









-
Gastric cancer is a digestive tract malignancy that causes many deaths worldwide, especially in China[1]. Gastric cancer has the third-highest incidence and third-highest mortality rates among all cancer types in China[2]. As a result, gastric cancer imposes a significant burden on economic development and the healthcare system. Over the past few decades, gastric cancer treatments, including surgical treatment, chemotherapy, targeted therapy, and immunotherapy, have advanced significantly[3]. However, these treatments have been found to fail in certain instances due to relapse or progression of gastric cancer. Therefore, this research set out to identify an effective biomarker for diagnosing or treating gastric cancer.
Glycosyltransferase 8 domain-containing protein 1 (GLT8D1) belongs to the glycosyltransferase family and is located on chromosome 3p21.1. Several studies have revealed that the expression of GLT8D1 is downregulated in schizophrenia[4,5] and amyotrophic lateral sclerosis[6,7]. The studies also state that GLT8D1 is correlated with cognitive function[5]. However, the biological function of GLT8D1 remained unclear. Recent studies have shown that GLT8D1 is related to several tumors, including human melanoma[8], glomangiopericytal tumor[9], glioblastoma[10], and head and neck squamous cell carcinomas[11]. In addition, studies have shown that poor prognosis in melanoma is caused by the overexpression of GLT8D1[8]. The studies have also shown that the mutation of GLT8D1 germline results in multiple sarcomas[9]. Ilina et al.[10] revealed that GLT8D1 promotes the migration of glioblastoma cells. Nonetheless, other reports have shown that the knockdown of GLT8D1 promotes apoptosis and cell cycle arrest and inhibits glioma stem cell self-renewal[12]. Collectively, these data suggest a tumor-promoting function for GLT8D1.
One of the most common changes that proteins undergo after translation is glycosylation. Glycosylation is involved in vital pathological processes, especially in cancers[13,14]. N- and O-glycosylation are the two forms of glycosylation that are linked to cancer[15,16]. GLT8D1, a member of the glycosyltransferase family, has been reported to influence cancer cell behavior by regulating glycosyltransferase activity[10]. Protein glycosylation is accompanied by oncogenic alteration in gastric cancer[17]. However, there are no studies elucidating the relationship between gastric cancer and GLT8D1. Therefore, this study performed multiple analyses to determine the biological role and prognostic significance of GLT8D1 in gastric cancer via multiple databases and experiments. The results of this study demonstrated that overexpression of GLT8D1 resulted in poor overall survival and exerted oncogenic function in gastric cancer, thereby revealing its potential as a promising therapeutic target for this disease.
-
The information on RNA sequencing for the cancer genome atlas (TCGA) was obtained from the UCSC Genome Browser database[18]. This study selected 33 types of malignancies. In this study, clinical details and 'Level 3' RNA-sequencing data on gastric cancer were gathered from the TCGA website. In addition, this study followed the publishing standards set by the TCGA project.
UALCAN analysis
-
We investigated how the expression of GLT8D1 influences cancer stage and tumor stage using the UALCAN database[19]. On the other hand, the TCGA-STAD dataset was explored in the UALCAN database. In order to analyze the variance between the two groups, a Student's t-test was used. A significant difference was one with a p value less than 0.05.
Differentially expressed gene (DEG) analysis
-
Using the median expression value of GLT8D1, the gastric cancer samples were divided into two groups: high and low expression levels of GLT8D1. With a log2 fold change threshold of 1.5 and adjusted p value (FDR) < 0.05, the DEGs were identified using the Limma R program.
STRING analysis
-
STRING database, an online analysis website for protein–protein interactions (PPIs), was used to show the potential correlation among proteins and GLT8D1[20].
GeneMANIA analysis
-
GeneMANIA database is an online website that integrates various algorithms to predict gene function or gene–gene interaction networks, including physical interactions, gene co-expression, functional gene prediction, gene co-localization, genetic interactions, pathways, and shared protein domains[21]. This study used GeneMANIA to predict the interaction between GLT8D1 and other functional genes.
Functional/pathway enrichment analysis
-
The study also explored the function or pathway in which GLT8D1 is a factor in the oncogenic process that causes stomach cancer. DEGs were chosen in accordance with the criteria of log2 FC > 1.5 and a p value less than 0.05 to carry out the functional/pathway enrichment analysis. Gene ontology (GO) was used to define the functional proteins in specific species[22]. Kyoto Encyclopedia of Genes and Genomes (KEGG) was used to confirm the potential pathway of a functional protein[23]. The R package ClusterProfiler was used to evaluate the GO and KEGG[24]. Terms with an FDR-adjusted p value < 0.05 were considered significantly enriched.
Gene set enrichment analysis (GSEA)
-
Gene group function and biological regulation were the focus of GSEA[25]. Therefore, to determine the potential role of GLT8D1, GSEA analysis was employed. The GSEA analysis was done using the ClusterProfiler package in R. Then, each gene set was permutated 1,000 times. In this experiment, significant differences were defined as those with an adjusted p value of 0.05 or lower and an FDR q-value of 0.25 or below.
Prognostic value evaluation of GLT8D1
-
Univariate and multivariate analyses were performed to identify the risk factors of overall survival. To ascertain the predictive importance of GLT8D1 in gastric cancer, the Kaplan–Meier analysis was conducted on the TCGA-STAD dataset to compare high- and low-expression GLT8D1 groups. In addition, time-dependent receiver operating characteristic (ROC) curve analysis was used to determine the predictive significance of GLT8D1 in gastric cancer for 1-, 3-, and 5-year overall survival.
Prognostic nomogram construction and evaluation
-
A prognostic nomogram containing clinical characteristics and GLT8D1 expression was constructed to forecast the overall chance of survival for people with stomach cancer. The concordance index (C-index) was used to discriminate the nomogram. It was determined whether the expected probabilities and observed probabilities generated by the nomograms were accurate by using calibration curves. ROC analysis was used to evaluate the performance of the prognostic nomogram.
External validation of prognostic nomogram
-
The gastric cancer dataset GSE62254 and the Gastrointestinal Surgery Center of the First Affiliated Hospital of Sun Yat-sen University (FAHSYSU) cohort were utilized as the external validation set of the prognostic nomogram. The FAHSYSU cohort was included in the research if the patients had undergone a radical gastrectomy, were followed up for at least 60 months following therapy, and if their tumor stage was between I and III. On the other hand, patients who had undergone neoadjuvant therapy and whose follow-up file was incomplete were excluded from this study. In addition, patients whose imaging indicated existing distant metastasis were also excluded from this study. In total, 60 gastric cancer patients sampled between January 2009 and January 2013 were included in this study. The random sampling method was used to collect participants for this study. Paraffin sections of gastric cancer tissue from these patients were collected for further immunohistochemistry staining experiments. ROC analysis was also conducted to validate the capability of the prognostic model.
Immunohistochemistry (IHC) staining
-
To perform immunohistochemistry staining, for 3 h at 65 °C, the slides were roasted. Then, the slides were deparaffinized using xylene and hydrated using a different gradient alcohol solution. The slides were washed thrice using phosphate buffer saline (PBS) solution. Then, antigen retrieval was performed using EDTA solution in a water bath at 95 °C for 45 min, and 3% H2O2 was used for nonspecific antigen blocking. The primary antibody used was anti-GLT8D1 (1:400, Solarbio, K009871P). After 24 h, secondary antibody incubation and DAB (Gene Tech, GK600510) staining, dehydration, and xylene soaking were performed step by step. Two senior pathologists, who were blinded to the clinical outcomes, evaluated the immunohistochemistry staining of slides. The staining score was calculated by multiplying the score of staining intensity (negative signal—0, weak signal—1, moderate signal—2, strong signal—3) by the score of staining area (negative—0, 1%–25%—1, 26%–50%—2, 51%–75%—3, 76%–100%—4).
Cell lines and reagents
-
SGC-7901, HGC-27, MGC-803, SNU-719, MKN45, KATO III, MKN28, and AGS were purchased from American Type Culture Collection. The culture medium used for SGC-7901, HGC-27, MGC-803, SNU-719, MKN28, and MKN45 cells was RPMI-1640 (Gibco, 11875119) supplemented with 10% fetal bovine serum (FBS; Biochannel, BC-SE-FBS01) and 1% penicillin-streptomycin (PS) solution (Biosharp, BL505A). The KATO III cell line was cultured in IMDM medium (Gibco, 12440053) supplemented with 10% FBS and 1% PS, and the AGS cell line was maintained in DMEM/F-12 medium (Gibco, 11320033) containing 10% FBS and 1% PS. Following this, the cell lines were incubated at 37 °C and 5% CO2. 20 ng/mL recombinant human IL-6 protein (ProteinTech, HZ-1019) was used for treating assigned cells.
Gastric cancer tissues
-
A total of eight matched sets of stomach cancer and nearby normal tissues were acquired for this investigation. Following collection, the tissues were frozen and kept in liquid nitrogen. Before participating in this study, the patients were given informed consent forms.
Western blot analysis
-
Cells or tissues were washed using PBS and then treated with RIPA lysis buffer (Beyotime, P0013). Afterward, the protein content was determined with the use of a BCA Protein Assay Kit (KeyGEN, KGP903). Primary antibodies were anti-GLT8D1 (1:1,000, Solarbio, K009871P), anti-GAPDH (1:100,000, ProteinTech, 60004-1-Ig), anti-STAT3 (1:5,000, ProteinTech, 60199-1-Ig), and anti-pSTAT3 (1:1,000, ProteinTech, 39613). Western blot experiment was carried out as previously reported[26]. Lastly, we used an electrochemiluminescence (ECL) device equipped with an enhanced chemiluminescence reagent (Meilunbio, MA0186-2) to observe immunoreactive proteins.
Cell viability assay
-
Using a Cell Counting Kit-8 (Meilunbio, MA0218-1), we were able to determine cell viability. A 96-well plate was seeded with 1 × 103 cells per well, and the plate was cultured for 4 d to measure cell viability. Throughout the incubation period, CCK-8 reagent was added to cells for 2 h. After incubation, cells in 450 nm absorbance value were measured using a multimode reader (BioTek Synergy 2).
Transwell assay of migration and invasion
-
To perform a transwell migration assay, 5 × 104 cells were resuspended in serum-free media in top chambers (8 µm pore), and media supplemented with 10% FBS was added to bottom chambers (24-well plate). For the transwell invasion assay, upper chambers (8 µm pore) were covered with Matrigel, while bottom chambers (24-well plate) were supplemented with media containing 10% FBS. After culturing for 48 h, the upper chamber was washed with PBS buffer thrice. Finally, 0.1% crystal violet was used to visualize cell invasion and migration after fixation with 4% paraformaldehyde. Five randomly selected fields were collected using an Olympus Inverted Microscope.
Cell transfection
-
Small interference RNA (siRNA) of GLT8D1 was purchased from GenePharma Company. The siRNA transfection was performed following the manufacturer's instructions. First, siRNA was added to the Opti-MEM medium (ThermoFisher, 31985070) and incubated at room temperature for 15 min. Afterward, siRNA was mixed with Opti-MEM-Lipofectamine 6000 (Beyotime, C0526). The transfection mixture was then treated with cells in a 6-well plate. Subsequently, cells in the 6-well plate were incubated in a cell incubator for 6 h. The media in the 6-well plate was replaced with fresh media. After 72 h of incubation, transfected cells were collected for subsequent experiments. The GLT8D1 siRNA (siGLT8D1-1: 5'-GCAAGGUUCUACUUGCCAATTUUGGCAAGUAGAACCUUGCTT-3', siGLT8D1-2: 5'-GCCAGCACUUGCUCAUUUATT UAAAUGAGCAAGUGCUGGCTT-3') and control (5'-UUCUCCGAACGUGUCACGUTTACGUGACACGUUCGGAGAATT-3') were used to transfect gastric cancer cells.
Human studies
-
The clinical specimens used in this study were reviewed and approved by the Medical Committee Ethics of the Seventh Affiliated Hospital of Sun Yat-sen University (No. KY-2020-024-01). In addition, patients who participated in this study had signed an informed consent.
Statistical analysis
-
We employed the Student's t-test to compare the two groups. R Studio (version 3.6.1) was used for all the statistical analyses. Wilcoxon signed-rank test was used to compare GLT8D1 expression levels between matched and unmatched samples. On the other hand, to determine whether there was a statistically significant difference, Kaplan–Meier analysis was subjected to a log-rank test. Hazard ratios were determined for each potential risk parameter using the Cox proportional hazard regression model. The univariate and multivariate analysis models were used to identify the independent risk factors of gastric cancer patients. A difference with a p value of less than 0.05 was considered statistically significant.
-
Data of pan-cancers obtained from the UCSC Genome Browser database demonstrated that GLT8D1 was overexpressed in BLCA, BRCA, CHOL, COAD, GBM, HNSC, KIRP, LIHC, LUAD, PRAD, READ, and STAD, as shown in Fig. 1a. In addition, GLT8D1 expression was found to be significantly greater in tumor tissues than in normal tissues when paired and unpaired gastric cancer samples were examined, as shown in Fig. 1b, c. Tumor stage and malignancy grade were both linked with GLT8D1 expression in gastric cancer. An analysis of the UALCAN database was performed to investigate how the expression of GLT8D1 influences tumor stage and cancer grade. These findings revealed that GLT8D1 expression was correlated with higher tumor stage and cancer grade, as shown in Fig. 1d, e.
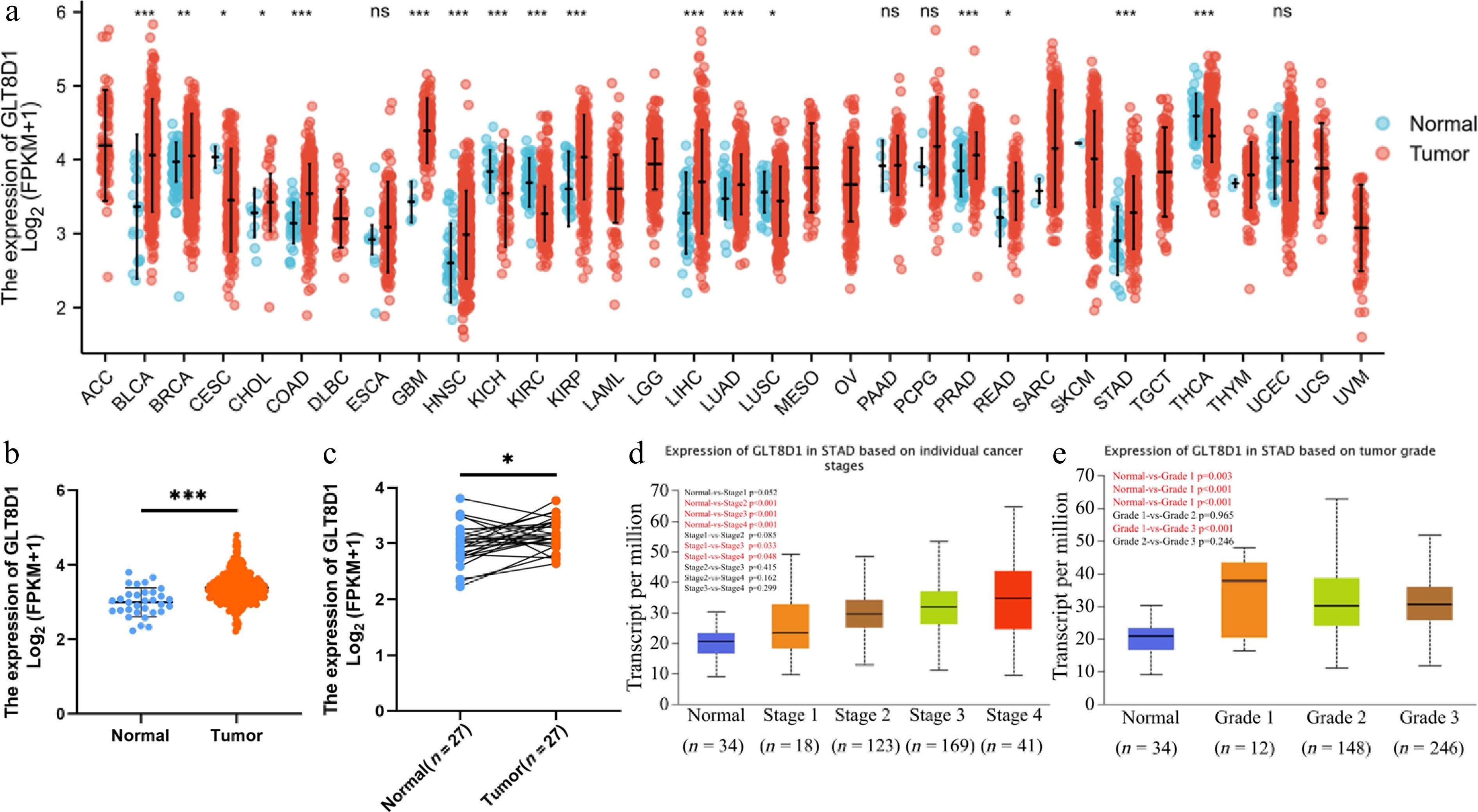
Figure 1.
Upregulated GLT8D1 expression in gastric cancer and GLT8D1 expression in different gastric cancer stages and grades. (a) mRNA expression of GLT8D1 in unpaired normal and pan-cancer. (b, c) GLT8D1 expression in unpaired and paired gastric cancer samples. (d, e) Evaluation of the expression of GLT8D1 in different tumor stages and grades in gastric cancer. * p < 0.05; ** p < 0.01; *** p < 0.001; ns, no significance.
Identification of DEGs based on GLT8D1 expression in gastric cancer
-
A comparison between the GLT8D1-high expression group and the GLT8D1-low expression group identified 161 DEGs. Of the 161 DEGs, 54 were upregulated genes, and 107 were downregulated (log2 fold change > 1.5, p < 0.05), as shown in Fig. 2a. A heatmap was used to visualize the expression of the top five upregulated DEGs and top five downregulated DEGs in gastric cancer samples, as shown in Fig. 2b.
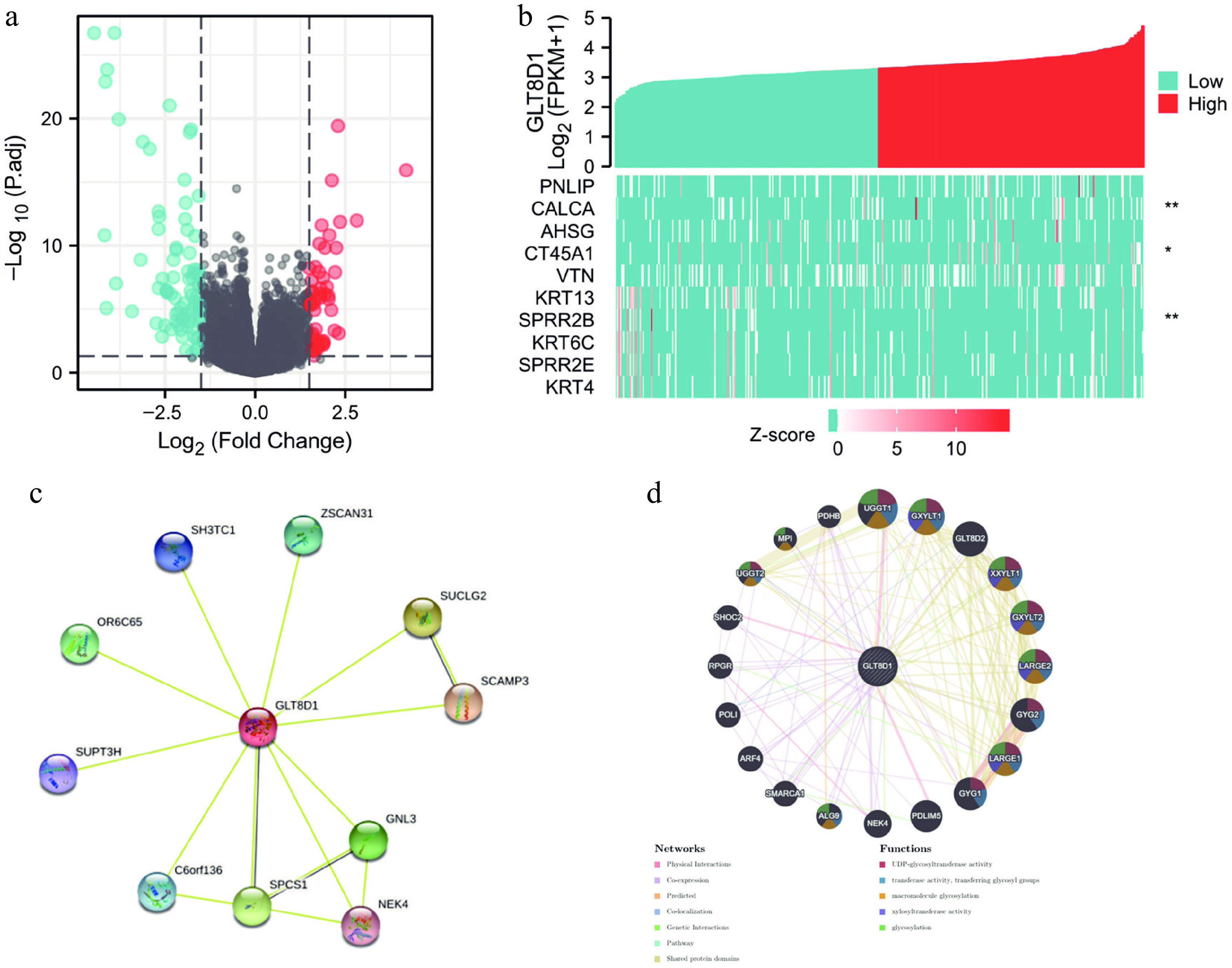
Figure 2.
Differentially expressed gene analysis of GLT8D1 in gastric cancer, protein–protein interaction, and gene–gene network of GLT8D1. (a) The volcano plot showed differentially expressed genes, including 54 upregulated (red) and 107 downregulated (blue) genes. (b) Heatmap showing the top five regulated DEGs and top five downregulated DEGs. (c) PPI network generated by the STRING database. (d) Gene–gene network constructed by the GeneMANIA database. * p < 0.05; ** p < 0.01.
Protein–protein interaction network
-
The STRING and the GeneMANIA databases were used to study the regulation of GLT8D1 in gastric cancer by exploring network interactions. An analysis of the STRING database found several nodes, including ZSCAN31, SUCLG2, SCAMP3, GNL3, NEK4, SPCS1, C6orf136, SUPT3H, OR6C65, and SH3TC1, were correlated with GLT8D1, as shown in Fig. 2c. The GeneMANIA database was used to establish a network of gene–gene interactions containing UGGT1, GXYLT1, GLT8D2, XXYLT1, GXYLT2, LARGE2, GYG2, LARGE1, GYG1, PDLIM5, NEK4, ALG9, SMARCA1, ARF4, POLI, RPGR, SHOC2, UGGT2, MPI and PDHB, as shown in Fig. 2d. The functions of these genes involve UDP-glycosyltransferase activity, transferring glycosyl groups, macromolecule glycosylation, xylosyltransferase activity and glycosylation.
GO and KEGG enrichment analyses and GSEA
-
The KEGG and GO enrichment analyses were conducted to reveal the function of GLT8D1 in gastric cancer. The GO enrichment analysis demonstrated that high GLT8D1 expression correlated with cellular components (CC), including intermediate filament cytoskeleton, intermediate filament and cornified envelope, as shown in Fig. 3a; biological process (BP), including epidermis development, skin development, and epidermal cell differentiation, as shown in Fig. 3b; molecular function (MF), including peptidase regulator activity, endopeptidase regulator activity, peptidase inhibitor activity, endopeptidase inhibitor activity, and serine hydrolase activity, as shown in Fig. 3c. On the other hand, KEGG enrichment analysis demonstrated pancreatic secretion, vascular smooth muscle contraction, protein digestion and absorption, and gastric acid secretion, as shown in Fig. 3d. In addition, GSEA showed that GLT8D1 is involved in the interferon-γ response, mTORC1 signaling pathway, IL6-JAK-STAT3 signaling pathway, and glycolysis, as shown in Fig. 3e–h.
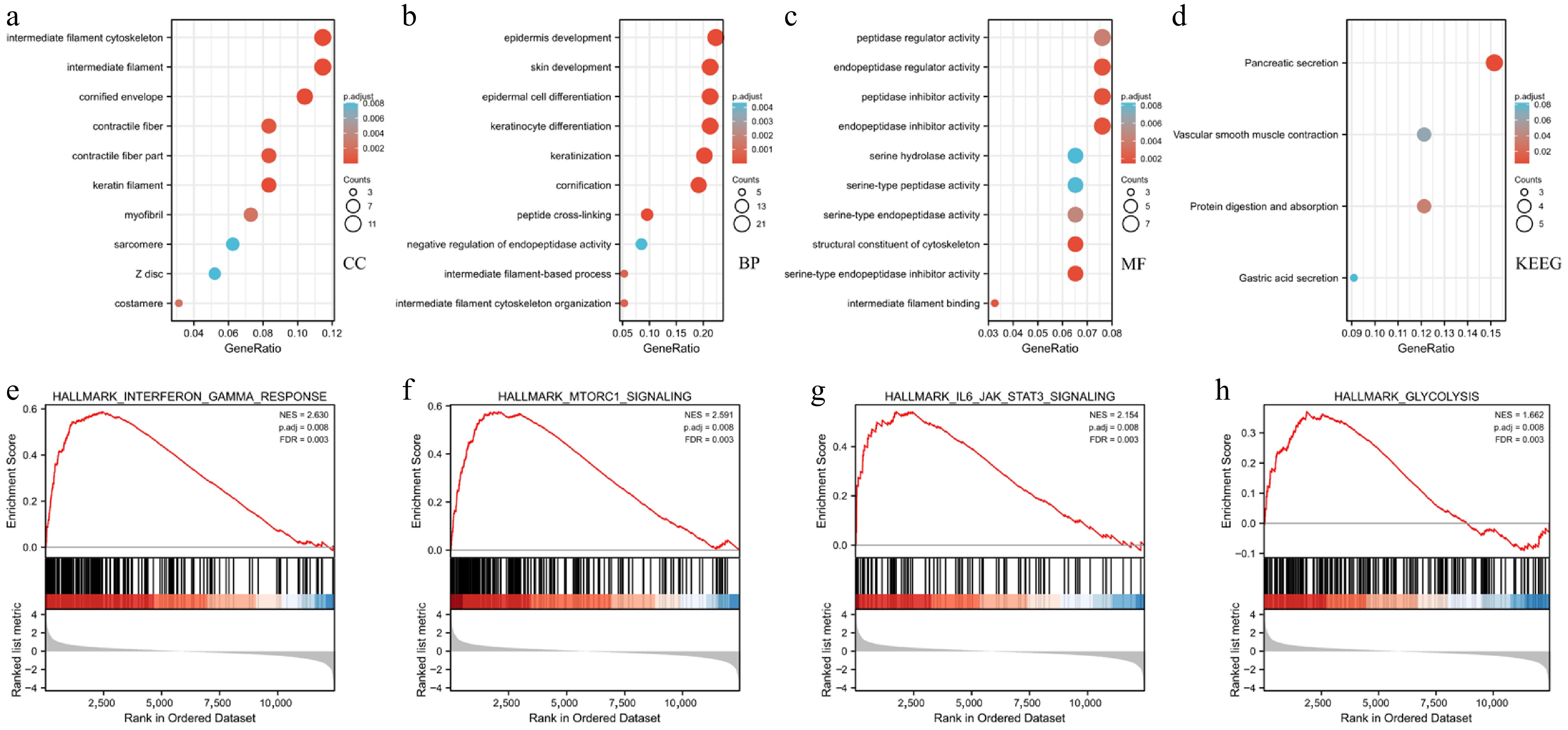
Figure 3.
GO, KEGG, and GSEA analysis of DEGs between high- and low-GLT8D1 expression in the TCGA-STAD dataset. (a) GO term of cellular component. (b) GO term of biological process. (c) GO term of molecular function. (d) Enrichment pathway analysis of KEGG. (e–h) Enrichment plot of GSEA.
GLT8D1 was an independent risk factor of overall survival in gastric cancer
-
The predictive value of GLT8D1 in gastric cancer was analyzed using the TCGA-STAD dataset for univariate and multivariate Cox regression analyses. Potential variables influencing overall survival were identified using the univariate Cox regression model. As a result, age (p = 0.005), T stage (p = 0.011), N stage (p = 0.002), M stage (p = 0.004), TNM stage (p < 0.001), and expression of GLT8D1 (p = 0.002), as shown in Table 1, are the risk factors of overall survival of gastric cancer patients. The multivariate analysis results showed that age (p = 0.005), M stage (p = 0.009), and GLT8D1 expression (p = 0.003) were the independent risk factors of overall survival in gastric cancer patients, as shown in Table 1. The multivariate analysis results also demonstrated a forest boxplot, as shown in Fig. 4a. Kaplan–Meier analysis revealed that the overall survival of patients was negatively correlated with the expression level of GLT8D1, as shown in Fig. 4b. In addition, the time-dependent ROC curve in Fig. 4c showed that GLT8D1 expression had promising prognostic power, with AUC values for predicting 1-, 3-, and 5-year overall survival of 0.573 (95% CI: 0.500–0.646), 0.633 (95% CI: 0.539–0.728), and 0.592 (95% CI: 0.443–0.742), respectively. The distribution of GLT8D1 expression, survival time, and risk score is shown in Fig. 4d.
Table 1. Univariate and multivariate analyses for the overall survival of TCGA-STAD patients.
Characteristics Total (N) Univariate analysis Multivariate analysis HR (95% CI) p value HR (95% CI) p value Age 367 ≤ 65 163 Reference > 65 204 1.620 (1.154–2.276) 0.005 1.701 (1.179–2.455) 0.005 Gender 370 Female 133 Reference Male 237 1.267 (0.891–1.804 0.188 Histologic grade 361 G1 10 Reference G2 134 1.648 (0.400–6.787) 0.489 G3 217 2.174 (0.535–8.832) 0.278 T stage 362 T1&T2 96 Reference T3&T4 266 1.719 (1.131–2.612) 0.011 1.335 (0.795–2.241) 0.275 N stage 352 N0 107 Reference N1&N2&N3 245 1.925 (1.264–2.931) 0.002 1.531 (0.859–2.731) 0.149 M stage 352 M0 327 Reference M1 25 2.254 (1.295–3.924) 0.004 2.236 (1.225–4.081) 0.009 Pathologic stage 347 Stage I&Stage II 160 Reference Stage III&Stage IV 187 1.947 (1.358–2.793) < 0.001 1.186 (0.688–2.046) 0.539 GLT8D1 370 1.868 (1.257–2.777) 0.002 1.937 (1.259–2.982) 0.003 HR, hazard ratio; CI, confidence interval. The significance of the bold values means p < 0.05. 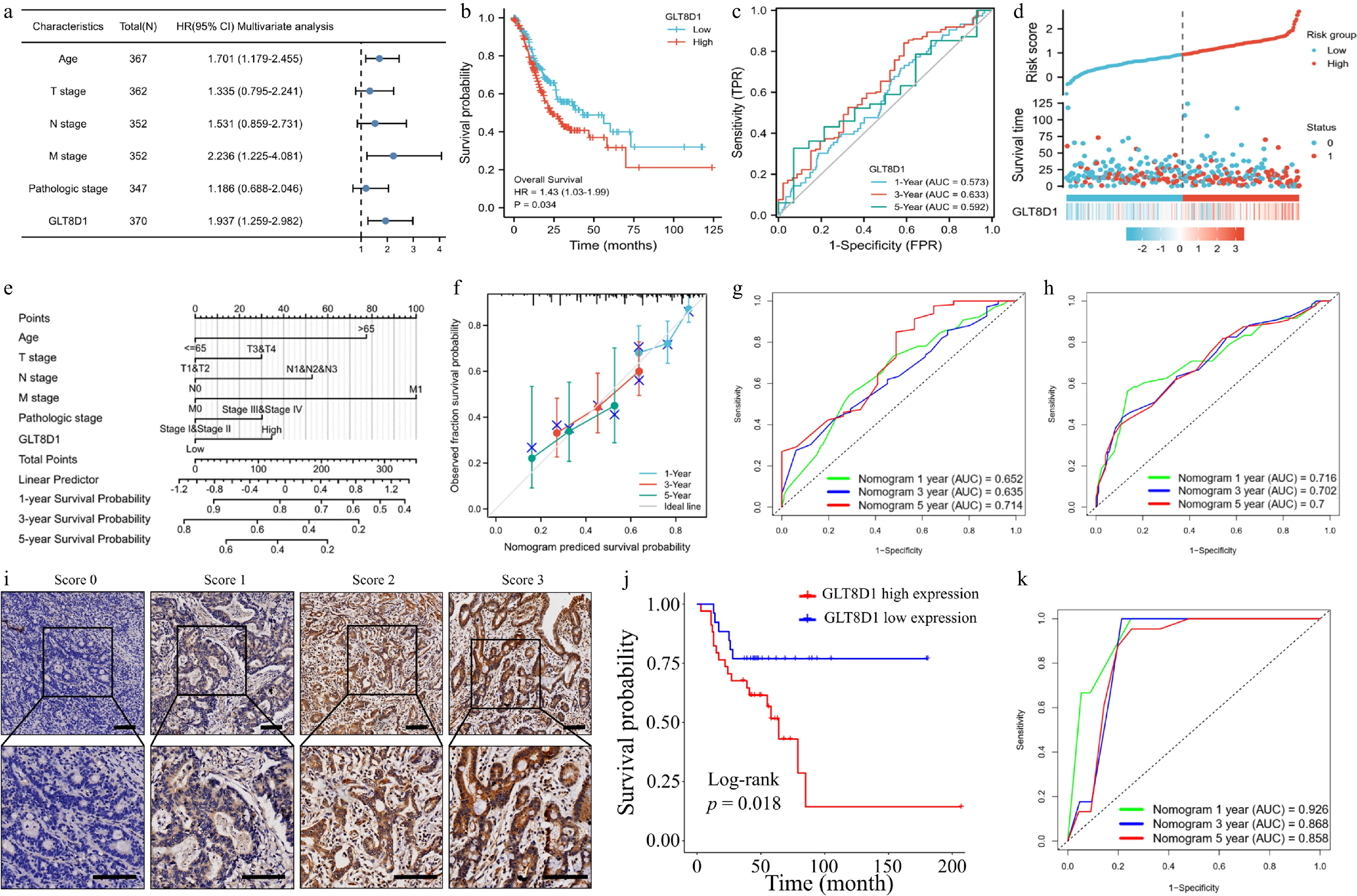
Figure 4.
Evaluation of the clinical significance of GLT8D1 in gastric cancer. (a) Forest plot showing multivariate analysis results. (b) Kaplan–Meier analysis between high- and low-GLT8D1 expression groups of TCGA-STAD samples. (c) ROC curve analysis predicted 1-, 3-, and 5-year overall survival probabilities. (d) Scatter plot showing survival time and risk score based on GLT8D1 expression. (e) Constructing a prognostic nomogram based on clinical factors and the expression level of GLT8D1. (f) Calibration curve analysis of the predicted nomogram. (g) Training set of TCGA-STAD dataset showing ROC values of the nomogram predicting 1-, 3-, and 5-year overall survival probabilities. (h) External validation of the GSE62254 dataset showing ROC values of the nomogram predicting 1-, 3-, and 5-year overall survival probabilities. (i) Immunohistochemical staining intensity of GLT8D1 in paraffin sections of the FAHSYSU cohort's gastric cancer tissues. Scale bar: 100 µm. (j) Kaplan–Meier analysis between groups of high- and low-GLT8D1 expression based on IHC results. (k) External validation of the FAHSYSU cohort showing ROC values of the nomogram predicting 1-, 3-, and 5-year overall survival probabilities.
Establishment and evaluation of prognostic nomogram
-
A nomogram was constructed to predict overall survival probability of 1, 3, and 5 years for gastric cancer patients (TCGA-STAD dataset) by combining GLT8D1 expression and clinical characteristics, as shown in Fig. 4e. The accuracy of the prognostic nomogram was assessed using the C-index and calibration curve (Fig. 4f). According to the nomogram's C-index, the average survival rate over 1, 3, and 5 years was 0.637 (CI: 0.611–0.664). The results in Fig. 4f show that the calibration curve of the prognostic nomogram exerted high coordination between the predictive and actual survival probabilities. ROC values of the TCGA-STAD cohort were 0.652, 0.635, and 0.714 for 1, 3, and 5 years of prognostic nomogram (Fig. 4g).
External validation for prognostic nomogram
-
GSE62254 dataset and FAHSYSU cohort were external validation cohorts. The ROC values of GSE62254 were 0.716, 0.702, and 0.7 for 1, 3, and 5 years of prognostic nomogram (Fig. 4h). Decision curve analysis also showed that our prognostic nomogram well predicted 1-, 3-, and 5-year overall survival in the GSE62254 dataset (Supplementary Fig. S1a–S1c). The expression level of GLT8D1 was evaluated by IHC staining. The typical images of GLT8D1 expression are shown in Fig. 4i. In accord with results of TCGA-STAD dataset, gastric cancer patients with high GLT8D1 expression level also have poorer survival than those with low GLT8D1 expression in the FAHSYSU cohort (Fig. 4j). Moreover, GLT8D1 expression was also found to be an independent risk factor for patients with gastric cancer in both univariate and multivariate analyses, as shown in Table 2. The ROC values of the FAHSYSU cohort were 0.926, 0.868, and 0.858 for 1, 3, and 5 years of prognostic nomogram (Fig. 4k).
Table 2. Univariate and multivariate analyses for overall survival of the First Affiliated Hospital of Sun Yat-sen University cohort.
Characteristics Total (N) Univariate analysis Multivariate analysis HR (95% CI) p value HR (95% CI) p value Age 60 < 60 48 Reference ≥ 60 12 2.229 (0.921–5.396) 0.076 Gender 60 Female 30 Reference Male 30 0.566 (0.247–1.295) 0.178 Gastrectomy 60 Proximal 1 Reference Distal 38 0.182 (0.023–1.466) 0.110 Total 21 0.722 (0.094–5.574) 0.755 Tumor diameter 60 < 5 cm 46 Reference ≥ 5 cm 14 3.103 (1.315–7.323) 0.010 1.997 (0.790–5.048) 0.144 Lymphovascular invasion 60 No 51 Reference Yes 9 4.702 (1.889–11.710) 0.0009 5.511 (1.814–16.740) 0.003 Perineural invasion 60 No 55 Reference Yes 5 2.895 (0.838–9.997) 0.093 T stage 60 T1&T2 23 Reference T3&T4 37 18.830 (2.539–139.6) 0.004 11.681 (1.240–110.067) 0.032 N stage 60 N0 22 Reference N1&N2&N3 38 19.99 (2.691–148.5) 0.003 7.748 (0.983–61.071) 0.052 Differentiation 60 Poor 48 Reference Well/moderate 12 0.365 (0.086–1.554) 0.173 Borrmann type 60 I-II 26 Reference III-IV 34 9.006 (2.644–30.67) 0.0004 1.401 (0.331–5.925) 0.647 GLT8D1 60 High expression 34 Reference Low expression 26 0.341 (0.134–0.869) 0.024 0.332 (0.126–0.874) 0.026 HR, hazard ratio; CI, confidence interval. The significance of the bold values means p < 0.05. GLT8D1 was upregulated in gastric cancer cell lines and tissues
-
To evaluate the expression level of GLT8D1 in gastric cancer cell lines, Western blot analysis results in Fig. 5a show that GLT8D1 was overexpressed in HGC-27, KATO III, SGC-7901, and SNU-719 cell lines. In addition, the results in Fig. 5b show that gastric tissues had higher expression of GLT8D1 than the nearby normal tissues.
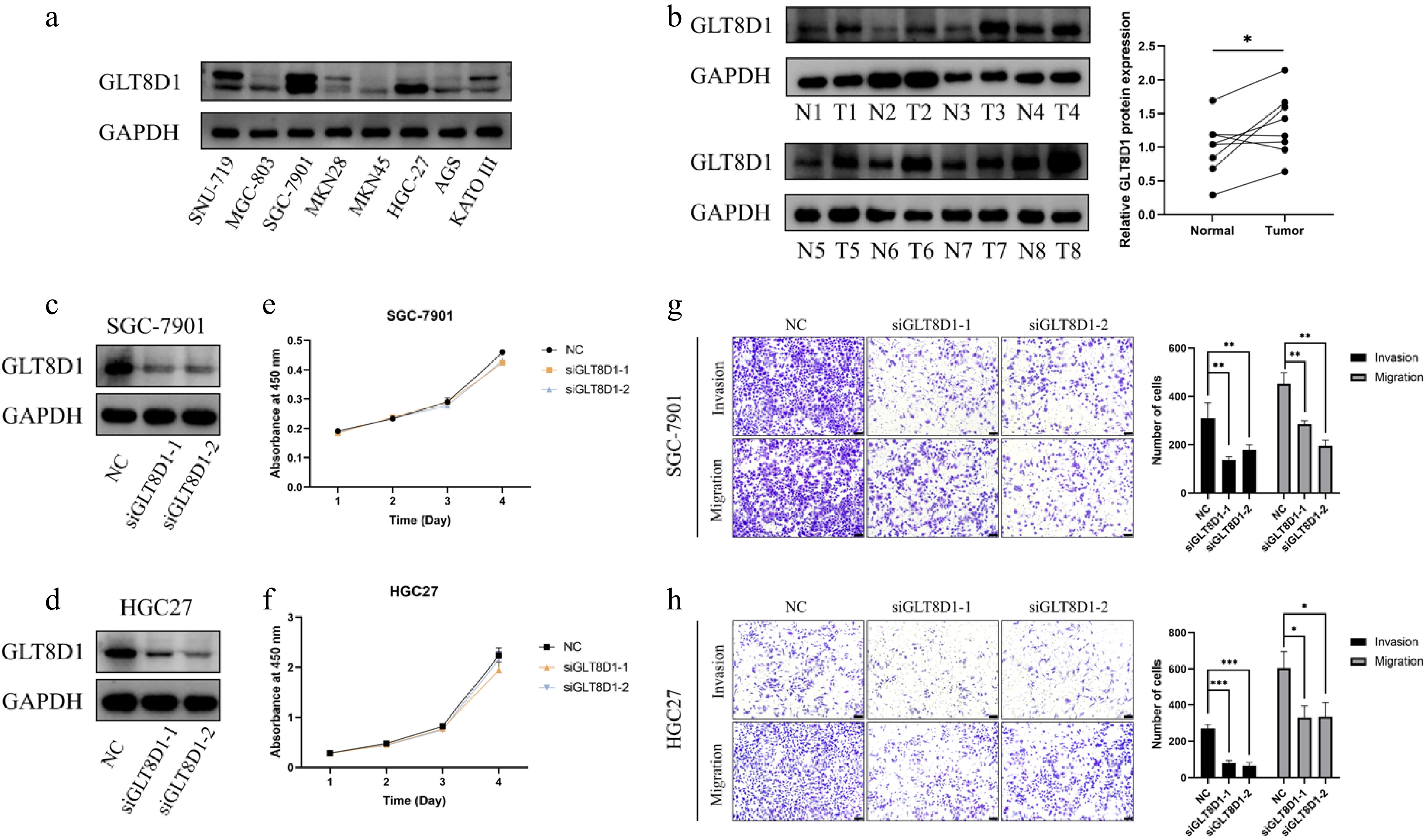
Figure 5.
GLT8D1 expression level and biological function analysis. (a) GLT8D1 expression in different gastric cancer cell lines. (b) GLT8D1 expression in gastric cancer tissue and adjacent normal tissue specimens. (c) Western blot analysis showed GLT8D1 knockdown in an SGC-7901 cell line. (d) Western blot analysis showed GLT8D1 knockdown in the HGC-27 cell line. (e) Cell viability assay of GLT8D1 in the SGC-7901 cell line. (f) Cell viability assay of GLT8D1 in the HGC-27 cell line. (g) Transwell assay of GLT8D1 in an SGC-7901 cell line. (h) Transwell assay of GLT8D1 in the HGC-27 cell line. * p < 0.05; ** p < 0.01; *** p < 0.001. Scale bar: 100 µm.
GLT8D1 does not affect gastric cancer cell proliferation but promotes migration and invasion in vitro
-
First, siRNA transfection was used to knock down GLT8D1 expression in HGC27 and SGC-7901 cells. The results in Fig. 5c, d show that the GLT8D1 expression in siGLT8D1group was significantly lower than that in the control group. Thus, we performed functional assays in vitro. The cell viability assay results in Fig. 5e, f show that GLT8D1 knockdown has little effect on the viability of gastric cancer cells. Transwell assay revealed that GLT8D1 knockdown inhibited cell migration and invasion, as shown in Fig. 5g, h.
GLT8D1 promotes gastric cancer metastasis, which is associated with activation of the IL6/STAT3 signaling pathway
-
To detect whether GLT8D1 gets involved in activating the IL6/STAT3 signaling pathway to promote gastric cancer metastasis, IL6 (20 ng/mL) was treated with GLT8D1 knockdown cells for 48 h. IL6 treatment rescued the expression level of p-STAT3 in GLT8D1 knockdown-induced downregulation (Fig. 6a). Transwell assay also showed that IL6 treatment significantly reversed the effect of inhibition GLT8D1 knockdown gastric cancer cells (Fig. 6b, c). The GSE206329 dataset contains the RNA-sequencing data of 3 cases of primary gastric cancer and the pair ovarian metastatic tissues. GLT8D1 was also found in DEG analysis (ovarian metastasis versus primary gastric cancer) (Fig. 6d). Besides, the expression level of GLT8D1 in ovarian metastasis was significantly higher than in primary gastric cancer tissues (Fig. 6e).
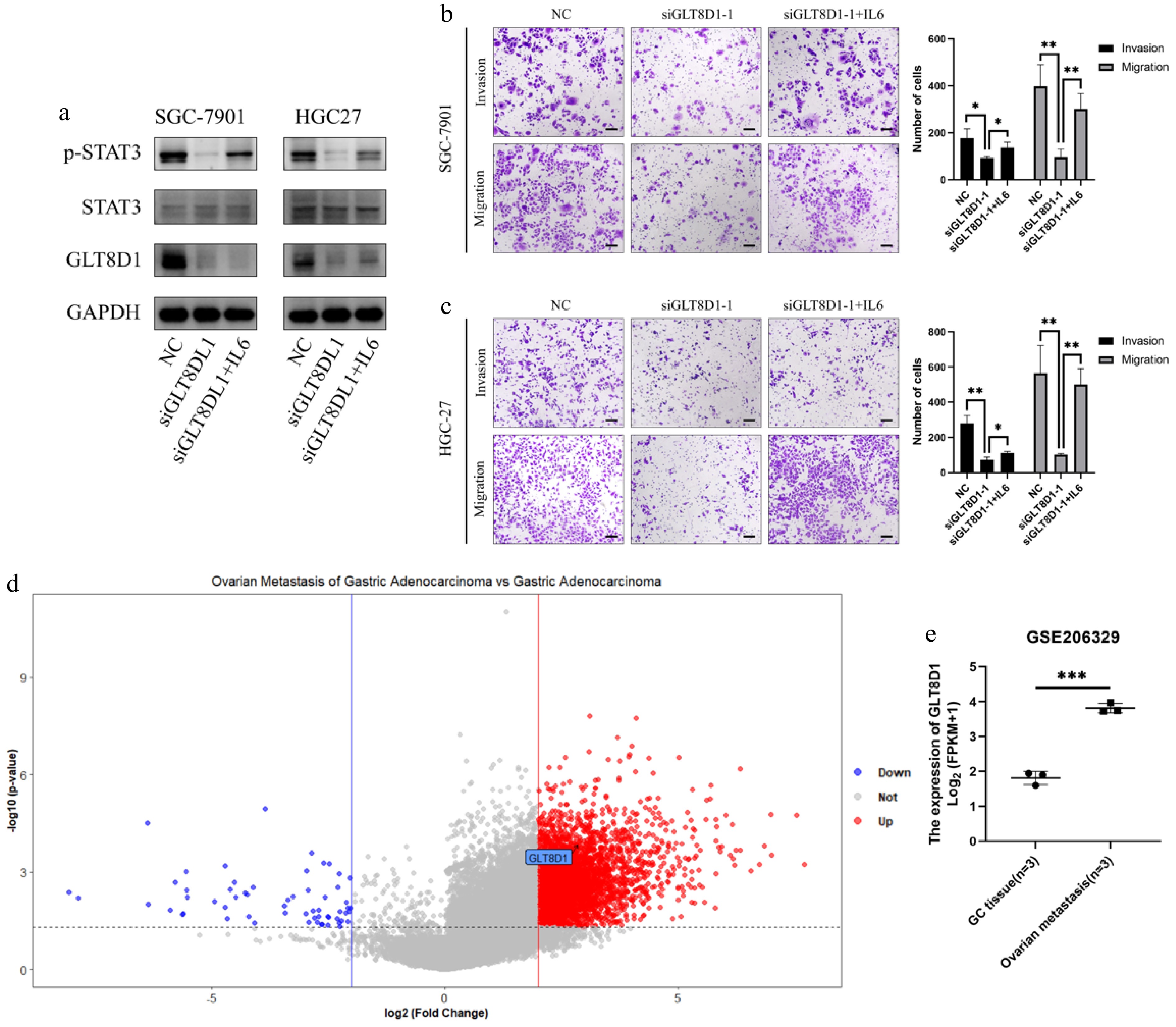
Figure 6.
GLT8D1 promotes gastric cancer cell invasion and migration, which is associated with activating the IL6/STAT3 signaling pathway. (a) Western blot analysis showed p-STAT3, STAT3, and GLT8D1 expression levels in GLT8D1 knockdown and GLT8D1 knockdown after IL6 treatment (20 ng/mL for 48-h incubation). (b, c) Transwell assay analysis showed GLT8D1 knockdown and GLT8D1 knockdown after IL6 treatment. (d) Volcano plot analysis of GSE206329 showed GLT8D1 was an upregulated gene (black arrow) in DEGs (ovarian metastasis versus primary gastric cancer). (e) GLT8D1 expression level in primary gastric cancer tissue and ovarian metastasis in the GSE206329 dataset. * p < 0.05; ** p < 0.01; *** p < 0.001. Scale bar: 100 µm.
-
Many studies state that gastric cancer is one of the most common malignancies in China[27]. Although various oncology and clinical research studies have been done on gastric cancer, its prognosis remains poorly understood[28]. However, the high mortality rate of gastric cancer has been attributed, in part, to the lack of reliable prognostic biomarkers. Recent studies indicate that glycosylation of tumor-associated proteins is an effective screening strategy to improve clinical outcomes[29]. Glycosylation is modulated by various glycosyltransferases in the endoplasmic reticulum and Golgi apparatus[30]. Several studies have reported that the glycosyltransferases of ppGalNAc-T6 and ST6GalNAc-I are upregulated in the serum of gastric cancer patients[31]. These studies also identified POFUT1, GALNT5, and C1GALT1 biomarkers in gastric cancer[32−34]. Glycosyltransferase 8 domain-containing protein, a family member of glycosyltransferases, was initially identified as a risk indicator of schizophrenia[5]. Literature investigating GLT8D1 and cancer remains limited. Only five studies found that GLT8D1 correlated with the progression of malignancies[8−12]. Moreover, the prognostic value and biological efficacy of GLT8D1 in gastric cancer have also not been elucidated. This study explored the expression of GLT8D1 in pan-cancers, especially in gastric cancer. High levels of GLT8D1 expression were shown to be an independent risk factor for poor overall survival in patients with gastric cancer, as determined by analysis of the TCGA-STAD dataset. These findings were validated in the FAHSYSU's gastric cancer patients. Gastric cancer patients with an increased expression of GLT8D1 had a poor prognosis. The study revealed that gastric cancer patients with high levels of GLT8D1 have a more advanced grade and tumor stage than those with low GLT8D1 expression.
In addition, a prognostic nomogram was developed by combining GLT8D1 expression with age, T stage, N stage, M stage, and TNM stage to exhibit the intended performance accurately. The C-index and calibration curve results confirmed that the nomogram exhibited robust predictive performance. External validation of GSE62254 and FAHSYSU cohorts showed the favorable performance of the prognostic nomogram. The nomogram helped clinicians predict the death and survival probabilities of patients, which assisted them in making therapeutic decisions. Collectively, these findings confirmed that GLT8D1 has clinical significance in making diagnostic decisions and evaluating prognoses for gastric cancer patients.
Previous studies have not fully elucidated the biological function of GLT8D1. As a result, there is limited information regarding the biological role and mechanism of GLT8D1. The bioinformatics analysis conducted in this study showed that the overexpressed GLT8D1 is involved in cytoskeleton-related cellular components, epidermal-related biological processes, and peptidase regulator activity. On the other hand, the GSEA results showed that high GLT8D1 expression correlates with cancer-related signaling pathways, especially the STAT3 signaling pathway. These findings confirmed that GLT8D1 supports its oncogenic role in gastric cancer. Ilina et al.[10] reported that GLT8D1 promotes the migration of glioma cells in vitro because of interactions with cytoskeleton-related proteins. In another study, Liu et al.[12] found that GLT8D1 is upregulated in glioma, promoting glioma stem cell maintenance and cell proliferation by N-glycosylation of CD133. The results in this study were in accord with the findings of Liu et al.'s study. This study also found that GLT8D1 promotes the progression of gastric cancer and migration of gastric cancer cells, and depletion of GLT8D1 does not inhibit the proliferation of gastric cancer cells. We hypothesize that this discrepancy was because GLT8D1 exerts a comprehensive function in different malignancies. This study also revealed that the STAT3 signaling pathway and glycosylation may be regulated by the expression of GLT8D1 in gastric cancer. For example, the glycosylated PD-L2 complex combined with EGFR can activate the STAT3 signaling pathway, resulting in immune evasion of cancer cells in cancer immunotherapy[35]. In addition, studies have also shown that a glycosylated mutant of PDGFA—PDGFA Y288C—can activate the STAT3 signaling pathway[36]. Therefore, these results may indicate that GLT8D1may contribute to activating the STAT3 signaling pathway through glycosylation modification.
The GLT8D1-related PPI network and gene–gene interactions were also analyzed using online databases, as shown in Fig. 2. The results of PPI network analysis revealed that GLT8D1 is capable of constructing interactions with other proteins, such as SUCLG2, SCAMP3, and ZSCAN31. Many studies have stated that ZSCCAN31 is a potential therapeutic target in hepatocellular carcinoma[37]. It has been reported that EGFR-LIFR signaling induces SUCLG2, which promotes neuroendocrine differentiation and glycolysis in prostate cancer[38]. Elsewhere, it was found that SCAMP3 is associated with the progression of breast cancer[39] and glioma[40]. The gene–gene interactions showed that GLT8D1 has multiple interactions with 20 genes, most of which are involved in glycosylation, including UGGT1, GXYLT1, and GLT8D2, as shown in Fig. 2d. Therefore, these findings suggest that GLT8D1 and these proteins or genes form complex interactions, thereby regulating the development of gastric cancer by glycosylation.
Although GLT8D1's potential significance in gastric cancer was investigated in this work, this study has several limitations. First, a limited number of patients were included in the clinical sample, which could have affected the reliability and validity of the results. Therefore, a larger cohort should be used in future studies. Second, more experiments should be performed to clarify the regulatory mechanism of GLT8D1 in gastric cancer.
In summary, GLT8D1 is a biomarker that may predict whether or not a patient would have a poor clinical outcome from gastric cancer. Moreover, according to the findings of the research, GLT8D1 expression was significantly upregulated in gastric cancer tissue compared to adjacent normal tissues that were matched. In addition, the study showed that the biological functions of GLT8D1, including glycosylation, can participate in gastric cancer. The study also found that GLT8D1 may contribute to regulating the migration of gastric cancer cells via the IL6/STAT3 signal pathway. GLT8D1 may become an effective biomarker to predict the prognosis of gastric cancer patients. Therefore, this study recommends that further investigations are warranted to uncover the regulatory mechanism of GLT8D1 in gastric cancer.
-
GLT8D1 is a prognostic biomarker that suggests an unfavorable prognosis for gastric cancer. GLT8D1 promotes gastric cancer cell migration and invasion, which is associated with activating the IL6/STAT3 signaling pathway.
-
The clinical specimens used in this study were reviewed and approved by the Medical Committee Ethics of the Seventh Affiliated Hospital of Sun Yat-sen University (No. KY-2020-024-01). In addition, the patients who participated in this study had signed an informed consent.
-
The authors confirm their contributions to the paper as follows: conception and experimentation: Chen S, Yu H, Liu Z, Huang Y, Feng Y; collected database data: Hong C; bioinformatic and statistical analysis: Chen S, Yu H, Liu Z; revision of manuscript: Zhang C, Sun C, Jiang J; study and manuscript supervision: Sun A, Jiang J. All authors reviewed the results and approved the final version of the manuscript.
-
Public datasets were described in this study. TCGA datasets supported our conclusion in this article and can be accessed via www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga. GEO datasets (GSE62254, GSE206329) can be downloaded at www.ncbi.nlm.nih.gov/geo/query/acc.cgi.
-
This study was funded by the Guangdong Provincial Key Laboratory of Digestive Cancer Research (No. 2021B1212040006), Shenzhen Science and Technology Program (No. JCYJ20230807110718037 and JCYJ20230807110411024), Scientific and Technological Innovation Capacity Building Project of Beijing Academy of Agricultural and Forestry Sciences (No. KJCX20230426), Medical Scientific Research Foundation of Guangdong Province, China (No. A2025131), and the Sanming Project of Medicine in Shenzhen (No. SZSM201911010).
-
The authors declare that they have no financial or other conflicts of interest.
-
accompanies this paper online at: https://doi.org/10.48130/git-0026-0008.
-
# Authors contributed equally: Songyao Chen, Hong Yu, Zhichao Liu
- Supplementary Fig. S1 Decision curve analysis of the prognostic nomogram in GSE62254 dataset.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen S, Yu H, Liu Z, Huang Y, Feng Y, et al. 2026. GLT8D1 promotes gastric cancer invasion and migration, which is associated with activation of the IL6/STAT3 signaling pathway. Gastrointestinal Tumors 13: e010 doi: 10.48130/git-0026-0008 

GLT8D1 promotes gastric cancer invasion and migration, which is associated with activation of the IL6/STAT3 signaling pathway
- Received: 22 September 2025
- Revised: 16 March 2026
- Accepted: 25 March 2026
- Published online: 12 June 2026
Abstract: Gastric cancer, one of the deadliest gastrointestinal tumors, has a high incidence rate worldwide. Many studies show that the glycosyltransferase 8 domain-containing protein 1 (GLT8D1) plays a crucial role in protein glycosylation. However, few scholars have explored the prognostic importance of GLT8D1 in gastric cancer. The purpose of this research is to investigate the clinical value and biological function of GLT8D1 by examining multiple databases and performing relevant experiments. This study obtained the transcriptomic information and clinically relevant data from the cancer genome atlas (TCGA) and evaluated GLT8D1 in pan-cancer and gastric cancer. The prognostic role of GLT8D1 in the TCGA-stomach adenocarcinoma (STAD) dataset and gastric cancer patients was explored. A predictive nomogram was developed based on GLT8D1 mRNA expression level and clinical features. The receiver operating characteristic curve (ROC) values of predicting overall survival probabilities in the training set (TCGA-STAD dataset) and external validation sets (GSE62254 and FAHSYSU's cohort) were investigated to evaluate the performance of the prognostic nomogram. The STRING database and GeneMANIA were used to describe the proteins and genes interacting with GLT8D1. Kyoto Encyclopedia of Genes and Genomes (KEGG), Gene ontology (GO), and gene set enrichment analysis (GSEA) were used to analyze the biological function of GLT8D1 based on the TCGA-STAD dataset. The study also performed in vitro assays to evaluate the proliferation, invasion, and migration abilities of GLT8D1 in gastric cancer cells. The results found that GLT8D1 is upregulated in gastric cancer. In addition, according to the study, those with stomach cancer who had increased GLT8D1 expression had decreased overall survival (OS). Moreover, GLT8D1 was identified as a separate risk factor for OS in gastric cancer by the univariate and multivariate analyses. Nomogram evaluation revealed that the predictive model was reliable for estimating the probability of overall survival over 1, 3, and 5 years. Functional enrichment analysis showed that interferon-γ response, mTORC1 signaling pathway, glycolysis, and IL6-JAK-STAT3 signaling pathways were enriched in patients with high GLT8D1 expression levels. Our in vitro assay experiment revealed that GLT8D1 promotes gastric cancer cell invasion and migration, which suggests the involvement of activating the IL6/STAT3 signaling pathway. The study concludes that GLT8D1 is a prognostic biomarker that suggests an unfavorable prognosis for gastric cancer.
-
Key words:
- GLT8D1 /
- Gastric cancer /
- Prognosis /
- Overall survival /
- IL6/STAT3 signaling pathway