








-
Apples (Malus domestica Borkh.), belonging to the Rosaceae family and the genus Malus Mill, have been widely cultivated worldwide with a long agricultural history[1]. Dwarf dense planting has emerged as the current trend in global apple production. While dwarfing clonal rootstocks are commonly adopted in foreign countries, China still faces a shortage of superior dwarfing clonal rootstocks[1,2]. Dwarfing rootstocks offer several advantages, including early fruiting, high yield, improved fruit quality, compatibility with mechanization, and reduced labor costs[1]. However, apple rootstock breeding is a time-consuming process. Conventional breeding methods often require extensive screening of progeny populations to ensure broad adaptability and multiple resistances, which typically takes several decades from the selection of superior lines to commercialization[3]. With the rapid advancements in molecular biology, bioinformatics, and genomics, modern breeding technologies have been increasingly applied to apple rootstock improvement. These technologies hold great promise for accelerating the breeding cycle, enhancing rooting ability, and improving resistance traits, thereby providing novel technical solutions for the genetic enhancement of apple rootstocks.
Among these modern breeding technologies, functional gene validation and precision trait improvement represent critical steps in accelerating the breeding process. To this end, the establishment of efficient and reliable genetic transformation systems is imperative. Within the spectrum of transformation methodologies, Agrobacterium-mediated transformation has been established as a fundamental technology for genetic manipulation in higher plants, owing to its capacity for stable gene integration and expression. Two primary Agrobacterium strains are commonly employed in plant genetic engineering: A. tumefaciens, harboring the Ti plasmid and inducing crown gall formation[4], and Agrobacterium rhizogenes, carrying the Ri plasmid and triggering hairy root development[4,5]. Among these, A. tumefaciens-mediated transformation is the most prevalent method for generating transgenic apples, renowned for its stable transgene expression. However, this approach is encumbered by a laborious workflow, protracted transformation period, and relatively low efficiency, typically yielding transformation rates of merely 0%–3%[6,7]. In recent years, genetic transformation in apple rootstocks has been progressively documented. However, apart from the well-established genetic transformation systems in a limited number of rootstocks such as M.26, M9-T337, and M.7, the development of such systems for other rootstocks remains challenging, which severely impedes the progress of apple rootstock breeding[8,9]. Agrobacterium rhizogenes-mediated genetic transformation has gained increasing application in genetic engineering due to its merits of high transformation efficiency, streamlined procedures, and shorter experimental duration[4,10]. A. rhizogenes-induced hairy roots provide an ideal platform for investigating gene function and secondary metabolism[11,12], particularly in dissecting the molecular mechanisms underlying soil stress responses[13,14]. Leveraging this system, researchers can effectively explore and validate the functions of stress-related genes, thereby enhancing plant adaptability and stress tolerance[4,5,15]. The Agrobacterium rhizogenes-mediated hairy root induction technique has been widely applied in transgenic research of various plants, such as Solanum lycopersicum L. (tomatoes)[16], Citrus reticulata Blanco (citrus)[17], Glycine max (L.) Merr (soybeans)[18], and Pyrus betulifolia Bunge (pears)[19]. Existing studies have revealed substantial disparities in hairy root induction rates when employing different A. rhizogenes infection strategies. For instance, in 22 citrus species, transformation efficiencies span from 0% to 95%[20]; soaking Phoebe bournei explants in the bacterial solution achieved a 41.2% hairy root induction rate[21]. In the case of 'Golden Delicious' apples, the rooting induction rate fluctuates between 0% and 20%, with efficiency being contingent upon the specific bacterial strain[22]. Notably, hairy root induction rates vary significantly among woody plants, and apples particularly demonstrate relatively low transformation efficiencies[10,22−24]. Multiple factors contribute to these variations, including the A. rhizogenes strain type, explant characteristics, infection methods, and infection conditions[24,25]. These findings underscore the complexity of optimizing hairy root induction protocols across different plant species and highlight the need for tailored approaches in apple genetic research.
CX5, a dwarf apple rootstock line, was selected by the Research Institute of Pomology, Chinese Academy of Agricultural Sciences (CAAS), from 34 progenies of the cross 193 × M9[26,27]. Previous studies demonstrated that grafting major Chinese apple cultivars onto CX5 rootstock resulted in a survival rate exceeding 90%, indicating excellent graft compatibility[26]. When used as an interstem with Malus baccata as the rootstock, CX5 exhibited superior overwintering capacity compared to M6, as evidenced by higher survival rates of grafted saplings in western Liaoning. Thus, CX5 has strong winter hardiness[28]. In a separate study using M. baccata as the base rootstock and CX5, 75-9-5, or SH3 as interstocks for 'Honeycrisp' grafting, CX5 significantly increased the proportion of short branches, improved yield, and advanced the age of first fruiting[29]. When grafting 'Huahong' apples with M. baccata as the base and CX5 as the interstock, the total phenolic content in both the pericarp and pulp of CX5-grafted fruits was lower than that of the control group at maturity. This reduction suggests that CX5 may enhance resistance to postharvest browning in 'Huahong' apples[30]. Collectively, these findings demonstrate that CX5 possesses multiple advantageous traits, including high graft compatibility, superior overwintering ability, enhanced productivity, and potential for improving fruit quality. As such, CX5 represents a promising dwarfing rootstock for apple cultivation, particularly in cold regions, and warrants further investigation and widespread application in apple breeding and horticultural practices. However, it also has issues such as difficulty in rooting and a low propagation rate. Therefore, we hope to address this issue through genetic improvement.
In previous studies, we have demonstrated that leaf explants of the dwarf apple rootstock CX5 exhibit recalcitrance to adventitious shoot regeneration, with persistent failure in obtaining viable shoots[27,31]. Given this limitation, we chose to employ Agrobacterium rhizogenes-mediated transformation as an alternative approach to establish a genetic transformation system in CX5. Liu et al. identified MD13G1252700 as the predicted ortholog of AtAIL5 through phylogenetic analysis of Arabidopsis and apple AIL (AINTEGUMENTA-LIKE) proteins, designating it as MdAIL5[7]. Subsequent overexpression of MdAIL5 in the model apple line GL-3 significantly enhanced adventitious shoot regeneration efficiency and rooting capacities[7]. This provides a pragmatic strategy to overcome technical bottlenecks in generating transgenic plants in apple and other horticultural tree species. Consequently, this study employed A. rhizogenes-mediated transformation for CX5 genetic transformation. Through systematic optimization of critical parameters, including Agrobacterium strain selection, explant types, and infection duration, based on their impact on hairy root induction efficiency in apple, we established a robust genetic transformation system. This platform achieved stable integration of the MdAIL5 transgene into CX5 lines and demonstrated its promoting effect on adventitious rooting, thereby providing a foundational framework for future efforts to develop self-rooting rootstocks, to facilitate genetic improvement of recalcitrant apple germplasm, and enhance genetic improvement.
-
The explants utilized in this study consisted of in vitro shoots of the CX5 apple cultivar, maintained within the tissue culture collection at the National Apple Breeding Center, Research Institute of Pomology, Chinese Academy of Agricultural Sciences. The A. rhizogenes strains employed were K599, C58C1, and MSU440 (Weidi Biotechnology Ltd., Shanghai, China). The MdAIL5 gene was cloned from the apple cultivar GL-3, and the pRI101-MdAIL5-GFP overexpression vector, harboring a green fluorescent protein (GFP) tag, was generated previously in our laboratory[7]. Primers utilized for vector construction are detailed in Table 1 and were synthesized by Genewiz (Beijing, China). All molecular biology reagents, including DNA markers, were procured from Takara Biomedical Technology (Beijing, China) Co., Ltd.
Table 1. Primer list.
Primers Primer sequence Purpose MdAIL5-F CCTTGGAACTTTCAGCACCCA qRT-PCR MdAIL5-R TAGCGGCTCATGTCGAAGTT MdAIL5-SalI-PRI101-GFP-F CACTGTTGATACATATGCCCGTCGACATGGATTCTTCTCCTCAGAACTGG Vector construction MdAIL5-SalI-PRI101-GFP-R TCGGATCCGGTACCCCCGGGGTCGACTTCCATCCCAAAAATTGGTGT 35S-F CCTCGGATTCCATTGCC Transgenic identification 35S-R GCTTGGGGCGAAGGATAG MdActin-F TGACCGAATGAGCAAGGAAATTACT qRT-PCR MdActin-R TACTCAGCTTTGGCAATCCACATC A. rhizogenes transformation and infection solution preparation
-
A. rhizogenes-mediated genetic transformation was performed according to methodologies established by Tao et al.[25] and Feng et al.[32] with minor modifications.
Agrobacterium transformation and positive clone preparation
-
For genetic transformation, 5 μL of the pRI101-MdAIL5-GFP overexpression vector plasmid was transferred to 100 μL of competent cells for each A. rhizogenes strain (K599, C58C1, MSU440). The cell-plasmid mixture was subjected to sequential incubation: 5 min on ice, flash-freezing in liquid nitrogen for 5 min, heat-shock at 37 °C for 5 min, and a final ice-incubation for 5 min. Subsequently, 400 μL of YEB medium was added, and the culture was incubated with shaking (200 rpm, 28 °C) for 3 h to facilitate recovery. A 100 μL aliquot was plated onto LB agar supplemented with kanamycin (50 mg·L−1) and rifampicin (50 mg·L−1). Plates were inverted and incubated at 28 °C for 48 h. Putative positive clones were screened by PCR amplification using gene-specific primers (Table 1) and cryopreserved at –80 °C in 20% (v/v) glycerol for downstream applications
Infection solution preparation
-
Positive Agrobacterium clones were inoculated into 50 mL of YEB medium supplemented with 50 mg·L−1 kanamycin and 50 mg·L−1 streptomycin. Cultures were grown at 28 °C with agitation at 200 rpm until the optical density at 600 nm (OD600) reached 0.8 ± 0.1. Bacterial cells were harvested by centrifugation at 5,000 × g for 5 min at 28 °C. The resulting pellet was resuspended in sterile infiltration buffer (1/2 MS medium, pH 5.8) to a final OD600 of 0.5. The resuspended culture was immediately used for plant infection experiments.
Optimization of explant type and infection duration
-
To determine the optimal conditions for hairy root induction, three types of explants-leaves, apical buds, and stem segments were excised from CX5 in vitro- grown plants. Each explant was superficially wounded using a sterile scalpel and immersed in the bacterial suspension prepared as described for the A. rhizogenes transformation and infection solution preparation for immersion periods of 1, 5, 10, 15, or 20 min. Following infection, explants were gently blotted dry on sterile filter paper and co-cultivated on MS basal medium supplemented with 100 μM acetosyringone. Co-cultivation proceeded in darkness at 25 °C for 48 h. Each treatment consisted of 20 explants and was replicated three times. After 30 d of culture on selection medium (MS + 500 mg·L−1 cefotaxime + 50 mg·L−1 kanamycin), the number of explants exhibiting hairy roots was recorded. Transformation efficiency was calculated as follows:
$ \begin{split}&\text {Transformation efficiency }({\text{%}})\\=\;&\dfrac{\text {Number of explants with hairy roots}}{\text {Total number of inoculated explants}} \times 100 {\text{%}}\end{split} $ Optimization of A. rhizogenes strain and inoculum concentration
-
To identify the optimal A. rhizogenes strain and bacterial cell density for hairy root induction, three A. rhizogenes strains (K599, C58C1, and MSU440) were evaluated across five optical density ranges (OD600): 0–0.2, 0.2–0.4, 0.4–0.6, 0.6–0.8, and 0.8–1.0. Pre-wounded leaf explants from CX5 in vitro-grown plantlets were immersed in the respective bacterial suspensions for the predetermined optimal duration, gently blotted dry on sterile filter paper, and co-cultivated on MS basal medium supplemented with 100 μM acetosyringone. Following a 48-h co-cultivation period in darkness at 25 °C, explants were transferred to selection medium (MS + 500 mg·L−1 cefotaxime + 50 mg·L−1 kanamycin). Each treatment consisted of 20 explants with three biological replicates. The hairy root induction rate was quantified 30 d after inoculation using the formula:
$ \text { Induction rate }({\text{%}})=\dfrac{\text { Number of explants with hairy roots }}{\text { Total number of inoculated explants }} \times 100 {\text{%}} $ Optimization of subculture duration
-
Leaves excised from CX5 shoots at 30, 40, and 50 d after the last subculture were inoculated with A. rhizogenes. Following co-cultivation on MS + 100 μM acetosyringone, explants were transferred to selection medium. Each treatment (20 explants per replicate, three biological replicates) was assessed for hairy root induction after 30 d post-inoculation.
Regeneration and transformation efficiency analysis
-
Hairy root segments (1, 2 cm in length) were excised and cultured on shoot regeneration medium supplemented with various combinations of plant growth regulators. A factorial experimental design comprising twelve treatments was employed, combining TDZ (0.5, 1.0, or 1.5 mg·L−1), 6-BA (1, 2, or 3 mg·L−1), and IAA (0.1 or 0.2 mg·L−1). Each treatment consisted of 30 root segments, with three independent replicates per treatment. Cultures were maintained at 23 ± 2 °C under a 14 h photoperiod with a light intensity of 2,000 lx. Callus formation and shoot regeneration were assessed at 15-d intervals. Transformation efficiency was evaluated based on the following metrics:
$ \begin{split}&\rm Callus\;formation\;rate\;({\text{%}})=\dfrac{Number\; of\;roots\;forming\;callus}{Total\;number\;of\;root\;segments}\times 100{\text{%}} \\ &\rm Shoot\;regeneration\;rate\;({\text{%}})=\dfrac{Number\;of\;explants\;with\;shoots}{Total\;number\;of\;explants}\times 100{\text{%}}\\ &\rm Transgene\;positivity\;rate\;({\text{%}})=\dfrac{Number\;of\;PCR{\text-}Positive\;shoots}{Total\;number\;of\;regenerated\;shoots}\\&\quad\rm\times 100{\text{%}}\end{split} $ Identification and statistical analysis of hairy roots and transgenic roots
Fluorescent screening
-
Transgenic hairy roots expressing GFP were identified using a portable fluorescence excitation system (LUYOR-3415, USA) and documented with a photograph.
DNA analysis
-
Genomic DNA was isolated from hairy root tissues using the DNeasy Plant Kit (Qiagen, Germany). PCR amplification was performed to detect the presence of the Cauliflower mosaic virus 35S promoter and MdAIL5 gene (primer sequences provided in Table 1). The thermocycling profile consisted of an initial denaturation at 98 °C for 3 min, followed by 36 cycles of denaturation at 98 °C for 10 s, annealing at 58 °C for 15 s, and extension at 72 °C for 5 min, with a final extension at 72 °C for 10 min (details in Table 2). The PCR products were electrophoresed on 1.2% (w/v) agarose gels and visualized under ultraviolet (UV) light.
Table 2. PCR reaction system.
Reagents Volumetric (μL) RTap 10 DNA 2 Forward primer 1 Reverse primer 1 ddH2O 6 RNA analysis
-
Total RNA was extracted using the Universal Plant RNA Kit (Hua Yue Yang, China). Complementary DNA (cDNA) was synthesized from total RNA using the PrimeScript™ RT Master Mix (Takara, Japan) under the following conditions: 37 °C for 15 min and 85 °C for 5 s (reverse transcription parameters detailed in Table 3). qRT-PCR was conducted using a CFX96™ Real-Time PCR Detection System (Bio-Rad, USA). The qRT-PCR cycling protocol included an initial denaturation at 95 °C for 3 min, followed by 39 cycles of denaturation at 95 °C for 5 s, and combined annealing/extension at 58 °C for 30 s (specific cycling parameters listed in Table 4). Each sample was analyzed in technical triplicate, and relative gene expression levels were calculated using the 2−ΔΔCᴛ method, with MdActin (gene ID: MD12G1140800) serving as the reference gene.
Table 3. Reverse transcription system.
Reagents Volumetric 5X PrimeScript RT Master Mix 2 μL Total RNA 500 mg of RNA volume RNase free water Make up to 10 μL Table 4. qRT-PCR reaction system.
Reagents Volumetric (μL) Forward primer 0.4 Reverse primer 0.4 TB Green 10.0 cDNA 1.0 ddH2O 8.2 Total RNA was isolated using the Universal Polysaccharide and Polyphenol Universal Plant RNA Extraction Kit (Beijing Hua Yue Yang Biotechnology Co., China). Complementary DNA (cDNA) synthesis was performed via reverse transcription using the PrimeScript™ RT Master Mix (Perfect Real Time) kit (Takara, Japan), according to the manufacturer's protocol (Table 3). The reaction was incubated in a thermal cycler under the following conditions: 37 °C for 15 min, followed by enzyme inactivation at 85 °C for 5 s.
$\begin{split}&\rm Induction\; rate\; of\; hairy \;roots\; ({\text{%}}) =\\&\quad\rm\dfrac{Number\; of\; explants\; producing\; hairy\; roots}{Total\; number \;of \;explants} \times 100{\text{%}}\end{split} $ $\rm Positive\; conversion\; rate\; ({\text{%}}) =\dfrac{Number\; of\; positive\; hairy\; roots}{Total\; number\; of\; explants}\times 100{\text{%}} $ Rooting treatment of rootstocks and efficiency evaluation
-
Tissue culture plantlets of apple that had been subcultured for approximately 40 d were transferred to rooting medium (1/2 MS medium supplemented with 0.6 mg·L−1 IBA). Each treatment consisted of ten culture bottles, with three plantlets per bottle. After 40 d, the rooting rate and the number of roots per plantlet were recorded. The statistical formulas are as follows:
$\rm Rooting\; rate\; ({\text{%}}) =\dfrac{Number \;of\; rooted\; plantlets }{Total\; number\; of \;plantlets} \times 100{\text{%}} $ $\begin{split}&\rm Average\; number \;of \;roots\; per\; rooted\; plantlet =\\&\quad\rm\dfrac{ Total\; number \;of \;roots}{Number \;of \;rooted \;plantlets}\end{split} $ $\begin{split}&\rm Adventitious\; root \;proliferation \;coefficient =\\&\quad\rm \dfrac{Total\; number \;of\; adventitious\; roots}{Total \;number\; of\; inoculated\; explants}\end{split} $ Statistical analysis
-
Data were analyzed using Origin 2024 software. Significant differences among multiple groups were assessed via One-way analysis of variance (ANOVA) followed by Tukey's post-hoc test. Pairwise comparisons were evaluated using Student's t-test. Statistical significance was defined as p < 0.05. Data are presented as mean ± standard error (SE) from three independent replicates. Significant differences relative to the control group are indicated by lower-case superscript letters or asterisks (p < 0.05) in figures/tables.
-
In this study, CX5 leaves, apical buds, and stem segments were employed as explants for hairy root induction. The results demonstrated distinct capabilities for hairy root formation among the explant types. Specifically, apical buds and stem segments exhibited significantly lower induction efficiencies compared to leaves. The maximum induction rate for leaves reached 36.67%, whereas rates for apical buds and stem segments were 8.33% and 5.00%, respectively (Table 5 and Fig. 1). Upon close observation, high quantity and quality of hairy roots were induced by leaves. It was noted that only a small number of hairy roots emerged from the apical buds, and these roots were characterized by shorter lengths and thicker diameters. Additionally, a tendency to form callus tissue was observed in apical bud explants (Fig. 1). Stem segments, on the other hand, were even less conducive to hairy root induction than apical buds and were more susceptible to browning and death. Therefore, considering the induction efficiency and the quality of hairy roots formed, leaves were identified as the most appropriate explants for inducing hairy root formation in CX5.
Table 5. The effect of infection time of explant types on the induction rate of hairy roots.
Explant Time of infection Total number of explants Induction
rate (%)Transformation efficiency (%) Leaf 1 60 6.67 ± 1.67 cd 3.33 ± 1.67 b 5 60 36.67 ± 1.67 a 21.67 ± 4.41 a 10 60 16.67 ± 1.67 b 8.33 ± 1.67 b 15 60 15.00 ± 2.89 bc 6.67 ± 3.33 b 20 60 3.33 ± 1.67 d 1.67 ± 1.67 b Apical bud 1 60 1.67 ± 1.67 d 0 5 60 5.00 ± 2.89 d 3.33 ± 1.67 b 10 60 8.33 ± 1.67 bcd 6.67 ± 1.67b 15 60 1.67 ± 1.67 d 1.67 ± 1.67 b 20 60 0 0 Stem segment 1 60 0 0 5 60 5.00 ± 0 d 3.33 ± 1.67 b 10 60 1.67 ± 1.67 d 1.67 ± 1.67 b 15 60 1.67 ± 1.67 d 0 20 60 0 0 Data in the same column represented by different letters are very significantly different (p < 0.05, one-way ANOVA test). 
Figure 1.
Morphological effect of explant types and Agrobacterium infection duration on hairy root induction efficiency in CX5. Scale bars, 1 cm.
Effects of infection duration on the induction rate of hairy roots
-
The induction of hairy roots in apple rootstock CX5 is significantly influenced by infection duration. The induction rate initially increases and then decreases as infection duration extends. Excessive infection periods may lead to explant browning or death (Figs. 1 and 2). When leaves were used as explants, hairy roots were successfully induced across all infection duration treatments (1, 5, 10, 15, and 20 min). The highest induction efficiency of 36.67% was achieved at 5 min of infection, which was significantly higher than the other treatments. For apical bud explants, the optimal induction rate of 8.33% occurred at 10 min of infection. In the case of stem segment explants, no hairy roots were observed at infection durations of 1 or 20 min, and even at 10 and 15 min, the induction rates remained extremely low. Only at 5 min of infection did the induction rate reach 5.00%, although this was still significantly lower than that of leaf explants (Fig. 2). The aforementioned results once again confirm that leaves serve as the optimal explants for hairy root induction in CX5, with the ideal Agrobacterium infection duration being 5 min.
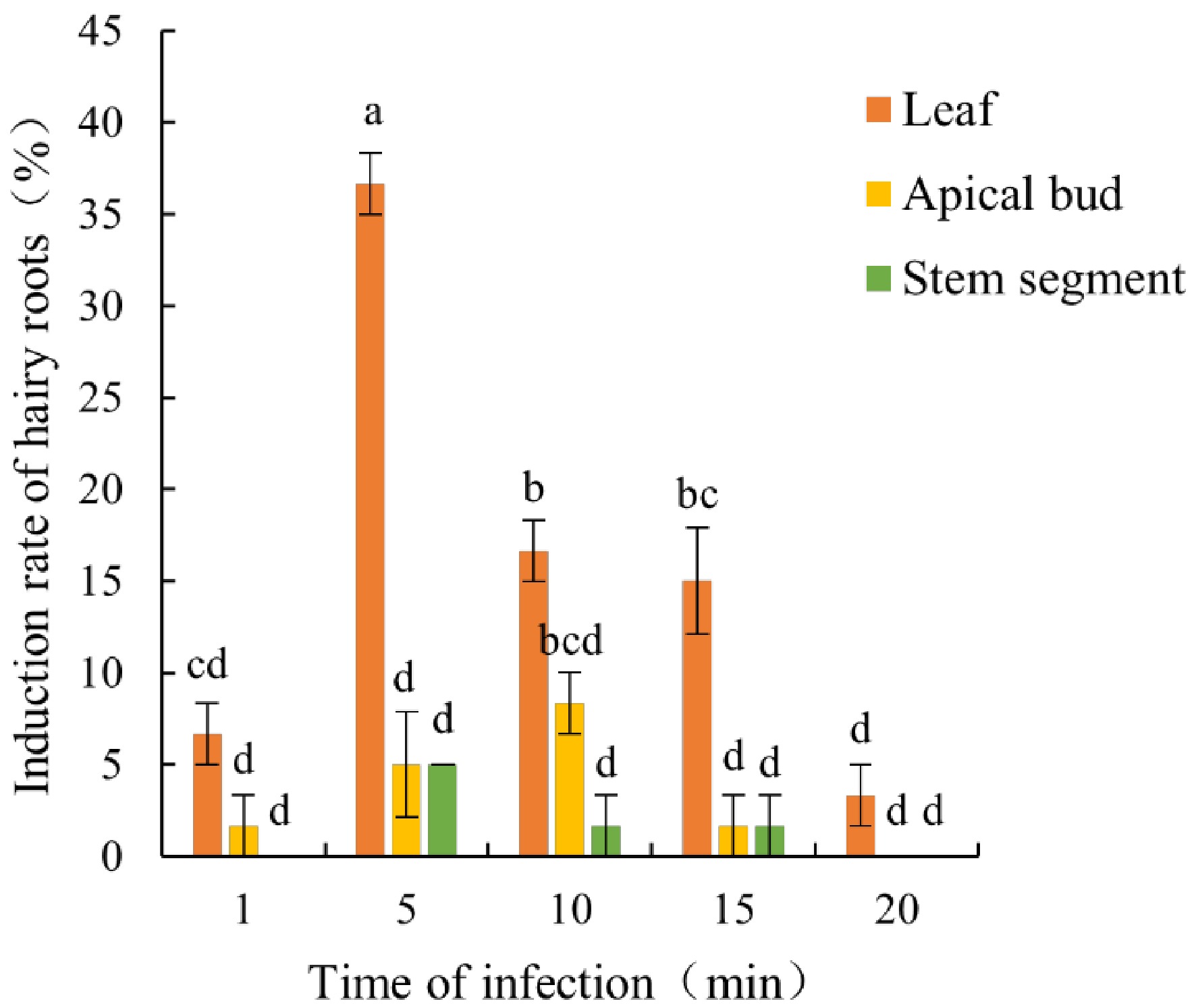
Figure 2.
Effects of explant types and Agrobacterium infection duration on hairy root induction efficiency in CX5 infection duration. Bars represent the mean value ± standard error (SE) (n = 3). Lowercase letters indicate significant differences among treatments (p < 0.05, one-way ANOVA test). Color codes denote explant types: leaf (orange), apical bud (yellow), and stem segment (green).
Effect of subculture duration on hairy root induction in CX5 genetic transformation
-
CX5 leaves at different subculture durations were used as explants for genetic transformation. The induction rate initially increased and then decreased with increasing subculture duration. At 40 d after the last subculture, the induction rate was significantly higher than for other ages, reaching 38.33% (Table 6). The hairy roots were long, and some developed small lateral roots. Therefore, leaves from shoots subcultured for 40 d are recommended as explants for inducing hairy roots.
Table 6. Effects of subculture duration on hairy root induction in CX5.
Subculture duration (d) Total number of explants Induction rate
(%)Transformation efficiency (%) 30 60 16.67 ± 4.41 b 8.33 ± 1.67 b 40 60 38.33 ± 1.67 a 18.33 ± 1.67 a 50 60 21.67 ± 4.41 b 11.67 ± 1.67 b Different lowercase letters indicate significant differences (ANOVA Duncan test, p < 0.05). Optimization of A. rhizogenes strain and bacterial solution concentration for hairy root induction in CX5
-
The induction rate of hairy roots in CX5 leaves varies significantly with different types of A. rhizogenes. As shown in Table 7, K599 achieved the highest induction rate of 36.67%, which was substantially higher than the maximum rates of 11.67% for C58C1 and 8.33% for MSU440. After inoculation with these three strains, the leaves exhibited distinct responses. K599-infected leaves were more prone to developing red spots at wound sites, followed by hairy root formation. In contrast, C58C1 and MSU440 were more likely to induce green callus tissue and less likely to form hairy roots, with prolonged storage even leading to leaf yellowing (Fig. 3). Overall, the induction and transformation rates of the three strains followed the order K599 > C58C1 > MSU440. Within the same strain, the induction rate was also greatly influenced by bacterial solution concentration. As the concentration increased, the induction rate first rose and then fell. Low concentrations resulted in poor induction or none at all, while high concentrations inhibited plant growth and reduced the induction rate. The optimal induction rate for all three strains was achieved when the bacterial solution OD600 was between 0.6 and 0.8. Based on these findings, A. rhizogenes K599 was selected as the preferred strain, with the optimal bacterial solution concentration range being OD600 = 0.6–0.8.
Table 7. Effects of A. rhizogenes strain types and bacterial concentration on hairy roots induction efficiency in CX5.
Strain type OD600 Total number of explants Induction rate
(%)Transformation efficiency (%) K599 0−0.2 60 6.67 ± 1.67 cd 3.33 ± 1.67 c 0.2−0.4 60 8.33 ± 1.67 cd 5.00 ± 2.89 c 0.4−0.6 60 23.33 ± 4.41 ab 16.67 ± 3.33 ab 0.6−0.8 60 36.67 ± 1.67 a 25.00 ± 2.89 a 0.8−1.0 60 16.67 ± 4.41 bc 8.33 ± 3.33 bc C58C1 0−0.2 60 0 0 0.2−0.4 60 1.67 ± 1.67 d 1.67 ± 1.67 c 0.4−0.6 60 3.33 ± 1.67 cd 1.67 ± 1.67 c 0.6−0.8 60 11.67 ± 1.67 bcd 6.67 ± 1.67 bc 0.8−1.0 60 1.67 ± 1.67 d 1.67 ± 1.67 c MSU440 0−0.2 60 0 0 0.2−0.4 60 1.67 ± 1.67 d 1.67 ± 1.67 c 0.4−0.6 60 5.00 ± 2.89 cd 3.33 ± 1.67 c 0.6−0.8 60 8.33 ± 1.67 cd 5.00 ±0 c 0.8−1.0 60 1.67 ± 1.67 d 1.67 ± 1.67 c Data in the same column represented by different letters are very significantly different (p < 0.05, one-way ANOVA test). 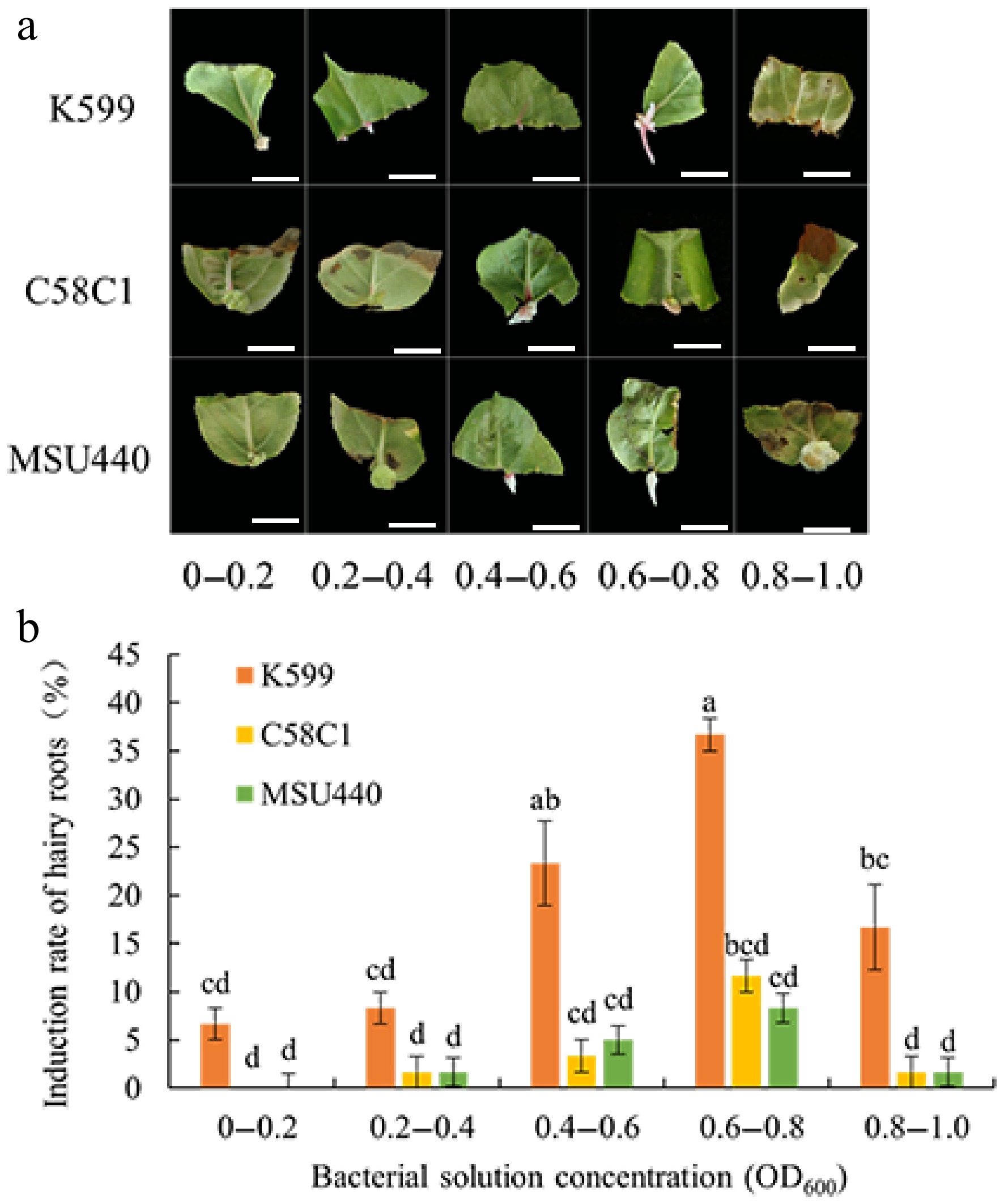
Figure 3.
Effects of A. rhizogenes strain types and bacterial concentrations on hairy root induction in leaves of CX5. (a) Morphological effects of infection concentrations mediated by strains K599, C58C1, and MSU440 on hairy root induction in CX5 leaves. (b) Effects of strain types and bacterial concentrations on hairy root induction efficiency in CX5 Bars represent the mean value ± standard error (SE) (n = 3). Lowercase letters indicate significant differences among treatments (p < 0.05, one-way ANOVA test). Color codes denote explant types: leaf (orange), apical bud (yellow), and stem segment (green). Scale bars, 1 cm.
Identification of CX5 transgenic hairy roots
-
Genetic transformation of CX5 was performed using 40-day-old CX5 leaf explants, A. rhizogenes K599, and a bacterial solution concentration of OD600 = 0.6–0.8. Under these conditions, the hairy root induction rate was 42.22% (Table 8). Fluorescent screening showed green fluorescent protein signals in some hairy roots (Fig. 4a). DNA extraction and PCR amplification confirmed the successful transfer of the MdAIL5-GFP gene into CX5 hairy roots, with a positive transformation rate of 28.89% (Table 8). Real-time fluorescence quantitative PCR revealed significant upregulation of MdAIL5 expression in several transgenic hairy roots compared to the wild-type, particularly in L2, L3, L4, L8, and L9, confirming the successful transfer of the MdAIL5 gene (Fig. 4).
Table 8. Hairy root induction efficiency in CX5 under optimized transformation conditions.
Total number of explants Induction
rate (%)Average (%) Transformation efficiency (%) Average (%) 30 36.67 42.22 ± 2.94 23.33 28.89 ± 2.94 30 46.67 30.00 30 43.33 33.33 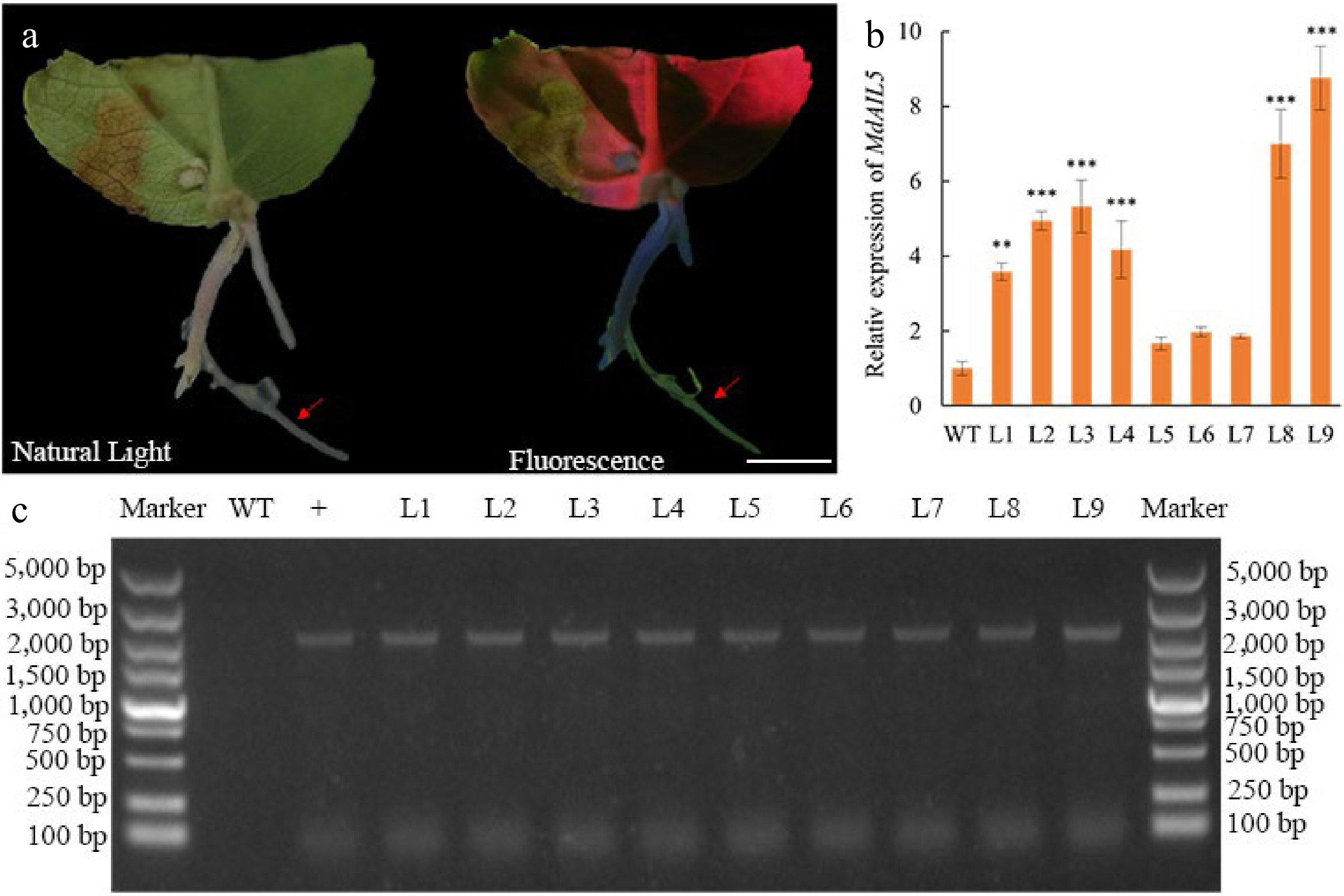
Figure 4.
Identification of MdAIL5 transgenic hairy roots. (a) Fluorescence detection of transgenic hairy roots. (b) Relative expression levels of MdAIL5 transcripts in hairy roots of WT and L1−9 lines based on qRT-PCR. For each sample, transcript levels were normalized with those of MdActin. Relative expression levels for each gene were obtained via the 2−ΔΔCᴛ method. The value of WT-Vector was set to '1'. (c) Identification of MdAIL5 transgenic hairy roots by PCR. WT, wild-type; +, positive control; arrows indicate transgenic hairy roots; L1−L9, transgenic hairy roots; Asterisks indicate significantly different values (*p < 0.05, ** p < 0.01, and *** p < 0.001; two-tailed t-tests). Scale bars, 1 cm.
The effect of different hormone ratios on the differentiation of hairy roots into buds
-
To obtain transgenic plants, hairy roots need to be induced to differentiate into shoots. To determine the optimal induction medium, twelve treatments with different TDZ, 6-BA, and IAA concentrations were set up. After 10 d on the induction medium, the root segments began to form callus. Most hairy roots failed to further differentiate into adventitious shoots after callus formation, with only a small portion of light green callus tissues eventually differentiating into shoots, which took a longer time (Fig. 5). When TDZ was used as the cytokinin, the callus induction rate initially increased and then decreased with increasing TDZ concentration. The highest callus induction rate was observed at a TDZ/IAA ratio of 10. The optimal concentrations for callus induction, germination rate, and positive rate were found to be 1.0 mg·L−1 TDZ and 0.1 mg·L−1 IAA, with rates of 85.56%, 4.44%, and 3.33%, respectively. One of the adventitious shoots was not a positive plant, suggesting that the hairy root may be chimeric. Most adventitious shoots differentiated from the root segments (Fig. 5). When TDZ concentration was 1.5 mg·L−1, and IAA was 0.2 mg·L−1, the adventitious shoots emerging from the callus were weak. With 6-BA as the cytokinin, the callus percentage initially increased and then decreased as 6-BA concentration rose to 88.89%. The average callus induction rate was higher than with TDZ, but the germination rate was lower. At a 6-BA concentration of 2.0 mg·L−1 and an IAA concentration of 0.2 mg·L−1, adventitious shoots grew from the root segments, accompanied by the appearance of white callus tissues on some root segments. In conclusion, 1.0 mg·L−1 TDZ and 0.1 mg·L−1 IAA were found to be optimal for inducing CX5 hairy roots to differentiate into shoots.

Figure 5.
The effect of different plant hormone ratios on the induction of adventitious buds from hairy roots. The hormone ratios from one to 12 are consistent with Table 9. Scale bars, 1 cm.
Characterization of CX5 transgenic lines
-
After the adventitious buds developed into transgenic lines, screening was conducted using the LUYOR-3415 Portable Fluorescent Protein Excitation Light Source. Green fluorescence was observed in the transgenic lines (Fig. 6a). Agarose gel electrophoresis and qRT-PCR were also performed. As shown in Fig. 6, the fragment size of the amplified product from the leaves matched the expected size, confirming the successful transfer of the MdAIL5 gene into CX5. MdAIL5 expression was significantly upregulated in all transgenic lines except line #4, indicating the successful integration of the MdAIL5 gene.
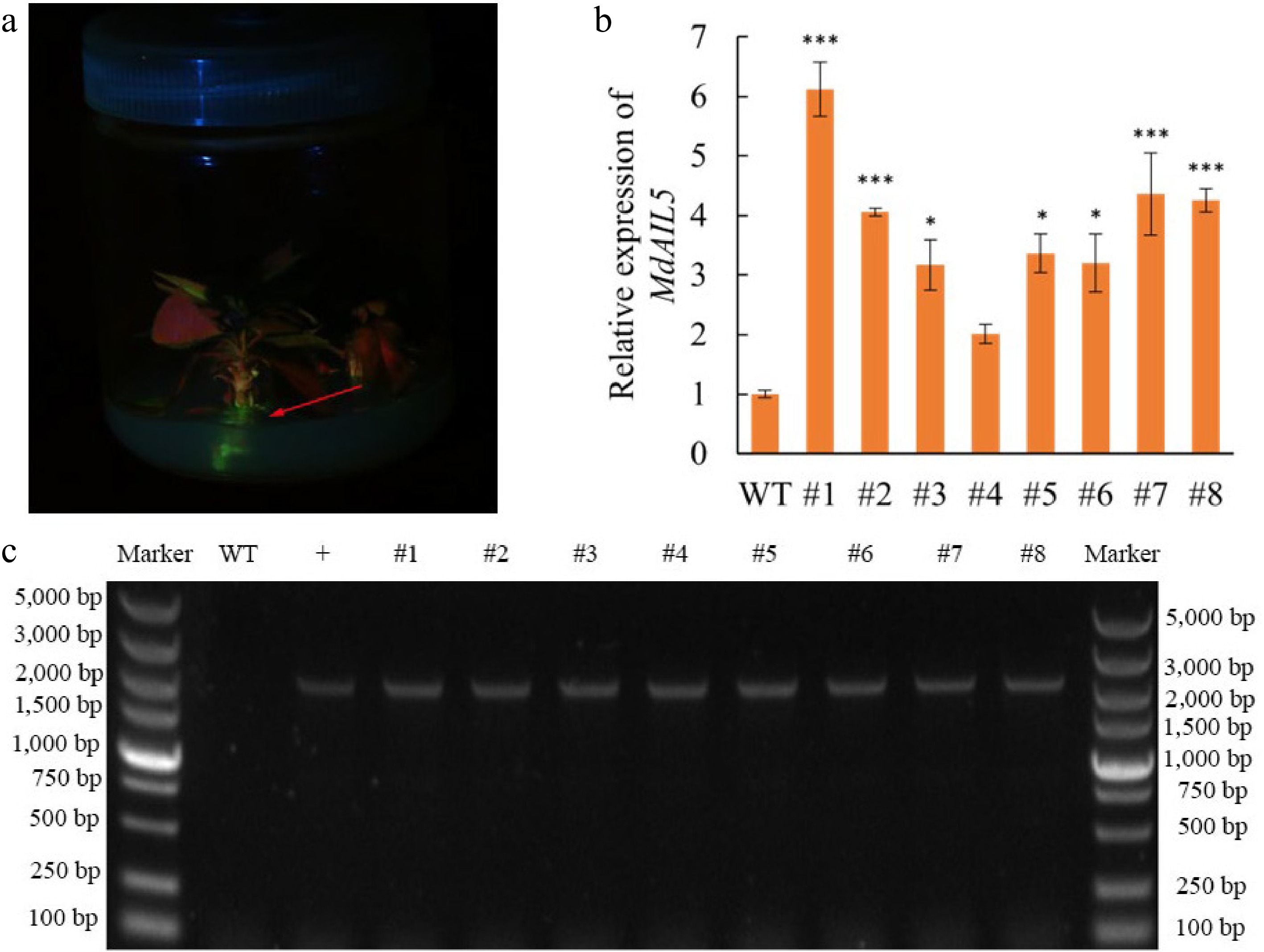
Figure 6.
Identification of transgenic buds. (a) Fluorescence detection of transgenic lines. (b) Relative expression levels of MdAIL5 transcripts in leaves of WT and #1–8 lines based on qRT-PCR. For each sample, transcript levels were normalized with those of MdActin. Relative expression levels for each gene were obtained via the 2−ΔΔCᴛ method. The value of WT-Vector was set to '1'. Asterisks indicate significantly different values (*** p < 0.001, ** p < 0.01, * p < 0.05; two-tailed t-tests). (c) Identification of MdAIL5 transgenic leaves by PCR (WT, wild-type; +, positive control; #1–8 are transgenic lines).
Effect of MdAIL5 overexpression on adventitious root formation in CX5
-
To investigate the effect of MdAIL5 overexpression on adventitious root formation in CX5, the wild-type (WT) and transgenic lines #1, #4, and #7 were subjected to root induction culture. After 40 d of culture, the rooting rate and other parameters were measured. As shown in Fig. 7, overexpression of MdAIL5 promoted adventitious root formation. Statistical analysis revealed that the rooting rates of transgenic lines #1, #4, and #7 differed significantly from that of the WT. The number of adventitious roots in the transgenic lines was at least 1.71-, 1.56-, and 1.56-fold higher than that of the WT, respectively, while the adventitious root proliferation coefficient was at least 2.70-, 2.23-, and 2.36-fold higher, respectively, showing significant differences. These results indicate that overexpression of MdAIL5 enhances the rooting capacity of CX5.
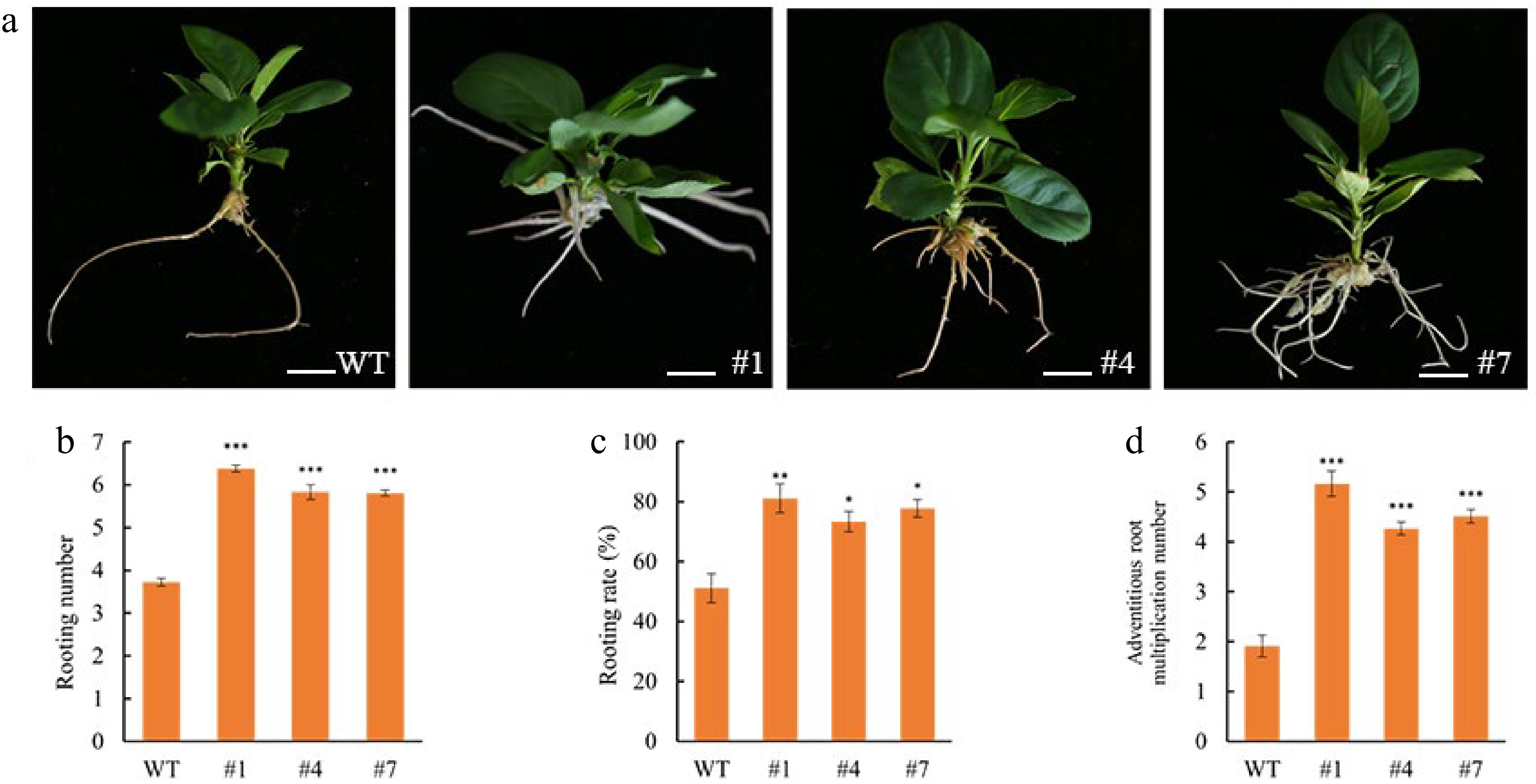
Figure 7.
Effect of MdAIL5 on Rooting of CX5 wild type and transgenic lines #1, #4, #7. (a) Overexpression of MdAIL5 and the rooting of WT apple plants were investigated. (b)–(d) the overexpression of MdAIL5 and the statistics of WT rooting number, rooting rate, and adventitious root proliferation number. The value in the figure represents the standard error of the mean value (n = 3), and the asterisk indicates that the difference is statistically significant. (*** p < 0.001, ** p < 0.01, * p < 0.05 t-test). Scale bars, 1 cm.
-
In the apple genetic transformation system, conventional methods predominantly rely on the leaf disc method, in which the Ti plasmid carried by Agrobacterium tumefaciens facilitates T-DNA integration into the plant genome, and the balance between cytokinins and auxins is utilized to directly induce adventitious shoot organogenesis. However, this approach often faces challenges such as low regeneration efficiency and strong genotype dependence in apple and other woody plants. A. rhizogenes-mediated hairy root genetic transformation offers advantages including high transformation efficiency, simple operation, and stable inheritance of transgenes[4,33]. It enables functional verification of target genes and is particularly valuable for plant species that are recalcitrant to transformation. In establishing such a transformation system, explant type, A. rhizogenes strain, infection duration, and bacterial solution concentration are key factors affecting hairy root induction and transformation efficiency[25].
Hairy root induction rates vary across plant species and explant types. For instance, using whole plants, rootless seedlings, and petioles as explants for Idesia polycarpa yielded induction rates of 71.91%, 71.5%, and 68.25%[34]. In contrast, for Ipomoea batatas, stem segments had a higher induction rate, while leaves and petioles hardly induced hairy roots[25]. Thus, selecting suitable explants for CX5 is crucial. Accordingly, our results indicated that CX5 leaves proved to be the most effective explants for inducing hairy roots compared to shoot tips and stem segments (Table 5 and Fig. 1). A few hairy roots could be induced both in shoot tips and stem segments; however, stem segments were prone to browning or death. Shoot tips induced short, thick hairy roots with a tendency to form callus tissue and a slower induction time than leaves (Fig. 1). Stem segments induced short hairy roots and grew new white leaves on leaf buds, possibly due to active stem cells at leaf growth sites, which may hinder hairy root differentiation and increase induction time. Yu et al. reported that leaves were the most suitable explants for inducing hairy roots in Malus xiaojinensis using A. rhizogenes[35]. The difference lies in the fact that Yu et al. did not use stem segments to induce hairy roots but achieved higher efficiency with apical buds and leaves[35] (Liu et al. successfully established a Gala genetic transformation system mediated by A. rhizogenes using leaves as explants, with a hairy root induction efficiency of 78.8%[5]. However, Wu et al. used stem segments in Agrobacterium-mediated genetic transformation of Malus baccata, achieving an 88.89% hairy root induction rate, which might be attributed to genotypic specificity[36].
Hairy root induction rates also vary with infection duration. Our study found that using leaves as explants with a 5-min infection period resulted in the highest induction rate of 36.67% (Table 5). An excessively short infection duration led to low induction efficiency, whereas an overly long one resulted in ineffective suppression of Agrobacterium, increased leaf mortality, and a decreased induction rate. The optimal infection durations differed for various explants, with the highest efficiency of 8.33% for shoot tips at 10 min and 5% for stem segments at 5 min (Table 5). Neither shoot tips nor stem segments yielded hairy roots after 20 min of infection, possibly due to prolonged infection duration and contact with bacterial solution, which inhibits explant growth and increases mortality. This aligns with previous research. Yu et al. reported that using leaves as explants to infect hairy roots for 10 min yielded the highest induction rate, while induction rates decreased when the time was too short or too long[35]. Pang et al observed that the induction rate of hairy roots in the basal part of Gerbera jamesonii Bolus leaves was obtained after 20 min of infection[37].
The infectivity of different A. rhizogenes strains vary. Our study of three A. rhizogenes strains infecting CX5 apple rootstock leaves showed that the hairy root induction rate followed the order of K599 > C58C1 > MSU440 (Table 6). Consistent with this, a study in Ipomoea batatas reported a similar gradient in the hairy root induction rate within its genetic transformation system, namely K599 > Ar Qual > C58C1 > MSU440 > Ar. 1193[25]. Likewise, during in vitro transformation in citrus, the efficiency of hairy root induction across different strains of A. rhizogenes was found to be A4 > R100 > 15,834[38]. Conversely, in the tomato hairy root transformation system, the order was reported as Ar Qual > MSU440 > K599[39]. In pigeon pea, the ranking followed K599 > C58C1 > MSU440 > ArA4[23]. These findings suggest that different plants may be compatible with different A. rhizogenes strains. Previous research further supports the genotype-dependent specificity; for instance, K599 was shown to be more effective in difficult-to-root apple rootstocks such as M9 and M26, inducing more robust rooting compared with MSU440. In contrast, MSU440 outperformed K599 in the relatively easy-to-root Malus xiaojinensis rootstocks[40]. This underscores that the suitability of different rhizogenic Agrobacterium strains can vary even within the same crop, depending on the genetic background.
Bacterial solution concentrations also differentially affect hairy root induction rates. Our research demonstrated that as the concentration of bacterial solution increased, the induction rate initially rose and then declined. For strains K599, C58C1, and MSU440, the induction rate of CX5 was significantly higher at OD600 values ranging from 0.6 to 0.8 compared to other concentrations (Table 7). Similarly, Wu et al. reported the highest hairy root induction rate in Phoebe bournei (Hemsl.) using a K599 bacterial solution with the OD600 of 0.8[21]. Consistent with this, Xie et al. also observed the highest induction rate when infecting Carya illinoinensis (Wangenh.) with a K599 bacterial solution with OD600 = 0.8, reaching 56.5%[41]. The optimal bacterial solution concentration OD600 for tomato hairy root transformation was reported between OD600 = 0.6-0.8[39]. In contrast, in pigeon pea, the highest induction rate was achieved at an OD600 of 0.3[23]. In M9, M26, and Malus xiaojinensis root induction, the optimum transforming bacterial solution concentration was OD600 of 0.5[40]. Collectively, these results indicate that optimal bacterial solution concentration varies considerably across different plant species, highlighting the necessity of selecting an appropriate OD600 for each specific plant system.
Subculture duration differences can also lead to varying induction rates. This study found that when using CX5 leaves as explants, the hairy root induction rate was highest at 40 d of subculture duration, reaching 38.33% (Table 6). The lowest induction rate was at 30 d of subculture duration, possibly because the leaves were too young and tender (Table 6). Additionally, growth on antibiotic-containing media was inhibited, further reducing hairy root induction rates. When seedlings are too old, leaf aging and weakened cell division ability may decrease induction rates. These results are consistent with those of Liu et al.[5], who found that in Gala hairy root induction, leaves from tissue-cultured in vitro shoots after three weeks of propagation were better than those after four weeks, and young leaves had higher transformation rates than old leaves[5]. Unlike CX5, the best subculture duration for Gala was three weeks, which is a shorter duration than CX5's 40 d. This may be related to the apple genotype, as CX5 rootstock grows more slowly. In the hairy root induction of Pisum sativum L 'Zhe Wan 1', seven-day-old seedlings showed the highest efficiency, producing more hairy roots. Short infection times resulted in decreased hairy root numbers and weakened plants. In Phoebe bournei (Hemsl.), the induction rate was higher in seedlings at the second and third leaf stages than in those at the one-leaf stage[21]. When inducing hairy roots in Malus hupehensis via injection, the induction rate at the eight-leaf stage was higher than at the three-leaf stage[15]. Using bacterial liquid infection, however, the induction rate at the three-leaf stage was higher than at the five-and eight-leaf stages[15]. For many plants undergoing hairy root induction, the early stage is preferred because cells are dividing vigorously and are more receptive to exogenous DNA. However, using overly young plants is not advisable. After bacterial infection, explants need to grow in media with added antibiotics. If explants are too young, their growth is inhibited, making them prone to death, which leads to decreased induction rates.
Different hormone concentration ratios used in this study significantly affect adventitious bud regeneration. This study found that the regeneration of roots in apple plants remains challenging, with a low regeneration rate of only 4.44%, which aligns with findings in Malus baccata results[36]. Adventitious buds were regenerated from transgenic hairy roots, which were preliminarily identified via fluorescence. The transgenic nature of the regenerated buds was confirmed by agarose gel electrophoresis, showing the highest positive rate at 3.33% (Table 9). Only one bud in treatment 2 was non -transgenic and died later. It is hypothesized that the hairy root was chimeric and that the site of adventitious shoot regeneration had not been successfully transformed with the target gene. Overexpression of the MdAIL5 gene significantly enhanced the rooting ability of CX5 apple rootstocks (Fig. 1), suggesting that this transcription factor plays a positive regulatory role in adventitious root formation.
Table 9. Effects of different hormone ratios on the differentiation of hairy roots into buds.
Number TDZ
(mg·L−1)6-BA
(mg·L−1)IAA
(mg·L−1)Total number
of explantsCallus induction
rate (%)Germination
rate (%)Positivity
rate (%)1 0.5 0 0.1 90 73.33 ± 1.92 ab 0 0 2 1.0 0 0.1 90 85.56 ± 1.11 a 4.44 ± 1.11 a 3.33 ± 0 a 3 1.5 0 0.1 90 71.11 ± 2.22 bc 1.11 ± 1.11 ab 1.11 ± 1.11 b 4 0.5 0 0.2 90 57.78 ± 2.94 c 0 0 5 1.0 0 0.2 90 63.33 ± 3.33 bc 0 0 6 1.5 0 0.2 90 71.11 ± 4.01 bc 1.11 ± 1.11 ab 1.11 ± 1.11 b 7 0 1 0.1 90 84.44 ± 2.94 abc 0 0 8 0 2 0.1 90 87.78 ± 1.11 ab 1.11 ± 1.11 ab 1.11 ± 1.11 ab 9 0 3 0.1 90 77.78 ± 2.94 c 0 0 10 0 1 0.2 90 81.11 ± 1.11 bc 0 0 11 0 2 0.2 90 88.89 ± 2.22 a 2.22 ± 1.11 a 2.22 ± 1.11 a 12 0 3 0.2 90 82.22 ± 2.22 abc 0 0 Different lowercase letters indicate significant differences (ANOVA Duncan test, p < 0.05). -
In conclusion, this study establishes a robust Agrobacterium rhizogenes-mediated transformation system for CX5. The optimized protocol employs leaf explants from shoots subcultured for 40 d, infection with A. rhizogenes K599 (OD600 = 0.6–0.8), and regeneration on medium supplemented with 1.0 mg · L−1 TDZ and 0.1 mg · L−1 IAA. This breakthrough provides a critical platform for the functional genomics and molecular breeding of CX5, enabling targeted improvement of root architecture and stress tolerance to support sustainable apple production in cold regions. Meanwhile, overexpression of MdAIL5 improves the rooting ability of CX5 rootstock.
-
The authors confirm their contributions to the paper as follows: study conception and design: Zhang C, Han X, Tian Y; experimental work: Tian Y, Han X, Yan J, Yang A, Kang L, Ma S, Zhang C; data analysis, draft manuscript preparation: Tian Y; manuscript editing, supervision: Zhang C, Ma S. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
-
This research was supported by the China Agriculture Research System of MOF and MARA (Grant No. CARS-25), the Agricultural Science and Technology Innovation Program (Grant No. CAAS-ASTIP-2021-RIP 02), and the National Key R & D Program of China (Grant No. 2022YFD1600500).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Tian Y, Han X, Yan J, Yang A, Kang L, et al. 2026. Establishment of a genetic transformation system for apple rootstock CX5 mediated by Agrobacterium rhizogenes. Fruit Research 6: e026 doi: 10.48130/frures-0026-0019 

Establishment of a genetic transformation system for apple rootstock CX5 mediated by Agrobacterium rhizogenes
- Received: 29 December 2025
- Revised: 08 April 2026
- Accepted: 21 May 2026
- Published online: 10 July 2026
Abstract: The apple rootstock CX5 is valued for its cold tolerance and dwarfing characteristics, positioning it as a promising candidate for overcoming cold stress and modernizing cultivation systems in China's apple industry. However, its broader adoption is hindered by challenges such as poor rooting ability, which limit the efficiency of conventional breeding approaches. Biotechnological breeding can offer a solution to this bottleneck, and establishing a genetic transformation system for CX5 is a pivotal step. Based on previous findings that MdAIL5 significantly enhances adventitious root regeneration, we introduced the PRI101-MdAIL5-GFP vector into CX5 using the Agrobacterium rhizogenes strain K599-mediated system and systematically optimized key transformation parameters. Results showed that using leaves from shoots subcultured for 40 d, infected with bacterial solution at an OD600 of 0.6−0.8 for 5 min, achieved a maximum hairy root induction rate of 28.89%. Furthermore, on medium supplemented with 1.0 mg·L−1 TDZ and 0.1 mg·L−1 IAA, the regeneration rate of adventitious buds from transgenic hairy roots reached 3.33%. Overexpression of MdAIL5 enhanced the rooting ability of CX5 tissue culture plantlets. In summary, we successfully established a genetic transformation system for CX5 and demonstrated that overexpression of MdAIL5 significantly enhances rooting ability. This system provides a crucial technical foundation for genetically improving the poor rooting phenotype and other target traits of this cold-tolerant dwarf rootstock.
-
Key words:
- Apple rootstock /
- Agrobacterium rhizogenes /
- Hairy root /
- CX5 /
- Genetic transformation