









-
Plants initiate a series of complex molecular and physiological responses when facing abiotic stress. On a global scale, the most significant natural disasters include drought, low temperature, and salt stress[1]. Drought stress leads to water imbalance and oxidative stress in plant cells, thereby inhibiting plant growth and development. Low-temperature stress causes cell freezing, dehydration, and oxidative stress[2], while salt stress disrupts ionic balance and causes osmotic dysregulation[3]. Plants respond to various abiotic stresses through different pathways and mechanisms, and these responses involve the regulation of many different types of genes. Transcription factors such as Dehydration Responsive Element Binding Protein (DREB)[4], NAC (NAM, ATAF1/2, CUC2)[5], Basic Leucine Zipper (bZIP)[6], and MYB[7] play crucial roles. Stress-responsive proteins such as Late Embryogenesis Abundant Protein (LEA)[8] and Heat Shock Protein (HSP)[9] are also involved.
The first NAC gene, NAM, was identified in petunia[10], followed by the discovery of Arabidopsis thaliana Transcription Activation Factor 1/2 (ATAF1/2) and Cup-Shaped Cotyledon 2 (CUC2)[11]. Based on predicted and known sequence similarity of NAC domains, Ooka et al. classified these members into two major groups and 18 subgroups[12]. Subsequent genome-wide analyses revealed varying numbers of NAC family members across plant species. For instance, rice and Arabidopsis contain at least 151 and 117 NAC genes, respectively[13]; maize has at least 157, wheat at least 210, and barley at least 48[14]. In dicot species, cotton has at least 220 members, tomato has at least 101, and soybean has been reported to contain 152 members[15,16]. In woody plants, poplar contains at least 163 NAC members[17], and 45 NAC genes were identified in the EST database of citrus[18], while apple has at least 180 members[19]. To date, the mango NAC gene family has not been systematically characterized. Existing reports are limited to preliminary functional studies of only a few genes. For instance, MiNAC1 and MiNAC7 have been implicated in abiotic stress responses[20,21], MiNAC6 is involved in carotenoid degradation during postharvest metabolism[22], and MiNAC25[23] has been identified as co-expressed with the Stay-Green Regulator 1 (MiSGR1) to regulate fruit coloration. Clearly, these findings have not been validated in mango, and the function of MiNAC2D is also unclear.
The structural diversity of NAC proteins endows them with various functions, enabling NAC transcription factors to play crucial roles in almost every stage of plant growth and development, as well as under various stress conditions[24]. In Arabidopsis, the NAC transcription factor 2 (AtNAC2) gene has extremely high expression levels in the roots and flowers, and is highly induced by high salt, abscisic acid (ABA), 1-aminocyclopropane-1-carboxylic acid (ACC), and α-naphthaleneacetic acid (NAA). Overexpression of AtNAC2 results in an increased number of lateral roots in transgenic plants[25]. Overexpression of the wheat TaNAC2 gene in Arabidopsis enhances tolerance to drought, salt, and freezing stresses[26]. Transgenic tomato plants overexpressing the SlNAC2 gene exhibit increased tolerance to salt and drought (PEG) stresses[27].
Mango (Mangifera indica) is a crucial tropical and subtropical fruit. Environmental factors such as drought, low temperature, and salt stress seriously affect mango physiological and biochemical processes and severely limit its growth and development and fruit quality. In this study, an NAC transcription factor, MiNAC2D, was selected from the drought and salt stress transcriptome data based on a differential expression fold change greater than 2 (|log2 FC| > 1) (unpublished). Therefore, this study aimed to investigate the function of MiNAC2D. The results indicated that the MiNAC2D gene plays a facilitating role in the response to abiotic stresses.
-
The mango variety 'SiJiMi' was grown in the experimental orchard of Guangxi University. Arabidopsis thaliana ecotype Columbia-0 (Col-0) and tomato (Solanum lycopersicum) variety Micro-Tom were used for genetic transformation.
Sequences and phylogenetic analyses
-
Physicochemical properties of MiNAC2D were analyzed using BioXM 2.7 software, and specific primers were designed with Primer3Plus. Gene structure (exon-intron organization) was determined using the Gene Structure Display Server 2.0 (GSDS 2.0). Chromosomal localization of MiNAC2D in 'SiJiMi' mango was visualized with TBtools, and a chromosome distribution density heatmap was generated. Secondary structure and three-dimensional structural models of the MiNAC2D protein were predicted using SOPMA and SWISS-MODEL, respectively. Multiple sequence alignment of MiNAC2D homologs based on their deduced amino acid sequences was performed using DNAMAN. Conserved domain analysis was conducted using Pfam and SMART. For cis-element analysis, the 2,000 bp sequence upstream of the start codon (ATG) was submitted to PLACE and PlantCARE, and the results were visualized with TBtools. Homologous MiNAC2D sequences were retrieved from NCBI using BLAST, and a phylogenetic tree was constructed using MEGA 11, with subsequent refinement using EvolView. Statistical analyses and graphical presentation were performed using GraphPad Prism 7, and image assembly was conducted using PowerPoint.
Sample extraction and validation of MiNAC2D gene expression
-
DNA of mango and Arabidopsis was extracted using the cetyltrimethylammonium bromide (CTAB) method. Total RNA was extracted from mango using the RNAprep Pure Plant Kit (Tiangen, Beijing, China) and from Arabidopsis using the HiPure Plant RNA Mini Kit (Magen, Guangzhou, China). RNA integrity and concentration were assessed by 1.5% agarose gel electrophoresis and UV spectrophotometry, respectively. cDNA was synthesized by reverse transcription using an oligo(dT)18 primer and M-MLV Reverse Transcriptase (RNase H−).
To investigate the expression pattern of MiNAC2D in different tissues and under abiotic stress conditions in 'SiJiMi' mango, specific primers were designed, and quantitative real-time PCR (RT-qPCR) was conducted using the ChamQ SYBR qPCR Master Mixture (Vazyme Biotech). MiActin1 was used as the reference gene, and fold changes in gene expression were calculated using the 2−ΔΔCᴛ method[28].
Subcellular localization analysis
-
The 35S::MiNAC2D-GFP fusion expression vector was constructed by removing the stop codon from the full-length coding sequence (CDS) of MiNAC2D, with the empty 35S::GFP vector serving as a negative control. The fusion construct was introduced into the Agrobacterium tumefaciens strain EHA105. Onion epidermal cells were then transformed via Agrobacterium-mediated transformation for the transient expression of 35S::MiNAC2D-GFP and 35S::GFP. After 16 h of incubation and DAPI staining, fluorescence signals were observed using a confocal laser scanning microscope (Leica TCS-SP8 MP, Germany).
Analysis of MiNAC2D protein self-activation
-
A bait vector pGBKT7 containing the full-length coding region of MiNAC2D was constructed and transformed into yeast cells. Single colonies of yeast transformed with pGBKT7-MiNAC2D or the empty pGBKT7 vector were selected and cultured in SD/-Trp liquid medium. The cultures were then diluted to 10−1, 10−2, 10−3, and incubated for 12 h. Aliquots of each dilution were plated onto SD/-Trp, SD/-Trp/X-α-Gal, and SD/-Trp/X-α-Gal/AbA solid media. Plates were incubated upside down at 30 °C for 2–3 d, and colony growth was observed. If colonies transformed with pGBKT7-MiNAC2D grew on SD/-Trp/X-α-Gal and SD/–Trp/X-α-Gal/AbA media and turned blue, this indicated that MiNAC2D activated the reporter gene, suggesting self-activation activity. Conversely, if colonies grew only on SD/-Trp and SD/-Trp/X-α-Gal media but failed to grow on SD/-Trp/X-α-Gal/AbA medium, no self-activation was detected.
Transformation of Arabidopsis with MiNAC2D and phenotypic analyses
-
The MiNAC2D gene fragment was amplified using primers containing XbaI and SmaI restriction sites and cloned into the pBI121-GUS empty vector. The sequence-verified plasmid was transformed into Agrobacterium tumefaciens strain EHA105. Arabidopsis plants were transformed using the floral dip method. T0 seeds were harvested and sown; T1 plants were screened by PCR amplification and electrophoresis, followed by sequencing to confirm the presence of the transgene. Positive T1 plants were propagated to the T3 generation to obtain homozygous transgenic lines, with WT Arabidopsis as controls.
Functions of MiNAC2D-OE Arabidopsis in terms of drought, salt, and low temperature tolerance
-
To comprehensively evaluate the physiological and molecular responses of transgenic Arabidopsis under abiotic stress conditions, phenotypic, physiological, and gene expression analyses were conducted.
For seed germination analysis, WT and MiNAC2D-overexpressing (OE) lines were grown on 1/2 MS medium supplemented with 300 mM mannitol, 100 mM NaCl, or 10 μM ABA, or subjected to 4 °C. Germination rates were recorded every 12 h over a 3-d period and documented photographically.
Root phenotype under abiotic stress was assessed by growing WT and MiNAC2D-OE seedlings on 1/2 MS medium containing various concentrations of mannitol (0, 200, 300, 400 mM), NaCl (0, 100, 150, 200 mM), or ABA (10, 20 μM). Plates were placed vertically at 22 or 4 °C for 7 d, after which primary root length and the number of lateral roots (> 5 mm) were measured and photographed. For seedling survival assays, seven-day-old seedlings were transplanted into soil pots and acclimated under normal watering for one week. Subsequently, three-week-old WT and MiNAC2D-OE plants were sprayed with 300 mM NaCl or 300 mM mannitol solutions to simulate salt and drought stress, respectively, with water as the control. Cold stress was imposed by transferring the seedlings to 4 °C for 10 d. Survival rates were recorded after treatment.
To evaluate oxidative stress and cell death under abiotic conditions, histochemical staining was performed. Two-week-old soil-grown rosette-stage plants were treated with water, 300 mM mannitol, or 200 mM NaCl for 6 h, after which leaves were stained with 0.1 mg/mL nitro blue tetrazolium chloride (NBT) for 3 h, 1 mg/mL 3,3'-Diaminobenzidine (DAB) for 24 h, and 2.5% Evans Blue for 6 h. Samples were decolorized in 95% ethanol and imaged using a super-depth field microscope. For physiological measurements, aerial parts of WT and MiNAC2D-OE plants were collected after treatment with 300 mM mannitol, 200 mM NaCl, or water. Malondialdehyde (MDA), proline (Pro), hydrogen peroxide (H2O2) contents, and superoxide dismutase (SOD) activity were quantified using commercial kits (Solarbio, Beijing, China).
To elucidate molecular mechanisms underlying abiotic stress responses, WT and transgenic plants were subjected to 300 mM NaCl for 12 h (salt stress), 20% PEG 6,000 for 24 h (drought stress), or 4 °C for 6 h (cold stress), with water treatment as a control. Total RNA was extracted from whole plants, reverse transcribed, and analyzed by RT-qPCR. AtActin2 was used as the internal control. Marker genes for drought stress included AtKIN1, AtRD29A, and AtCOR15A; for salt stress: AtSOS1, AtNHX1, and AtDREB2A; and for cold stress: AtRD29A and AtRD29B. The primer sequences are listed in Supplementary Table S1. All experiments were performed with three biological replicates.
Transformation of tomato with MiNAC2D and phenotypic analyses
-
Transgenic tomato plants (T1 generation) were generated by Agrobacterium-mediated transformation using WT tomato as the recipient. The lines were propagated to the T3 generation, and homozygous lines were identified via PCR screening of genomic DNA extracted from leaf tissue. Based on expression analysis, the three transgenic lines with the highest target gene expression levels were selected for further analysis, using WT tomato plants as controls. All plants were grown in soil substrate under controlled conditions in a growth chamber (16 h light/8 h dark, 25 °C), and phenotypic traits were monitored and recorded.
Functions of MiNAC2D-OE tomato in terms of drought, salt and low temperature tolerance
-
To evaluate the abiotic stress responses of transgenic tomato plants, seed germination assays, root growth analyses, seedling survival tests, and endogenous gene expression profiling were performed. For the germination assay, T3 generation transgenic and WT tomato seeds were sown on 1/2 MS medium supplemented with various stress agents, including 300 mM mannitol, 200 mM NaCl, or 10 μM ABA, or subjected to 4 °C. Seeds were vernalized at low temperature for 2 d, then transferred to a 22 °C growth chamber under a 16 h light/8 h dark photoperiod. Germination was assessed every 12 h based on the emergence of a radicle ≥ 1 mm, and images were captured on the tenth day after sowing.
To assess the impact of abiotic stress on root development, seeds from transgenic and WT lines were sown on square 1/2 MS plates containing different concentrations of mannitol (0, 200, 300, 400 mM), NaCl (0, 100, 150, 200 mM), or ABA (10, 20 μM). Cold stress was simulated using plates without chemical additives maintained at 4 °C. After vernalization, all plates were placed vertically in growth chambers at 22 or 4 °C for 7 d. Primary root length and the number of lateral roots were measured. Each treatment was performed with three biological replicates.
For seedling survival assays, seven-day-old seedlings germinated on 1/2 MS medium were transplanted into 16 × 16 cm square pots containing soil. After normal watering and growth to the five to six true-leaf stage, plants were subjected to abiotic stress treatments: 300 mM NaCl foliar spray for salt stress; natural drought for 7 d followed by rewatering for 5 d for drought stress; and cold stress by incubation at 4 °C for 3 d. Seedling survival rates were recorded after treatments, and each condition was repeated in three independent biological replicates.
To investigate molecular responses to abiotic stresses, RT-qPCR was conducted to examine the expression of endogenous stress-responsive genes in transgenic and WT tomatoes. Total RNA was extracted from the aboveground parts of homozygous seedlings after 24 h of stress treatment and reverse-transcribed into cDNA. The tomato SlActin gene was used as an internal reference. Salt-responsive genes included Solanum lycopersicum Chitinase 9 (SlCHI9), Aspartate Aminotransferase-Related Protein 1 (SlAPA1), and PIN-Formed 2 (SlPIN2); drought-responsive genes included Protein Phosphatase 2C 1 (SlPP2C1), Abscisic Acid-Responsive Element Binding Protein 1 (SlAREB1), and Dehydration-Responsive Element Binding Protein A4 (SlDREBA4); cold-responsive genes included WHIRLY 1 (SlWHY1), C-Repeat Binding Factor 2 (SlCBF2), and C-Repeat Binding Factor 3 (SlCBF3). The primer sequences are listed in Supplementary Table S1. All analyses were conducted using three biological replicates per treatment.
Transformation of mango roots with MiNAC2D and phenotypic analyses
-
To induce hairy root formation in mango, Agrobacterium rhizogenes was used to infect the hypocotyl region of mango seedlings. A 1 mL aliquot of the confirmed positive bacterial culture was added to 100 mL of TY liquid medium supplemented with 10 mM CaCl2, 50 μg/mL kanamycin, and 50 μg/mL streptomycin under sterile conditions, and incubated until the optical density at 600 nm (OD600) reached 1.0. The bacterial culture was then transferred to 50 mL centrifuge tubes and centrifuged at 6,000 rpm for 10 min at room temperature. The resulting pellet was resuspended in an infection buffer composed of 0.05 M MES, 2 mM Na3PO4, 0.1 mM acetosyringone, and 0.5% (w/v) glucose, and adjusted to an OD600 of 0.8. Healthy mango seedlings were selected, and the hypocotyl region (approximately 1 cm below the cotyledons, near the root zone) was ring-cut to a length of 2 cm using a sterile scalpel. The cambium was gently scraped off, and a vertical incision was made above the girdled region to create a wound. The wound was covered with sterile cotton soaked in the prepared bacterial suspension for 30 min. Subsequently, moist cotton was wrapped around the wound to maintain humidity for 2 d. The treated area was then enclosed in a layering box filled with a moist substrate and vermiculite mixture to support root induction. Water and nutrient solution were applied as needed. After 60 to 100 d, transgenic hairy roots emerged above the wounded site. Genomic DNA was extracted from the regenerated roots for molecular analysis.
DNA was extracted from the hairy roots of putative transgenic mango plants and used as the template for PCR amplification. A forward primer targeting the common upstream region of the vector (35SFu) and a gene-specific reverse primer (MiNAC2Dd) were designed. PCR products showing the expected amplicon size were subjected to cloning and sequencing. Plants yielding sequences that included both vector-specific and target gene regions were identified as transgenic positives and further cultivated. For confirmation, PCR products with the same expected size as that of the recombinant plasmid were sequenced and aligned with the MiNAC2D gene. Positive sequences containing vector elements, restriction sites, and the MiNAC2D coding sequence confirmed the successful integration of the transgene. The total RNA was subsequently extracted from the hairy roots of confirmed transgenic seedlings, reverse transcribed into cDNA, and used for RT-qPCR to assess the expression level of MiNAC2D. Specific primers were designed for MiNAC2D, and expression levels were normalized using the internal reference gene MiActin1. The primer sequences are listed in Supplementary Table S1.
Functions of MiNAC2D mango roots in terms of drought and salt tolerance
-
Transgenic mango seedlings (OE1-OE3) were subjected to abiotic stress treatments by soaking in a 300 mM NaCl solution for 48 h, while transgenic mango seedlings (OE4-OE6) were soaked in 20% PEG 6000 solution for 48 h, with sterile water as the control. Due to the limited availability of transgenic mango plants and the inability to clonally propagate the same line, the control for each genotype (WT and transgenic) was the same individual plants sampled immediately before stress treatment (0 h). After treatment, plant roots were collected for transcriptome sequencing and RT-qPCR analysis. For transcriptome sequencing, the control group (WT and OE1-OE6) was submitted, along with OE1-OE3 under NaCl stress and OE4-OE6 under PEG stress. The primer sequences are listed in Supplementary Table S1. The WT and MiNAC2D transgenic mango root was submitted to Gidiou Company for transcriptome sequencing, with three biological replicates. Sample RNA was extracted and then its purity and integrity were assessed using an Agilent 2100 Bioanalyzer. cDNA libraries were constructed and sequenced on the Illumina HiSeq platform. Raw image data were converted into sequences for analysis via CASAVA base calling. Raw reads were quality-filtered using the fastp software. Gene expression levels were quantified as FPKMs using StringTie (v2.0.4). Differential gene expression analysis was performed using DESeq2 (v1.26.0). Gene functional annotation was carried out using eight major databases: COG, GO, KEGG, KOG, NR, PFAM, SWISS, and EGGNOG; HOMER (v4.11;
https://homer.ucsd.edu/homer/ ) and Mfuzz (v2.46.0;https://bioconductor.org/packages//2.7/bioc/html/Mfuzz.html ) were used for transcription factor analysis and time-series clustering analysis, respectively. A heatmap was subsequently generated using TBtools for visualization.Data analysis
-
GraphPad Prism 9 was used to create the graphs, and SPSS 25.0 was used to organize and statistically analyze the data, and Tukey's t-test was used to test for significance at the p < 0.05 (* or lower case letters in the graphs) and p < 0.01 (**) levels.
-
Bioinformatics analysis revealed that the MiNAC2D gene comprises three exons and two introns, and its structure was visualized using IBS 1.0 software (Fig. 1a). The NAM conserved domain of MiNAC2D was localized to amino acids 11-132 at the N-terminus (Fig. 1b). The MiNAC2D gene was mapped to chromosome 17 of 'SiJiMi' (Fig. 1c). The open reading frame is 885 bp in length, encoding 295 amino acids, with a theoretical isoelectric point of 6.63 and a molecular weight of 34.13 kDa. The three-dimensional conformation of the MiNAC2D protein revealed that irregular coils accounted for 80.27% of the structure, followed by extended strands (11.22%), with α-helices representing the smallest proportion (8.5%) (Fig. 1d). A large number of stress-responsive and multiple hormone-responsive elements were identified in the promoter sequence of MiNAC2D (Fig. 1e). Phylogenetic analysis showed that MiNAC2D clustered most closely with Pistacia vera PvNAC2 (Fig. 1f).
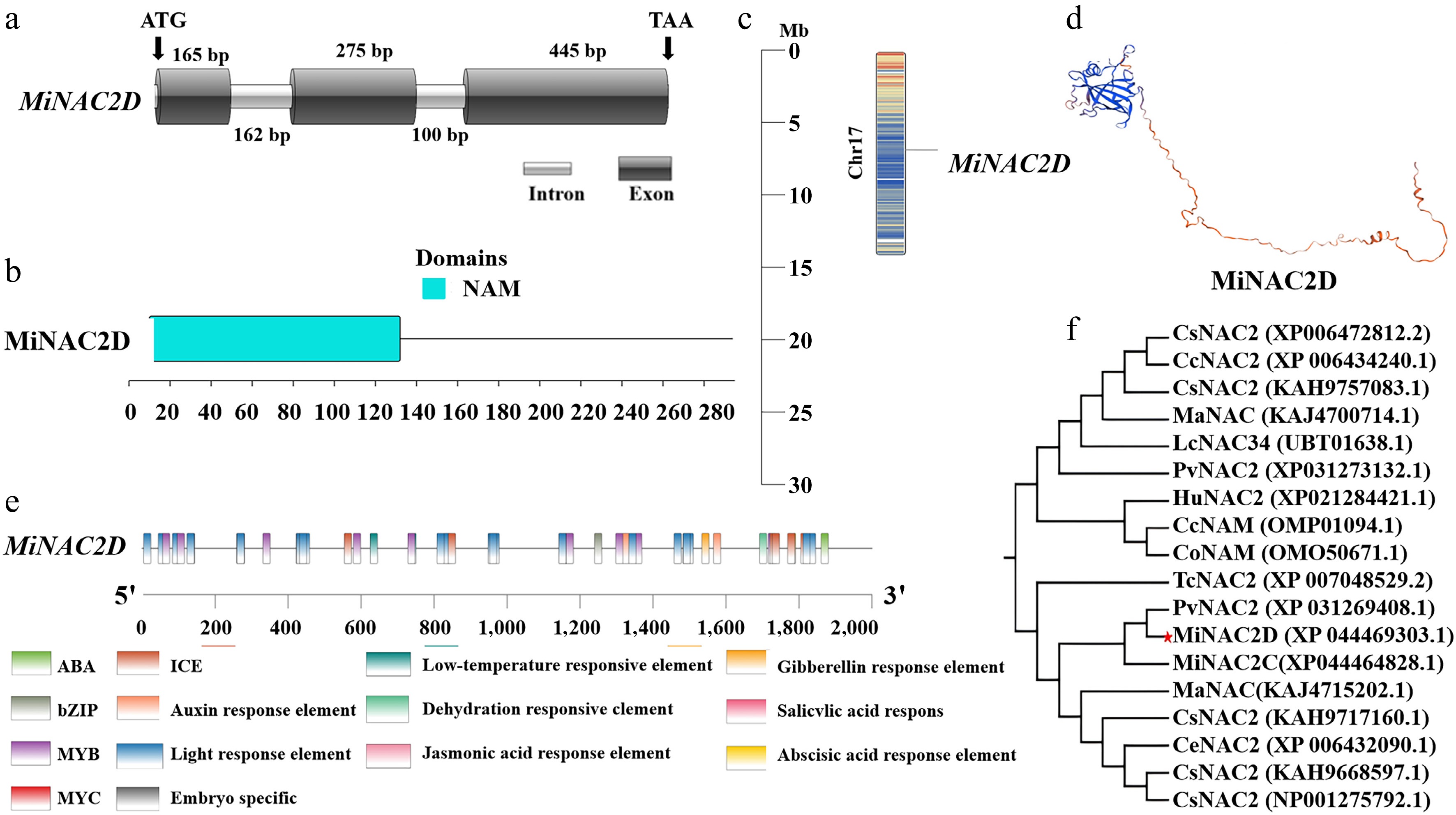
Figure 1.
Identification of MiNAC2D gene. (a) Gene structure of MiNAC2D. (b) Conserved structural domains of MiNAC2D. (c) Chromosomal localization of MiNAC2D. (d) Structure of MiNAC2D protein. (e) Cis-element analysis of MiNAC2D. (f) Phylogenetic tree of NAC proteins. Pv, Pistacia vera; Cs, Cucumis sativus; Cc, Cymbopogon citratus; Ma, Melia azedarach; Lc, Litchi chinensis; Cu, Citrus unshiu; Tc, Theobroma cacao; Hu, Hylocereus undatus; Tc, Theobroma cacao.
Subcellular localization and transcriptional activation activity
-
To examine subcellular localization, the 35S::GFP construct driven by the 35S promoter was used as a control. In unloaded 35S::GFP cells, green fluorescence was widely distributed across the cell membrane, cytoplasm, and nucleus. After DAPI staining, blue fluorescence was observed in the nucleus, indicating that GFP alone exhibited no specific localization. In contrast, in cells expressing the 35S::MiNAC2D-GFP fusion protein, green fluorescence was exclusively localized to the nucleus and completely overlapped with the blue fluorescence from DAPI staining, suggesting that the MiNAC2D protein is localized in the nucleus (Fig. 2a).
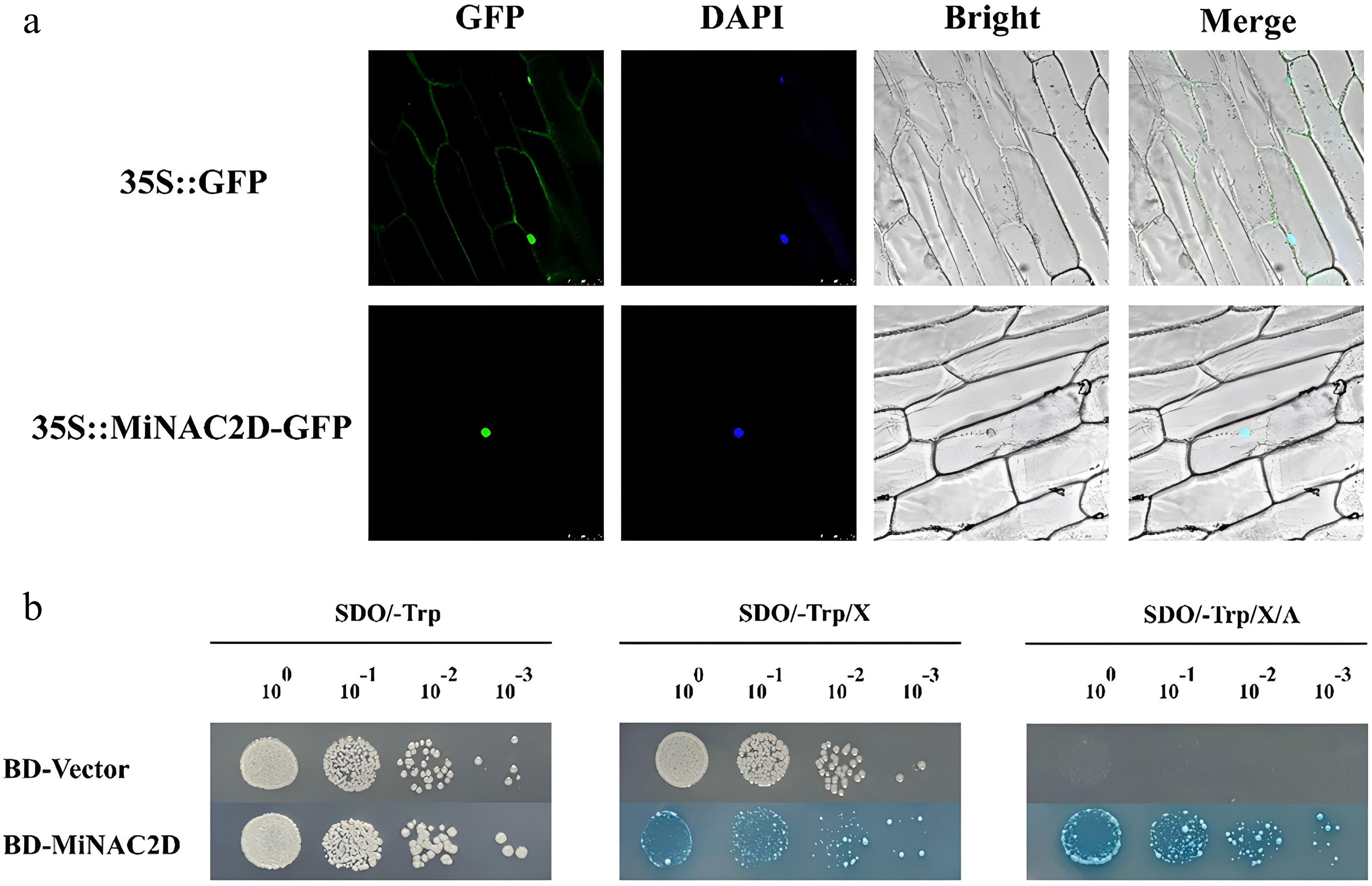
Figure 2.
Subcellular localization and transcriptional activation activity analysis. (a) Subcellular localization of MiNAC2D. Bars = 100 μm. (b) Transcriptional activation activity analysis of MiNAC2D.
To assess transcriptional activation activity, the pGBKT7-MiNAC2D bait vector and the pGBKT7 empty vector were constructed and transformed into yeast cells. The transformed yeast cultures were spotted onto SD/-Trp, SD/-Trp/X, and SD/-Trp/X/A plates at dilutions of 10−1, 10−2, and 10−3, respectively. The results showed that yeast harboring the pGBKT7 empty vector grew as full, round yellow colonies on SD/-Trp and SD/-Trp/X plates but failed to grow on SD/-Trp/X/A plates. Yeast containing the pGBKT7-MiNAC2D bait vector exhibited growth on SD/-Trp medium comparable to that of the pGBKT7 empty vector, indicating that the bait plasmid did not confer toxicity to Y2HGold yeast cells. However, colonies grown on SD/-Trp/X and SD/-Trp/X/A plates turned blue, suggesting that the MiNAC2D protein possesses self-activating activity (Fig. 2b).
Expression analysis of MiNAC2D in different tissues and under stress in mango
-
To investigate the expression pattern of the MiNAC2D gene across different tissues, RT-qPCR analysis was performed. The results showed that MiNAC2D expression was significantly higher in leaves than in other tissues, followed by primary roots, seeds, and fibrous roots, with varying expression levels also detected in stems, flowers, and fruits (Fig. 3a).
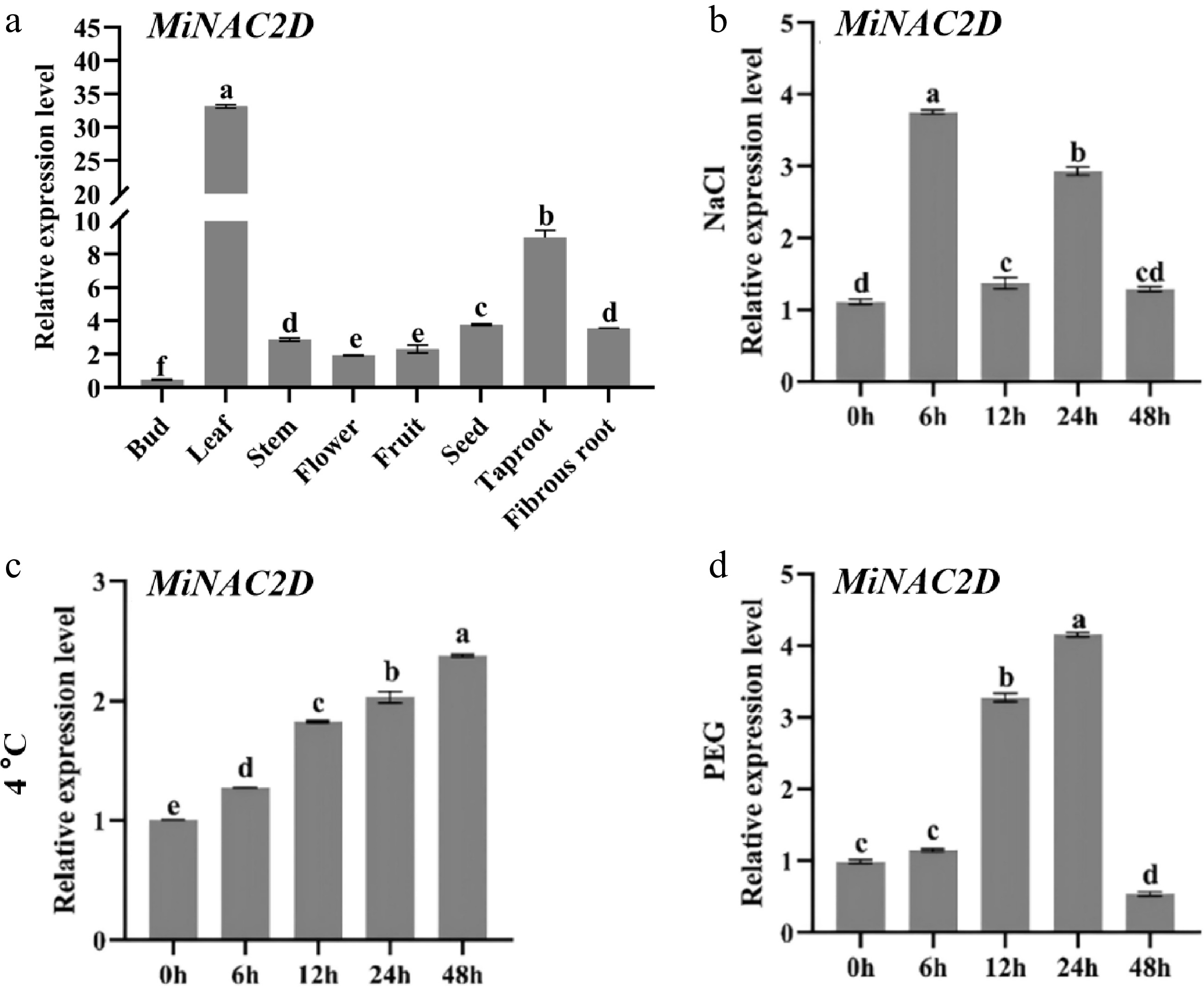
Figure 3.
Expression analysis of the MiNAC2D gene. (a) Tissue expression pattern analysis of the MiNAC2D gene. (b) Expression pattern analysis of the MiNAC2D gene under salt stress. (c) Expression pattern analysis of the MiNAC2D gene under low temperature stress. (d) Expression pattern analysis of the MiNAC2D gene under drought stress. Significant differences are indicated when p < 0.05, with a, b, c, d, e, and f representing significant differences.
The expression pattern of MiNAC2D under different stress conditions was further examined. MiNAC2D responded to drought, salt, and low-temperature stresses to varying degrees. Under salt stress, MiNAC2D expression increased rapidly, reaching the highest level at 6 h. Expression decreased significantly at 12 h but increased again at 24 h, and although it declined at 48 h, it remained higher than that of the control throughout the 6-48 h treatment period (Fig. 3b). Under low-temperature treatment at 4 °C, expression gradually increased from 0 to 48 h, reaching a maximum at 48 h that was approximately three times that of the control group (Fig. 3c). Under 20% PEG 6000 treatment, MiNAC2D expression gradually increased at 6 and 12 h, peaked at 24 h at approximately four times that of the control, and then decreased markedly at 48 h (Fig. 3d).
Germination rate and root parameters of WT and MiNAC2D-OE Arabidopsis under abiotic stresses
-
WT and MiNAC2D-OE lines were incubated for 3 d under treatments of 300 mM mannitol, 100 mM NaCl, 4 °C low temperature, and 10 μM ABA. The results showed that under 100 mM NaCl, the germination rate of the MiNAC2D-OE lines was significantly higher than that of WT, and similar results were observed under 4 °C and 300 mM mannitol treatments. However, under 10 μM ABA treatment, the germination rate of the MiNAC2D-OE lines was significantly lower than that of WT (Fig. 4a; Supplementary Fig. S1). These results suggest that the MiNAC2D gene promotes seed germination under drought, salt, and low-temperature stress conditions, but inhibits germination in response to ABA treatment.
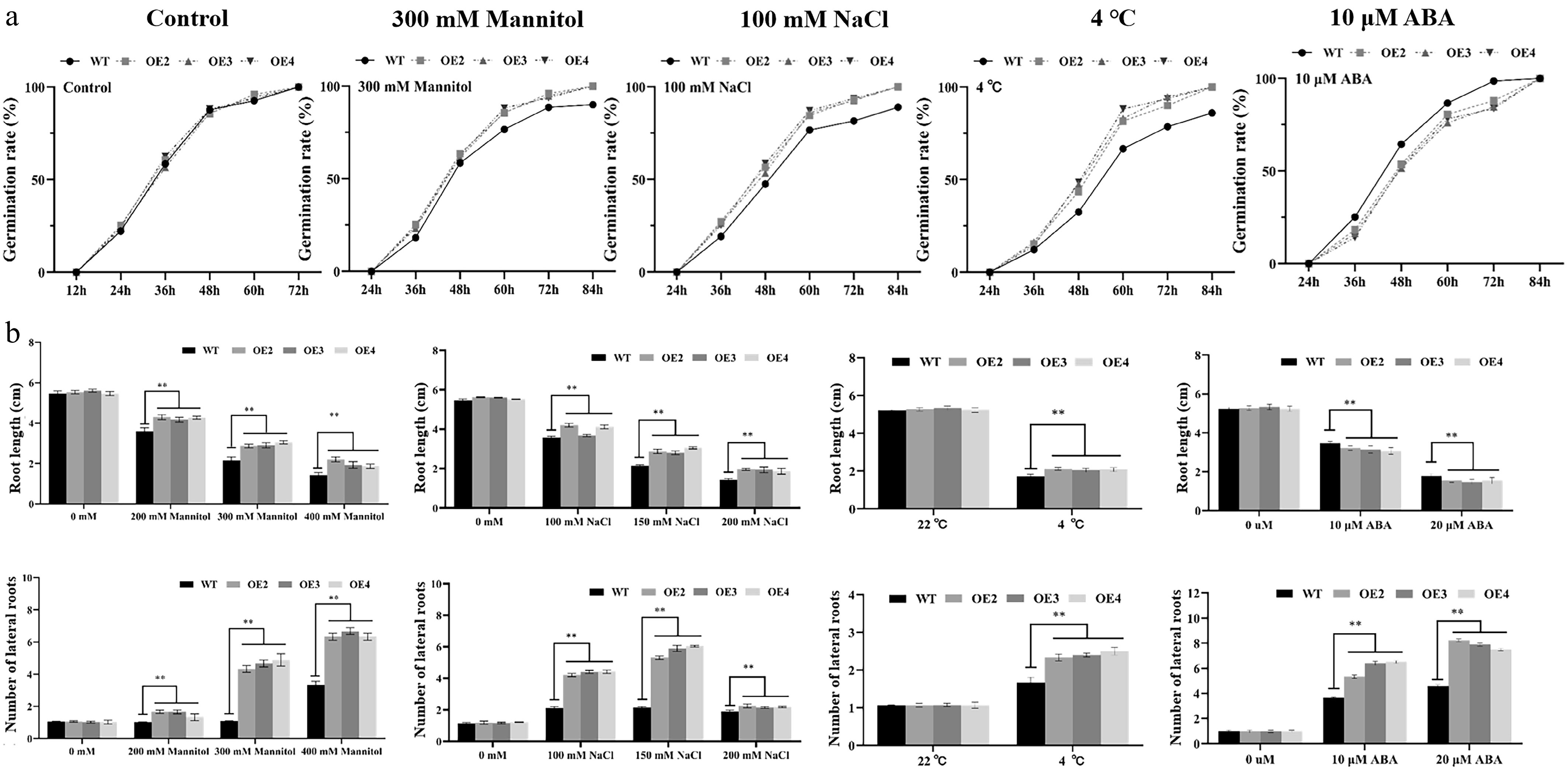
Figure 4.
Performance of WT and MiNAC2D-OE Arabidopsis under drought, salt, 4 °C, and ABA treatment. (a) Germination rate analysis of WT and MiNAC2D-OE. (b) Analysis of root length and lateral root numbers in MiNAC2D-OE and WT after treatment with different sodium chloride concentrations, ABA, and temperature gradients. Significant differences were defined as p < 0.01 (**) according to Tukey's t-tests.
To assess root growth under drought stress, WT and MiNAC2D-OE lines were treated with 0, 200, 300, and 400 mM mannitol, and primary root length and lateral root number were measured. Under control conditions (0 mM mannitol), no significant differences were observed between WT and MiNAC2D-OE lines in either parameter. With increasing mannitol concentrations, growth and development were inhibited in both genotypes, as evidenced by leaf yellowing, narrowed petioles, suppression of primary root elongation, and an increase in lateral root number. However, the MiNAC2D-OE lines exhibited less pronounced growth inhibition. Under mannitol treatment, the MiNAC2D-OE lines had significantly longer primary roots and more lateral roots than WT, indicating that MiNAC2D may enhance drought stress tolerance in Arabidopsis.
Under salt stress conditions, similar trends were observed. In the absence of NaCl, no significant differences were detected between WT and MiNAC2D-OE lines in primary root length or lateral root number. With increasing NaCl concentrations, both genotypes showed growth inhibition, characterized by leaf yellowing and whitening, reduced primary root length, and increased lateral root number. Nevertheless, the MiNAC2D-OE lines displayed milder growth suppression, with significantly longer primary roots and more lateral roots than WT under salt treatment, suggesting that MiNAC2D significantly improves salt stress tolerance in Arabidopsis.
For low-temperature stress, WT and MiNAC2D-OE lines were grown at 22 °C (control) and 4 °C. At 22 °C, no significant differences were observed between the two genotypes. At 4 °C, both exhibited small and thickened leaves, reddening of leaf petioles, and overall reduced vigor. However, under low-temperature treatment, the MiNAC2D-OE lines had significantly longer primary roots and more lateral roots than WT, indicating that MiNAC2D enhances tolerance to cold stress in transgenic Arabidopsis.
Under ABA treatment, both MiNAC2D-OE lines and WT showed growth alterations, but the MiNAC2D-OE lines exhibited smaller and more yellowed leaves and weaker growth vigor compared with WT. Statistical analysis revealed that the MiNAC2D-OE lines had significantly shorter primary roots and significantly more lateral roots than WT, suggesting that MiNAC2D increases sensitivity to ABA treatment in Arabidopsis (Fig. 4b; Supplementary Fig. S2).
MiNAC2D enhanced abiotic stresses tolerance in Arabidopsis
-
Plant survival rates were measured in 14-day-old WT and MiNAC2D-OE Arabidopsis plants after 3 d of spray treatment with 300 mM NaCl, 10 d of low-temperature treatment at 4 °C, and 3 d of rehydration following 12 d of drought. Under normal conditions, MiNAC2D-OE plants remained green and healthy. After salt treatment and after drought followed by rehydration, most MiNAC2D-OE plants remained green, while only a few lost their green color and dried up; in contrast, most WT plants wilted and died. Under low-temperature treatment at 4 °C, the survival rate of the MiNAC2D-OE lines was higher than that of WT (Fig. 5a, b).
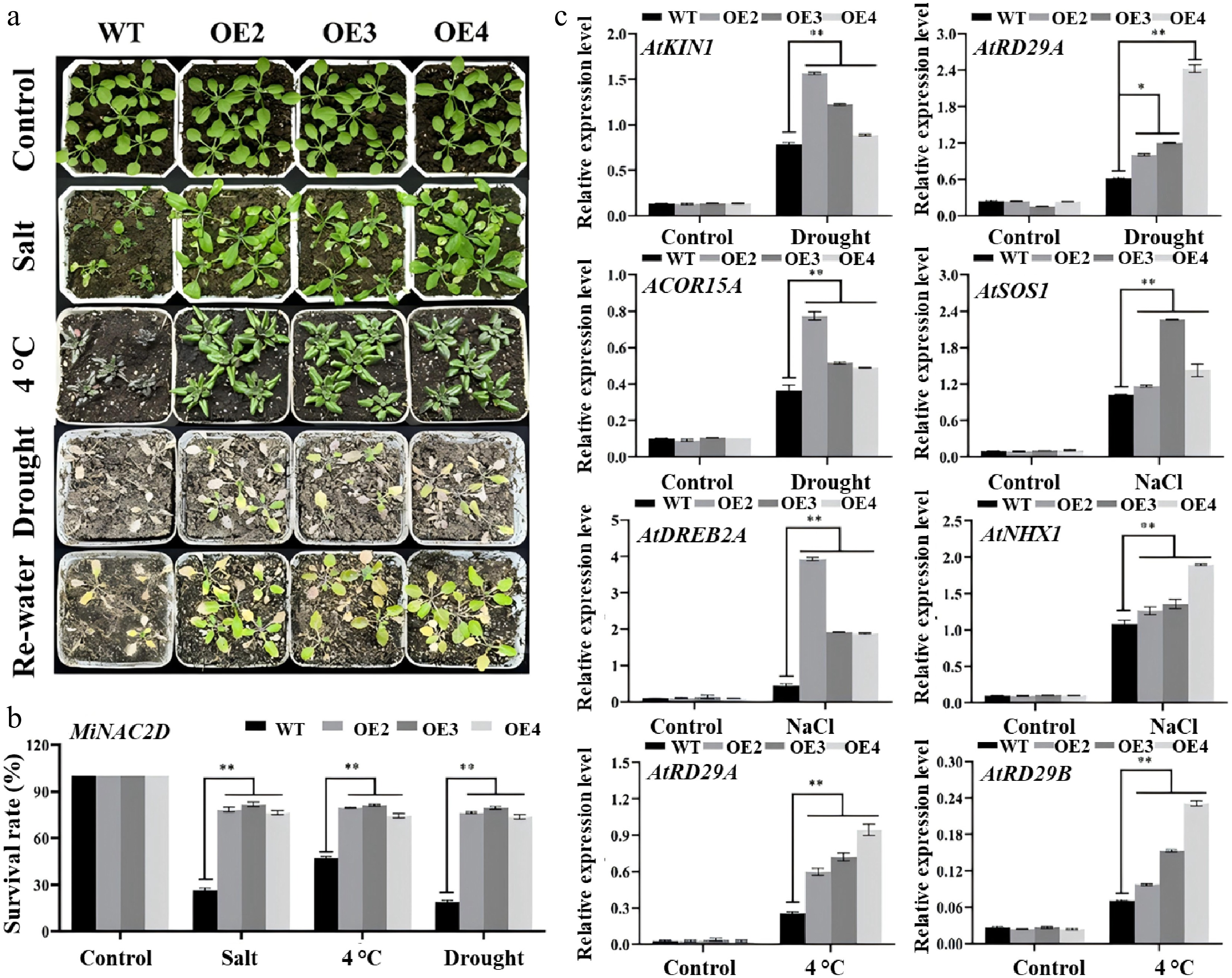
Figure 5.
MiNAC2D enhanced abiotic stress tolerance in Arabidopsis. (a) Growth phenotype of WT and MiNAC2D-OE lines. (b) Survival rate analysis of WT and MiNAC2D-OE lines. (c) Expression levels of the stress-related genes in WT and MiNAC2D-OE lines. Significant differences were defined as p < 0.01 (**) according to Tukey's t-tests.
The expression levels of drought stress-related genes (AtKIN1, AtRD29A, AtCOR15A), salt stress-responsive genes (AtSOS1, AtDREB2A, AtNHX1), and cold-regulated genes (AtRD29A and AtRD29B) were examined in MiNAC2D-OE and WT plants by RT-qPCR. Under normal conditions, most of these genes were expressed at low levels. Under salt, drought, and cold stress conditions, the expression levels of most of these genes were elevated, and their expression levels in the MiNAC2D-OE lines were significantly higher than those in WT. Collectively, these results suggest that MiNAC2D positively regulates salt, drought, and cold stress responses in Arabidopsis (Fig. 5c).
MiNAC2D enhances Arabidopsis tolerance by boosting antioxidant enzymes and modulating osmotic homeostasis
-
Under normal conditions, no significant differences were observed in DAB or NBT staining between the WT and MiNAC2D-OE lines. However, under drought or salt stress, the staining intensity in WT was stronger than that in the MiNAC2D-OE lines. Evans blue staining revealed less blue product accumulation in MiNAC2D-OE plants than in WT plants, indicating that cell death was more severe in WT leaves than in MiNAC2D-OE leaves (Fig. 6a).
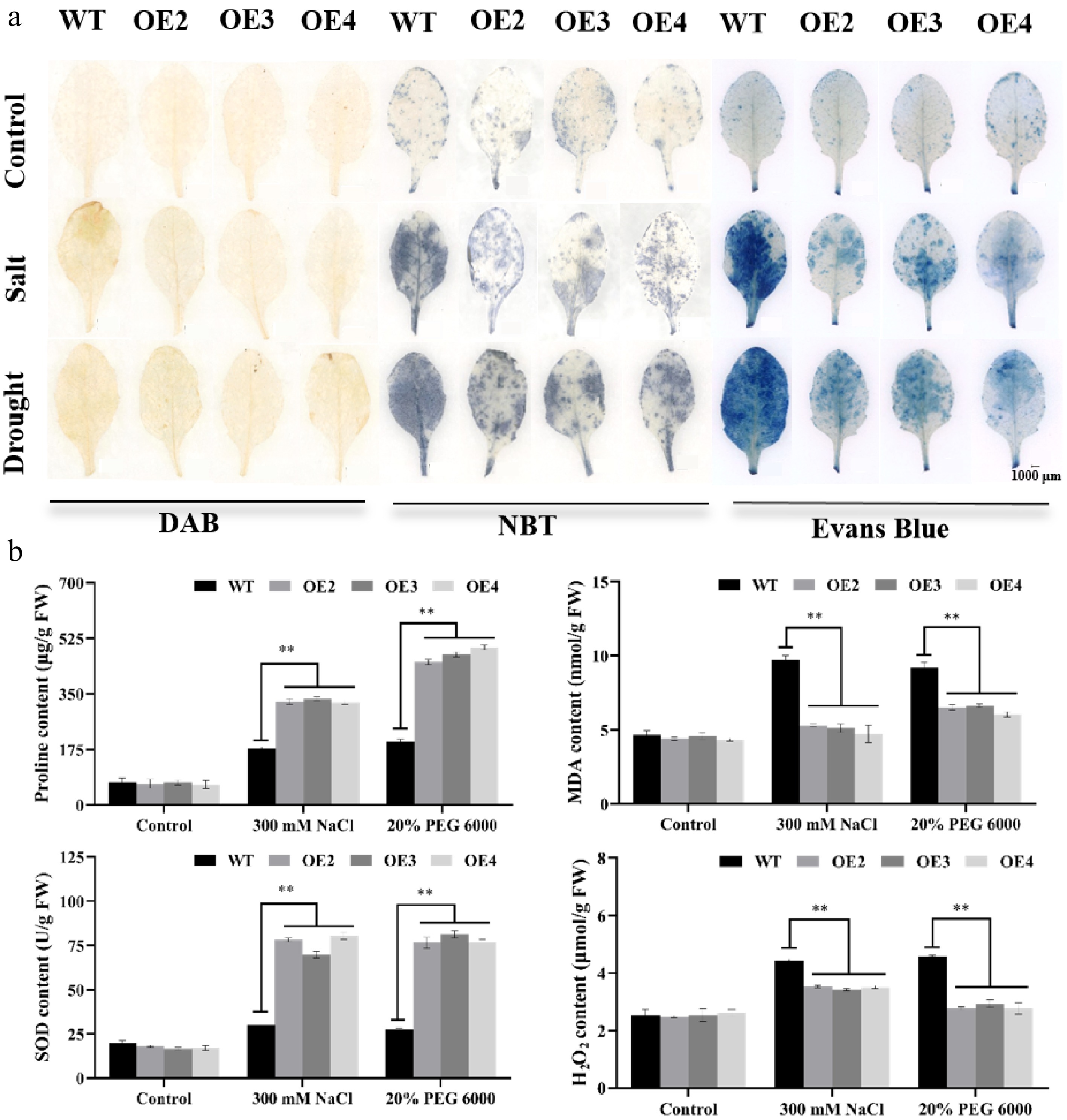
Figure 6.
MiNAC2D enhances Arabidopsis tolerance by boosting antioxidant enzymes and modulating osmotic homeostasis. (a) Phenotypes of WT and MiNAC2D-OE leaves stained with DAB, NBT, and Evans Blue. (b) SOD activity as well as the content of Pro, H2O2, and MDA in WT and MiNAC2D-OE lines. Significant differences were defined as p < 0.01 (**) according to Tukey's t-tests.
SOD activity, as well as Pro, H2O2, and MDA contents, were measured. Under normal conditions, no significant differences were detected between WT and MiNAC2D-OE lines in any of these parameters. Under salt or drought stress conditions, MDA and H2O2 contents in the MiNAC2D-OE lines were significantly lower than those in WT, whereas SOD activity and Pro content were significantly higher. These results suggest that MiNAC2D enhances tolerance to salt and drought stresses, potentially through the enhancement of antioxidant enzyme activity and the regulation of osmotic homeostasis (Fig. 6b).
Germination rate and root parameters of WT and MiNAC2D-OE tomato under abiotic stresses
-
WT and MiNAC2D-OE transgenic tomato lines were treated with 300 mM mannitol, 100 mM NaCl, low temperature (4 °C), and 10 μM ABA. The results showed that under NaCl, 4 °C, and mannitol treatments, the germination rate of MiNAC2D-OE lines was significantly higher than that of WT, whereas under ABA treatment, the germination rate of MiNAC2D-OE lines was significantly lower than that of WT (Fig. 7a; Supplementary Fig. S3). These findings in MiNAC2D transgenic tomatoes were consistent with those observed in MiNAC2D transgenic Arabidopsis, further supporting the role of MiNAC2D in regulating germination under stress conditions.
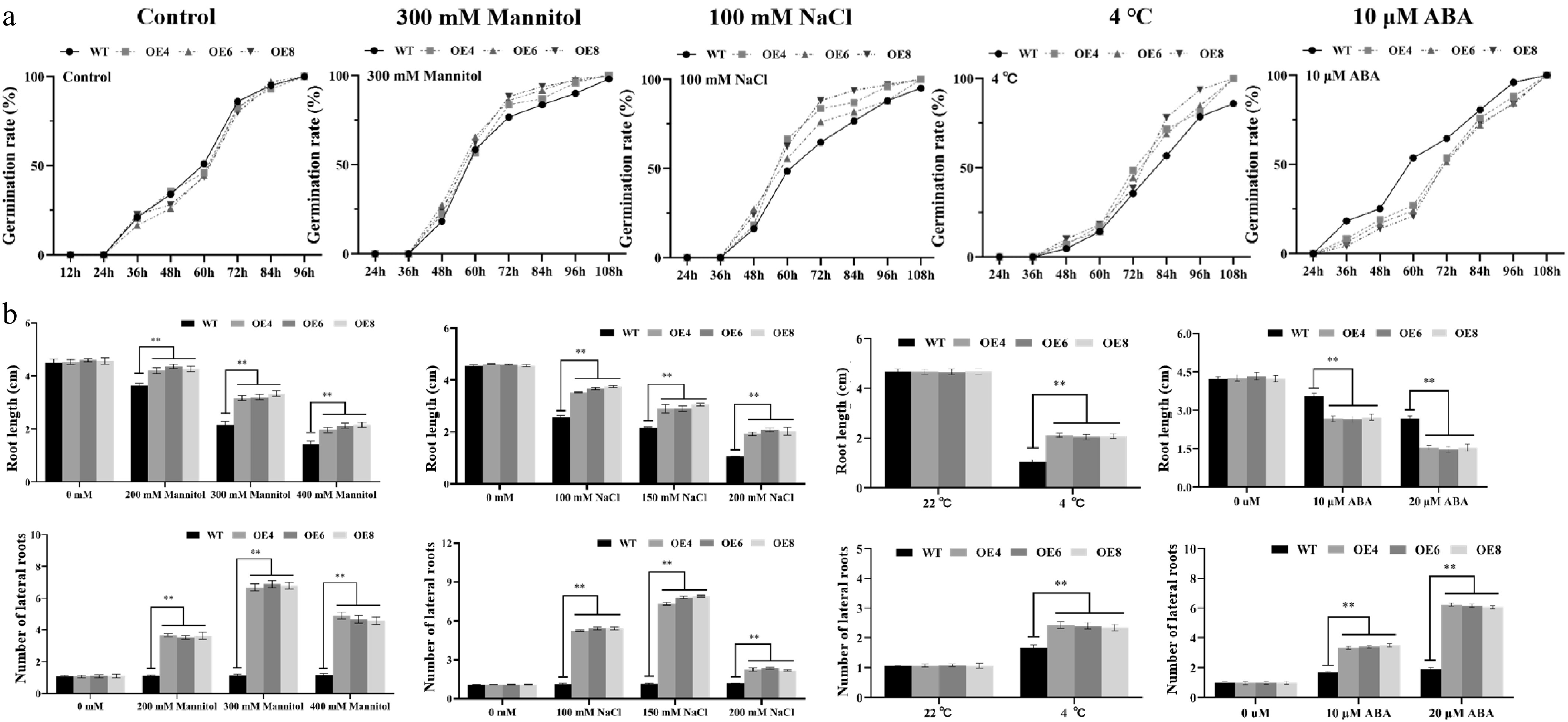
Figure 7.
Performance of WT and MiNAC2D-OE tomato under drought, salt, 4 °C and ABA treatment. (a) Germination rate analysis of WT and MiNAC2D-OE lines. (b) Root length and lateral root number of WT and MiNAC2D-OE lines. Significant differences were defined as p < 0.01 (**) according to Tukey's t-tests.
Subsequently, WT and MiNAC2D transgenic tomato seedlings grown in petri dishes were subjected to the same stress treatments as those applied to MiNAC2D transgenic Arabidopsis, and their root system characteristics were measured. The results were consistent with those obtained in MiNAC2D transgenic Arabidopsis, further confirming the function of MiNAC2D in root development under stress conditions (Fig. 7b; Supplementary Fig. S4).
MiNAC2D enhanced abiotic stresses tolerance in tomato
-
The survival rate was evaluated in WT and MiNAC2D-OE tomato plants subjected to three stress treatments: Spraying with 300 mM NaCl for 3 d, low temperature at 4 °C for 3 d, and natural drought for 7 d followed by 5 d of rehydration. Under normal growth conditions, both WT and MiNAC2D-OE plants remained turgid and green, with no signs of wilting or drying. Under salt stress, most transgenic plants retained green leaves, with only a few showing drying and yellowing, whereas the majority of WT plants wilted, lost their green color, and eventually dried up and died. Following drought treatment and subsequent rehydration, most MiNAC2D-OE plants remained green, with only a few exhibiting loss of green color and desiccation, while most WT plants wilted and perished. Under low temperature treatment, MiNAC2D-OE plants displayed downward leaf curling and deepened leaf color; in contrast, WT plants showed stunted growth, more pronounced leaf curling, and a change in color of the main stems and leaf undersides from dark green to purplish-red. Across all treatments, the survival rate of MiNAC2D-OE lines was higher than that of WT (Fig. 8a, b).
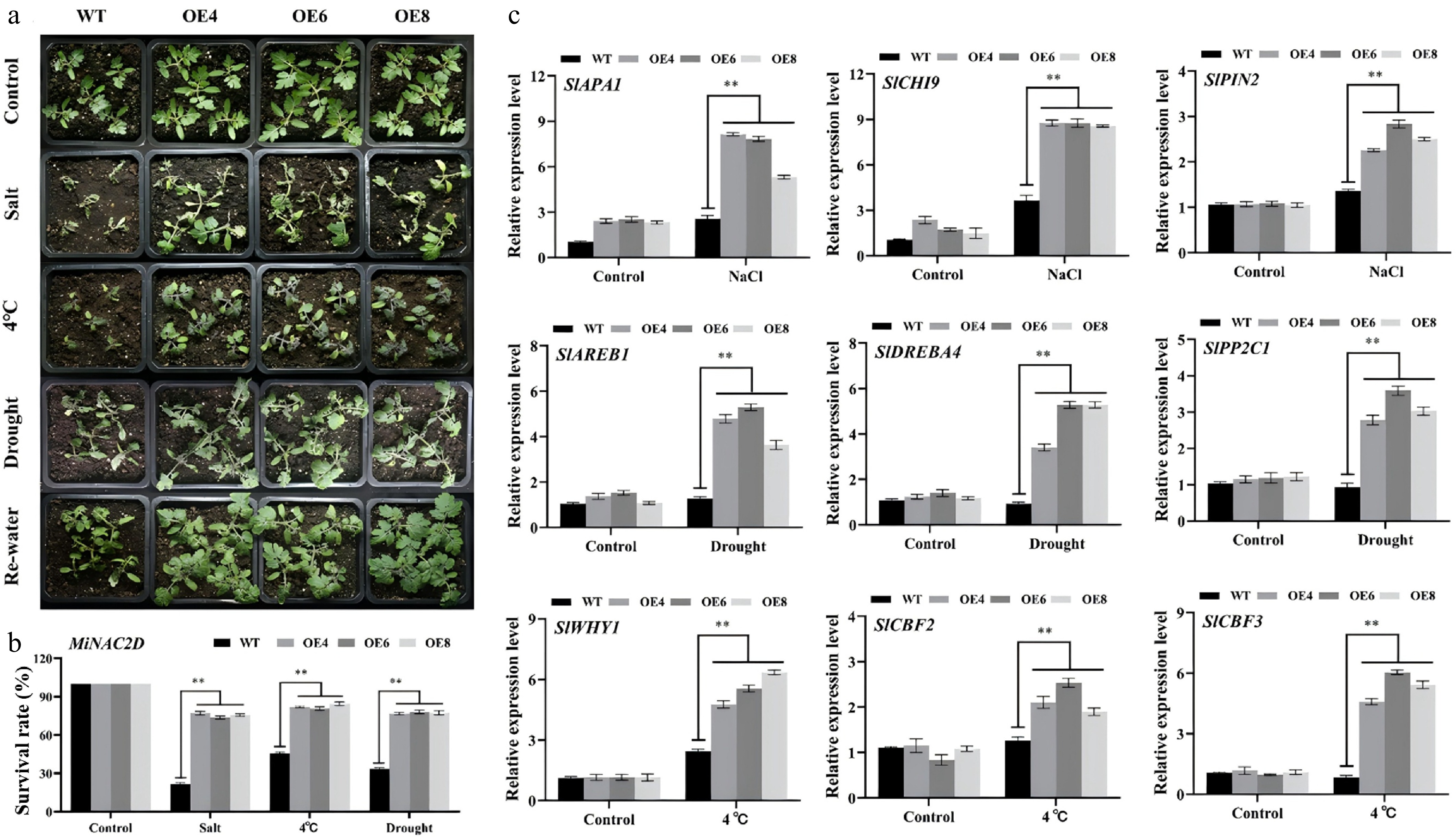
Figure 8.
MiNAC2D enhanced abiotic stress tolerance in tomato. (a) Growth phenotype of WT and MiNAC2D-OE lines. (b) Survival rate analysis of WT and MiNAC2D-OE lines. (c) Expression levels of the stress-related genes in WT and MiNAC2D-OE lines. Significant differences were defined as p < 0.01 (**) according to Tukey's t-tests.
RT-qPCR was used to examine the expression levels of stress-related endogenous genes in MiNAC2D-OE and WT plants. Salt stress-related genes included SlCHI9, SlAPA1, and SlPIN2; drought stress-related genes included SlPP2C1, SlAREB1, and SlDREBA4; and low temperature stress-related genes included SlWHY1, SlCBF2, and SlCBF3. Under normal conditions, no significant differences in the expression levels of these genes were observed between WT and MiNAC2D-OE lines. However, following stress treatment, the expression levels of these stress-related genes were significantly higher in MiNAC2D-OE lines than in WT plants, suggesting that MiNAC2D contributes to abiotic stress adaptation (Fig. 8c).
MiNAC2D enhanced abiotic stress tolerance in mango
-
While heterologous expression of MiNAC2D in Arabidopsis and tomato established its role in enhancing abiotic stress tolerance, its function in the native mango background remains to be confirmed. To this end, we generated transgenic mango plants expressing MiNAC2D (OE1-OE7) (Fig. 9a, b; Supplementary Fig. S5). Subsequently, both WT and transgenic plants were separately subjected to salt and drought treatments to verify the gene's tolerance function and elucidate its underlying mechanism in mango. The activity of SOD, together with the contents of Pro, H2O2, and MDA, was measured in WT and transgenic mango (OE2 under salt stress, OE5 under drought stress). Under normal conditions, no significant differences were observed between the WT and OE line in terms of SOD activity or the levels of Pro, H2O2, and MDA. However, under both salt and drought stress conditions, the OE line exhibited significantly lower MDA and H2O2 contents compared to the WT, while SOD activity and Pro content were significantly higher. These findings indicate that MiNAC2D enhances salt and drought stress tolerance in mango, likely through boosting antioxidant enzyme activity and regulating osmotic homeostasis (Fig. 9c, d).
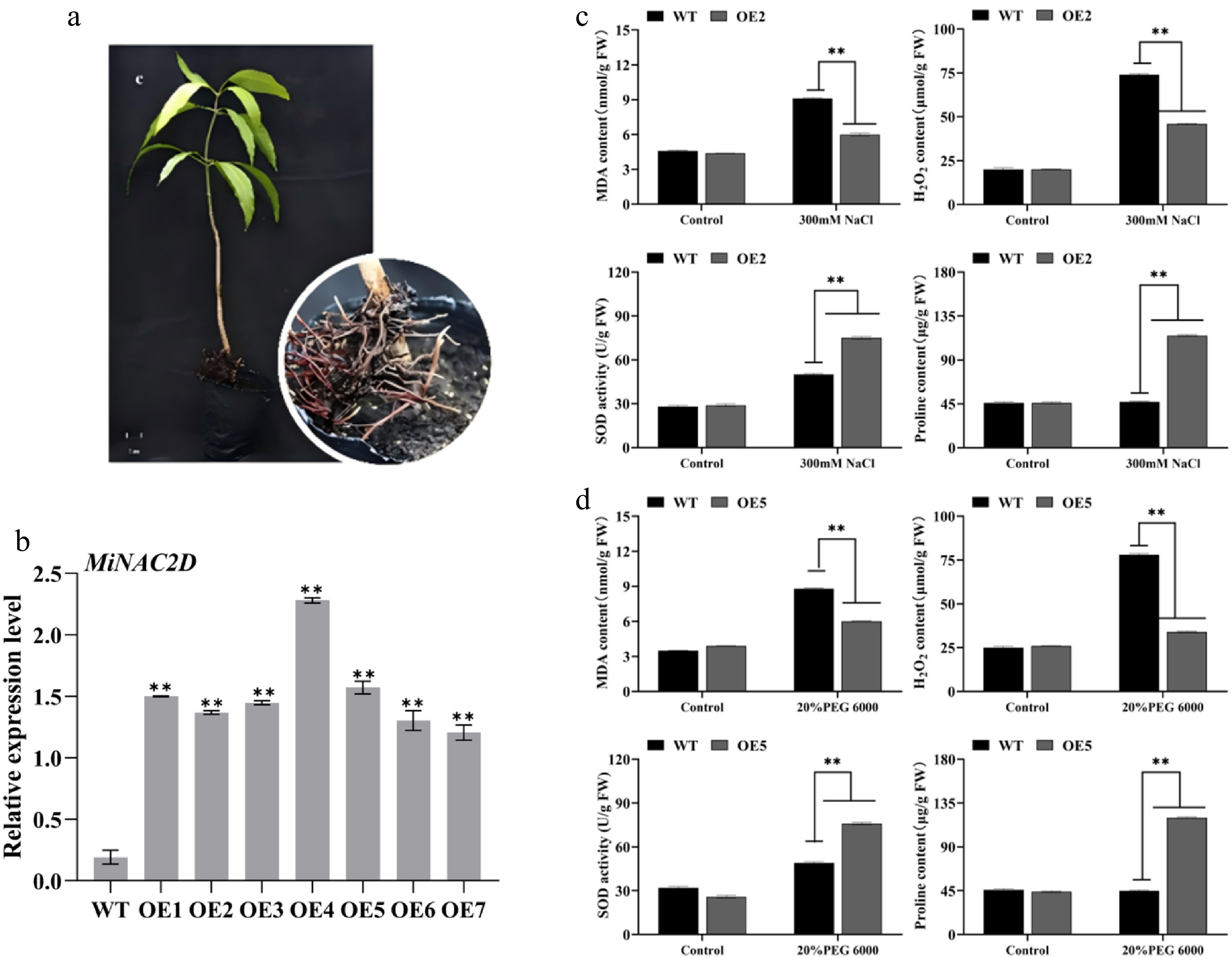
Figure 9.
MiNAC2D enhanced abiotic stress tolerance in mango. (a), (b) Overexpression of MiNAC2D in the root system of mango. (c) Physiological index changes of WT and OE2 under 300 mM NaCl salt stress. MDA content, H2O2 content, SOD activity and proline content were determined in control and salt-treated seedlings. (d) Physiological index changes of WT and OE5 under 20% PEG 6000 drought stress. MDA content, H2O2 content, SOD activity and proline content were determined in control and drought-treated seedlings. Significant differences were defined as p < 0.01 (**) according to Tukey's t-tests.
MiNAC2D mediates drought and salt tolerance through distinct signaling pathways
-
Through transcriptome sequencing combined with KEGG pathway analysis, we systematically examined gene distribution characteristics in WT and transgenic mango lines. Under normal growth conditions, OE lines showed 1,104 genes in the 'Global and overview maps' category of metabolic pathways compared to WT, with significant differential expression in secondary metabolic pathways such as carbohydrate and amino acid metabolism. Additionally, pathways including genetic information processing, environmental information processing, cellular processes, and organism systems exhibited notable gene expression differences (Fig. 10a), providing molecular evidence for MiNAC2D's regulation of fundamental physiological processes in mango. Under drought stress conditions, OE lines showed 787 genes differentially expressed in the 'Global and overview maps' category compared to WT. Although the number of differentially expressed genes in carbohydrate and amino acid metabolism pathways decreased, it remained significantly higher than in other pathways. It is worth noting that during the processing of environmental information, the 'signal transduction' pathway of the OE lines contains 145 genes, which show significant differences from the WT levels at the same time, while maintaining specific gene expression patterns in pathways like genetic information processing and cellular processes (Fig. 10b). The differential expression of pathway genes under drought stress indicates that MiNAC2D overexpression activates mango's drought response mechanism through remodeling the gene expression network, which regulates signal transduction and metabolic processes.
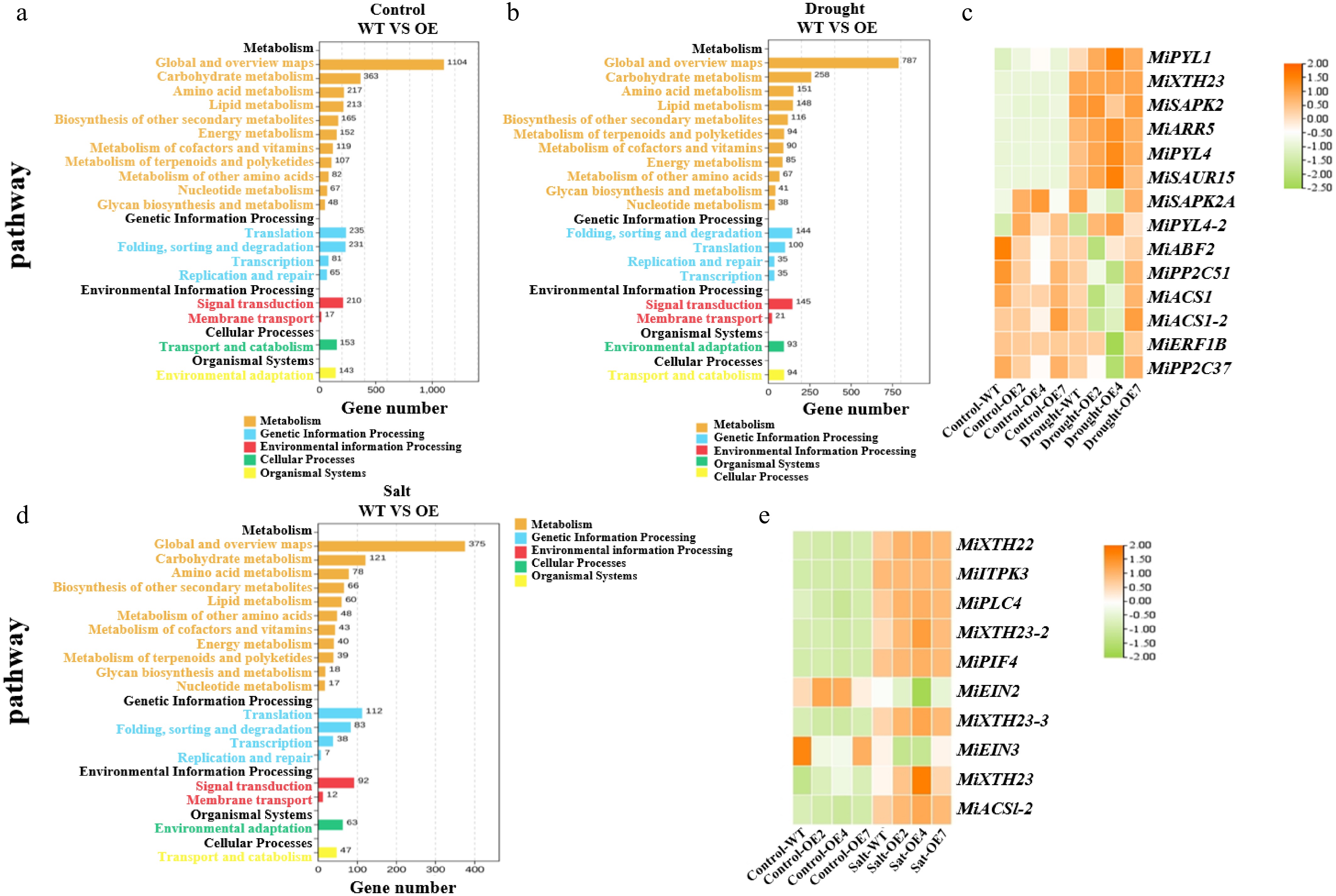
Figure 10.
MiNAC2D mediates drought and salt tolerance through distinct signaling pathways. (a) KEGG pathway enrichment histogram in WT and OE lines under control conditions. (b) KEGG pathway enrichment histogram in WT and OE lines under drought conditions. (c) Heatmap analysis of candidate gene expression patterns under control and drought conditions. (d) KEGG pathway enrichment histogram in WT and OE lines under salt conditions. (e) Heatmap analysis of candidate gene expression under different treatment conditions.
Studies have demonstrated that plants can resist biotic and abiotic stresses through signal transduction[29]. Therefore, we selected 14 significantly differentially expressed genes from the signaling pathway analysis for expression pattern evaluation. Results showed that compared to the WT, OE lines exhibited significantly upregulated expression of genes including Pyrabactin Resistance 1-Like 1 (MiPYL1), MiPYL4, Xyloglucan Endotransglucosylase/Hydrolase 23 (MiXTH23), Stress-Activated Protein Kinase 2 (MiSAPK2), Arabidopsis Response Regulator 5 (MiARR5), and Small Auxin-Up RNA 15 (MiSAUR15), while ABA-responsive Element Binding Factor 2 (MiABF2) and Protein Phosphatase 2C 51 (MiPP2C51) genes showed no significant differences or slight downregulation under normal conditions. This indicates that MiNAC2D can regulate the basal expression of certain ABA signaling and growth-related genes under normal conditions. After drought stress treatment, the upregulation trend of key ABA signaling genes such as MiPYL1, MiXTH23, and MiSAPK2 was further enhanced in OE lines, while expression patterns of genes including MiABF2, MiPP2C51, and 1-Aminocyclopropane-1-Carboxylate Synthase 1 (MiACS1) underwent significant adjustments. Comparative analysis of expression patterns under normal and drought-stressed conditions revealed that both WT and OE lines exhibited significant expression changes in most target genes under drought stress. For instance, the upregulation of MiPYL1 and MiXTH23 under drought stress was significantly higher than under normal conditions, while genes like MiABF2 and MiPP2C51 showed more pronounced downregulation trends (Fig. 10c). This clearly demonstrates the drought stress-induced induction of ABA signaling and related pathway gene expression in mango.
To investigate the molecular regulatory mechanisms of MiNAC2D in mango's salt stress response, transcriptome sequencing was performed on WT and OE lines under salt stress, followed by KEGG pathway analysis. Compared with normal growth conditions, the 'Global and overview maps' in metabolic pathways contained 375 genes, with significant differences observed in secondary metabolic pathways such as carbohydrate metabolism (121 genes) and amino acid metabolism (78 genes). The signal transduction pathway in environmental information processing involved 92 genes, playing a central role in salt stress signal perception and transmission. Meanwhile, pathways related to genetic information processing, organism systems, and cellular processes also exhibited substantial gene expression variations (Fig. 10d), indicating that salt stress induces molecular responses in mango through multiple dimensions, including metabolic regulation, signal transduction, and genetic information processing.
We selected ten significantly differentially expressed genes from the signaling pathway analysis for expression pattern evaluation. Under control conditions, compared to the WT, OE lines exhibited varying degrees of upregulated expression of genes such as MiXTH22, MiXTH23-2, Inositol-Trisphosphate 3-Kinase 3 (MiITPK3), Phospholipase C 4 (MiPLC4), and Phytochrome Interacting Factor 4 (MiPIF4), while Ethylene Insensitive 2 (MiEIN2) showed no significant difference or slight downregulation. This indicates that MiNAC2D can regulate the basal expression of certain signal transduction and metabolism-related genes under normal conditions. After salt stress treatment, the upregulation trend of genes like MiXTH23 and MiACS1-2 in OE lines was further enhanced, with significant adjustments in the expression patterns of genes such as MiEIN2 and MiEIN3. Meanwhile, the upregulation levels of genes including MiXTH23-2, MiXTH23-3, MiPIF4, and MiPLC4 in OE lines were significantly higher than in WT (Fig. 10e). Comparative analysis of expression patterns under normal and salt-stressed conditions revealed that salt stress significantly induced expression changes in genes related to ion concentration regulation, such as MiITPK3, MiPLC4, and MiPIF4. In summary, MiNAC2D can enhance gene regulation in ion metabolism-related signaling pathways, modulate XTH family genes (involved in cell wall modification), EIN family genes (participating in ethylene signal transduction), and ACS genes (involved in ethylene synthesis), thereby activating downstream salt stress response networks and improving mango's salt stress tolerance.
-
The remarkable functional diversity of NAC transcription factors stems from their conserved-variable architecture. While they share a highly conserved N-terminal DNA-binding domain of about 150–160 amino acids, their C-terminal ends contain transcriptional activation regions that are highly diverse. This structural design allows a single protein family to govern pivotal roles across virtually all phases of growth and development, alongside adaptation to numerous stress conditions[30,31]. The MiNAC2D gene was identified in the mango genome. NAC2 genes have been identified and analyzed in several species and vary among species, but the mango MiNAC2 gene clustered with the Pistacia vera PvNAC2 gene because they belong to the same family, Anacardiaceae.
Drought stress is a major natural disaster that severely impacts agricultural production, particularly in water-scarce regions, thereby limiting agricultural development[32]. Research on drought stress in crops is crucial for mitigating its harmful effects. Temperature, another key factor, also affects plant growth; extreme cold can stunt growth or even kill plants[33]. Soil salinization is globally recognized as one of the geo-environmental problems that poses a threat to the biosphere and ecosystems. In high-salt environments, the reduction in soil water potential induces water stress in plants, leading to cellular water loss, reduced physiological activity, and even death[34].
The NAC family of transcription factors is not only widely involved in plant growth and development but also plays an important role in stress responses. In this study, the expression level of MiNAC2D in leaves was significantly higher than that in other tissues. In rice, the Oryza sativa NAC transcription factor 19 (OsNAC19) gene was expressed at relatively high levels in the roots, culms, and leaf sheaths of seedlings[35,36] identified two salt-sensitive NAC genes from tomato, Solanum lycopersicum NAC transcription factor 1 (SlNAC1) and Solanum lycopersicum NAM transcription factor 1 (SlNAM1). Tissue expression analysis showed that SlNAC1 was expressed at high levels in flowers and ripe tomato fruits[36]. SlNAC2 was expressed in a variety of tissues, with the highest expression in leaves, and expression analysis indicated that it was strongly induced when leaves were subjected to salt stress and low temperature stress[37]. In the present study, MiNAC2D transcript levels initially increased and then decreased under low temperature stress, but overall showed an up-regulated trend, suggesting its involvement in abiotic stress responses. Similarly, the Saccharum spp. NAC transcription factor 23 (SsNAC23) gene was induced at 4 °C but not at 12 °C, indicating its role in the response of sugarcane to extreme low temperature stress[38]. Overexpression of the Triticum aestivum NAC transcription factor 69 (TaNAC69) gene resulted in increased transcript levels of drought stress-upregulated genes in the leaves and roots of transgenic plants, thereby enhancing drought tolerance[39]. Additionally, Triticum aestivum root-preferential NAC transcription factor 1 (TaRNAC1) is specifically expressed in wheat roots; transgenic wheat overexpressing TaRNAC1 in roots exhibited increased root length, biomass, and drought resistance[40]. Genetic engineering was used to preliminarily characterize the function of MiNAC2D overexpression in Arabidopsis, tomato, and mango. The results showed that this gene confers significant tolerance to abiotic stress across different species[41]. Under normal growth conditions, Arabidopsis plants overexpressing MiNAC2D did not differ from WT in germination rate. Under salt, drought, and low temperature stress, MiNAC2D overexpression plants exhibited higher germination rates than WT plants; root length and survival rates similarly indicated enhanced tolerance to these stresses. Under drought stress, salt stress, and ABA treatment, transgenic Arabidopsis overexpressing the peanut NAC transcription factor 2 (AhNAC2) showed increased ABA sensitivity, but exhibited significantly higher growth and survival rates than WT plants, along with up-regulated expression of stress-related genes[42]. The tomato NAC transcription factor 35 (SlNAC35) gene was induced by drought stress and salt stress. Overexpression of SlNAC35 in tobacco increased lateral root number and root length in transgenic plants[43]. Rice transgenic plants overexpressing the Eleusine coracana NAC transcription factor 67 (EcNAC67) gene exhibited greater drought and salinity tolerance and grew more roots than WT plants under stress[44]. Salt stress, drought stress, and ABA treatment all inhibit the transcriptional expression of SlNAC3. This gene may interact with environmental and endogenous signals and function as a transcriptional regulator in plant responses to salt and drought stress via the ABA signaling pathway[45]. Expression of the rice Stress-Responsive NAC 1 (SNAC1) gene results in increased ABA sensitivity, which further leads to significant inhibition of transgenic plant growth[46]. Oryza sativa NAC transcription factor 2 (OsNAC2) regulates rice responses to abiotic stress by binding to the promoters of key genes in the ABA signaling pathway and repressing their expression[47].
Physiological index and histochemical staining analyses revealed that Evans blue, DAB, and NBT staining were significantly more intense in WT plants than in MiNAC2D-OE Arabidopsis lines. In contrast, SOD activity and proline content were higher in the OE lines than in WT plants, whereas MDA and H2O2 contents were lower. Overall, overexpression of MiNAC2D enhanced the regulation of stress-responsive substances and improved tolerance to drought and salt stress. Overexpression of Oryza sativa NAC transcription factor 6 (OsNAC6) induced the expression of a peroxidase gene, and transgenic plants showed improved tolerance to dehydration and high salt stress[48]. Under stress conditions, H2O2 and MDA levels were reduced in plants overexpressing the Stress-Responsive NAC 3 (SNAC3) gene, and the expression of genes related to reactive oxygen species (ROS) scavenging was up-regulated[49]. Arabidopsis plants overexpressing the Triticum aestivum NAC transcription factor 29 (TaNAC29) gene exhibited reduced MDA and H2O2 contents, increased SOD and CAT activities, and enhanced tolerance to drought and salt stress[50].
In our study, the expression of genes such as MiPYL1 and MiPYL4 was significantly upregulated under drought stress, consistent with previous reports that PYL functions as an ABA receptor in drought response[51,52]. This suggests that these genes may participate in drought stress response by sensing ABA signals. MiABF2 showed significant upregulation in control WT plants but downregulation in the drought-treated OE lines, aligning with the proposal that ABF acts as a binding factor for ABA-responsive elements, highlighting its transcriptional regulatory role in the ABA signaling pathway[53]. Meanwhile, the differential expression of MiPP2C51 and MiPP2C37 under drought treatment is consistent with previous findings that PP2C family genes negatively regulate drought resistance via the ABA pathway[54,55]. The expression changes of MiACS1 and MiACS1-2 support the hypothesis by Wu et al. that ACS accelerates ethylene synthesis to negatively regulate drought resistance[56], while the expression pattern of MiSAUR15 corresponds to the previously proposed mechanism in which SAUR inhibits stomatal closure, thereby reducing drought tolerance[57]. SAPK2 primarily regulates drought resistance through nitrogen metabolism[58], and the upregulated expression of MiSAPK2 in this study further supports this perspective. These findings demonstrate that the drought-stress-induced differential gene expression patterns in this study are highly consistent with the reported ABA signaling pathway and related metabolic and structural maintenance pathways. The expression differences among genotypes (WT and OE lines) in the heat map further suggest genotype-specific regulation of these genes, providing valuable insights for exploring drought tolerance mechanisms involving gene-gene and genotype-gene interactions.
In the salt stress treatment groups, the high expression of MiITPK3, MiXTHs, and MiPLC4 aligns with previous reports that ITPK3 regulates Zn2+ concentration and PLC4 positively modulates salt-induced Ca2+ elevation[59,60]. This suggests that these genes may participate in salt stress response through ion concentration regulation and calcium signaling. The expression changes of MiPIF4 also support the view of Xu et al. that PIF4 plays a key role in multiple signaling pathways, indicating its potential integration of light and hormone signals under salt stress[61]. For ethylene-related genes, the differential expression of MiEIN2 and MiEIN3 under salt treatment corresponds to the proposed mechanism by Zhang et al.[62], in which EIN2 and EIN3 counter-regulate salt stress through ethylene-induced stomatal closure. The MiACS1-2 gene shows significantly upregulated expression under salt stress. Xu et al. demonstrated that ethylene synergizes with melatonin to regulate plant salt tolerance[63]. Meanwhile, the genotype-specific expression differences in the heatmap indicate genotype-specific regulation of ethylene signaling in salt stress response, providing insights for future research on gene-gene interaction mechanisms underlying salt tolerance.
-
Transgenic plants overexpressing MiNAC2D exhibited enhanced survival rate, growth recovery ability, and stability of physiological indexes under various stress conditions, including drought, salt, and low temperature stress. This effectively strengthened the plant stress tolerance mechanism by regulating stress-responsive signaling pathways, providing a new strategy for molecular breeding aimed at enhancing resilience in multiple species, and offering theoretical support for dissecting the molecular network underlying plant stress adaptation.
-
The authors confirm contributions to the paper as follows: conceptualization, data curation, writing-original draft: Guo T, Xu F; methodology: Guo T, Xu F, Yang X, Liu Y, Liu H, Zhou X, He X, Luo C; investigation: Yang X, Liu Y, Liu H, Zhou X; writing-review & editing: Guo T, Xu F, He X, Luo C; project administration, resources: He X, Luo C. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
This work was supported by Guangxi Natural Science Foundation (Grant No. 2023GXNSFAA026268), Guangxi Science and Technology Base and Talent Special Project (Grant No. GuikeAD25069107), the CARSGIT-Guangxi Mango Industry Project (Grant No. nycytxgxcxtd-2021-06-02), the six one's special action of 'strengthening agriculture and enriching people' by science and technology vanguard (Grant No. 202604) and Funded by the Second Batch of Guangxi Qingmiao Talent Inclusive Support Policy (Research Start-up Fund).
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/frures-0026-0018.
-
# Authors contributed equally: Tianli Guo, Fang Xu
- Supplementary Table S1 Primer sequences used in this study.
- Supplementary Fig. S1 Germination phenotype of WT and MiNAC2D-OE lines.
- Supplementary Fig. S2 Growth phenotype of WT and MiNAC2D-OE lines.
- Supplementary Fig. S3 Germination phenotype of WT and MiNAC2D-OE lines.
- Supplementary Fig. S4 Growth phenotype of WT and MiNAC2D-OE lines.
- Supplementary Fig. S5 PCR assay of transgenic mango.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Guo T, Xu F, Yang X, Liu Y, Liu H, et al. 2026. Mango MiNAC2D enhances drought, low temperature, and salt stress tolerance in transgenic plants. Fruit Research 6: e025 doi: 10.48130/frures-0026-0018 

Mango MiNAC2D enhances drought, low temperature, and salt stress tolerance in transgenic plants
- Received: 01 April 2026
- Revised: 14 May 2026
- Accepted: 22 May 2026
- Published online: 10 July 2026
Abstract: The NAC transcription factor family represents a unique class of plant-specific regulators involved in abiotic stress responses. In this study, a MiNAC2D gene was obtained from the mango genome and transcriptome. Subcellular localization confirmed the nuclear localization of MiNAC2D. Expression analysis revealed that its transcript levels were highest in leaves and were significantly upregulated under drought, low temperature, and salt stresses. To investigate its biological function, MiNAC2D was overexpressed in Arabidopsis, tomato, and mango. Transgenic Arabidopsis and tomato plants showed enhanced performance under abiotic stress conditions, including higher seed germination rates, increased root length, and improved survival rates. In addition, transgenic plants displayed significantly reduced leaf damage compared to wild-type (WT) controls. Transcriptome profiling of WT and transgenic mango roots under drought stress identified 145 differentially expressed genes involved in signaling pathways, 14 of which were validated by RT-qPCR. Under salt stress, 92 differentially expressed signaling pathway-related genes were identified, with ten confirmed by RT-qPCR. These results suggest that MiNAC2D may enhance drought tolerance through the abscisic acid (ABA) pathway and improve salt tolerance via mechanisms involving ion homeostasis and the ethylene pathway. Overall, this study demonstrates the role of MiNAC2D in enhancing abiotic stress tolerance and provides a foundation for further exploration of its regulatory network in mango.
-
Key words:
- Mango /
- MiNAC2D /
- Expression analysis /
- Functional analysis