








-
Colon cancer (CC) is one of the most common human malignancies worldwide and the fifth leading cause of cancer-related deaths[1]. Many factors are involved in the development and progression of CC, including the activation of oncogenes and inactivation of tumor suppressor genes. Despite advances in treatment in the last decades, tumor recurrence is common[2,3] and most patients with CC have a limited response to immune checkpoint inhibitor (ICI) therapy[4]. Emerging biomarkers such as tumor mutational load (TMB), inflammatory tumor microenvironment (TME), and microsatellite instability (MSI) have been identified to predict the therapeutic benefit of CC[5]. Therefore, the discovery of additional diagnostic and prognostic biomarkers and therapeutic targets will likely lead to better treatment strategies for CC patients and improve long-term outcomes.
Cuproptosis is a newly discovered mode of cell death that differs from all known processes regulating cell death, including apoptosis, ferroptosis, pyroptosis, and necroptosis. The main process of cuproptosis is that excess copper induces copper-induced cell death by promoting aberrant oligomerisation of copper-dependent lipoylated proteins in the tricarboxylic acid cycle as well as decreasing Fe-S cluster protein levels[6]. Dihydrolipoamide dehydrogenase (DLD) is a component of the glycine cleavage system and an E3 component of the three α-ketoacid dehydrogenase complexes (pyruvate, α-ketoglutarate, and branched-chain amino acid dehydrogenase complexes), and the DLD gene is a key regulator of copper-induced cell death[6−9]. DLD has been studied in pan-cancer, showing that high expression of DLD is associated with a better clinical prognosis in colon adenocarcinoma (COAD)[10]. However, the impact of the cuproptosis-associated gene DLD on clinicopathological factors and immune cell infiltration in CC remains largely unknown.
This study highlights the expression, prognostic and immunological significance of DLD in CC. Data from The Cancer Genome Atlas (TCGA) and Gene Expression Database (GEO) were used to assess the role of DLD in CC. Bioinformatic analysis confirmed the expression profile and prognostic value of DLD in CC. The association of DLD with somatic mutations and immune cell infiltration was assessed. Gene Ontology (GO) and Kyoto Encylopedia of Genes and Genomes (KEGG) enrichment analysis was used to analyze the potential mechanisms that differed between the high- and low-DLD groups. Finally, we determined that high expression of DLD improved the survival of CC patients, which may be associated with the regulation of DLD in different functional states of the tumor cells and the immune microenvironment.
-
In this study, we downloaded mRNA sequencing data in fragments per kilobase per million mapped fragments (FPKM) format from the TCGA database (
https://portal.gdc.cancer.gov/ , as of December 28, 2022) for normal and primary tumor samples, including 446 COAD samples and 93 normal colon samples. We also downloaded the GSE39582 dataset from Gene Expression Omnibus (GEO) (www.ncbi.nlm.nih.gov ) for validation[11]. Clinical data of COAD patients from the TCGA database and the GEO database were also obtained. Patients were classified into high and low groups according to the cut-off point of mRNA expression of DLD. A total of 1,011 COAD samples (TCGA: 446; GSE39582: 562) and 112 normal tissues (TCGA: 93; GSE39582: 19) were included in this study.DLD expression profiles in CC
-
The mRNA expression of DLD in CC was analyzed using the University of Alabama at Birmingham Cancer data analysis portal (UALCAN) (
http://ualcan.path.uab.edu ), a comprehensive, user-friendly, interactive web-based resource for analyzing cancer omics data[12,13]. The immunohistochemistry of DLD proteins in normal human colon tissue and CC tissue was obtained using the Human Protein Atlas (HPA) (www.proteinatlas.org )[14,15]. In addition, the diagnostic power of DLD for CC was calculated using receiver operating characteristic (ROC) curves.Relationship between DLD expression and CC survival prognosis
-
Overall survival (OS) data were downloaded from the TCGA database's COAD project and the GEO database GSE39582. Kaplan-Meier (K-M) survival curve analysis was implemented by the R packages "Survival" (version 3.2-10) and "Survminer" (visualization, version 0.4.9). Cox regression or log-rank tests were used to analyze the relationship between DLD expression and the survival of CC patients, and the effect of DLD expression on the prognosis of different clinical variables.
Enrichment analysis of the DLD gene's co-expression network in CC
-
First, we identified co-expressed genes associated with DLD expression in the TCGA-COAD dataset and GSE:39586 in R software. We used Pearson's correlation coefficient to test the statistical correlations, and the ggplot2 package of R software to draw heat maps for presentation. We performed GO function and KEGG pathway enrichment analysis using the DAVID website (
https://david.ncifcrf.gov/ ), and enriched gene terms with false disovery rate (FDR) q-values < 0.05 were considered to be statistically significant. The results were displayed as bar graphs using R software[16].Gene set enrichment analysis
-
To explore the potential regulatory mechanisms of DLD, we divided the samples in the TCGA-COAD dataset into two groups based on the cut-off points of DLD expression levels. The R package "DESeq2" was run to filter differentially expressed genes (DEGs) between CC samples with high and low DLD expression[17]. A fold change (FC) of > 1.2 or < 0.83 and p < 0.05 were defined as the filtering criteria. The results of the DEGs were displayed as volcano plots using the R package "ggplot2". Gene set enrichment analysis (GSEA) analysis was performed using OmicShare tools, a free online platform for data analysis (
www.omicshare.com/tools ). FDR (q-value) < 0.05 was considered to be statistically significant.Tumor Mutation Load Analysis
-
We downloaded somatic mutation and somatic copy number variation (CNA) data for COAD from the TCGA database. Mutation details of genes with the 30 highest mutation frequencies between the high- and low-DLD groups were represented as waterfall plots using the "maftools" R package.
Analysis of tumor immune infiltration
-
The Tumor Immunity Evaluation Resource (TIMER) (
http://timer.cistrome.org/ ) is a comprehensive resource for the systematic analysis of immune infiltration in different cancer types[18]. The correlation between DLD expression in CC and six types of immune cells (CD8 T cells, B cells, NK cells, CD4 T cells, macrophages, and neutrophils) was assessed using TIMER. A single-sample gene set enrichment analysis (ssGSEA) was applied to compare 28 tumor-infiltrating immune cells in CC between the high- and low-DLD groups.Single-cell sequencing data analysis
-
CancerSEA (
http://biocc.hrbmu.edu.cn/CancerSEA/home.jsp ) was used to analyze the effect of DLD expression levels on the different functional states of cancer cells at the single-cell level[19]. We downloaded the colorectal cancer expression and functional status files from the Cancer SEA database (database ID: EXP0051) and the corresponding CC single-cell sequencing files from the Gene Expression Omnibus (GEO) database (GSE81861) and visualized the correlation between DLD and the functional status of CC using the R package "ggplot2".Statistical analysis
-
Statistical analyses for this study were performed using R language (version 4.3.0), which is a common language and environment used for statistical calculations (R Foundation, Vienna, Austria,
www.R-project.org ) and GraphPad Prism software (version 8.0.2). The ROC curve of the pROC package was applied to assess the potential diagnostic value of DLD expression and the area under the curve (AUC) was obtained[20]. AUC > 0.7 showed good diagnostic accuracy. The chi-square (χ2) test was used to assess the correlation between DLD expression and the clinicopathological characteristics of the patients. The significance of differences between groups was determined by Student's t-test (unpaired, two-tailed) and one-way analysis of variance (ANOVA). The "Survminer" and "survivor" R packages were used in R language for the Kaplan-Meier analysis, and the log-rank test was used to determine the cut-off point for the difference in OS between the high- and low-DLD groups, and p < 0.05 was considered statistically significant. -
We analyzed the expression levels of DLD in CC samples from the TCGA database, the UALCAN database, and GSE39582. The results showed that DLD expression was low in CC (Fig. 1a−c). Immunohistochemical results from the HPA database showed that DLD was expressed in the mucosal epithelium, lamina propria, and submucosa of normal colon tissues (Fig. 1d). However, compared with normal colon tissue, there was no obvious expression in CC tissue (Fig. 1e). More importantly, the ROC curves showed that DLD expression had the ability to identify CC and normal colon tissues in both cohorts (Fig. 1f, g).
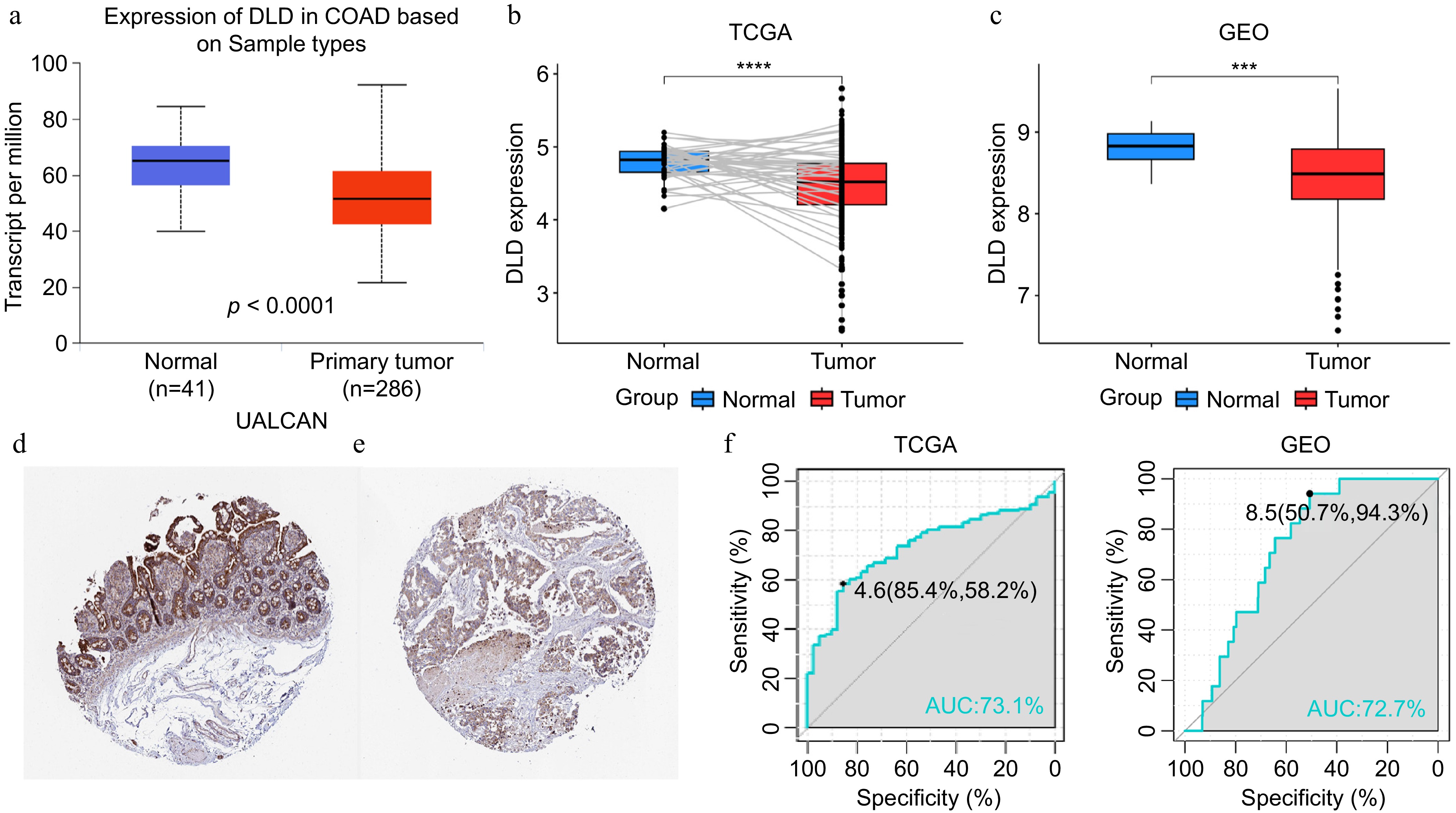
Figure 1.
DLD expression in colon cancer (CC). (a) mRNA expression of DLD in normal tissues and CC tissues, sourced from the the University of Alabama at Birmingham Cancer data analysis Portal (UALCAN) database. (b) Paired expression analysis of DLD in normal and colon adenocarcinoma (COAD) tissues sourced from The Cancer Genome Atlas (TCGA) database. (c) Expression analysis of DLD in normal and CC tissues sourced from the Gene Expression Omnibus (GEO) database. (d) Levels of DLD protein in normal colon tissues sourced from the Human Protein Atlas. (e) Levels of DLD protein in COAD tissues in the Human Protein Atlas database. (f), (g) The receiver operating characteristic (ROC) curve showed the high expression specificity of DLD in the TCGA database and GEO databases for CC. AUC, area under the curve. *** p < 0.001, **** p < 0.0001.
Relationship between DLD expression and clinical characteristics of CC patients
-
Patients with different levels of DLD expression showed different patterns of clinical and pathological features. In the TCGA dataset and the GSE39582 dataset, DLD expression levels, different clinical stages (T), lymph node invasion (N), and different pathological stages showed an asymmetric distribution (Fig. 2a, b). A comparative analysis was performed for different groups of these samples. In the TCGA database, the expression of DLD decreased with increasing lymph node invasion (N) and pathological stage level in the samples (p < 0.05; Fig. 2d, e), with no significant difference in the subgroup analysis of patients at different clinical stages (T) (Fig. 2c). The same trend was observed for DLD expression in the GSE39582 dataset, although this difference was not statistically significant (Fig. 2g, h); however, in the subgroup analysis of patients at different clinical stages (T), DLD expression decreased at higher clinical stages (T) (p < 0.05; Fig. 2f). These results suggest that DLD is lacking in CC with greater malignancy.
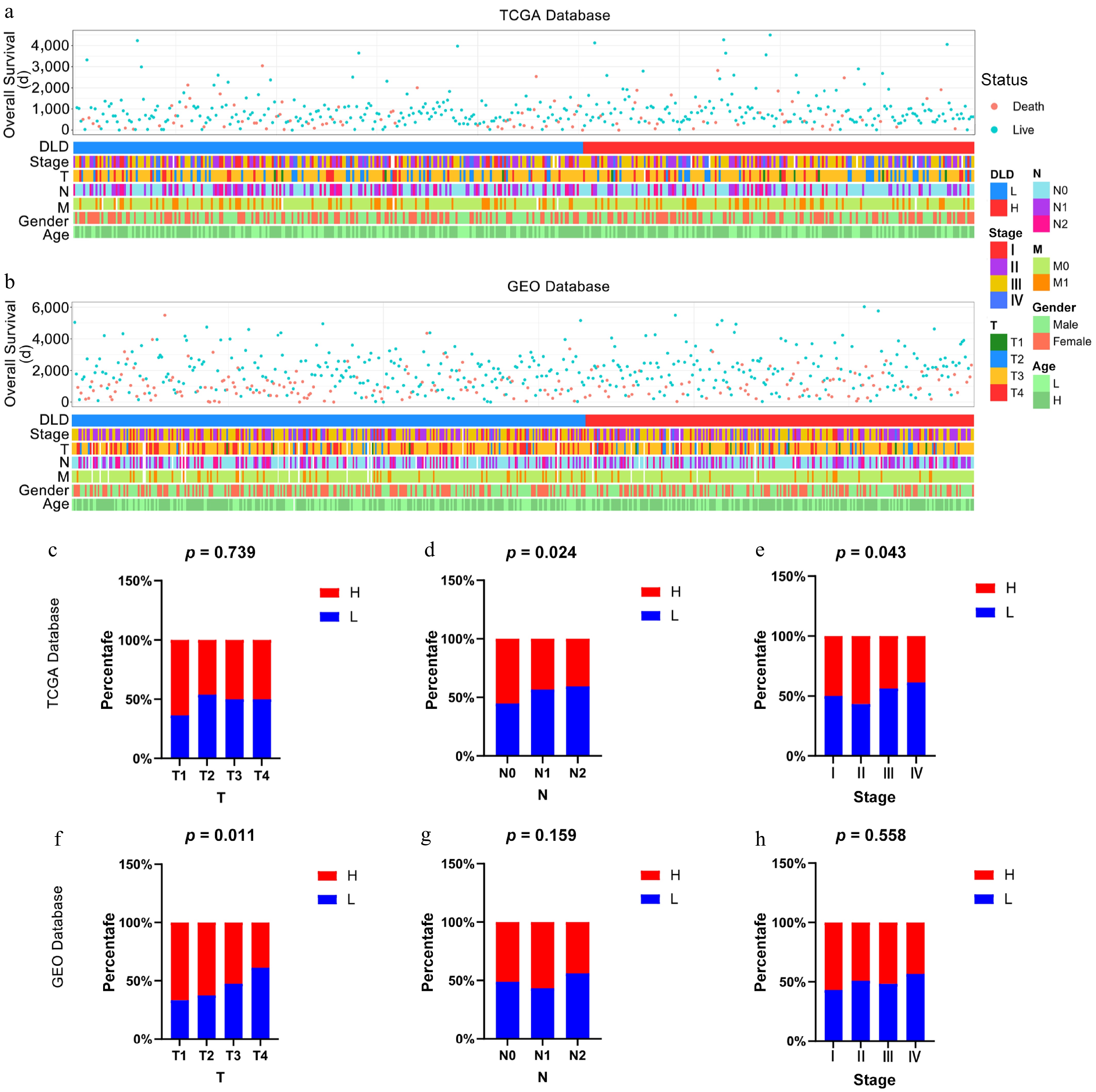
Figure 2.
Relationship between DLD and clinicopathological features of colon cancer (CC). (a) Status of clinicopathological features of CC associated with DLD in The Cancer Genome Atlas (TCGA) database. (b) Status of clinicopathological features of CC associated with DLD in the Gene Expression Omnibus (GEO) database. (c)–(h) Correlation of DLD expression with clinicopathological parameters including clinical stages, lymph node invasion, and pathological stage in CC patients in the TCGA database and GEO database. The significance of differences was tested by the χ2-test.
High DLD expression implies better prognosis for CC patients
-
To investigate the prognostic value of DLD in both cohorts, patients were divided into high- and low-DLD expression groups according to the optimal cut-off points calculated by the "survival" and "survminer" software packages. We then used Kaplan-Meier analysis to assess the prognostic significance of DLD expression in the TCGA-COAD database and the GSE39582 dataset. Kaplan-Meier survival curves were generated, based on the cut-off points for DLD expression and showed that CC patients expressing high levels of DLD showed better OS rates (p < 0.05; Fig. 3a, b).
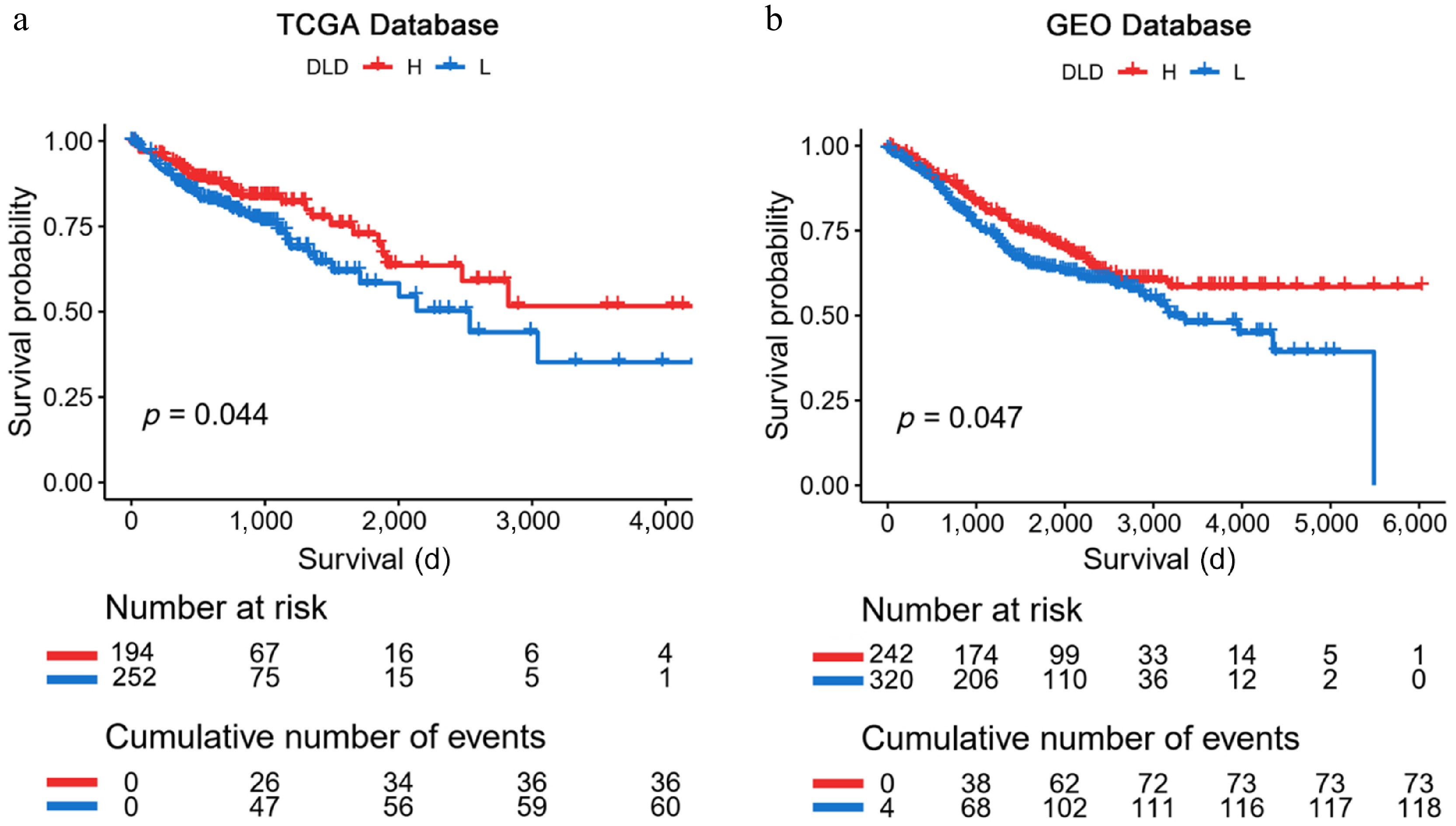
Figure 3.
Kaplan-Meier analysis of DLD expression in The Cancer Genome Atlas (TCGA) database and the Gene Expression Omnibus (GEO) database. (a), (b) Kaplan-Meier plots verified that high DLD mRNA levels are associated with better survival outcomes in patients with CC. Significance of prognostic value was tested by log-rank test.
Enrichment analysis of DLD co-expressed genes and DEGs in CC
-
To explore the biological functions associated with DLD, the 302 genes most associated with DLD were screened by Pearson correlation analysis (r > 0.5, p < 0.05) in the TCGA database. GO and KEGG analyses were performed on genes co-expressed with DLD via the R package and the DAVID website. Fifty genes were screened in the GSE:39582 dataset using the same screening method. The biological processes most associated with DLD in the TCGA database include protein transport, mitochondrial adeosine triphosphate (ATP) synthesis coupled proton transport, and mitochondrial transport (Fig. 4a). In addition, the most relevant cellular component for DLD is the mitochondrion (Fig. 4c). Molecular functions include guanosine triphosphate (GTP) binding, GTPase activity, and protein serine/threonine phosphatase activity (Fig. 4e). The most relevant signaling pathways for DLD are metabolic pathways and oxidative phosphorylation (Fig. 4g). The biological processes, cellular components, molecular functions, and signaling pathways associated with DLD in the GSE:39582 dataset were similar to those in the TCGA database (Fig. 4b, d, f, h). These findings suggest that DLD in CC cells could play an important role in the energy metabolism of the disease. The genes with the strongest associations were CYCS (r = 0.737, p = 8.93E–89), NDUFA5 (r = 0.691, p = 5.78E–74), SDHD (r = 0.654, p = 6.94E–64), and SNX4 (r = 0.653, p = 1.37E–63), where r > 0.5 and p < 0.05 are the cut-off values (Fig. 4i).
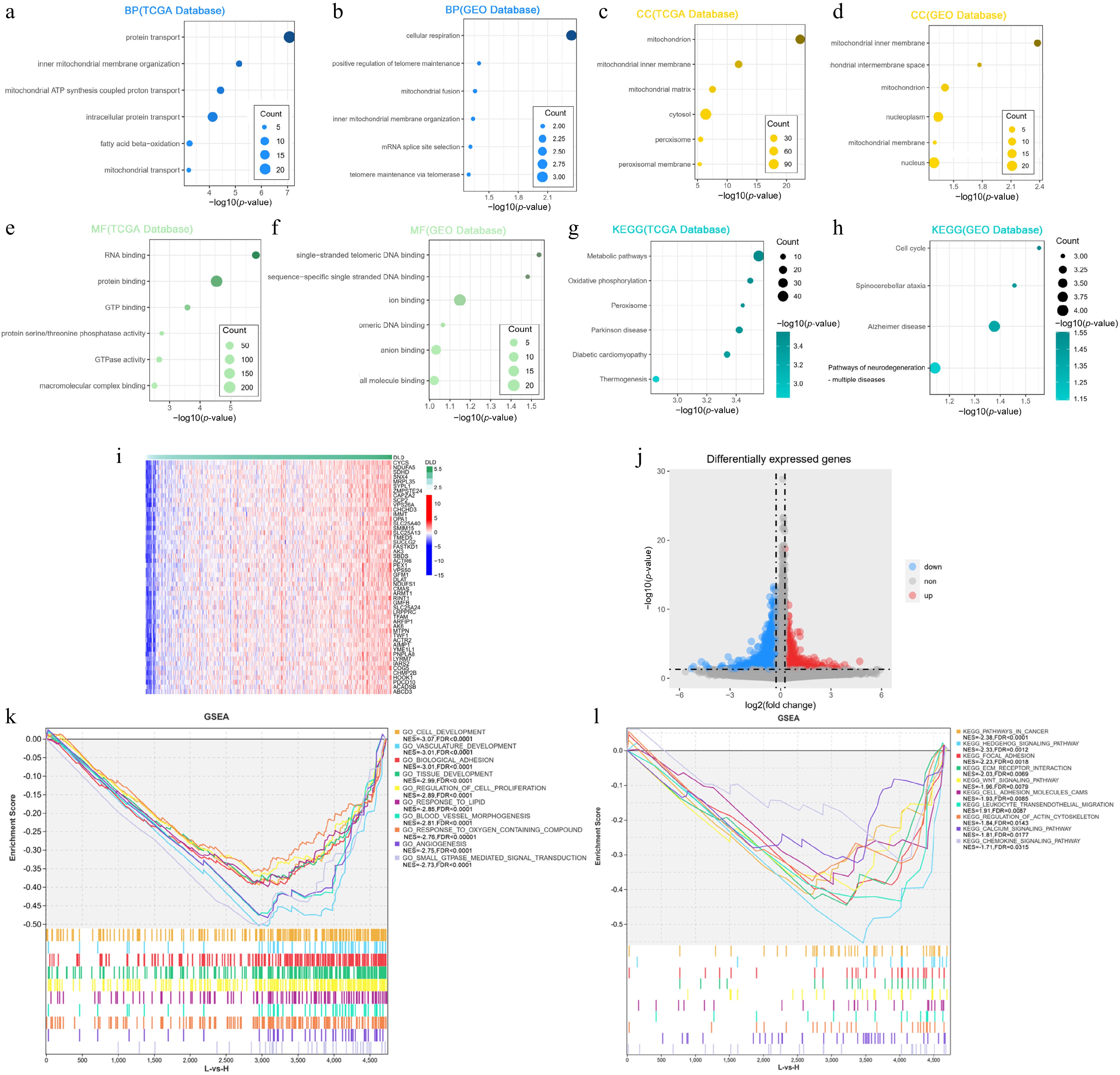
Figure 4.
DLD is closely related to the regulation of metabolic processes in CC. (a), (b) Biological processes (BP) mostly associated with DLD in The Cancer Genome Atlas (TCGA) database and the Gene Expression Omnibus (GEO) database. (c), (d) Cellular components (CC) mostly associated with DLD in the TCGA database and GEO database. (e), (f) The most relevant molecular functions (MFs) of DLD in the TCGA database and the GEO database. (g), (h) KEGG pathway analysis of DLD in the TCGA database and GEO database. (i) Heat map showing the top 50 co-expressed genes positively associated with DLD expression in the TCGA dataset. (j) Volcano plot showing differentially expressed genes in the high-DLD group versus the low-expression group in the TCGA dataset. (k), (l) Gene Ontology (GO) function and KEGG pathway enrichment analysis of differentially expressed genes in the high- versus low-DLD groups in the TCGA dataset.
We divided the samples in the TCGA COAD dataset into two groups according to the cut-off points of DLD expression levels, with 713 upregulated genes and 4,018 downregulated genes (Fig. 4j). To further investigate the potential functions of DLD in CC, we performed a GSEA based on the DLD levels in the TCGA database. A large set of genes were negatively enriched in the high-DLD expression group, including pathways of tumors (Normalized Enrichment Score (NES) = –2.38, FDR < 0.0001; Fig. 4l), Hh signaling (NES = –2.33, FDR = 0.001; Fig. 4l), wingless-type MMTV integration site family (WNT) signaling pathway (NES = –1.96, FDR = 0.008; Fig. 4l), the extracellular matrix (ECM) (NES = –2.03, FDR = 0.007; Fig. 4l), angiogenesis (NES = –2.75, FDR < 0.0001; Fig. 4k), blood vessel morphogenesis (NES = –2.8, FDR < 0.0001; Fig. 4k), cell development (NES = –3.06, FDR < 0.0001; Fig. 4k), cell proliferation (NES = –2.89, FDR < 0.0001; Fig.4k), and response to oxygen-containing compound (NES = –2.7, FDR < 0.0001; Fig. 4k).
DLD expression levels are associated with tumor mutation burden
-
To determine whether DLD expression levels correlate with specific genomic features in CC, we performed a somatic mutation analysis based on DLD expression levels using the "maftools" package in the TCGA-COAD database, which shows the top 30 mutated genes. In the high-DLD group, 66.3% (173/261) of the samples were mutated, with a high frequency of mutations in APC (38%), TP53 (32%), TNN (32%), and KRAS (30%) (Fig. 5a). In contrast, in the low-DLD group, up to 99.5% (201/202) of samples were mutated, with TTN (55%), APC (53%), TP53 (49%), KRAS (39%), and MUC16 (30%) being mutated more frequently (Fig. 5b).
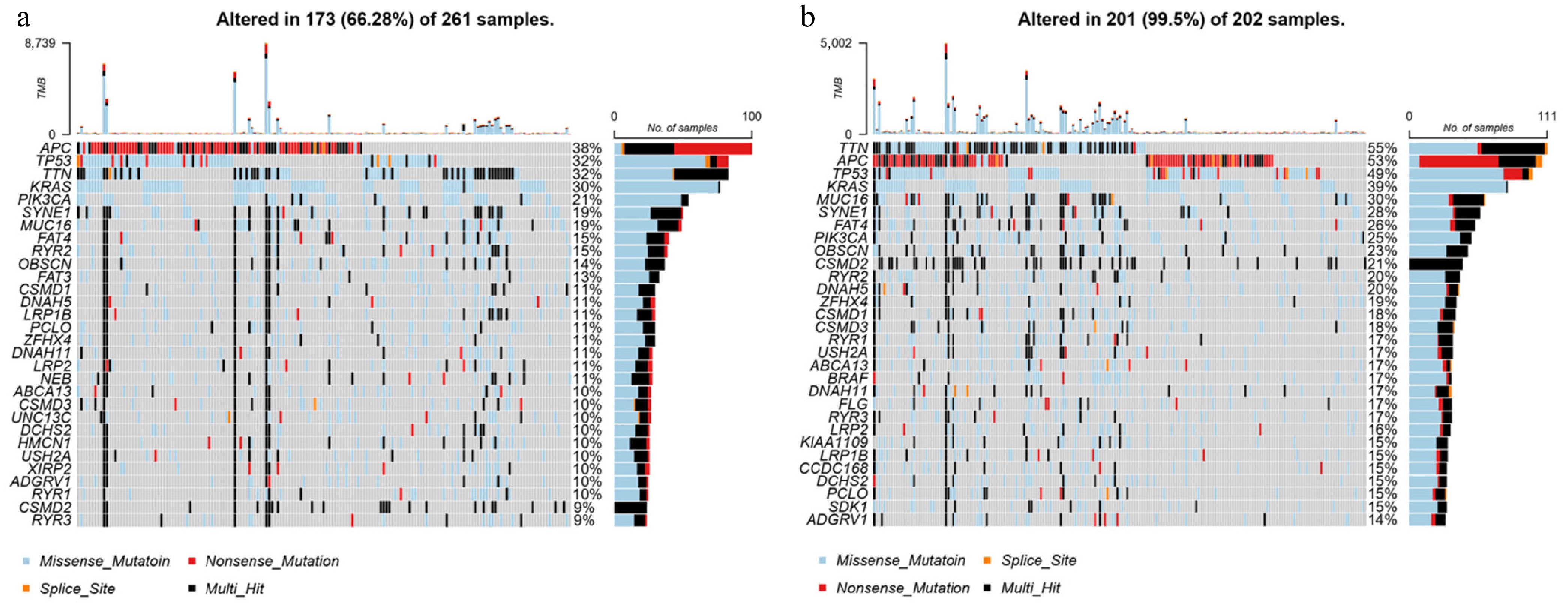
Figure 5.
Somatic mutation analysis in groups with high and low expression of DLD. Waterfall plot of the top 30 mutational genes in the (a) high-DLD group and (b) low-DLD group.
The relationship between DLD expression levels in single cells and tumors' functional status
-
We analyzed the expression of DLD in COAD (single cells) and its relationship with the tumor's functional status by using the CancerSEA database. We found that DLD expression in COAD was positively correlated with "invasion" (r = 0.39, p < 0.05, Fig. 6a) and "DNA repair" (r = 0.29, p < 0.05, Fig. 6b). In contrast, DLD expression was negatively correlated with "angiogenesis" (r = −0.28, p < 0.05, Fig. 6c).
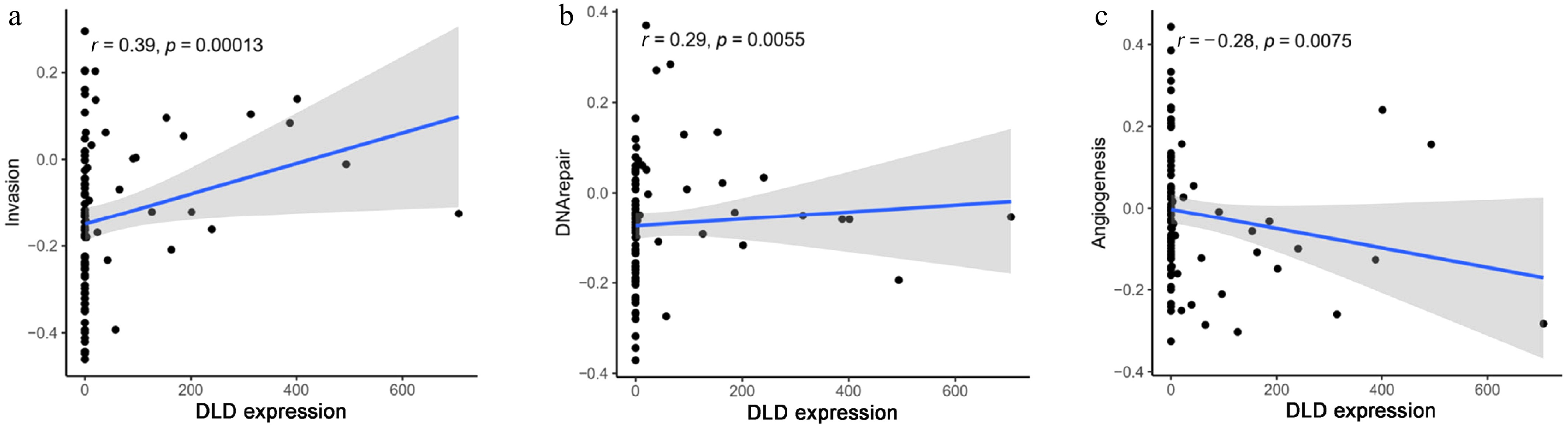
Figure 6.
Correlation between DLD expression and significantly different functional states in CC: (a) invasion, (b) DNA repair, and (c) angiogenesis.
Relationship between DLD and immune cell infiltration
-
The relationship between DLD expression and immune infiltration of CC was analyzed by the "Gene" module of the TIMER website. The results showed that DLD expression was associated with CD8 T cells (r = 0.225, p < 0.05), B cells (r = 0.214, p < 0.05), NK cells (r = 0.15, p < 0.05), CD4 T cells (r = 0.101, p < 0.05), macrophages (r = 0.193, p < 0.05), neutrophils (r = 0.124, p < 0.05), and dendritic cell infiltration levels (Fig. 7a).
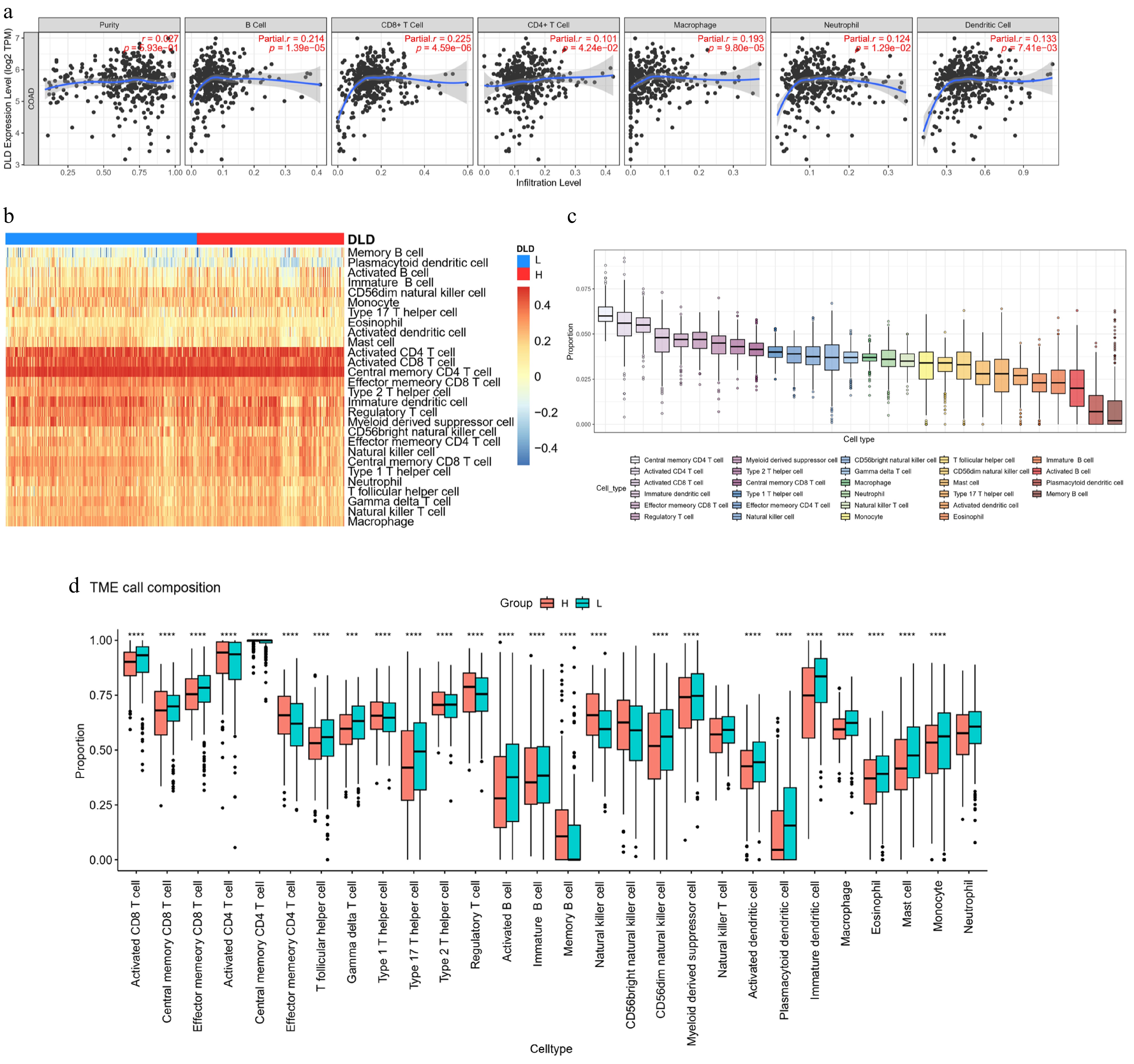
Figure 7.
DLD expression correlates with the degree of immune infiltration in CC. (a) Spearman's correlation between DLD expression and the abundance of six types of infiltrating immune cells. (b) Heat map showing differences in immune cell infiltration between the high- and low-DLD groups. (c) Proportion of 28 immune cell subtypes infiltrating in CC. (d) Changes in 28 immune cell subtypes between the high- and low-DLD groups in CC tumor samples. * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001.
To confirm the role of DLD in the tumor immune microenvironment, we assessed the levels of 28 immune cells using the ssGSEA algorithm (Fig. 7b–d). The TCGA-COAD samples were assigned to either the high-DLD-expressing or low-DLD-expressing groups on the basis of the optimal cut-off point. In the high-DLD samples, CD4 T cells, Type 1 T helper cells, Type 2 T helper cells, regulatory T cells, memory B cells, and NK cells were significantly increased. However, in samples with low DLD expression, CD8 T cells, Tfh cells, gamma delta T cells, Type 17 T helper cells, activated B cells, immature B cells, CD56dim natural killer cells, myeloid-derived suppressor cells, activated dendritic cells, plasmacytoid dendritic cells, immature dendritic cells, macrophages, eosinophils, mast cells, monocytes, and neutrophils decreased (p < 0.05; Fig. 7d). DLD potentially regulates the immune microenvironment of CC by regulating NK cells.
-
CC is a fatal malignancy of the digestive system, and its prognosis remains very poor[21]. Currently, conventional treatment options also have limitations in improving patient survival outcomes, including surgery and chemotherapy[22−24]. At the same time, immune checkpoint inhibitor therapy offers patients better monitoring opportunities, unique treatment options, and greater hope for prolonged survival[4]. Therefore, the discovery of new biomarkers associated with immunomodulation in CC is crucial for its diagnosis, treatment, and prognosis.
The DLD gene encodes a member of the Class I pyridine nucleotide-disulfide oxidoreductase family, which plays multiple roles in the decarboxylation of pyruvate and the oxidation of dihydrolipoamide and is involved in constituting α-keto acid dehydrogenase complexes (e.g., α-ketoglutarate dehydrogenase, α-ketoadipate dehydrogenase, and glycine decarboxylase)[7,25−27]. DLD is one of the key genes for copper-induced cell death, and DLD is an important component of the mitochondrial aerobic respiration process. It plays a key role in the process of copper-induced cell death, which may provide new ideas on the therapeutic effects of certain cancer-targeting drugs[6,28]. Previous studies have found that DLD is involved in constituting the pyruvate dehydrogenase complex and the α-ketoglutarate dehydrogenase complex, which are associated with several fatal metabolic diseases and have been identified as important metabolic targets in cancer[29−32]. Evidence from other studies suggests that integrins mediate the rapid uptake of DLD by cells, upregulate DLD levels, likely affect mitochondrial metabolism, and induce regression and a reduction in the number of melanoma cells in humans and mice by promoting reactive oxygen species (ROS) generation and altering the energy metabolism[33]. However, more bioactivities of DLD as a promising therapeutic target need to be found in different diseases.
Expression profiling based on the TCGA-COAD database and the GSE:39582 dataset showed that DLD expression was significantly lower in CC than in normal tissues (Fig. 1c). We obtained DLD protein expression levels in normal human colon tissues and CC tissues from the HPA database, and the results were consistent with the mRNA expression levels of DLD (Fig. 1c, d). In our study, low DLD expression was associated with malignant clinicopathological features such as staging and grading. We then evaluated the prognostic value of DLD in CC using two cohorts and found that high DLD expression was significantly associated with the OS prognosis of CC patients (Fig. 3a, b).
To reveal the hidden mechanism behind their reduced invasive growth pattern, we constructed gene co-expression network for DLD using the TCGA-COAD dataset and performed GO and KEGG enrichment analysis. In this study, the expression of CYCS, NDUFA5, SDHD, and SNX4 in CC showed the highest correlation with DLD. Copper ion-induced ROS leads to the release of Cytochrome C (CYCS) release from mitochondria into the cytoplasm, resulting in caspase activation and DNA breakage in PC12 pheochromocytoma cells[34]. Deubiquitinated CYCS affect the progression of liver cancer by regulating apoptosis, metabolism and other pathways[35]. Moreover, NDUFA5 (ubiquinone oxidoreductase subunit A5) is involved in encoding the B13 subunit of the mitochondrial respiratory chain complex I, a protective prognostic gene in ovarian cancer[36,37]. Succinate Dehydrogenase Complex Subunit D (SDHD) benefits patients with COAD and is a novel biomarker[38]. A retrospective analysis based on GSE66229 data showed that SNX4 was associated with good survival outcomes in gastric cancer[39]. GO and KEGG enrichment analysis based on co-expression correlated with many classical signaling pathways such as metabolic pathways, oxidative phosphorylation, and the peroxisome. GSEA showed that differential genes in the high-DLD group were mainly enriched in some important tumor-associated KEGG pathways. This study is the first to reveal that DLD is associated with pathways in CC in cancer, the hedgehog signaling pathway, focal adhesion, the ECM, WNT signaling, cell adhesion molecules, leukocyte transendothelial migration, regulation of the actin cytoskeleton, the calcium signaling pathway, and the chemokine signaling pathway.
Tumor mutation burden (TMB) reflects the number of mutations in the tumor and generates immunogenic neoantigens, thus increasing the likelihood of T cell recognition and correlating with the response to immune checkpoint inhibitors (ICIs)[40]. Consistent with PD-L1 expression, the TMB might provide a reference for the selection of ICIs for the treatment of tumor patients[41,42]. In this study, a somatic mutation analysis was performed in the TCGA database based on DLD expression levels. We listed the top 30 genes with the highest mutation rates in the high- and low-DLD groups (Fig. 5a, b). TTN, APC, TP53, KRAS, MUC16, and SYNE1 were the most frequently mutated genes in both groups, while PIK3CA appeared in the top five in the high-DLD group. PIK3CA mutations identified colorectal cancer as a poor prognostic factor associated with solid organ transplantation and mismatch repair deficiency, and PIK3CA mutations predict a reduced response to anti-EGFR therapy in patients with metastatic colorectal cancer[43,44]. In conclusion, low expression of DLD has a higher tumor mutation rate compared with high expression, which would provide some direction for the selection of immunotherapy regimens for patients with low expression DLD.
This study investigated the potential mechanisms of high DLD expression for prognosis, using cell function and immune infiltration levels in the COAD single-cell database. The CancerSEA database reveals the functional states of different cancer cells at the single-cell level, involving 25 functional states of 41,900 cancer cells from 14 cancer types[45]. In this study, DLD expression was positively correlated with tumor cell invasion and DNA repair, as analyzed in the CancerSEA database. In contrast, DLD expression was negatively correlated with angiogenesis. These results suggest that high expression of DLD plays a crucial role in the suppression of colon tumors. Recent evidence suggests that DLD is highly correlated with the abundance of lipid acylated protein in a variety of human tumor cells and that cell lines with high levels of lipid acylated proteins are sensitive to copper-induced cell death[6]. This may be another important factor in the suppression of tumor invasion by DLD.
Immune infiltrating cells are an important component of the tumor microenvironment and play an important role in influencing tumors' growth and progression, treatment outcome, and patient prognosis[46−48]. Higher immune infiltration in colorectal cancer is associated with improved time to recurrence, OS, and disease-free survival[49,50]. TIMER was used to explore the correlation between DLD expression and the level of immune cell infiltration in tumors and showed that samples with high DLD expression levels tended to infiltrate more immunocytes. By analyzing the infiltration levels of 28 immune cells in the high- and low-DLD groups using the ssGSEA algorithm, we also observed that the infiltration levels of CD4 T cells, Type 1 T helper cells, Type 2 T helper cells, regulatory T cells, memory B cells, and NK cells in the high-DLD group were elevated, whereas infiltration levels in CD8 T cells, Tfh cells, gamma delta T cells, Type 17 T helper cells, activated B cells, immature B cells, CD56dim natural killer cells, myeloid-derived suppressor cells, activated dendritic cells, plasmacytoid dendritic cells, immature dendritic cells, macrophages, eosinophils, mast cells, monocytes, and neutrophil were reduced. A higher percentage of NK cells were activated in the high-DLD group, a cell particularly known in oncology for its innate ability to recognize and spontaneously kill tumor cells[51]. Thus, a greater proportion of NK cells being activated may reduce the tumor's immune escape in the tumor immune microenvironment, which may be a potential reason for the prognostic benefit of high DLD expression in CC patients. Interestingly, the high-DLD group also possessed a higher proportion of Tregs. It is known that Tregs contribute to suppressing excessive immune activation and act as immunosuppressive cells to coordinate the tumor's immune escape and are considered targets of systemic immunotherapy[52]. These analyses suggest that DLD is involved in regulating the immune mechanisms of the CC tumor microenvironment, particularly the regulation of NK cells. We hypothesize that DLD plays an important role in recruiting infiltrating immune cells and regulating immunity in CC, thereby affecting prognosis. However, more studies are needed to confirm this hypothesis, especially the effect of DLD on NK cells in the CC microenvironment.
-
In conclusion, our study shows that DLD has low expression levels in CC tissues, and these levels correlate with clinicopathological features. Furthermore, high DLD expression was significantly associated with beneficial prognosis in CC, as validated by an analysis of the TCGA database and the GEO database. In terms of biological function, we demonstrated that DLD was negatively enriched in cancer, the hedgehog signaling pathway, focal adhesion, the ECM, and the WNT signaling pathway in CC. In addition, DLD expression correlated with the levels of the tumor's genetic mutation genes and various immune cells in the tumor microenvironment. Taken together, these results suggest a potential role for DLD as a prognostic marker and therapeutic target for future CC.
This article was supported by Shaanxi Province Key Research and Development Program (2021SF-129), Medical Project of Xi'an Science and Technology Bureau [2019114713YX002SF035(1)], and Youan Medical Alliance for the Liver and Infectious Diseases Research Special Fund (LM202028).
-
Ethical review and approval were waived for this study because all data used were obtained from publicly available databases, especially The Cancer Genome Atlas (TCGA) and Gene Expression Omnibus (GEO). Both the TCGA and GEO databases provide open access to de-identified datasets for research purposes. TCGA data are processed in compliance with ethical guidelines such as the Health Insurance Portability and Accountability Act (HIPAA), with all patient identifiers removed; GEO datasets also adhere to standard ethical practices, where individual-level information is anonymized prior to public release. Since this study utilized only these pre-existing, de-identified public data without involving any direct interaction with human participants or access to identifiable private information, ethical review was not required.
-
The authors confirm their contributions to the paper as follows: study conception and design: Fan L, Zhang M; data collection: Lu N; analysis and interpretation of results: Li A, Li J; draft manuscript preparation: Zhang L, Cui M. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Ning Lu, Ling Fan, Lingmin Zhang
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Lu N, Fan L, Zhang L, Li A, Li J, et al. 2025. High expression of the cuproptosis-related gene DLD in relation to good prognosis and immune cells infiltration in colon cancer. Gastrointestinal Tumors 12: e016 doi: 10.48130/git-0025-0017 

High expression of the cuproptosis-related gene DLD in relation to good prognosis and immune cells infiltration in colon cancer
- Received: 06 March 2025
- Revised: 17 June 2025
- Accepted: 05 September 2025
- Published online: 22 October 2025
Abstract: Cuproptosis is a new mode of cell death that involves a copper-dependent process, and dihydrolipoamide dehydrogenase (DLD) is a relevant positive regulator of cuproptosis. However, the role of DLD in the pathogenesis of colon cancer (CC) remains to be investigated. Correlations between DLD expression and clinicopathological parameters were analyzed in the University of Alabama at Birmingham Cancer database. Kaplan-Meier (K-M) survival curves were used to elucidate the relationship between DLD expression and survival time of CC patients, which was validated in the GSE:39582 dataset. Then gene set enrichment analysis (GSEA) was performed to explore the biological function of DLD. The relationship between DLD expression and cancer-related immune infiltration was analyzed using the Tumor Immunization Estimation Resource database (TIMER), single-sample gene set enrichment analysis (ssGSEA), and others. We found that DLD was downregulated in CC samples and that increased DLD expression was associated with better overall survival. Tumor-related immune infiltration analysis revealed that DLD potentially recruits infiltrating immune cells and modulates immunity in CC, thus affecting prognosis. DLD plays a role in inducing cuproptosis and regulating immunity in CC, which can be considered as a new biomarker for CC.
-
Key words:
- DLD /
- Cuproptosis /
- Prognosis /
- Immune infiltrate /
- Colon cancer