 promoter was significantly enhanced when co-transformed with the PagAREB1-2, PagAREB1-3, and PagAREB1-4 effector. In contrast, the relative luciferase activity was not significantly different from the empty vector (EV) when co-transformation with PagABF3 (Fig. 2e). Our previous studies found that PagAFP2a, a protein that negatively regulates salt stress, antagonizes the function of PagAREB1-3 by repressing the transcriptional activity of PagAREB1-3 at its target genes in salt stress signaling[27]. Therefore, whether PagAFP2a negatively affects the transcriptional activation of PagHB7a was tested by PagAREB1-3 with dual-luciferase transient expression assays. The results showed that the co-transformation of PagAFP2a with PagAREB1-3 significantly inhibited the activation of PagHB7a by PagAREB1-3 (Fig. 2f). Collectively, these results demonstrate that PagAREB1-2, PagAREB1-3, and PagAREB1-4 directly bind to the PagHB7a promoter and activate its expression.
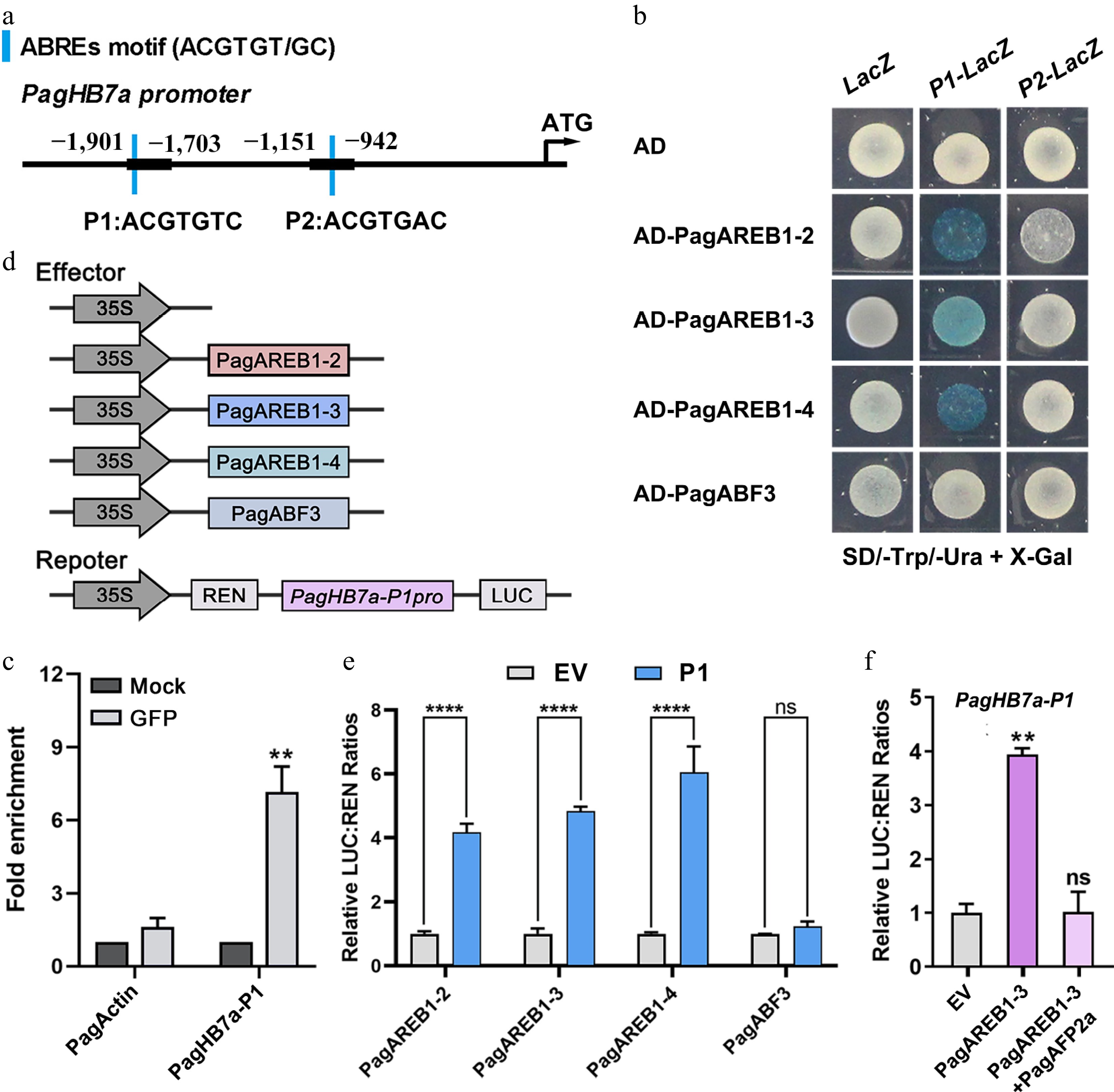
Figure 2.
PagHB7a is directly bound and activated by PagAREB1s. (a) Schematic diagram of the PagHB7a promoter. Blue bars indicate ABRE cis-elements; P1 and P2 denote promoter fragments containing ABRE motifs. (b) Yeast one-hybrid (Y1H) assay shows that PagAREB1-2, PagAREB1-3 and PagAREB1-4 bind to the PagHB7a promoter region containing P1. (c) ChIP-qPCR demonstrates that PagAREB1-3 binds to the PagHB7a promoter region containing P1 in vivo. Statistical comparisons between the Mock (no anti-GFP), and anti-GFP groups were performed using Student's t-test. ** p < 0.01. (d) Schematic representation of effector and reporter constructs used in dual-luciferase assays. (e) Dual-luciferase assay shows that PagAREB1-2, PagAREB1-3, and PagAREB1-4 activate the expression of firefly luciferase (LUC) gene driven by the PagHB7a promoter region containing P1 in tobacco leaves. (f) Dual-luciferase assays show that PagAREB1-3 activates LUC expression driven by the PagHB7a promoter region containing P1, while co-expression of PagAFP2a inhibits the activation. The relative LUC:REN ratio in control samples (transformed with the empty vector, EV) was normalized to a value of 1. Error bars represent SD (n = 3). Asterisks indicate significant differences (Student's t-test): ** p < 0.01; **** p < 0.0001.
PagHB7a is a positive regulator of salt tolerance and ABA sensitivity in Populus
-
To investigate the function of PagHB7a in response to salt stress, PagHB7a overexpression lines (PagHB7a-OE), and CRISPR/Cas9-mediated knockout mutants (PagHB7a-KO) in poplar were generated. Two PagHB7a-OE (OE22, OE26) lines with high transgene expression level, and two homozygous PagHB7a-KO (KO2, KO6) lines were selected for further analysis (Supplementary Fig. S2). First, the short-term salt stress responses of PagHB7a-OE, WT, and PagHB7a-KO plants were tested in liquid-culture with 150 mM NaCl. After 2 h of salt treatment, PagHB7a-KO plants exhibited stem tip bending and leaf curling, whereas WT plants showed only moderate leaf drooping. PagHB7a-OE plants were minimally affected, displaying only slight drooping. This phenotypic divergence became more pronounced after 48 h, with severe wilting and necrosis appearing in most PagHB7a-KO leaves, and WT plants showed noticeable damage. In contrast, PagHB7a-OE plants remained largely undamaged (Fig. 3a).
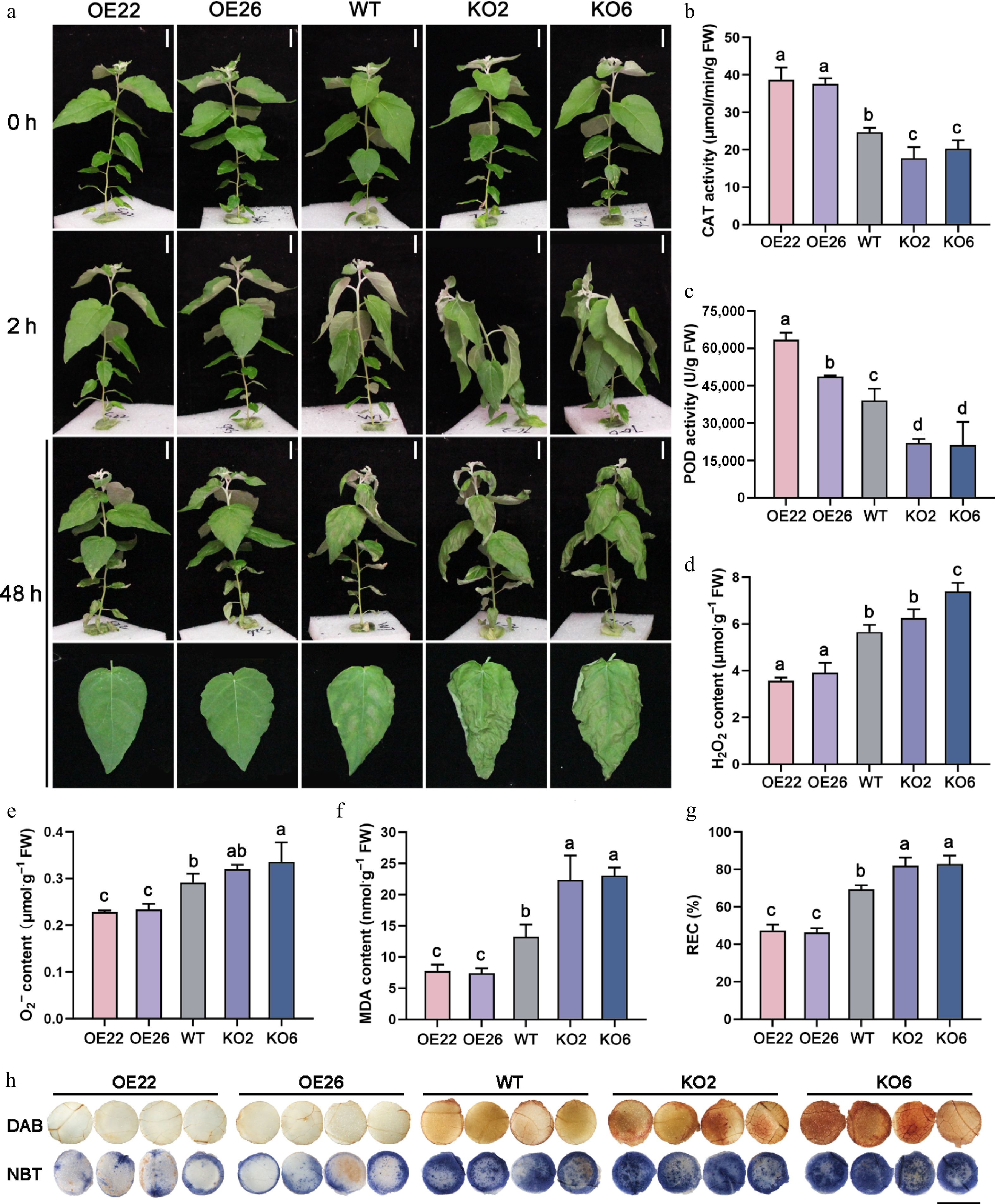
Figure 3.
PagHB7a is a positive regulator of short-term salt tolerance in poplar. (a) Phenotype of PagHB7a-OE, WT, and PagHB7a-KO plants before and after treated with 150 mM NaCl. Scale bars, 5 cm. The (b) catalase activity (CAT), (c) peroxidase activity (POD), (d) H2O2 content, (e) O2·− content, (f) malondialdehyde (MDA) content, and (g) relative electrical conductivity (REC) in PagHB7a-OE, PagHB7a-KO, and WT plants with 150 mM NaCl treatment. Error bars represent ± vSD (n = 3). Statistical significance was determined by one-way ANOVA with Tukey's post hoc test. Different letters represent significant differences. (h) Histochemical staining of reactive oxygen species using DAB (3,3'-diaminobenzidine), and NBT (nitroblue tetrazolium) in leaves of plants under 150 mM NaCl treatment. Scale bars = 6 mm.
Salt stress induces excessive accumulation of reactive oxygen species (ROS), causing oxidative damage to conductive cells and consequently leading to elevated malondialdehyde (MDA) content, and increased relative electrical conductivity (REC)[61]. Thus, MDA content, REC values, and the ROS-scavenging activities of catalase (CAT) and peroxidase activity (POD) serve as key indicators of plant salt tolerance. Quantification analysis found that under salt stress, PagHB7a-OE plants exhibited significantly higher CAT and POD activities, along with markedly lower levels of H2O2, O2·–, MDA, and REC compared to WT plants (Fig. 3b–g). In contrast, PagHB7a-KO plants showed reduced CAT and POD activities, and elevated accumulation of ROS (H2O2 and O2·–), MDA content, and REC (Fig. 3b–g). Additionally, ROS staining analysis was performed on leaf tissues from each plant genotype using 3,3'-diaminobenzidine (DAB) and nitroblue tetrazolium (NBT). Consistently, the leaves of PagHB7a-KO displayed stronger ROS staining while the leaves of PagHB7a-OE displayed weaker staining compared to the WT under salt stress (Fig. 3h). These results suggested that PagHB7a positively regulates salt tolerance in poplar.
To investigate the role of PagHB7a in response to long-term salt stress, we treated soil-grown PagHB7a-OE, WT, and PagHB7a-KO plants with 100 mM NaCl solution. After 20 d of salt treatment, PagHB7a-KO plants exhibited severe wilting and leaf desiccation, while WT plants exhibited moderate wilting. In contrast, the upper and middle leaves of PagHB7a-OE plants exhibited only slight drooping (Fig. 4a). Under control conditions (without NaCl), after 20 d of growth, PagHB7a-OE plants exhibited slight growth inhibition, with plant height reduced by 4.8% and 5.1% compared to WT, whereas PagHB7a-KO plants showed a slight increase in height (Fig. 4b). Prolonged salt stress eliminated these growth differences, resulting in similar plant heights across all lines. At the end of the treatment, several physiological indicators associated with salt stress were measured. These results showed that the PagHB7a-OE plants accumulated greater biomass (fresh weight of stems and leaves, and dry weight of roots) compared to WT under salt stress, while the PagHB7a-KO plants exhibited a reduction in biomass (Fig. 4c, d). Additionally, the chlorophyll content and the relative water content (RWC) in the PagHB7a-OE plants leaves were significantly higher than those in WT leaves under salt stress, whereas PagHB7a-KO plants showed the opposite trend (Fig. 4e, f). Under normal growth conditions (Control), no significant differences were observed in physiological parameters among all tested poplar lines (Fig. 4b–f). Together, these results further support that PagHB7a is a positive regulator of salt tolerance in poplar.
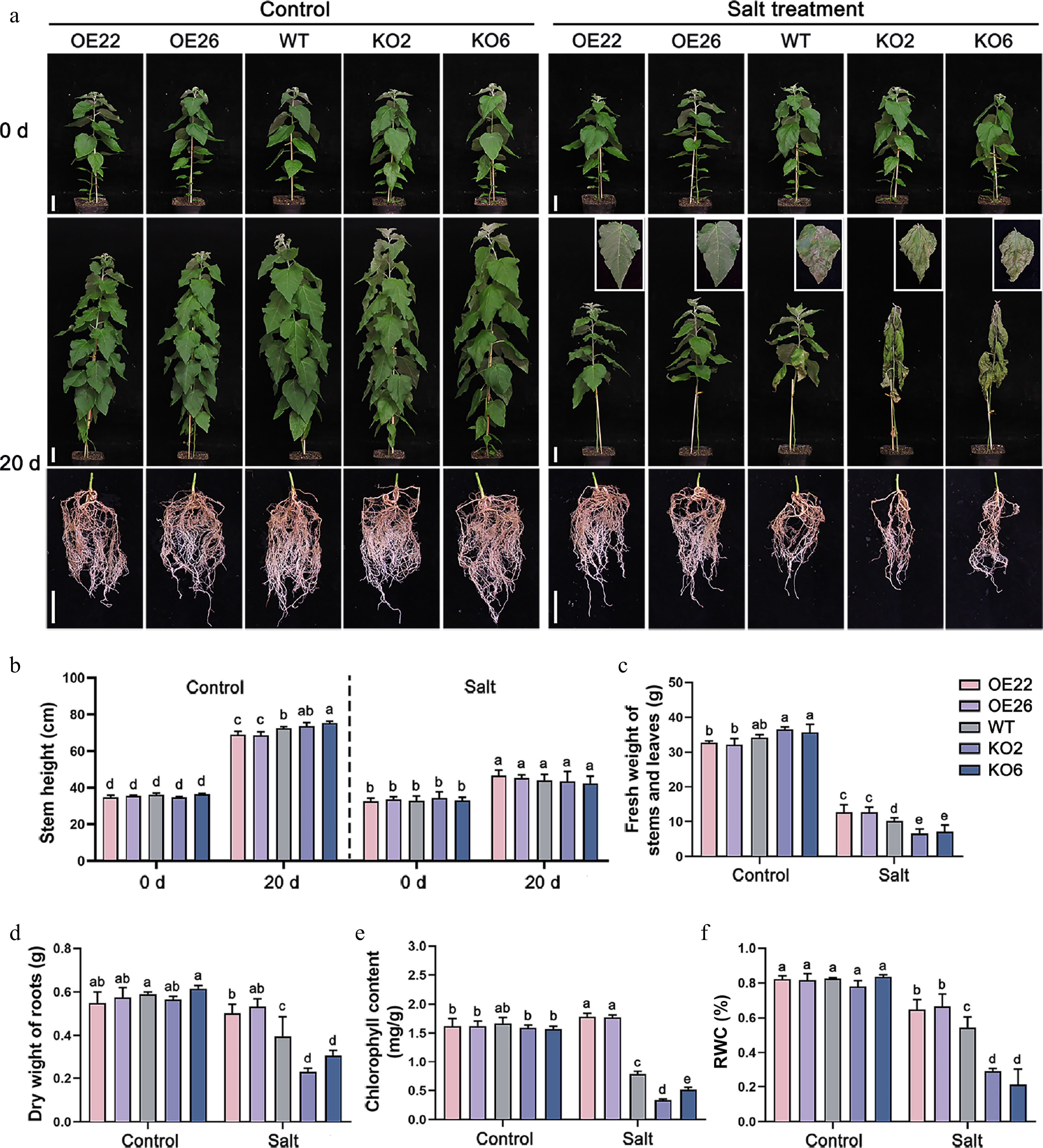
Figure 4.
PagHB7a is a positive regulator of long-term salt tolerance in poplar. (a) Phenotypes of PagHB7a-OE, WT, and PagHB7a-KO plants treated with or without (Control) 100 mM NaCl treatment in soil. Scale bar = 5 cm. (b) Stem height of PagHB7a-OE, WT, and PagHB7a-KO plants under control conditions, or after treatment with 100 mM NaCl treatment. Quantification analysis of (c) fresh weight of stems and leaves, (d) dry weight of roots, (e) chlorophyll content, and the (f) relative water content (RWC) of PagHB7a-OE, WT, and PagHB7a-KO plants under control conditions or after treatment with 100 mM NaCl for 20 d. Statistical significance was determined by one-way ANOVA with Tukey's post hoc test. Different letters represent significant differences. Error bars represent SD values (n = 3).
Given the central role of ABA signaling in abiotic stress responses and the significant induction of PagHB7a by ABA, its function in ABA sensitivity was investigated. Growth assays on medium containing 5 μM ABA revealed that PagHB7a-OE plants were hypersensitive, exhibiting significant reductions in root fresh weight and length, compared to WT (Supplementary Fig. S3a–S3c). Although the PagHB7a-KO plants showed a consistent trend toward reduced sensitivity, the difference was not statistically significant. Collectively, these results suggested that PagHB7a is a positive regulator of ABA signaling.
To assess the impact of PagHB7a on ABA homeostasis, the expression of ABA metabolic genes in the leaves of PagHB7a transgenic and WT plants following ABA treatment were further analyzed. In PagHB7a-OE plants, the expression of ABA biosynthesis genes PagNCED3 and PagNCED5[62] were significantly upregulated compared to WT, while the expression of PagCYP707A1[63], which encodes a key enzyme in ABA oxidative catabolism, remained unaltered (Supplementary Fig. S3d). Conversely, PagHB7a-KO plants exhibited significant upregulation of PagCYP707A1, with no discernible change in PagNCED3 or PagNCED5 expression (Supplementary Fig. S3d). These results suggest that PagHB7a fine-tunes ABA homeostasis through coordinated regulation of both synthetic and degradative pathways.
Transcriptome analysis of PagHB7a overexpression and knockout plants
-
To explore the regulatory network of PagHB7a in response to salt stress, RNA-seq was performed to profile transcriptomic changes in the roots of PagHB7a-OE (OE22), WT and PagHB7a-KO (KO2) plants. A total of 7,035 differentially expressed genes (DEGs) were identified between PagHB7a-OE and WT plants under salt stress (p-value < 0.05), comprising 3,642 upregulated and 3,411 downregulated genes (Fig. 5a, Supplementary Table S2). In contrast, only 304 DEGs were detected between PagHB7a-KO and WT plants under the same condition (p-value < 0.05), including 212 upregulated and 92 downregulated genes (Fig. 5b, Supplementary Table S3). Comparative analysis identified 228 common differentially expressed genes (DEGs) in PagHB7a-OE and PagHB7a-KO plants under salt stress conditions (Fig. 5c, Supplementary Table S4). A comparative analysis was further conducted using DEG sets from both PagHB7a-OE and PagHB7a-KO plants under control and salt-stressed conditions, alongside a reference set of 3,747 salt-responsive DEGs previously identified in the poplar salt-stress transcriptome (Supplementary Table S5)[59]. Integration across these five datasets revealed that 1,480 DEGs from salt-stressed PagHB7a-OE plants, and 89 DEGs from salt-stressed PagHB7a-KO plants overlapped with DEGs in salt-stressed WT plants (Fig. 5d, Supplementary Table S6, S7). Additionally, 488 DEGs were common between control and salt-stressed PagHB7a-OE plants, while 23 DEGs were shared between the two conditions in PagHB7a-KO plants. (Fig. 5d, Supplementary Tables S8, S9).
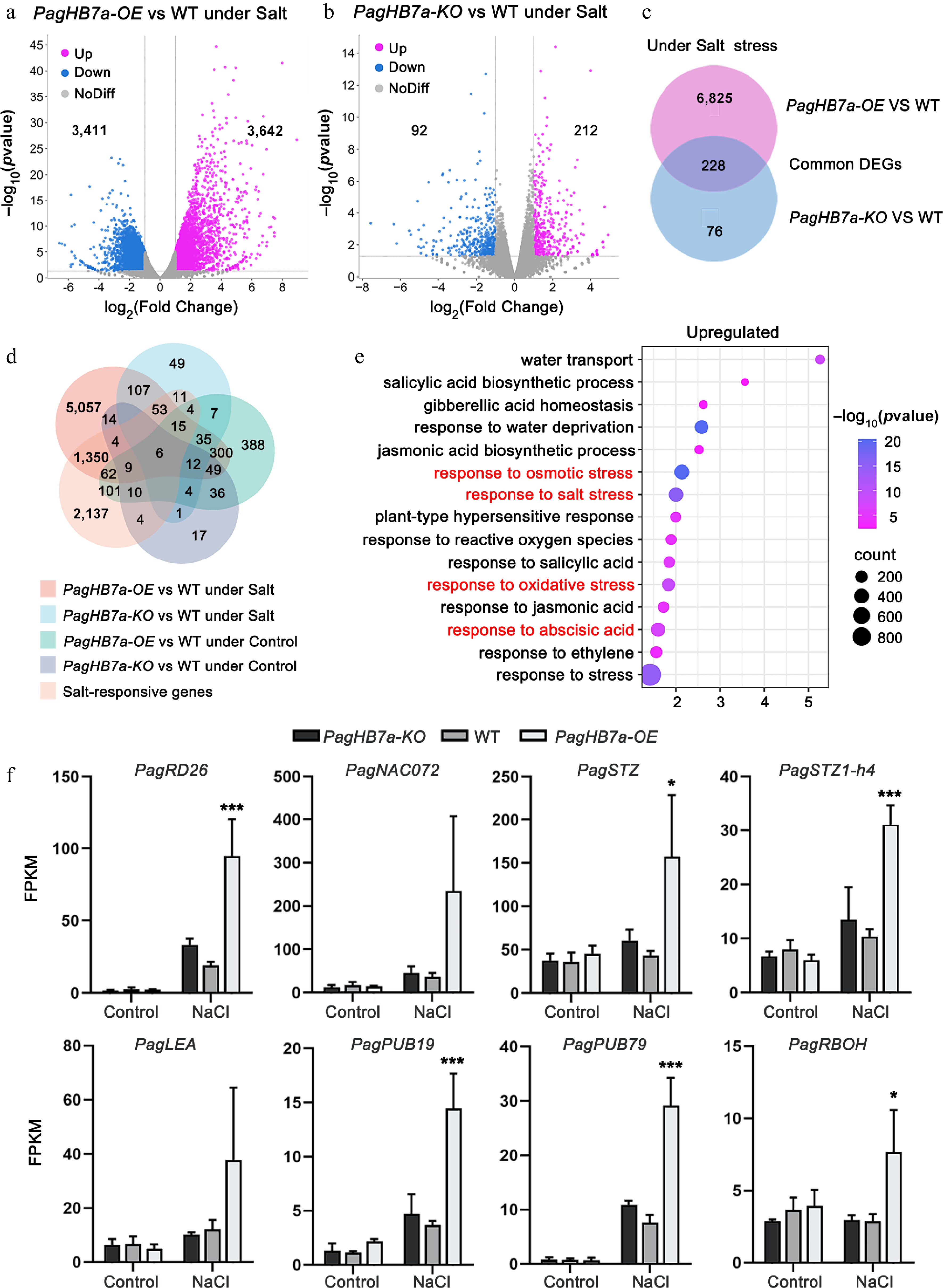
Figure 5.
Transcriptome analysis of PagHB7a-OE, PagHB7a-KO, and WT plants. Volcano plots showing differentially expressed genes (DEGs) in (a) PagHB7a-OE, and (b) PagHB7a-KO plants compared to WT under NaCl treatment (p-adjust < 0.05, log2(Fold Change) ≥ 2.0). The x-axis represents log2 transformed gene expression level, and the y-axis represents the −log10 transformed p-adjust. Each dot represents a single gene. Purple and blue dots denote significantly upregulated and downregulated genes, respectively. Grey dots represent genes without significant changes in expression. (c) Venn diagram illustrating the overlap of DEGs between PagHB7a-OE and PagHB7a-KO plants under NaCl treatment conditions. (d) Venn diagram showing DEGs shared among five comparison groups. (e) Gene Ontology (GO) enrichment analysis of upregulated DEGs in PagHB7a-OE under NaCl treatment. Terms with large gene hit and low p-adjust. The size of the circles indicates the number of hit genes, and the color represents the significance level based on the adjusted p-value. (f) Expression of representative stress-responsive DEGs in PagHB7a-OE, PagHB7a-KO, and WT plants under 100 mM NaCl treatment for 6 h. Gene expression levels are shown as mean FPKM from RNA-seq data. Asterisks denote significant differences compared to values of WT, * p < 0.05; ** p < 0.01; *** p < 0.001.
Gene Ontology (GO) analysis revealed that upregulated DEGs in PagHB7a-OE under salt stress were significantly enriched in pathways related to response to salt stress (GO: 0009651), response to abscisic acid (GO:0009737), response to osmotic stress (GO:0006970), and oxidative stress (GO:0006979) (Fig. 5e, Supplementary Table S10). Consistent with the high salt tolerance of PagHB7a-OE plants, key salt- and osmotic-stress-responsive genes (including PagRD26, PagSTZ, and PagSTZ1-h4)[64,65] were significantly upregulated under salt stress. PagNAC072[66] and PagLEA[67], which were key genes involved in osmoregulation and ion homeostasis, also exhibited a consistent upregulation trend in PagHB7a-OE plants, further supporting the coordinated activation of the stress response network, even though these changes did not reach statistical significance (Fig. 5f). In contrast, PagHB7a-KO plants showed only marginal, non-significant changes in the expression of these genes after NaCl treatment (Fig. 5f). In addition, two genes involved in the ABA signaling pathway (PagPUB19 and PagPUB79)[68,69] were also upregulated in PagHB7a-OE plants, but only induced slightly in PagHB7a-KO plants after NaCl treatment (Fig. 5f). Furthermore, the expression levels of an ortholog of the respiratory burst oxidase homolog (RBOH) gene, a key player in systemic ROS signaling under salinity stress was analyzed. This gene was significantly upregulated in PagHB7a-OE plants, but remained unchanged in PagHB7a-KO plants[70,71] (Fig. 5f). Together, the transcriptional data support the conclusion that PagHB7a positively regulates salt- and ABA-associated genes.
-
Understanding how plants respond to salt stress and balance growth with salt tolerance has long been a major focus of plant research[30,72−76]. HD-ZIP transcription factors are known to play crucial roles in conferring abiotic stress tolerance, with several HB7 genes implicated in the regulation of plant responses to salt and drought stress[28,41,43,77]. In this study, it was demonstrated that PagHB7a is a positive regulator of salt tolerance in poplar. Overexpression of PagHB7a enhanced salt tolerance, while CRISPR/Cas9-generated mutants exhibited increased sensitivity to salt stress (Figs. 3a, 4a).
Abscisic acid (ABA) plays a central role in plant stress perception, particularly in salt-sensitive Populus species[78]. Building on previous identification of salt-responsive regulatory genes and co-expression modules in poplar[59], it was found that PagHB7a expression is significantly induced under both salt and ABA treatments (Fig. 1b–e, Supplementary Fig. S1a), suggesting its potential involvement in ABA-mediated salt stress adaptation. AREB-type transcription factors, members of the bZIP family, typically function in abiotic stress responses through ABA-dependent pathways[79−81], by binding to ABRE cis-elements associated with stress-responsive gene expression[82−84]. In this study, it was demonstrated through yeast one-hybrid (Y1H), ChIP–qPCR, and dual-luciferase reporter assays, that PagAREB1s binds directly to an ABRE-rich segment (P1) in the PagHB7a promoter and activates its expression (Fig. 2a–e). Furthermore, extending earlier reports that PagAREB1s-PagAFP2a forms a negative feedback loop to fine-tune salt-responsive gene activation[27], it is shown that PagAFP2a also suppresses PagAREB1-3-driven transcription of PagHB7a (Fig. 2f).
Reactive oxygen species (ROS) levels, malondialdehyde (MDA) content, relative electrical conductivity (REC), and antioxidant enzyme activities are established biochemical indicators of plant salt tolerance[45,85−87]. Accordingly, these parameters were measured in PagHB7a-OE, PagHB7a-KO, and WT plants under short-term salt stress. Under salt stress, PagHB7a-OE plants exhibited significantly increased catalase (CAT) and peroxidase activity (POD), along with markedly reduced levels of ROS (H2O2 and O2·−), MDA, and REC, whereas PagHB7a-KO plants showed opposite trends (Fig. 3c–h). These findings suggest that PagHB7a enhances the enzymatic antioxidant capacity under salinity, thereby promoting redox homeostasis and membrane stability. This aligns with previous studies showing that modulation of PagHB7a expression influences ROS-scavenging ability and abiotic stress tolerance[28,43,88].
Under normal soil conditions, PagHB7a-OE plants exhibited reduced growth compared to WT after 20 d, whereas PagHB7a-KO plants showed a slight increase in height (Fig. 4b). Notably, long-term salt stress alleviated the growth inhibition observed in PagHB7a-OE plants and the growth enhancement in PagHB7a-KO plants under control conditions (Fig. 4b). These findings suggest that PagHB7a fine-tunes growth responses under salinity, thereby modulating the trade-off between growth and tolerance. Moreover, PagHB7a-OE plants accumulated more biomass in both shoots and roots under salt stress than WT, whereas PagHB7a-KO plants exhibited suppressed growth and increased salt sensitivity, resulting in reduced biomass (Fig. 4b–d). These results strongly suggest that salt-induced PagHB7a plays a critical role in salt tolerance. It should be noted that due to the relatively short duration of the long-term salt treatment applied in this study, the phenotypic differences, although statistically significant, were relatively modest. Further field-based studies under long-term natural saline conditions will be essential to fully elucidate the role of PagHB7a in balancing growth and stress adaptation in poplar.
Through RNA sequencing, genes that were differentially expressed (DEGs) in PagHB7a-OE and PagHB7a-KO plants compared to WT plants under both salt stress and normal growth conditions were identified (Fig. 5a–c). Notably, the significantly lower number of DEGs in PagHB7a-KO plants, compared to PagHB7a-OE plants (304 vs 7,035) under salt stress, suggest functional redundancy within the HD-ZIP I transcription factor family. Given the high sequence similarity (98%), and co-induction under salt stress between PagHB7a and its close homolog PagHB7b (Supplementary Fig. S1), it is speculated that PagHB7b may compensate for the loss of PagHB7a in knockout plants, maintaining expression of critical downstream targets, and thus buffering the transcriptional impact of PagHB7a knockout. Such genetic redundancy is common in plant transcription factor families[89−92] and helps ensure robustness in biological systems. Therefore, although PagHB7a-KO plants exhibit clear salt sensitivity, the full regulatory potential of PagHB7a is more evident from overexpression studies. Gene Ontology (GO) analysis revealed that under salt stress, the upregulated DEGs in PagHB7a-OE plants were significantly enriched in biological processes related to salt stress (GO: 0009651), response to abscisic acid (GO:0009737), response to osmotic stress (GO:0006970), and oxidative stress (GO:0006979) (Fig. 5e, Supplementary Table S10), which is consistent with the observed salt-tolerant phenotype and physiological changes. Furthermore, many genes involved in salt, osmotic, and oxidative stress responses, as well as ABA signaling, were significantly up-regulated in PagHB7a-OE plants under salt stress (Fig. 5f), confirming that PagHB7a acts as a positive regulator in the salt stress response.
A mechanistic model is proposed, illustrating the PagAREB1s-PagHB7a-mediated regulatory network underlying salt tolerance in poplar (Fig. 6). Under salt stress, PagHB7a expression is directly activated by PagAREB1s. The increased PagHB7a protein enhances salt tolerance by boosting ROS scavenging capacity. Furthermore, PagHB7a may also directly regulate certain salt-responsive genes by binding to unidentified cis-elements, thereby promoting transcriptional reprogramming and salinity adaptation. It would be of interest to further investigate whether PagHB7a interacts with other key regulators to form protein complexes that coordinately modulate salt tolerance in poplar.
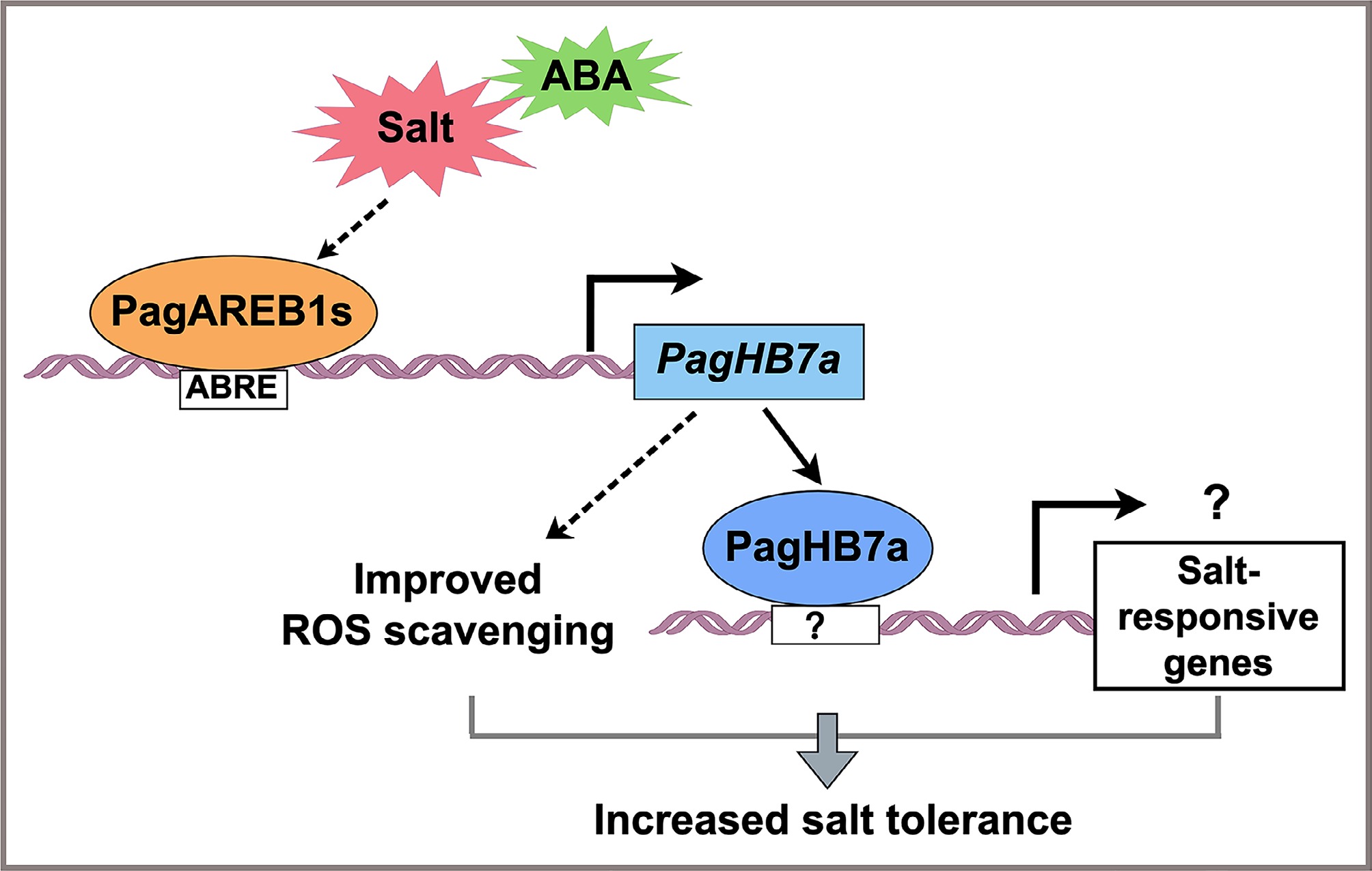
Figure 6.
Proposed model for the regulatory role of PagHB7a in salt stress response. Under salt stress, PagAREB1s, key transcription factors within the ABA signaling pathway, directly bind to the promoter of PagHB7a and activate its expression. The upregulation of PagHB7a enhances reactive ROS scavenging capacity, thereby improving salt tolerance in poplar. Additionally, PagHB7a may also contribute to salinity adaptation through direct activation of unidentified salt-responsive genes.
-
In this study, the HD-ZIP transcription factor PagHB7a was identified as a key positive regulator of salt tolerance in poplar. Its expression is directly activated by PagAREB1s, establishing it as a key downstream component of the ABA signaling pathway. Genetic analyses confirmed that PagHB7a overexpression significantly enhances tolerance to both NaCl and ABA treatment, whereas its knockout increased sensitivity. Furthermore, transcriptome profiling revealed that PagHB7a coordinates a molecular network governing responses to salt stress, ABA signaling, and oxidative stress. Future research should focus on identifying the direct transcriptional targets of PagHB7a and conducting their functional characterization. Overall, this research uncovers a key PagAREB1s-PagHB7a regulatory network, elucidates the molecular mechanism of PagHB7a-mediated salt tolerance, and provides a valuable candidate gene for breeding salt-tolerant poplar varieties.
This work was supported by the Key R&D Program of Shandong Province, China (2024LZGC025).
-
The authors confirm their contributions to the paper as follows: designed the research: Zhou B, Xu N, Liu LJ; performed the experiments and data analysis: Zhou B, Xu N, Yang Z, Sun X, Sun Y, Ji Z; wrote the manuscript: Zhou B, Liu LJ. All authors reviewed the results and approved the final version of the manuscript.
-
The RNA-seq data underlying this article are available in the Genome Sequence Archive (GSA) at https://ngdc.cncb.ac.cn, and can be accessed with CRA032891.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primers used for gene cloning and qPCR.
- Supplementary Table S2 Summary of DEGs between PagHB7a-OE and the WT plants under 100 mM NaCl treatment for 6 h.
- Supplementary Table S3 Summary of DEGs between PagHB7a-KO and the WT plants under 100 mM NaCl treatment for 6 h.
- Supplementary Table S4 The overlapped DEGs between PagHB7a-OE and PagHB7a-KO by 100 mM NaCl treatment for 6 h.
- Supplementary Table S5 Summary of salt-responsive DEGs in WT poplar stems after 100 mM NaCl treatment for 6 h.
- Supplementary Table S6 The overlapped genes between DEGs of PagHB7a-OE vs WT under salt stress and salt-responsive genes.
- Supplementary Table S7 The overlapped genes between DEGs of PagHB7a-KO vs WT under salt stress and salt-responsive genes.
- Supplementary Table S8 The overlapped DEGs between PagHB7a-OE vs WT under control conditions and PagHB7a-OE vs WT under salt stress.
- Supplementary Table S9 The overlapped DEGs between PagHB7a-KO vs WT under control conditions and PagHB7a-KO vs WT under salt stress.
- Supplementary Table S10 Gene Ontology (GO) analysis of the up-regulated DEGs from RNA-seq (PagHB7a-OE under 100 mM NaCl treatment for 6 h).
- Supplementary Fig. S1 Characterization of PagHB7a in Populus.
- Supplementary Fig. S2 Identification of PagHB7a overexpression (PagHB7a-OE) and knockout (PagHB7a-KO) transgenic plants.
- Supplementary Fig. S3 PagHB7a is a positive regulator in response to ABA treatment.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhou B, Xu N, Yang Z, Sun X, Sun Y, et al. 2025. The Class I HD-ZIP transcription factor PagHB7a functions as a positive regulator of salt tolerance in Populus. Forestry Research 5: e030 doi: 10.48130/forres-0025-0030 

The Class I HD-ZIP transcription factor PagHB7a functions as a positive regulator of salt tolerance in Populus
- Received: 28 September 2025
- Revised: 20 November 2025
- Accepted: 03 December 2025
- Published online: 31 December 2025
Abstract: Homeodomain leucine zipper (HD-ZIP) proteins are plant-specific transcription factors that play important roles in plant development and abiotic responses. In our previous study, the PagHB7a gene was identified, which belongs to the Class I HD ZIP family, and was among the most significantly induced genes by salt stress in poplar. In the present study, the role of PagHB7a was functionally characterized in salt stress responses. Expression analysis confirmed that PagHB7a was significantly induced by salt and abscisic acid (ABA) treatments; moreover, PagHB7a was directly regulated by the ABA-responsive element (ABRE) binding proteins (PagAREB1s). Genetic analysis showed that overexpression of PagHB7a (PagHB7a-OE) significantly enhanced salt tolerance, whereas CRISPR/Cas9-mediated knockout of PagHB7a (PagHB7a-KO) significantly reduced it. Transcriptome analysis revealed that biological pathways responding to salt stress, ABA, and oxidative stress were significantly upregulated in PagHB7a-OE plants. Collectively, our results demonstrate that PagHB7a, a salt stress- and ABA-inducible transcription factor, acts as a positive regulator of salt tolerance in Populus.
-
Key words:
- Class I HD-ZIP /
- PagHB7a /
- Salt tolerance /
- Populus