









-
Grapes (Vitis vinifera L.), valued for their nutritional composition and antioxidant content, represent the world's third most important fruit crop with significant economic impact spanning cultivation, processing, and distribution sectors[1], 'Shine Muscat' has emerged as a premium table grape variety, renowned for its distinctive aroma, texture, and nutritional quality[2]. However, its postharvest preservation is challenged by physiological traits, including thin skin, high moisture content, and susceptibility to mechanical damage, leading to water loss, microbial infection, and quality deterioration[3]. Particularly, physical injury during handling promotes Aspergillus niger (A. niger) infection, resulting in decay and potential mycotoxin contamination[4]. Postharvest losses present substantial economic challenges, with global fruit and vegetable losses reaching 20%–50% and grape losses exceeding 30% in regions with inadequate cold chain infrastructure[5]. In China, annual economic losses from postharvest diseases amount to billions of yuan[6].
Current preservation strategies encompass physical[7], chemical[8], and emerging technologies[9], yet face limitations regarding residues, costs, and scalability. This has accelerated research toward sustainable alternatives, including biological preservatives derived from plant and microbial sources[10]. Among these, salicylic acid (SA) and chitosan (CTS) have garnered significant attention. SA, a naturally occurring phenolic phytohormone, is known to regulate plant growth, stress responses, and senescence. In postharvest applications, SA can be absorbed by fruit tissues to systemically induce defense responses, including the activation of antioxidant enzymes and pathogenesis-related proteins, thereby enhancing resistance to oxidative stress and pathogens[11]. CTS, a polysaccharide derived from chitin deacetylation, is approved as a food additive and postharvest treatment in multiple jurisdictions[12]. Its primary mechanism involves forming a semi-permeable edible coating on the fruit surface, which acts as a physical barrier against moisture loss, gas exchange, and pathogen penetration. Additionally, its inherent cationic properties can directly disrupt fungal cell membranes, exhibiting intrinsic antimicrobial activity. Although both SA and CTS have demonstrated preservative effects in various fruits[13−15], their application to 'Shine Muscat' grapes under commercial storage conditions, and a comparative analysis of their distinct modes of action remains insufficiently studied.
To address existing research gaps, this study aims to elucidate the mechanisms by which separately applied SA and CTS coatings preserve 'Shine Muscat' grapes under two typical storage temperatures (4 and 26 °C), by examining their respective impacts on antimicrobial activity, storage quality, and the antioxidant system. The research seeks to: (1) determine the optimal preservative concentration; (2) characterize the physiological and biochemical response pathways induced by the treatment, and (3) provide theoretical insights and a practical framework for developing sustainable grape preservation technologies. These findings indicate that SA and CTS, though applied separately, collectively contribute to a coordinated enhancement of fruit resilience by simultaneously targeting interconnected deterioration pathways.
-
Shine Muscat grapes were harvested from the experimental base of Yangtze University. The three-year-old plants were cultivated using a continuous trellis system under rain-shelter cultivation. Only healthy fruit clusters with consistent morphological indices, including cluster mass (1.5 ± 0.2 kg), and absence of mechanical damage, pests, or diseases, were selected. On the day of harvest, the samples were transported to the laboratory under cold-chain conditions. After standardized processing, clusters were trimmed into 800 g standardized units and horizontally pre-cooled at 4 °C for 24 h before subsequent experimental steps.
A. niger (preserved at the College of Horticulture and Gardening, Yangtze University), CTS (deacetylated ≥ 90%), SA, glacial acetic acid, sodium hydroxide, ascorbic acid, and assay kits for peroxidase and superoxide dismutase were purchased from Nanjing Jiancheng Technology (Nanjing, China).
Edible coatings treatment and storage conditions
-
A 1% glacial acetic acid solution was prepared. CTS powder was dissolved in the solution to obtain final concentrations of 1.0%, 1.5%, and 2.0%. The mixture was stirred continuously at 50 °C in a water bath until fully dissolved, and then cooled to room temperature. Separately, SA powder was dissolved in anhydrous ethanol and diluted to concentrations of 0.5, 1.0, and 1.5 mmol/L, maintaining a final anhydrous ethanol concentration of 0.05% to prevent phytotoxicity. The pH of all solutions was adjusted to 6.5 using NaOH or HCl. The grape samples were randomly divided into seven groups and immersed in the following solutions for 3 min: CK (ddH2O), SA (0.5, 1.0, 1.5 mmol/L), and CTS (1.0%, 1.5%, 2.0%). Each treatment group consisted of three replicates, with each replicate containing 10 bunches of grapes and 200 individual berries. After treatment, the samples were drained and placed on a sterile workbench to air dry naturally. Once a uniform coating had formed on the surface, the samples were stored under two constant temperature conditions: room temperature (26 °C), and refrigerated temperature (4 °C).
In vitro antifungal activity of SA and CTS against A. niger
-
To evaluate the in vitro inhibitory effects of SA and CTS on A. niger, the agar plate diffusion method was used. Potato dextrose agar (PDA) plates were prepared by incorporating various concentrations of SA (0.5, 1.0, and 1.5 mmol/L), and CTS (1.0%, 1.5%, and 2.0%) into the medium prior to solidification. A spore suspension of A. niger was adjusted to a concentration of 1 × 105 spores/mL, and centrally inoculated onto each prepared plate. The inoculated plates were then incubated at 25 °C. Colony diameters were regularly measured to calculate the mycelial growth inhibition rate. After 5 d of incubation, hyphal morphology was examined under a microscope. Each treatment was performed in triplicate, and the entire experiment was independently repeated three times.
Colony diameter (cm) = Control group diameter (cm) − Mycelial plugs (cm)
Inhibition rate = (Control group colony diameter − Treatment group colony diameter)/Control group colony diameter × 100%
In vivo efficacy of SA and CTS against A. niger infection in grapes
-
The A. niger spore suspension was prepared as described earlier. Grape berries, pretreated according to the method outlined earlier, were punctured with a sterile needle and inoculated with 10 µL of the spore suspension per berry. The inoculated grapes were then incubated at 25 °C to assess the inhibitory effect. Each treatment was performed with three biological replicates, each consisting of 30 grape berries. The lesion area (cm2) was measured using ImageJ software.
Measurement of quality and biochemical parameters in grapes
Determination of decay, weight loss, and fruit abscission rates
-
Measurements were taken every 7 d throughout the experiment. The cumulative decay rate (DR), weight loss rate (WLR), and fruit abscission rate (FAR) were assessed using a counting method for both the treatment and control groups. Each treatment consisted of 10 fruit clusters, with three replicates.
Determination of total soluble solids and titratable acidity
-
Total soluble solids (TSS) were measured using an ATAGO PAL-BX/ACID2 refractometer. Ten berries were randomly selected per biological replicate. Grape pulp was homogenized in a juicer, and the extract thoroughly mixed. TSS was determined immediately in triplicate, and the arithmetic mean was recorded. Titratable acidity (TA) was determined by acid-base titration: 5.0 g of pulp was homogenized in a mortar, transferred to a 50 mL volumetric flask, dilute to volume with ddH2O, and allowed to stand for 30 s. The solution was filtered, and 20 mL of filtrate was mixed with 2−3 drops of phenolphthalein and titrated with 0.1 mol/L NaOH to a persistent pink endpoint (≥ 30 s). Titration was repeated three times, TA was calculated from the average NaOH consumption. Each treatment consists of 10 grapes, repeated three times.
Determination of total soluble protein and ascorbic acid of grapes
-
Total soluble protein (TSP) was quantified via the Coomassie Brilliant Blue method[16]. Briefly, 1 g grape tissue (FW) was homogenized in 10 mL ddH2O, diluted to 50 mL, and centrifuged (5,800 × g, 5 min). The supernatant (0.5 mL) was mixed with 5 mL Coomassie Brilliant Blue reagent, incubated for 5 min, and absorbance was measured at 595 nm. Triplicate measurements were averaged (mg/100 g FW). Ascorbic acid (AsA) content was determined using a commercial kit (Nanjing Jiancheng Technology, China), following the manufacturer’s protocol. Absorbance (525 nm) was recorded on a microplate reader, with triplicate values expressed as mean ± SD (mg/100 g FW). Each treatment consists of 10 grapes, repeated three times.
Determination of antioxidant enzyme activities and MDA content
-
Malondialdehyde (MDA) was measured using the thiobarbituric acid dual-wavelength method[17], with absorbance readings taken at 532 and 600 nm. Catalase (CAT) activity was determined by UV absorption at 240 nm[18]. Peroxidase (POD) and superoxide dismutase (SOD) activities were assessed using commercial assay kits (Nanjing Jiancheng Technology, China) according to the manufacturer’s instructions, with absorbance measured at 420 nm for POD, and 550 nm for SOD using a microplate reader. All assays were performed in triplicate, and enzyme activities are expressed as U/g FW, where one unit (U) corresponds to the amount of enzyme that catalyzes 1 μmol of substrate per min.
RNA extraction and cDNA synthesis
-
Total RNA was extracted using the TaKaRa MiniBEST Plant RNA Extraction Kit (Takara Bio, Dalian, China). Fresh grapes tissues were rapidly ground into a fine powder in liquid nitrogen. All subsequent procedures were performed on ice to minimize RNA degradation, strictly following the manufacturer's instructions.
cDNA for quantitative real-time PCR (qPCR) was synthesized from the extracted RNA using the HiScript III 1st Strand cDNA Synthesis Kit (+gDNA wiper), (Vazyme Biotech, Nanjing, China). The resulting cDNA was appropriately diluted with sterile nuclease-free water, aliquoted, and stored at −40 °C to avoid repeated freeze-thaw cycles.
Quantitative real-time PCR (qRT-PCR) analysis
-
To elucidate the molecular mechanisms of CTS and SA treatments, we analyzed the expression of key genes involved in three critical postharvest pathways: antioxidant defense (VvCAT, VvAPX, VvGR, VvPOD), phenylpropanoid metabolism and enzymatic browning (VvPAL, VvPPO), senescence and membrane lipid peroxidation (VvLOX), and pathogen defense (VvBGLU, VvGST)[19]. These target genes were selected based on their well-established functional roles in grape stress responses and postharvest physiology. To ensure amplification specificity for the intended gene isoforms—given the presence of multigene families—all primer pairs (Supplementary Table S1) were designed to target unique, conserved coding sequences, with specificity verified using the BLAST tool on the NCBI website (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ).Quantitative real-time PCR was performed using the ChamQ Universal SYBR qPCR Master Mix (Vazyme Biotech Co., Ltd., Nanjing, China), strictly following the manufacturer's protocol. The amplification protocol consisted of an initial denaturation at 95 °C for 30 s, followed by 40 cycles of denaturation at 95 °C for 10 s and annealing/extension at 60 °C for 15 s. A melt curve analysis was subsequently conducted by increasing the temperature from 60 to 95 °C with an increment of 0.5 °C every 6 s. The relative expression levels of target genes were calculated using the 2−ΔΔCᴛ method. For each gene, the assay was performed with three biological replicates.
Data analysis
-
Experimental data was organized using Microsoft Excel 2019. Statistical analyses, including correlation analysis, principal component analysis (PCA), and cluster analysis were performed using SPSS 27 (IBM Corp., USA). Data was analyzed by one-way analysis of variance (ANOVA), and differences between means were compared using Duncan's new multiple range test at a significance level of p < 0.05. Data visualization was conducted using Origin 2024 (Origin Lab Corp., USA).
-
To investigate the inhibitory effects of SA and CTS on the growth of A. niger, this study evaluated the mycelial growth and inhibition rates under different concentrations of SA and CTS. Plate inhibition assay results indicated that both SA and CTS treatments inhibited the mycelial expansion of A. niger in a concentration-dependent manner (Fig. 1a, d). The control group exhibited vigorous colony growth with neat edges and dense mycelia, covering the entire plate after 5 d of cultivation, while all treatment groups showed significantly inhibited colony growth. The SA2 and CTS3 treatments demonstrated the most notable inhibitory effects, with inhibition rates reaching 74.77% and 86.26%, respectively (Fig. 1e). Microscopic observation revealed normal mycelial morphology in the control group, characterized by intact conidial heads and chain-like spherical conidia, with mature conidia appearing carbon-black due to pigment deposition. The SA treatment group showed a significant reduction in the number of mature conidia, whereas the high-concentration CTS treatment group (e.g., CTS3) exhibited obvious mycelial malformation, featuring localized swelling and constriction that formed numerous nodular structures (Fig. 1b), thereby impeding mycelial growth and indicating that CTS not only delayed, but substantially inhibited mycelial development. In vivo inoculation experiments further confirmed the efficacy of SA and CTS in controlling postharvest rot of grape fruits caused by A. niger (Fig. 1c). Rot symptoms began to appear in the control group 2 d after inoculation. By day 6, the lesion area in the control group reached 1.47 cm2, while those in the SA2 and CTS3 treatment groups were only 0.21 and 0.19 cm2, respectively (Fig. 1f), demonstrating that SA and CTS treatments effectively suppressed A. niger-induced fruit rot in grapes.
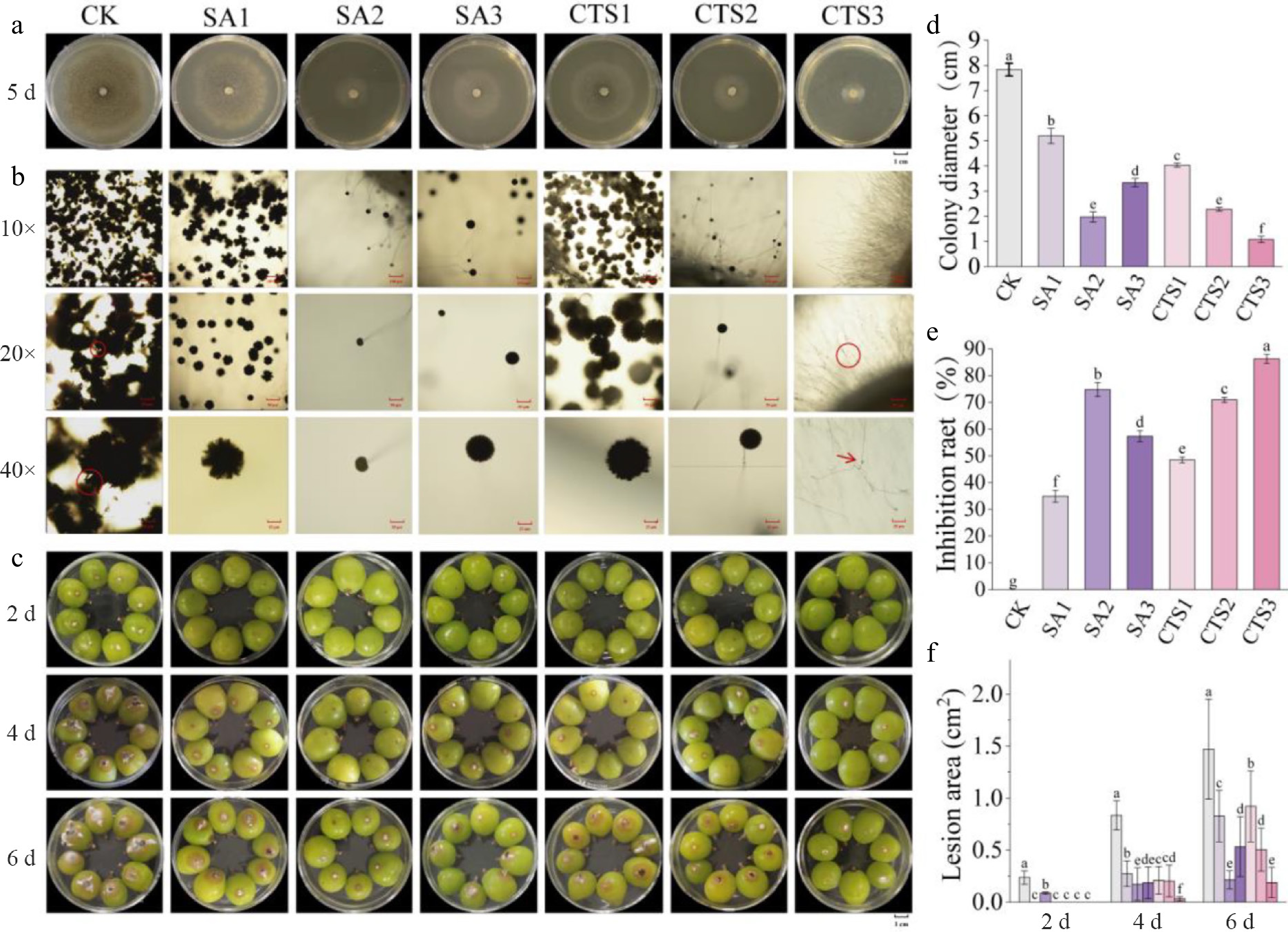
Figure 1.
Antifungal activity of SA and CTS against Aspergillus niger. (a) Inhibition of A. niger mycelial growth by SA and CTS; (b) effects of SA and CTS treatments on microscopic morphology of A. niger spores; the arrow is pointing to a pathological nodule. (c) To observe pathogenicity, grapes were inoculated with A. niger; (d) colony diameter; (e) inhibition rate; (f) lesion area. Data is presented as mean ± SD, n = 3. Different letters indicate significant (p < 0.05) differences between the control and treated samples.
Effects on external quality parameters
Reduction of grape decay rate by SA and CTS treatments
-
The CTS3 treatment was the most effective in controlling grape decay, resulting in the lowest recorded DR at both 4 °C (0.96%), and 26 °C (8.81%) after 42 d of storage (Fig. 2a, b). Throughout the storage period, all treatment groups maintained a consistently lower DR than the control (CK). At 4 °C, decay onset was delayed until day 21 in treated groups, while the CK showed a significantly higher DR, reaching 11.97% by day 42. At 26 °C, although decay was initiated by day 14 in all treatments, the CK DR reached 27.63% at the endpoint, substantially higher than the maximum observed in any treatment group.
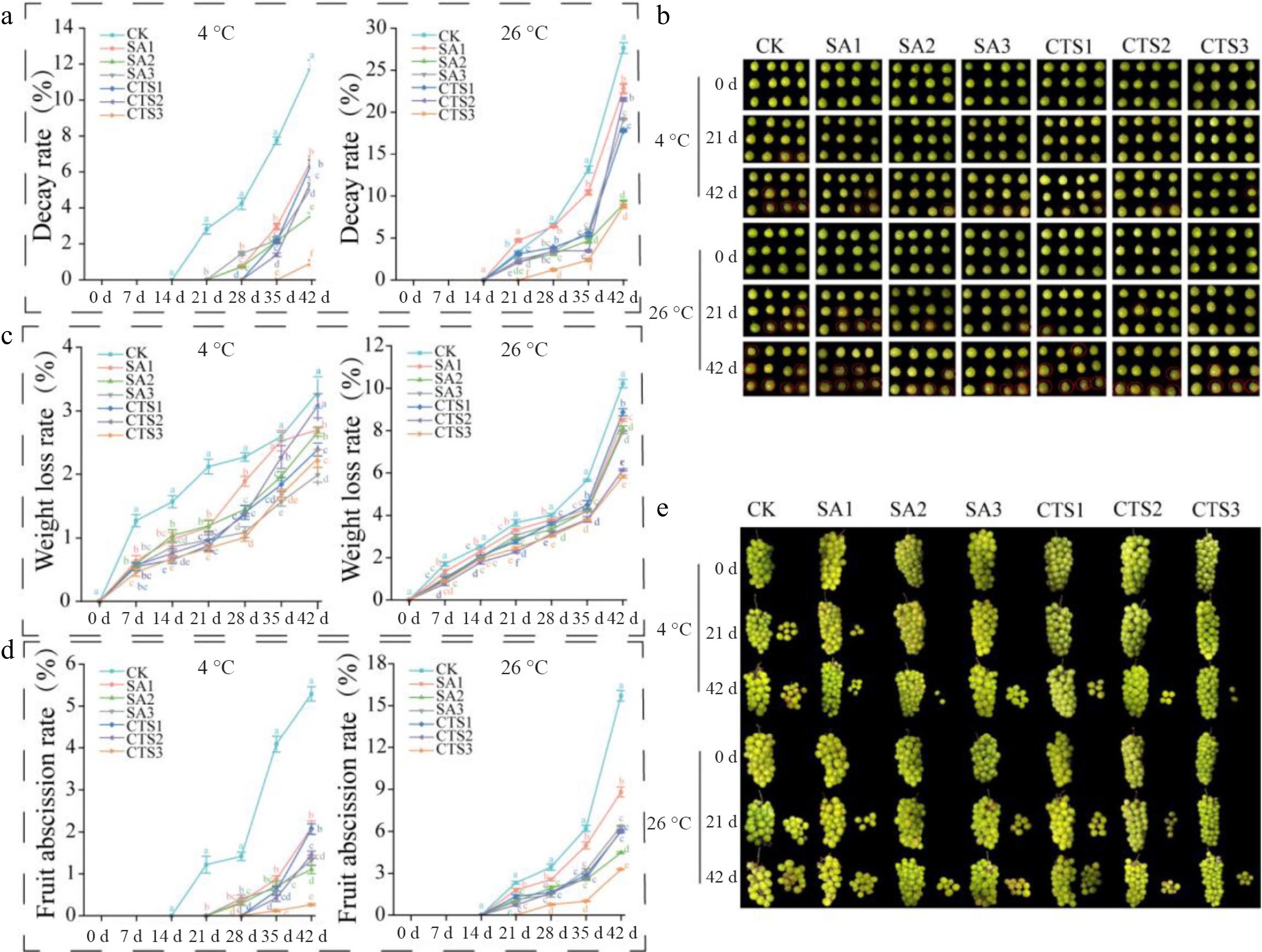
Figure 2.
Effects of different treatments on the external quality of grapes during storage. (a) Decay rate; (b) changes of fruit decay rate in control group and treatment group during storage; (c) weight loss rate; (d) fruit abscission rate; (e) changes of fruit abscission rate in the control group and treatment group during storage. Data is presented as mean ± SD, n = 3. Different letters indicate significant (p < 0.05) differences between the control and treated samples.
Preservation treatment significantly reduces the weight loss rate of grapes
-
The optimal treatment for minimizing grape WLR was found to be temperature-dependent. At 4 °C, SA3 and CTS3 were most effective, limiting WLR to 1.99% and 2.25%, respectively, after 42 d. In contrast, at 26 °C, CTS3 (5.83%) provided the best protection, reducing the WLR by 4.39% absolutely, compared to the CK (10.22%). Overall, all treatments significantly suppressed WLR relative to the CK throughout storage, despite a general increasing trend over time (Fig. 2c).
Preservation treatment alleviates fruit abscission in grapes
-
Grape FAR exhibited a progressive increase throughout the storage period (Fig. 2e). After 42 d, the efficacy of the treatments was temperature-dependent. At 4 °C, the FAR across groups followed the descending order: CK > SA1 > CTS1 > CTS2 > SA3 > SA2 > CTS3. In contrast, at 26 °C, the SA2 and CTS3 treatments resulted in the lowest FAR values (4.5% and 3.3%, respectively; Fig. 2d). Notably, the CTS3 treatment consistently demonstrated the greatest efficacy in minimizing fruit abscission at both storage temperatures.
Effects on internal quality parameters
Effect of different treatments on changes in TSS in grapes during storage
-
TSS consists of water-soluble sugars, organic acids, vitamins, and minerals, serving as a key indicator of fruit and vegetable juice quality. During storage, chemical degradation and molecular transformations of these components adversely affect key fruit quality attributes. Smaller changes in TSS before, and after storage indicate better preservation efficacy. Over extended storage periods, TSS levels in the fruit generally exhibited a gradual upward trend (Fig. 3a). After 42 d, the CK treatment showed the most significant TSS increase, while the SA3 treatment exhibited minimal change. At 26 °C, TSS levels increased more markedly. After 42 d, the CK group had substantially higher TSS than the SA and CTS treatments, whereas the SA3 treatment maintained the smallest TSS fluctuation.
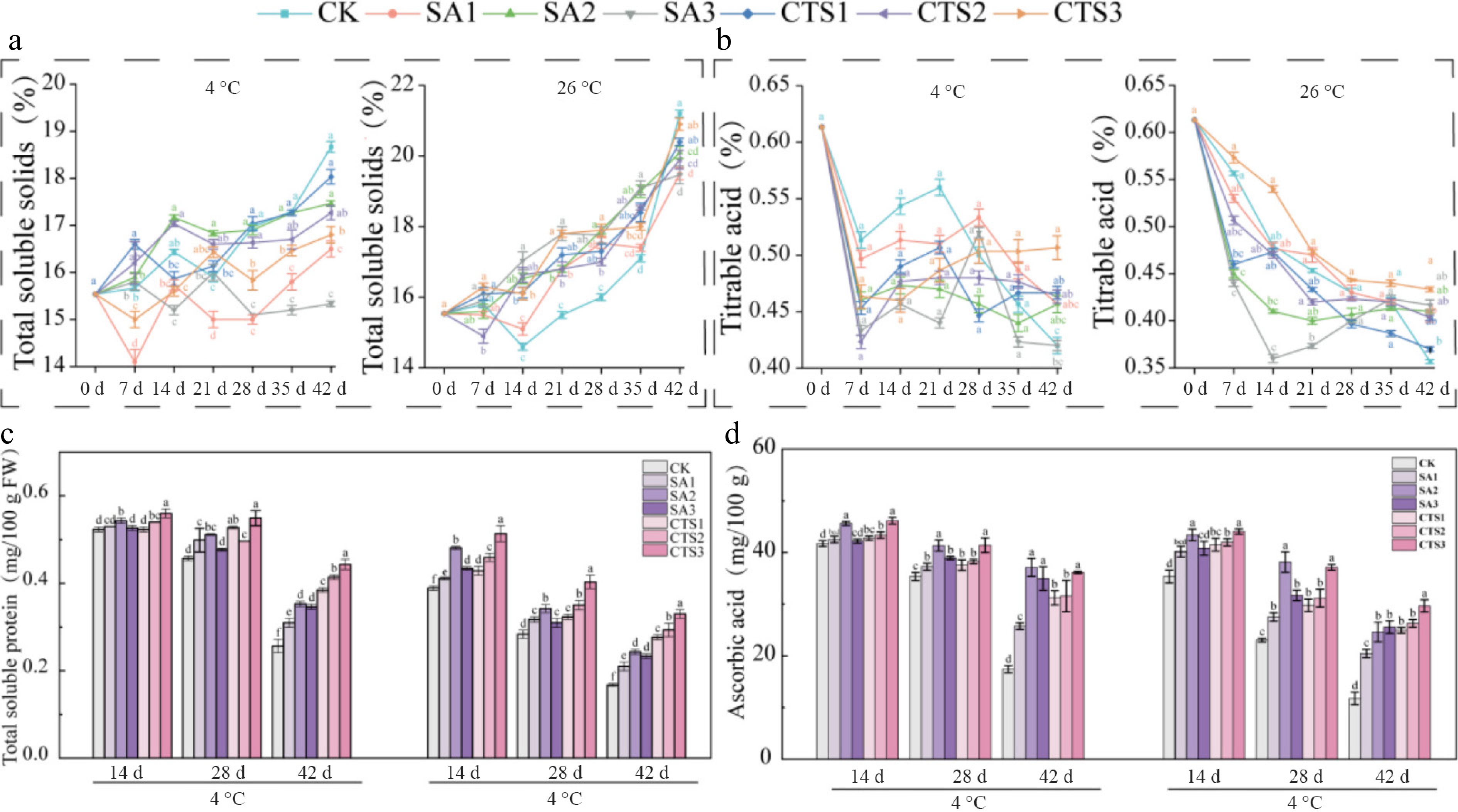
Figure 3.
Effects of different treatments on the internal quality of grapes during storage. (a) Total soluble solids content; (b) titratable acid content; (c) total soluble protein; (d) ascorbic acid. Data is presented as mean ± SD, n = 3. Different letters indicate significant (p < 0.05) differences between the control and treated samples.
Variations in grape TA under different treatments
-
TA, a key intrinsic trait influencing grape berry flavor, decreased consistently across all treatments under both storage conditions (Fig. 3b). At 4 °C, after 42 d of storage, TA decreased by 0.02% (SA1), 0.05% (SA2), and 0.01% (SA3), in the SA treatment group. The CTS treatment group showed slightly greater decreases: 0.03% (CTS1), 0.03% (CTS2), and 0.01% (CTS3). At 26 °C, decreases in the SA group were 0.25% (SA1), 0.23% (SA2), and 0.25% (SA3), and in the CTS group were 0.24% (CTS1), 0.22% (CTS2), and 0.19% (CTS3). Notably, the CK treatment retained the highest TA level after 42 d at both temperatures.
Temperature-dependent efficacy of preservation treatments in maintaining TSP stability
-
TSP, a key biochemical marker associated with fruit disease resistance, ripening, and senescence, decreased during storage across all treatments (Fig. 3c). At 4 °C, the CTS3 treatment maintained the highest TSP content (0.44 mg/100 g) after 42 d, which was significantly higher than that in other treatments. At 26 °C, CTS3 resulted in the highest TSP content (0.33 mg/100 g) after the same period, differing significantly from the other groups. The CK group showed the most rapid decline in TSP. Both SA and CTS treatments significantly retarded TSP degradation compared with CK. These results suggest that the efficacy of all experimental treatments is temperature-dependent: it helps preserve protein content by inhibiting hydrolase activity at 4 °C, whereas at 26 °C, accelerated metabolism likely outweighs the protective effect. Again, CTS3 resulted in the highest TSP at both temperatures.
SA and CTS coatings retarded the degradation of AsA
-
AsA, a major antioxidant in berries, is highly susceptible to degradation during storage. Throughout the storage period, all treatments exhibited a continuous decline in AsA content, with the CK group showing a significantly faster reduction than the SA and CTS treatments (Fig. 3d). After 42 d at 4 °C, the SA2 and CTS3 treatments retained markedly higher AsA contents (37.13 and 36.15 mg/100 g, respectively) than the other groups. At 26 °C, SA2 maintained the highest AsA level (38.16 mg/100 g) on day 28, while CTS3 showed the highest content (29.67 mg/100 g) on day 42. Overall, low-temperature storage effectively slowed the degradation of AsA, and both SA and CTS coatings significantly reduced AsA loss compared with CK.
Effects on oxidative damage and antioxidant enzymes
Suppression of MDA accumulation by SA and CTS coatings
-
MDA concentration, an indicator of membrane lipid peroxidation, reflects oxidative damage severity during fruit storage. Under both 4 and 26 °C storage, grape MDA concentrations increased over time. However, the increase was significantly lower in film-coated (SA, CTS) groups than in the uncoated CK (Fig. 4a). At 4 °C, after 42 d, the CTS3-treated group showed the lowest MDA concentration (0.25 U/g FW). At 26 °C, the CK group exhibited significantly higher MDA than SA and CTS groups. CTS3 treatment performed best, showing the most effective suppression of MDA accumulation. Collectively, both CTS3 and SA2 coatings effectively inhibited grape membrane lipid peroxidation, with CTS3 demonstrating the strongest inhibitory effect.
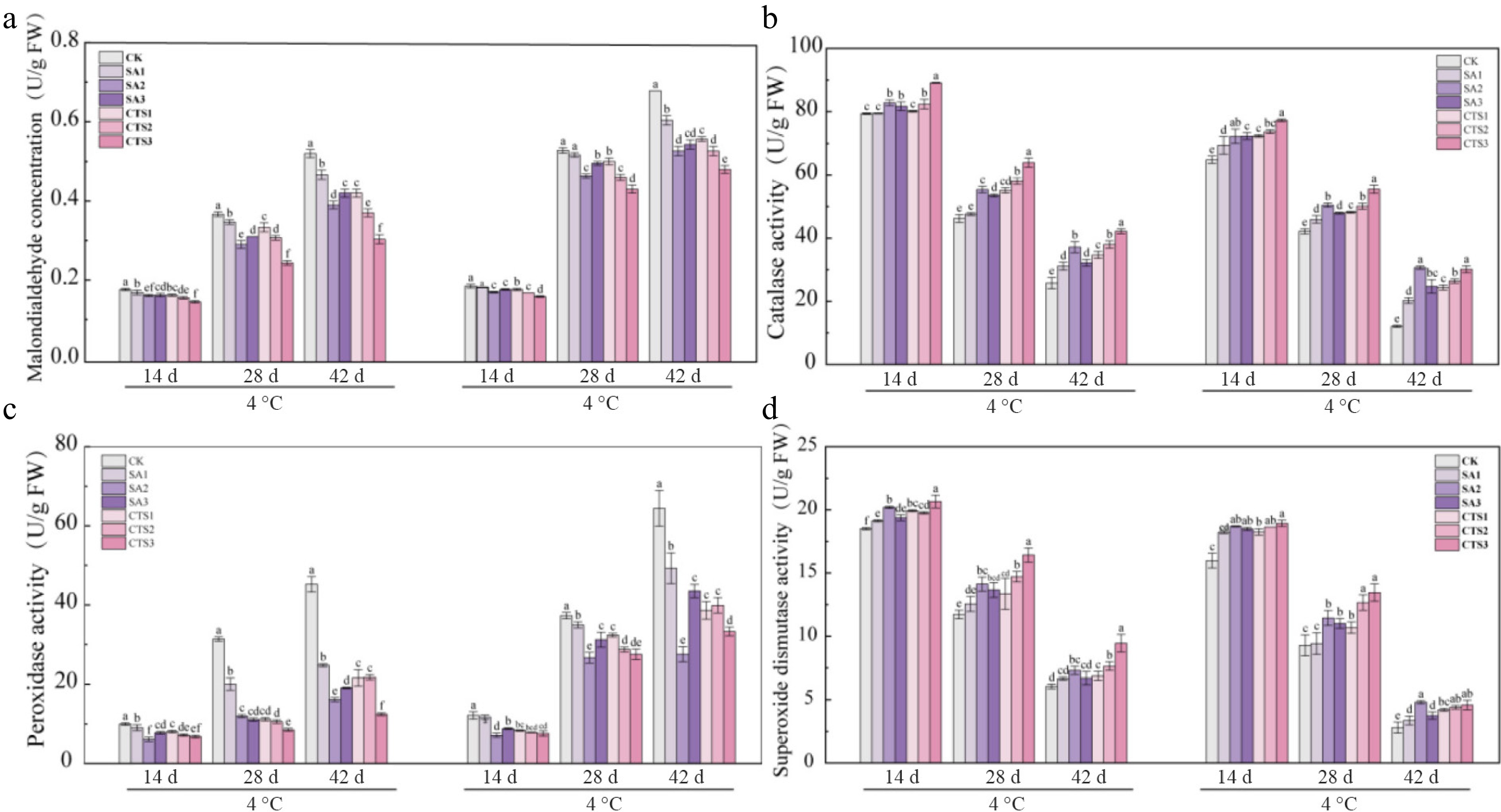
Figure 4.
Modulation of grape antioxidant defense by chitosan and salicylic acid coatings during storage at 4 and 26 °C. (a) Malondialdehyde concentration; (b) catalase activity; (c) peroxidase activity; (d) superoxide dismutase activity. Data is presented as mean ± SD, n = 3. Different letters indicate significant (p < 0.05) differences between the control and treated samples.
Effects of SA and CTS coatings on antioxidant enzyme activities
-
The activities of key antioxidant enzymes (CAT, POD, and SOD), were modulated by both SA and CTS treatments in a temperature-dependent manner (Fig. 4b–d). At 4 °C, the CTS3 treatment showed the most pronounced effects. It maintained the highest activities of CAT (42.21 U/g FW), and SOD (9.45 U/g FW) at the end of storage (day 42). Concurrently, this treatment most effectively suppressed the rise in POD activity, resulting in the lowest final POD activity (6.83 U/g FW), which was 63% lower than that in the control (CK) group. At 26 °C, the SA2 treatment exhibited the strongest regulatory effect. It sustained the highest SOD activity (4.81 U/g FW) throughout storage, and maintained POD activity at a relatively low level (27.52 U/g FW) compared to the sharp increase observed in the CK group. Furthermore, the SA2 treatment induced an earlier peak in CAT activity during the storage period. In summary, the CTS3 treatment most effectively enhanced the activities of CAT and SOD while suppressing POD activity at 4 °C. In contrast, the SA2 treatment was most effective in modulating these enzyme activities at 26 °C. Both treatments mitigated the changes in antioxidant enzyme activities observed in the control group, contributing to reduced oxidative stress in the grapes during storage.
Correlation analysis of physiological and biochemical indices during postharvest storage of grapes
-
Data standardization was performed on the original values of 11 physiological and biochemical indicators (DR, WLR, FAR, TSS, TA, TSP, AsA, MDA, CAT, SOD, and POD) using SPSS 27 to enable comparative analysis across different measurement units (Fig. 5a, b). Correlation analysis of these indicators under both 4 and 26 °C storage conditions identified a highly significant positive correlation between DR and FAR (r = 0.97 at 4 °C), and a significant negative correlation between CAT and MDA (r = −0.98 at 4 °C). Consequently, these correlations demonstrate a functional interplay between oxidative stress (represented by MDA and CAT) and water metabolism (represented by WLR and FAR) in postharvest grapes.
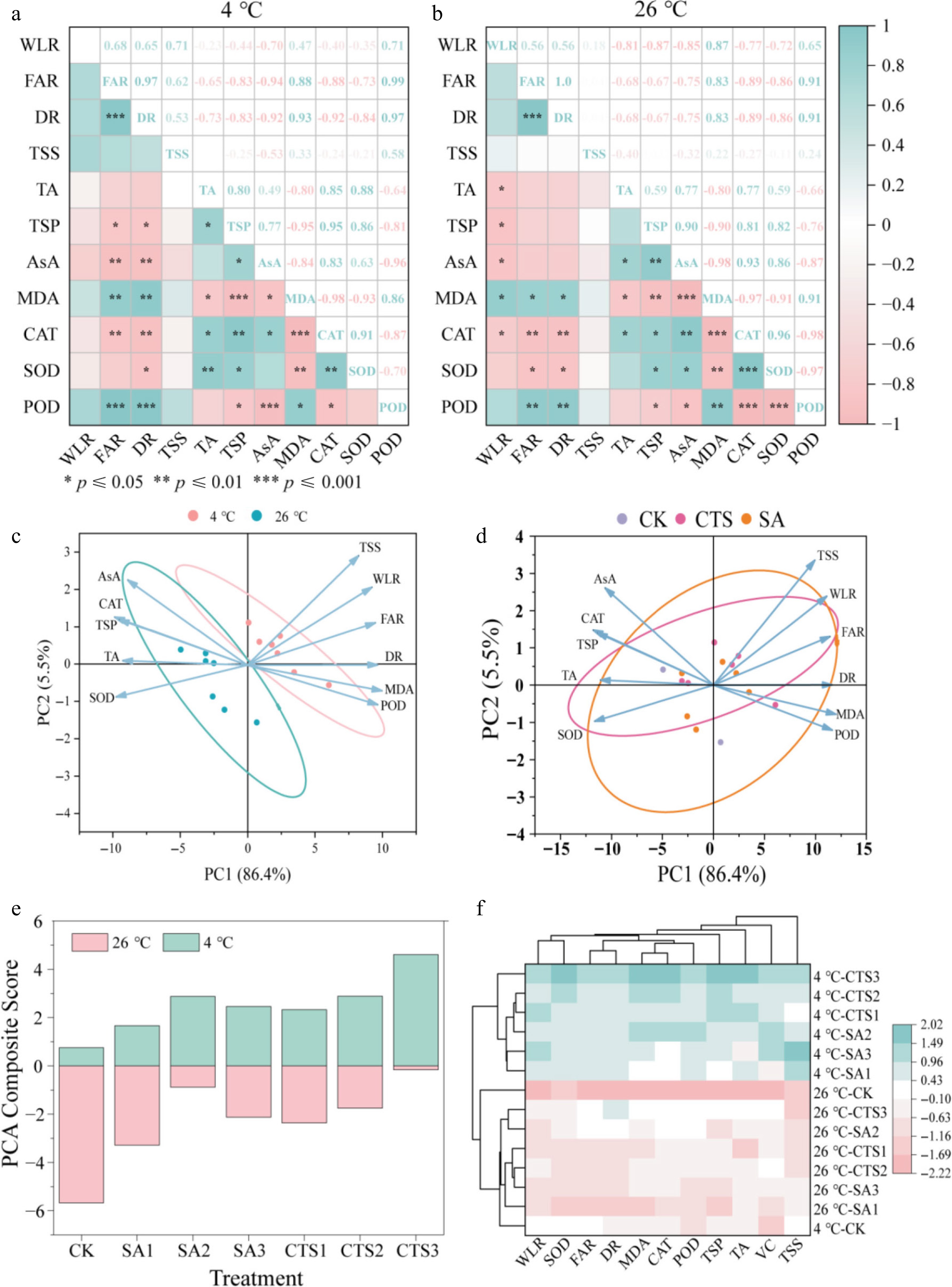
Figure 5.
Multivariate analysis of grape preservation mechanisms. (a) Correlation analysis; (b) PCA effect of storage temperature (4 °C) on storage quality of grapes during storage; (c) PCA effect of storage temperature (26 °C) on storage quality of grapes during storage; (d) PCA of grape storage quality with different coating treatments under two storage temperatures; (e) PCA composite score; (f) cluster analysis of different coating treatments and storage temperature of grapes.
Principal component analysis of preservation efficacy
-
PCA of 11 quality indicators revealed that coating preservation efficacy for grapes was predominantly governed by two components explaining 91.9% of the total variance (Fig. 5c, d). PC1, associated with external quality, showed positive loadings for TSS, WLR, FAR, DR, MDA, and POD, and negative loadings for AsA, TSP, TA, CAT, and SOD. Quantitative evaluation based on weighted PCA scores confirmed that preservation efficiency was significantly greater at 4 than at 26 °C for all treatments (Fig. 5e). Notably, the CTS3 coating consistently achieved the highest comprehensive score across both storage temperatures, identifying it as the most effective treatment, followed by SA2.
Cluster analysis identifies temperature as the primary factor in grape storage
-
To decipher the complex relationships governing grape storage quality, we performed a cluster analysis on standardized data from 11 quality indicators (Fig. 5f). This analysis synthesizes multidimensional data into intuitive groupings, overcoming the limitation of any single index in fully describing postharvest dynamics. The results clearly established storage temperature as the primary determinant of shelf life. Furthermore, the CTS3 and SA2 treatments formed a distinct cluster associated with superior storage performance, primarily by effectively reducing water loss-related indices and enhancing the antioxidant defense system compared to other treatments.
Gene expression profiles and correlation with antifungal efficacy
-
Based on comprehensive in vitro antifungal tests, in vivo inoculation validation, and multivariate analysis, SA2 and CTS3 treatments were identified as the most effective for storage and preservation. Consequently, the expression levels of key disease resistance-, and senescence-related genes were quantified for these two treatments. The results showed that SA2 and CTS3 treatments significantly upregulated the expression of genes involved in phenylpropanoid metabolism (VvPAL), antioxidant defense (VvCAT, VvAPX, VvGR), and direct pathogen response (VvBGLU, VvGST). Concurrently, the treatments downregulated genes associated with enzymatic browning, senescence, and stress response (VvPPO, VvPOD, VvLOX). This transcriptional reprogramming suggests a coordinated shift towards a defense-primed and senescence-delayed state (Fig. 6).
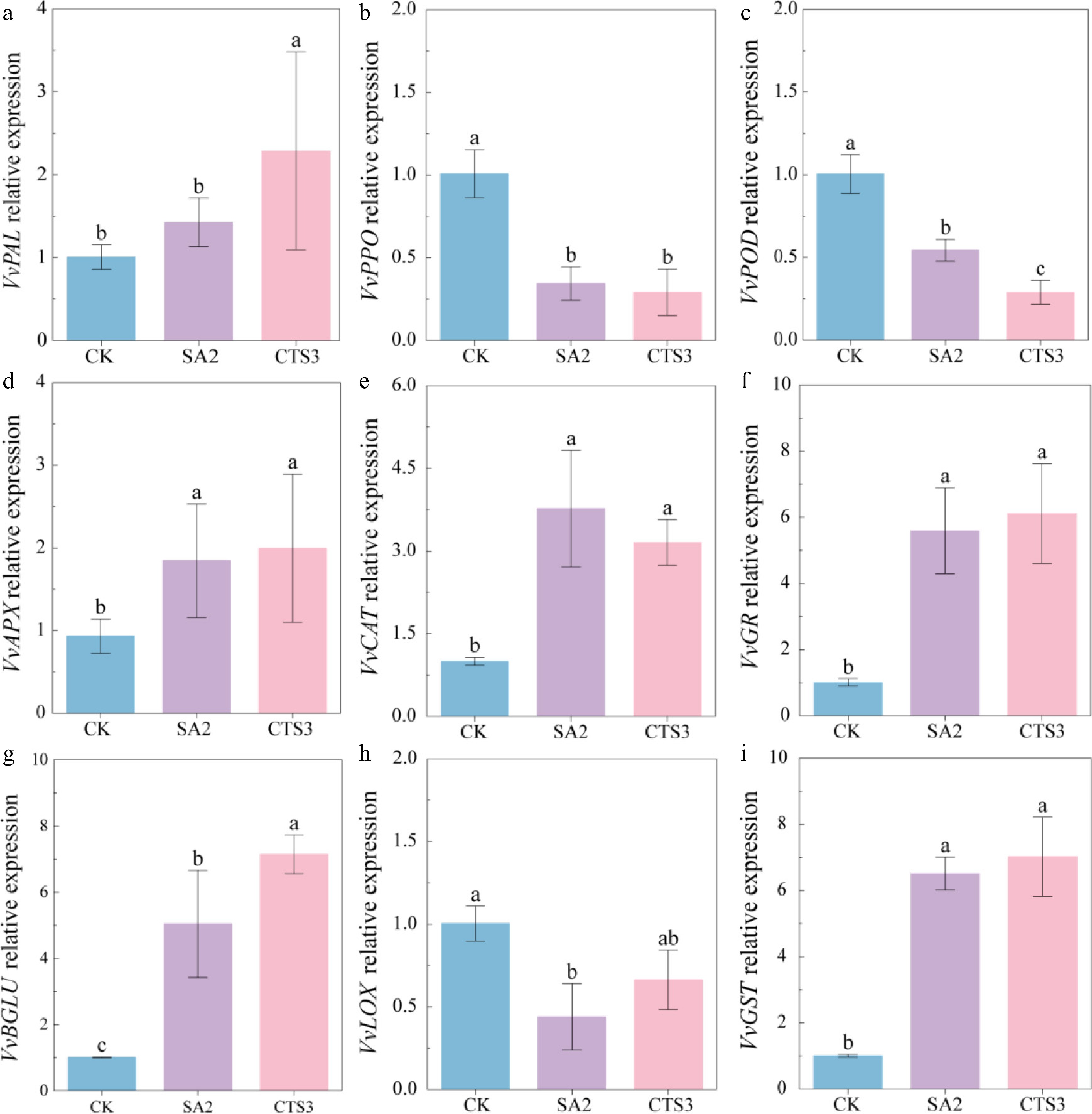
Figure 6.
Effects of SA2 and CTS3 treatments on the relative expression levels of disease-response genes in grapes inoculated with Aspergillus niger. (a) VvPAL; (b) VvPPO; (c) VvPOD; (d)VvAPX; (e) VvCAT; (f) VvGR; (g) VvBGLU; (h)VvLOX; (i) VvGST. Data is presented as mean ± SD, n = 3. Different letters indicate significant (p < 0.05) differences between the control and treated samples.
To quantitatively assess the functional relevance of these gene expression changes to the observed antifungal outcome, Pearson correlation analysis was performed between the relative expression levels of each gene and the lesion area caused by A. niger inoculation. The analysis revealed highly significant positive correlations for most of the upregulated defense and antioxidant genes with reduced lesion area: VvPAL (R = 0.61, p < 0.05), VvAPX (R = 0.83, p < 0.01), VvCAT (R = 0.76, p < 0.01), VvGR (R = 0.86, p < 0.001), VvBGLU (R = 0.83, p < 0.01), and VvGST (R = 0.93, p < 0.001). Conversely, the downregulated genes VvPPO and VvPOD also showed significant positive correlations with lesion area (R = 0.79 for both, p < 0.01), meaning that a greater reduction in their expression was associated with less disease severity. VvLOX showed a weaker but significant positive correlation (R = 0.49, p < 0.01) (Supplementary Fig. S1). These correlation results provide strong quantitative support for the functional role of the observed gene regulation. The strong positive correlations of VvBGLU and VvGST with disease resistance underscore their direct contribution to antifungal defense. Similarly, the correlations of antioxidant genes (VvCAT, VvAPX, VvGR) with reduced lesion area suggest that enhanced antioxidant capacity is integrally linked to limiting pathogen spread. The positive correlations of the downregulated genes (VvPPO, VvPOD, VvLOX) confirm that suppressing senescence and browning pathways is beneficial for overall fruit resistance against fungal decay.
-
The independent antifungal activities of CTS and SA against A. niger underscore their distinct primary modes of action. The hyphal deformities induced by CTS provide direct morphological support for its established mechanism of disrupting fungal cell wall and membrane integrity via cationic interaction[20,21]. This aligns with the broader understanding of chitosan's antimicrobial activity across diverse fungal species, confirming its role as a direct contact agent. In contrast, while SA is widely recognized as a defense signaling molecule[22], our observation of its direct inhibition of conidiation at the applied concentration highlights an often-overlooked contact-level antifungal property in postharvest contexts. The strong correlation between this in vitro activity and the reduced lesion expansion in vivo establishes a clear causal link: direct pathogen suppression is a significant and independent component of each treatment's overall efficacy, contributing substantially to the reduction of macroscopic decay during storage. While the present study focuses on these fundamental inhibitory effects on growth and morphology, the potential influence of SA and CTS on specific fungal virulence factors (e.g., hydrolytic enzyme secretion or secondary metabolism) presents an important avenue for future mechanistic investigation.
Effects on appearance quality under different temperatures
-
The high susceptibility of table grapes to postharvest quality loss is well-established, with thin skin and high moisture content predisposing them to mechanical injury, microbial decay, and pedicel abscission, factors that collectively drive significant economic losses globally[23]. Storage temperature is a critical determinant of this deterioration, directly modulating the rate of water loss, berry softening, and pedicel senescence, which ultimately governs FAR[24,25]. Our observations on 'Shine Muscat' grapes, which showed a progressive increase in DR, WLR, and FAR during storage, align with this general pattern reported across grape varieties[26]. However, the efficacy of preservative treatments in mitigating these losses varied notably between our study and previous reports, pointing to cultivar-specific physiological factors.
In particular, the superior performance of 1.0 mmol/L SA in reducing WLR, contrasts with the 2.0 mmol/L concentration identified as optimal in certain other table grape studies[27]. This discrepancy may be attributed to the characteristically thinner cuticle of 'Shine Muscat', which could enhance SA permeability and lower the effective concentration threshold required to elicit a physiological response, a hypothesis supported by similar findings in 'Cuibao' seedless grapes[28]. Conversely, the strong performance of 2.0% CTS across temperature conditions aligns with its established role as a physical barrier, though its pronounced effect under fluctuating storage regimes suggests that environmental variability may influence coating integrity or fruit responsiveness, as noted in citrus systems[29]
Differential effects of SA and CTS on grape nutritional metabolites under temperature fluctuation
-
Preserving nutritional quality postharvest requires interventions that target specific metabolic vulnerabilities. The attenuation of the common postharvest rise in total soluble solids by both treatments indicates a shared, indirect influence on carbohydrate metabolism[30,31]. A fundamental mechanistic divergence, however, is evident in their effects on key quality metabolites. In stark contrast, the exceptional efficacy of CTS in retaining ascorbic acid underscores its role as a microenvironment modulator[32]. By forming a semi-permeable coating that limits oxygen diffusion, CTS creates a low-oxygen apoplastic environment that directly slows the oxidation of this labile compound, a physical-barrier mechanism less pertinent to SA's action. This distinction is consistent with comparisons showing chitosan's superior AsA retention over certain elicitor-based treatments in berries[33]. The differential impact on TSP further supports this mechanistic model. CTS treatment has been shown to enhance total protein content in postharvest fruit through the elicitation of biochemical defense responses. For example, in tomato fruit, CTS treatment increased total protein levels while decreasing polyphenoloxidase activity, suggesting a role in modulating defense-related proteins[34]. In contrast, SA has been reported to moderately preserve soluble protein content during postharvest storage, with levels in SA-treated groups (28.22%–28.69%) exceeding those of the control (26.47%) in Pleurotus ostreatus. This protein retention is attributed to SA's ability to inhibit ethylene synthesis, delay senescence, and maintain membrane integrity. Nevertheless, compared to more effective treatments such as methyl jasmonate and citric acid, SA's efficacy in protein preservation is intermediate[35], highlighting its role as a biochemical modulator rather than a physical barrier agent.
Temperature-dependent antioxidant regulation by SA and CTS in postharvest grapes
-
The progression of postharvest senescence is fundamentally governed by the collapse of redox homeostasis, in which the overproduction of ROS overwhelms cellular scavenging capacity, leading to oxidative damage[36,37]. Effective preservation strategies must therefore reinforce the antioxidant system, yet the optimal approach can vary depending on the primary mode of action and the storage environment[38]. Our findings elucidate that CTS and SA employ contrasting, temperature-dependent strategies to achieve this reinforcement, offering a clear mechanistic framework for their differential efficacy.
At 4 °C, the superior antioxidant profile observed with 2.0% CTS—characterized by sustained high SOD activity and minimized MDA accumulation—can be interpreted through the lens of its primary physical mode of action. Low temperature intrinsically suppresses respiratory metabolism and ROS production[39]. In this context, the CTS coating provides a complementary physical barrier that further restricts oxygen influx, thereby reducing the substrate for ROS generation at its source[40]; this strategy contrasts with treatments that primarily enhance scavenging. While CTS may also possess radical-scavenging properties, its efficacy at 4 °C appears predominantly driven by this synergistic limitation of oxidative challenge, a mechanism that aligns with reports of chitosan's success in cold-stored commodities[41].
Conversely, under ambient storage (26 °C), where metabolic and oxidative rates are inherently higher, the 1.0 mmol/L SA treatment showed greater efficacy. This aligns with SA's established role as a potent inducer of systemic acquired resistance and antioxidant priming[42,43]. The early and sustained upregulation of key antioxidant enzymes (CAT, SOD) reflects a priming mechanism, whereby SA signaling pre-activates the fruit's enzymatic scavenging machinery to better cope with accelerated stress. This mechanism explains the significant reduction in MDA accumulation compared to the control. Furthermore, the observed modulation of POD activity by SA warrants attention. While POD activity generally rises with stress, its specific suppression by SA may reflect a targeted regulation to prevent its pro-oxidant and browning-related functions, thereby providing a dual benefit of defense and quality retention[44]. This nuanced regulation highlights the sophistication of SA-mediated signaling compared to a purely barrier-based approach.
Synergistic regulation of defense pathways by SA and CTS
-
The ultimate measure of a postharvest treatment's efficacy lies in its capacity to shift the fruit's transcriptional landscape from a senescence-prone to a defense-primed state. Our gene expression analysis reveals a pivotal finding: despite initiating from distinct biochemical and physical entry points, the optimal concentrations of SA (1.0 mmol/L) and CTS (2.0%) orchestrate a convergent transcriptional program. This common defense signature provides a unified molecular logic underlying their independent success in conferring physiological benefits.
The coordinated downregulation of key deterioration genes VvPPO and VvLOX constitute a fundamental mechanism for delaying aging-related quality loss[45]. Polyphenol oxidase (PPO) and lipoxygenase (LOX), central catalysts for enzymatic browning and membrane lipid peroxidation, respectively, are processes directly linked to quality loss[46]. The suppression of these genes by both SA and CTS aligns with findings where other elicitors, such as Burdock Oligosaccharide, similarly extends shelf life by inhibiting these key deterioration pathways[47]. This suggests the existence of conserved regulatory nodes in the senescence network that can be independently targeted by different external stimuli.
Concurrently, the upregulation of defense-related genes (VvBGLU, VvGST) and core antioxidant system genes (VvCAT, VvAPX, VvGR) indicates a concerted activation of both direct and indirect resistance mechanisms[48]. The induction of VvBGLU (β-glucosidase) and VvGST points to enhanced pathogen defense capacity, a response pattern also observed in ethylene- or jasmonate-mediated resistance[49]. The upregulation of the ascorbate-glutathione cycle genes (VvCAT, VvAPX, VvGR) mirrors mechanisms reported in papaya and other fruit treated with resistance inducers, confirming the critical role of redox homeostasis in postharvest persistence[50]. While SA and CTS initiate their actions independently, their convergence on downregulating senescence genes (VvLOX, VvPPO) while upregulating both antioxidant (VvCAT, VvAPX) and defense genes (VvBGLU, VvGST), suggests they are orchestrating a coordinated shift in the fruit’s physiological state[51]. The activation of the antioxidant pathway likely creates a cellular environment less conducive to senescence-related oxidation and pathogen proliferation, indirectly mediating the observed anti-senescence and antifungal effects. Thus, 'coordination' here refers to the functional synergy of these pathways in achieving overall resistance, even if triggered by separate agents.
Both chitosan and salicylic acid offer high practical suitability for commercial adoption. Chitosan holds a 'Generally Recognized as Safe' (GRAS) status from the US FDA specifically for postharvest coating applications[52]. Salicylic acid, being an endogenous plant signal molecule, presents a minimal residue concern as it integrates into natural metabolic pathways[53]. Their safety profiles, combined with the clear, temperature-specific efficacy demonstrated in this study (2.0% CTS at 4 °C; 1.0 mmol/L SA at 26 °C), provide a robust and eco-friendly framework for reducing postharvest losses in the grape industry, serving as viable alternatives to synthetic fungicides (Fig. 7).
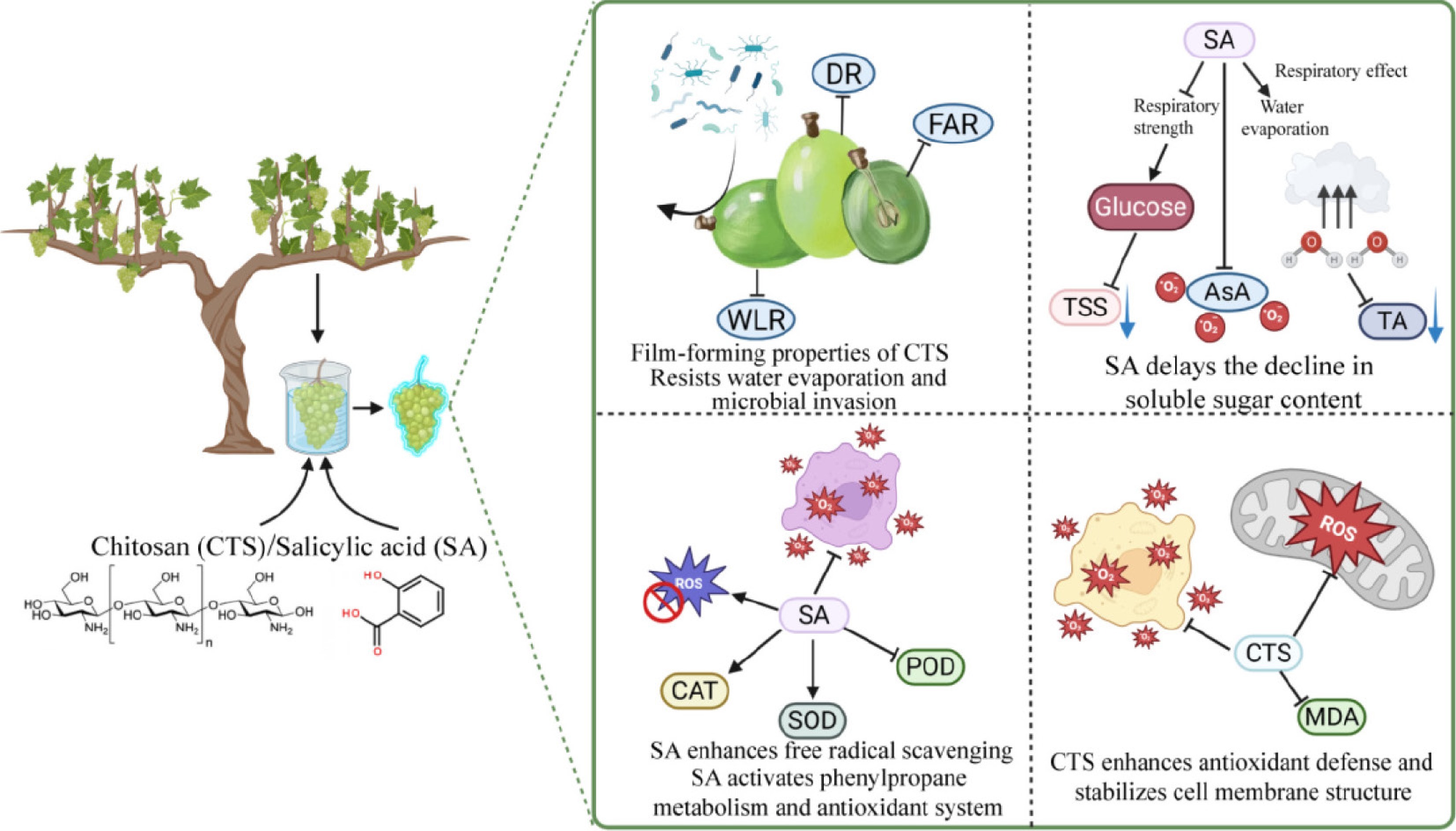
Figure 7.
Effects of postharvest coating with chitosan and salicylic acid on grape storage quality. CTS (chitosan); SA (salicylic acid); DR (decay rate); WLR (weight loss rate); FAR (fruit abscission rate); TSS (total soluble solids); TA (titratable acidity); AsA (ascorbic acid); CAT (catalase); SOD (superoxide dismutase); POD (peroxidase); MDA (malondialdehyde); ROS (reactive oxygen species). The figure was created on BioRender (
www.biorender.com ). -
This study demonstrates that chitosan and salicylic acid are effective standalone treatments for preserving 'Shine Muscat' grapes, with efficacy strictly dependent on storage temperature. At 4 °C, a 2.0% CTS coating forms a protective barrier that minimizes weight loss and decay, while boosting antioxidant defense. At 26 °C, 1.0 mmol/L SA excels by systemically inducing antioxidant enzymes and priming pathogen resistance. Molecular analysis confirmed that both treatments convergently downregulated senescence genes (VvPPO, VvPOD, VvLOX), and upregulated defense and antioxidant genes (VvPAL, VvCAT, VvAPX, VvGR, VvBGLU, VvGST), explaining the physiological benefits. For practical use, both safe, food-compatible compounds offer a clear protocol: apply 2.0% CTS for cold storage, and 1.0 mmol/L SA for ambient conditions. This eco-friendly strategy provides the grape industry with a viable, science-based alternative to synthetic fungicides for reducing postharvest loss.
-
The authors confirm their contributions to the paper as follows: study conception and design, draft manuscript preparation: Geng W; data collection: Gao X; analysis and interpretation of results: Geng W, Yang J, Zhang W, Xu F, Wang L, Ye J. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study is included in this published article and its supplementary information files.
-
This work was supported by grants from the National Natural Science Foundation of China (Grant No. 32202422), and the Shishou City Pioneering County 'Advanced Technology Integration Demonstration Base Construction and Targeted Research Initiatives' tender-based project (Grant No. SS202306).
-
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
-
accompanies this paper online at: https://doi.org/10.48130/frures-0026-0006.
- Supplementary Table S1 Sequences of primers used by RT-qPCR.
- Supplementary Fig. S1 Correlation analysis between lesion area and the relative expression levels of defense-related genes in grapes after inoculation with Aspergillus niger and treatment with SA2 or CTS3.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Geng W, Gao X, Yang J, Zhang W, Xu F, et al. 2026. Chitosan and salicylic acid enhance postharvest resistance in grapes via integrated modulation of antioxidant, antifungal, and anti-senescence pathways. Fruit Research 6: e011 doi: 10.48130/frures-0026-0006 

Chitosan and salicylic acid enhance postharvest resistance in grapes via integrated modulation of antioxidant, antifungal, and anti-senescence pathways
- Received: 04 January 2026
- Revised: 24 February 2026
- Accepted: 27 February 2026
- Published online: 25 March 2026
Abstract: The limited postharvest life of grapes poses a significant challenge to their commercial value. This study employed an integrated approach, combining in vitro and in vivo antifungal assays, storage experiments at two temperatures (4 and 26 °C), and physiological and molecular analyses, to elucidate the effects and underlying mechanisms of chitosan (CTS), and salicylic acid (SA) on preserving the quality of 'Shine Muscat' grapes. The results demonstrated significant antifungal activity of both CTS and SA against Aspergillus niger, with 2% CTS inducing hyphal deformities, and 1.0 mmol/L SA strongly inhibiting conidiation. Under storage conditions, 2.0% CTS at 4 °C most effectively reduced decay, fruit drop, and weight loss, while enhancing antioxidant enzyme activities (SOD, CAT) and minimizing malondialdehyde accumulation. Conversely, at 26 °C, 1.0 mmol/L SA was superior in alleviating quality deterioration by activating the antioxidant system and delaying acid degradation. Quantitative analysis revealed that the optimal treatments (2.0% CTS and 1.0 mmol/L SA) upregulated genes related to phenylpropanoid metabolism (VvPAL), antioxidant defense (VvCAT, VvAPX, VvGR), and defense responses (VvBGLU, VvGST), while downregulating senescence- and browning-related genes (VvPPO, VvPOD, VvLOX). Mechanistically, CTS primarily formed a protective semipermeable coating to reduce water loss and pathogen contact, whereas SA enhanced membrane stability and systemic antioxidant capacity. These findings elucidate the distinct mechanisms of CTS and SA and provide a theoretical basis for developing targeted, eco-efficient preservation strategies based on storage temperature.
-
Key words:
- Shine muscat grapes /
- Food preservation /
- Chitosan /
- Salicylic acid /
- Aspergillus niger