









-
Due to global climate change and environmental pollution, abiotic factors such as drought, salinity, and high temperature drastically hinder the yield, quality, and geographic distribution of economical crops worldwide[1]. Perceiving stress signals and adaptation to adverse environmental conditions are fundamental biological functions associated with plants[2,3]. Numerous studies have proven that plant transcription factors play a crucial role under abiotic and biotic stresses by acting as important regulators of stress responsive genes and potentially improving crop yield[4−6]. Specific TF families such as NAC[7,8], MYB[9,10], and bHLH[11,12] perform a critical role as regulators of plant responses to various abiotic stresses such as heat, cold, drought, and salt[13]. TFs interact with the cis-regulatory elements like promoter regions of stress-related genes, hence serve as a molecular switch in regulating the transcriptional output of their target genes[14]. For instance, GhbHLH18 from Gossypium hirsutum regulates the expression of GhPER8, OsbHLH062 from Oryza sativa mediates the expression of OsHAK21, whereas CsbHLH18 binds to the CsPOD promoter and regulates its expression in Citrus sinensis[15−17].
Basic-helix-loop-helix (bHLHs) proteins belong to a key family of transcription factors widely dispersed across the plants, fungi, and animal kingdoms[18−20]. The first plant in which bHLHs TF were found to regulate plant responses was maize (Zea mays L.)[21]. Since then, numerous bHLH TFs have been reported in different plant species, mediating responses under unfavorable environmental conditions[22,23]. For example, the bHLH TF 'MsbHLH115' from Medicago sativa mediates the cadmium stress tolerance[23], AhbHLH112 regulates drought tolerance mechanism in peanuts[21], and AtbHLH92 enhances the salt and osmotic stress tolerance in Arabidopsis thaliana[24]. As an illustration of the size of the bHLHs family in various plant species, 164 bHLH TFs have been found to be involved in Arabidopsis (Arabidopsis thaliana L.), 180 in rice (Oryza sativa L.), 190 in tobacco (Nicotiana tabacum L.), 102 in walnut (Juglans regia L.), 191 in grapes (Vitis vinifera L.), 85 in Ginkgo biloba (Ginkgo biloba L.), 440 in rapeseed (Brassica napus L.), 268 in wild cabbage (Brassica oleracea L.) and 251 in mustard (Brassica rapa L.), respectively[25]. The TF bHLHs are structurally divided into two specified regions, the first one is 'basic' and the second is the helix region (HLH). Both regions are highly specific in terms of their function, such as DNA binding and protein formation[13]. The specific domain of bHLHs consists of 60 amino acids, where both DNA-binding and protein regions are separated by the loop[26]. The bHLH transcription factor comprises the N-terminal basic region, which is directly followed by the HLH (helix-loop-helix) domain[26]. More than 50% of bHLHs that have been reported in plants so far possess a highly conserved HER motif (His5-Glu9-Arg13) to achieve DNA-binding and transcriptional regulation of their target genes[27].
The bHLH transcription factors have been reported to play vital roles in growth and development as well as abiotic stress tolerance of plants[25,28,29]. Previous studies have reported the mediatory role of these TFs in different plant species subjected to environmental hazards, such as drought[30], salinity[25], low temperature[25], heavy metal toxicity[31], and osmotic stress[32]. Moreover, bHLH TFs also contribute significantly to plant developmental functions, including iron homeostasis[21], flower and fruit development[33], stomatal initiation[34], root vascular cell proliferation[35], grain yield[33], and secondary metabolites biosynthesis, such as anthocyanin[36].
Pearl millet (Pennisetum glaucum L.), with a diploid genome (2n = 2x = 14), is an important warm-season (C4) grass species of the Panicoideae subfamily, exhibiting superior vitality under various environmental conditions[37,38]. It is highly palatable, easily digestible, and nutritionally rich, thereby contributing significantly to the livestock, food, and beverage industries[39]. In contrast to other cereal crops, it has a higher rate of photosynthetic capacity and biomass production, thus making it a highly valuable crop suitable for resource-limited and under-developed regions worldwide[40]. The crop covers over 31 million hectares of land globally and is cultivated in more than 30 countries across arid, semi-arid, tropical, and sub-tropical regions of Asia, Africa, and Latin America[41]. Pearl millet is a cross-pollinating crop with a genome size of 1.79 Gb[42] and has been shown to exhibit enhanced tolerance against abiotic stresses[43]. It is domesticated in regions with low soil fertility[44], drought[45], and heat stress[46,47], therefore making it naturally adapted to cope with the challenges associated with climate change[48,49]. Pearl millet exhibits strong adaptability to environmental resilience and possesses excellent potential for genetic improvement through breeding techniques[50,51]. The present study focuses on the bHLH TF family in pearl millet because other TFs gene families, such as NAC, MYB, WRKY, and bZIP, have been reported to play a role in defining stress responses in this crop[52−55].
Many studies have reported the involvement of the bHLH TFs family in various plant species such as Arabidopsis thaliana[56], Oryza sativa[57], Phaseolus vulgaris[58], Solanum tuberosum[59], and Malus domestica[60]. Notably, the role of bHLH TFs in pearl millet, an important cereal crop worldwide, remains largely unexplored. In the face of increasing environmental stresses such as drought, salinity, and heat stress, there is an urgent need to develop more resilient pearl millet cultivars. However, the bHLH transcription factor family, which plays key roles in plant development and abiotic stress responses, has not yet been characterized in this crop. By performing an in silico analysis to identify bHLH orthologous and paralogous groups and examining their expression under drought, salt, and heat stress conditions, this study aimed to uncover candidate genes that could be harnessed to improve pearl millet's tolerance to these stresses, ultimately contributing to the development of stress-resilient varieties. A genome-wide analysis of predicted 110 PgbHLH transcription factor-encoding genes was conducted, revealing both evolutionary conservation and divergence of this TF family through phylogenetic analysis, chromosomal localization, gene structure, conserved motifs, and collinearity analysis. Moreover, putative cis-regulatory element analysis highlighted an enrichment of stress-responsive elements. Furthermore, the study also demonstrated the in silico expression profiling of the PgbHLH genes in roots and leaves of pearl millet under various abiotic stresses. This study provides a valuable resource for future functional validation studies aimed at improving abiotic stress resilience in pearl millet through molecular breeding.
-
The bHLH genes in pearl millet (PgbHLH) were identified using the HMMER software (
http://hmmer.org ) with the hidden Markov model (HMM) having profile PF00010 (bHLH domain) from Pfam (http://pfam.xfam.org/ )[61]. The nucleotide as well as protein sequences of pearl millet bHLH (PgbHLH) genes were retrieved from the millet database 'Milletdb' (http://milletdb.novogene.com )[62]. For evolutionary analysis, protein sequences of Arabidopsis thaliana and Oryza sativa were obtained from 'The Arabidopsis Information Resource TAIR' (www.arabidopsis.org ) (Supplementary Table S1) and 'The Rice Genome Annotation Project' (https://rice.uga.edu ), respectively (Supplementary Table S2). A total of 110 PgbHLH genes were renamed according to their position on the pearl millet chromosomes as PgbHLH0.1 to PgbHLH7.9. Furthermore, the important protein features, including amino acid length, isoelectric points, and molecular weight, were calculated by using the ExPASy tools (www.expasy.org )[63].Gene structure and conserved motif analysis
-
To analyze the gene structures and conserved motifs of PgbHLH genes, several bioinformatics tools were utilized. For gene structure analysis, the genome and coding DNA (CDS) sequences of the identified PgbHLHs genes were used as a query for the exon-intron illustration in the Gene Structure Display Server (GSDS 2.0), online software (
http://gsds.cbi.pku.edu.cn )[64]. Multiple alignment of the retrieved PgbHLH genes was performed using the CLUSTALW (www.genome.jp/tools-bin/clustalw )[65] with default parameters. For conserved motif analysis, conserved domains were analyzed and displayed using UniGene[66]. The MEME software ( https://meme-suite.org/meme/doc/ame.html )[67] was used to identify conserved motifs in the PgbHLHs using amino acid sequences. The analysis parameters were set as follows: maximum number of motifs (20) and minimum sites per motif (two).Chromosomal mapping and collinearity analysis
-
To locate the positions of bHLH genes, the gene names were used as queries in the millet database 'Milletdb' (
http://milletdb.novogene.com )[62]. The genes were then mapped onto pearl millet's chromosomes using the TB-tools online platform (https://github.com/CJ-Chen/TBtools )[68]. For the collinearity analysis, whole-genome sequence and annotation files for Setaria italica (Foxtail Millet), Oryza sativa (Rice), Solanum lycopersicum (Tomato), Zea mays (Maize), and Arabidopsis thaliana (Thale Cress) were obtained from the Phytozome v13.0 (http://phytozome-next.jgi.doe.gov).For Pennisetum glaucum (pearl millet), annotation and genomic sequences were downloaded directly from the Milletdb database (http://milletdb.novogene.com )[62]. The genomic sequences and annotation files for all species were then uploaded to the MCScanX tool (https://github.com/wyp1125/MCScanX )[69] to generate a collinearity map. Subsequently, a pairwise synteny plot (dual synteny) was created using the TB-tools to identify the homology of Pennisetum glaucum (Pearl millet) bHLH transcription factors (PgbHLHs) with those in the other selected species.Evolutionary phylogenetic analysis of PgbHLH TFs
-
For the evolutionary phylogenetic analysis, Clustal Omega (
www.ebi.ac.uk/jdispatcher/msa/clustalo )[70] was used to align the protein sequences of Pennisetum glaucum (110 PgbHLH proteins) with bHLH proteins from two model species, Arabidopsis thaliana (145 AtbHLH proteins) and Oryza sativa (139 OsbHLH proteins). The neighbor-joining method with 1,000 bootstrap replicates was utilized to infer the evolutionary history. During the pairwise and multiple alignment, no sequences were deleted or excluded, and no gap positions were changed. All alignment parameters were set to their default values. This analysis resulted in the generation of an unrooted circular phylogenetic tree.Putative cis-regulatory element (CREs) analysis
-
Predicting the possible physiological functions of PgbHLH genes, the 1 Kb upstream genomic sequences from the translational start site (ATG) of each gene were directly retrieved, using the database NCBI (
www.ncbi.nlm.nih.gov ) (Supplementary Table S3). The extracted 1Kb upstream region of 110 PgbHLHs was then submitted to a promoter identified database PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html )[71], which identified 30 distinct putative cis-acting regulatory elements (CREs) of PgbHLH genes.In silico expression analysis
-
For the investigation of the expression profile of PgbHLH genes, the fragments per Kilobase of transcript per million mapped reads (FPKM) expression pattern data were directly downloaded from millet data base (
http://milletdb.novogene.com )[62] from the gene expression tool under different stresses such as heat stress, drought stress, and salt stress compared with the control (CK) for both leaves and root tissues (Supplementary Table S4). First, the datasets were aligned and rearranged, then 110 PgbHLH genes were selected for the heat map visualization. Heat maps were generated using the R packages pheatmap (https://github.com/raivokolde/pheatmap ) and tidyerse (www.tidyverse.org ). -
A total of 110 PgbHLH genes were identified and renamed as PgbHLH0.1 to PgbHLH7.9 based on their positions on pearl millet chromosomes (Supplementary Table S5). Different properties of the PgbHLHs were identified, such as their amino acid length, molecular weight, and iso-electric points. The length of the PgbHLH proteins ranged from 102 amino acids (PgbHLH1.2) to 814 amino acids (PgbHLH3.4). The molecular weight of the PgbHLH proteins varied from 10.76 (PgbHLH1.2) to 89.27 kDa (PgbHLH3.4). Moreover, the isoelectric point of the PgbHLH proteins ranged from 4.81 (PgbHLH5.3) to 11.45 (PgbHLH2.2).
Chromosomal distribution of PgbHLH genes
-
To determine the existence of PgbHLH genes on their respective chromosomes, a diagrammatic map was constructed to show the identified PgbHLH genes on all seven chromosomes. All chromosomes have different lengths, varying from 179.32 Mb (chromosome 5) to 336.26 Mb (chromosome 3). Identified PgbHLH (110) genes were distributed on seven chromosomes unevenly, with the highest enrichment (almost 19 PgbHLH genes) on chromosome 2, followed by chromosomes 3, 5, and 6, each of which contained a total of 18 PgbHLH genes. Chromosome 1 contained almost 11 PgbHLH genes, while the least number of PgbHLH genes (nine) were present on chromosome 7. Hence, the disproportionate abundance of PgbHLH genes on chromosomes 2, 3, 5, and 6 indicated a suitable hotspot region for the distribution of the PgbHLH family members; moreover, the blue lines represented the paralogous or duplicated PgbHLH genes on the same or different chromosomes. These duplications suggested that some PgbHLH genes might have retained ancestral functions, while others could have undergone neofunctionalization or pseudogenization, contributing to functional divergence within the gene family. This pattern underscores the potential role of gene duplication events in expanding the repertoire of bHLH functions relevant to stress responses and developmental processes in pearl millet (Fig. 1).
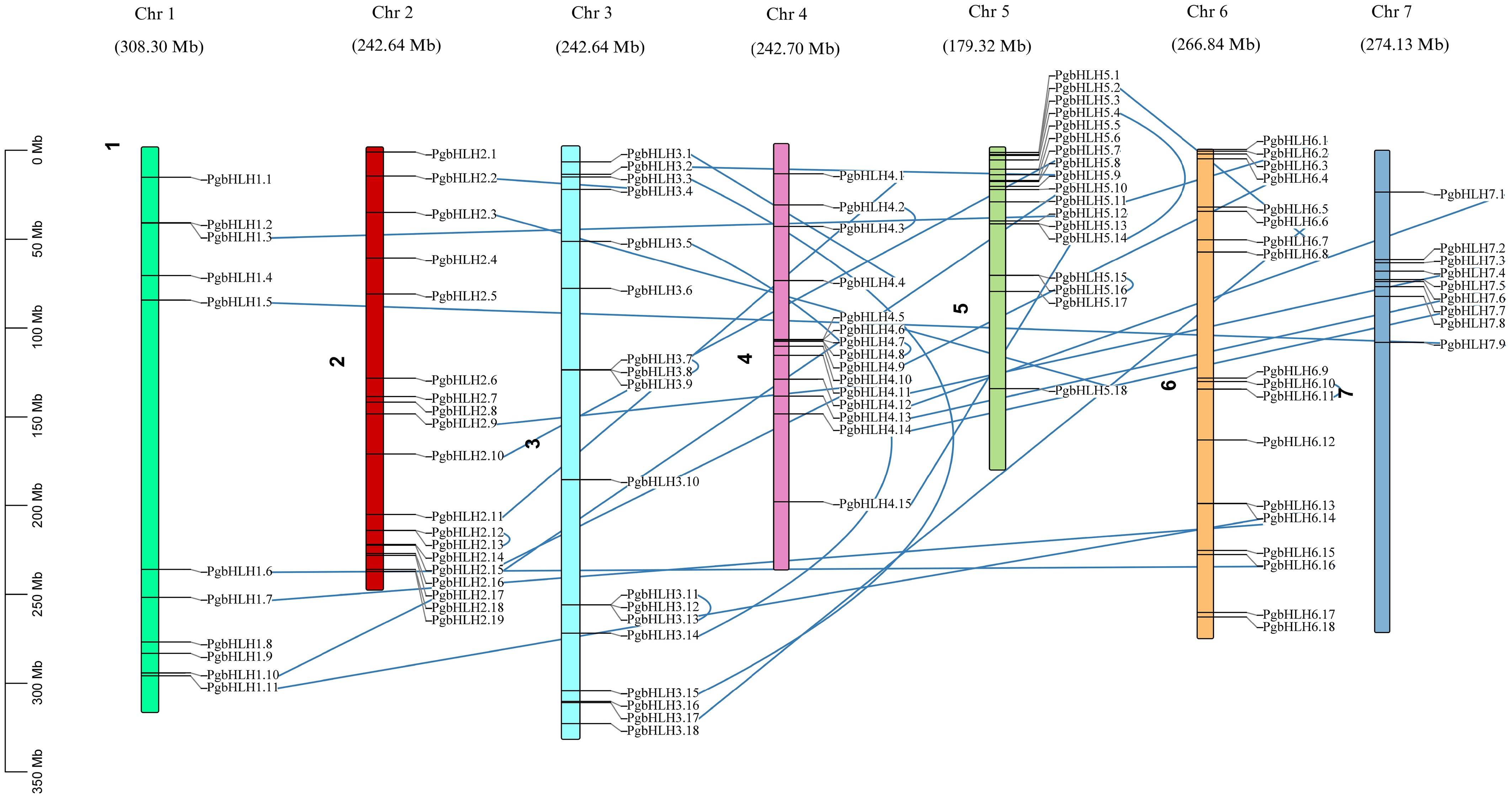
Figure 1.
Arrangement of 110 PgbHLH genes on the seven chromosomes of Pennisetum glaucum. Chromosomal distribution map represented as colored bars, with chromosomes showing different lengths in million base pairs (Mb). The PgbHLH gene distribution is based on the exact location on their respective chromosomes. The blue lines in the map indicate the paralogous genes, highlighting the presence of a gene duplication event within the pearl millet genome.
Conserved domain analysis of the PgbHLHs
-
The PgbHLH conserved domain analysis revealed the ability of DNA-binding and selection. Multiple sequence alignment of their amino acid sequences showed a partial conservation of the bHLH domain throughout all observed sequences (Fig. 2). The PgbHLH domain is generally around 40–60 amino acids; the analysis revealed conserved domain structure of the putative PgbHLH transcription factor, with distinct features at the N and C-terminal regions. Analysis of the N-terminus region revealed a consensus sequence characterized by partially conserved amino acid residues, such as essential amino acid leucine (L) and semi-essential amino acid arginine (R). Moreover, the C-terminal region also contained partial conservation of amino acid sequences, including lysine (K), alanine (A), tyrosine (Y), and valine (V). Moreover, the aligned PgbHLH domains showed other slightly conserved residues, including acidic amino acids glutamic acid (E) and aspartic acid (D), as well as essential amino acid isoleucine (I) and non-essential amino acid proline (P). The study's analysis revealed that E-box (CANNTG) binding sites were also partially conserved among PgbHLHs, although variations in neighboring residues might influence binding specificity.

Figure 2.
Multiple sequence alignment of pearl millet PgbHLH TFs. Alignment was created by using the online program CLUSTAL W and displayed using UniGene. The conserved amino acid sequences are shown in different colors. The sequence logo illustrates the conservation of the domain among all 110 PgbHLH genes.
Phylogenetic distribution and protein motif analysis of PgbHLHs
-
To determine the existing relationship among 110 PgbHLHs, the neighbor-joining (NJ) method was used to construct the phylogenetic tree (Fig. 3a). The tree was further classified into 12 subgroups (clusters) named cluster-I to cluster-XII. The largest cluster was found to have 15 and 16 PgbHLH genes named as cluster-VII and cluster-X, followed by the two clusters I and V that contain 14 and 12 PgbHLH genes, respectively. Moreover, 20 conserved motifs of the PgbHLH genes were identified (Fig. 3c). By identifying the motifs, paralogs were found within the clusters showing the same motif arrangements belonging to the same phylogenetic group, with only a slight difference being shown within the groups. Based on the arrangements, cluster-VII and cluster-IX contained eight and seven motifs, respectively, followed by motif 6 within cluster-X. The clusters I, II, III, IV, VIII, and XII contained four conserved motifs, and clusters V, VI, and XI also comprised the same three conserved motifs. The motif number 1, 2, 3, 4, 5, 14, and 15 showed their major presence within the clusters (Fig. 3b). This phylogeny and protein motif analysis revealed that the genes within the same group shared similar motif numbers and patterns, highlighting their evolutionary relationships and supporting their classification.

Figure 3.
Phylogenetic distribution and protein motif analysis of PgbHLH in pearl millet. (a) Phylogenetic tree is classified into 12 distinct clusters (cluster-I to cluster-XII). All the clusters are shown in different colors. (b) The conserved motifs are arranged according to phylogenetic relationships. (c) The legends list contains 20 different motifs, represented by different colors, allowing the identification of specific patterns.
Gene structural diversity of PgbHLHs
-
To analyze the diversity among the structure of 110 PgbHLHs genes, the 'Gene Structure Display Server' was used. The analysis revealed that arrangements of PgbHLH genes among the various subgroups (clusters) illustrated the differences among gene structures within these clusters. Gene length was managed according to the range of 0–26 Kb. The organization of the intron and exon in the gene sequence played an important role by providing the evolutionary hint representing the characteristics of the same gene family. The intron calculated ranged from about one to ten. Moreover, about 12.72% (14) PgbHLHs did not contain any intron, while 6.36% (seven) PgbHLHs contained only one intron. The maximum numbers of introns (nine) were usually observed in PgbHLH2.15 from cluster-I and PgbHLH3.4 from cluster-XII, showing its abundance among all 110 PgbHLH genes. The results revealed the remarkable degree of structural heterogeneity among the PgbHLHs genes in pearl millet, indicating the high level of genetic diversity within the bHLH TF family (Fig. 4).

Figure 4.
Gene structure representing the exon-intron structure of the PgbHLHs genes differentiated by the subgroups (cluster-I to cluster-XII). Gene structure display server was used to perform the analysis. CDS shown in red color (exons), dark black lines represent the introns, and dark blue color boxes depict the upstream/downstream regions. Moreover, the x-axis at the bottom provides the scale bar representing the length of the PgbHLH genes. The scale is marked with a numerical value indicating the size in Kilobase pairs (Kb).
Evolutionary analysis of bHLHs among pearl millet, Arabidopsis and rice
-
To analyze an evolutionary relationship of PgbHLH genes among various other species of plants, a multi-species neighbor-joining phylogenetic tree was constructed, comprising all bHLH genes from three plant species, Pennisetum glaucum (110 PgbHLHs), Arabidopsis thaliana (145 AtbHLH genes) (Supplementary Table S1), and Oryza sativa (139 OsbHLH genes) (Supplementary Table S2). The tree's structure and classification followed the method of Gabriela & Pires taken from[72]. Of all the bHLH genes present within the tree, 394 genes were subdivided into 15 subgroups and one orphan group (UC). All species were differentiated by different symbols, where circles represent Pennisetum glaucum, triangles represent Arabidopsis thaliana, and squares represent Oryza Sativa. The unclassified group (UC) contained one AtbHLH (AtbHLH111) and one OsbHLH (LOC Os06g50910.3) gene. Clades were designated as bHLH-I (blue color) to bHLH-XV (purple color), respectively. The bHLH-II clade was designated as the major clade, having 70 PgbHLH members, followed by PgbHLH-IV with 43 PgbHLH genes. Analysis showed that four clades (bHLH-V, bHLH-VI, bHLH-VII, and bHLH-XI) contained one ortholog pair between Pennisetum glaucum and Oryza sativa with genes PgbHLH3.10 and LOC Os02g23823.1, PgbHLH1.2 with LOC Os03g07540.1, PgbHLH2.2 and LOC Os05g06520.1, and PgbHLH7.8 with LOC Os09g28210.1, respectively. Only two clades showed ortholog pairs between Pennisetum glaucum and Arabidopsis thaliana: bHLH-I (PgbHLH6.7 with AtbHLH152) and bHLH-VII (PgbHLH2.16 and AtbHLH110, PgbHLH3.4 with AtbHLH117), which might suggest a distant evolutionary relationship (Fig. 5). The largest clades, bHLH-X (ten) and bHLH-IV (nine), formed ortholog pairs between Pennisetum glaucum and Oryza sativa, which may indicate a closer evolutionary relationship between these two species. No ortholog pairs were found in the clades bHLH-VIII and bHLH-XII among all three species (Fig. 5). Analysis revealed that PgbHLH contained the maximum number of orthologous gene pairs with O. sativa, suggesting that these proteins might have been conserved during the long-term evolution of the P. glaucum.
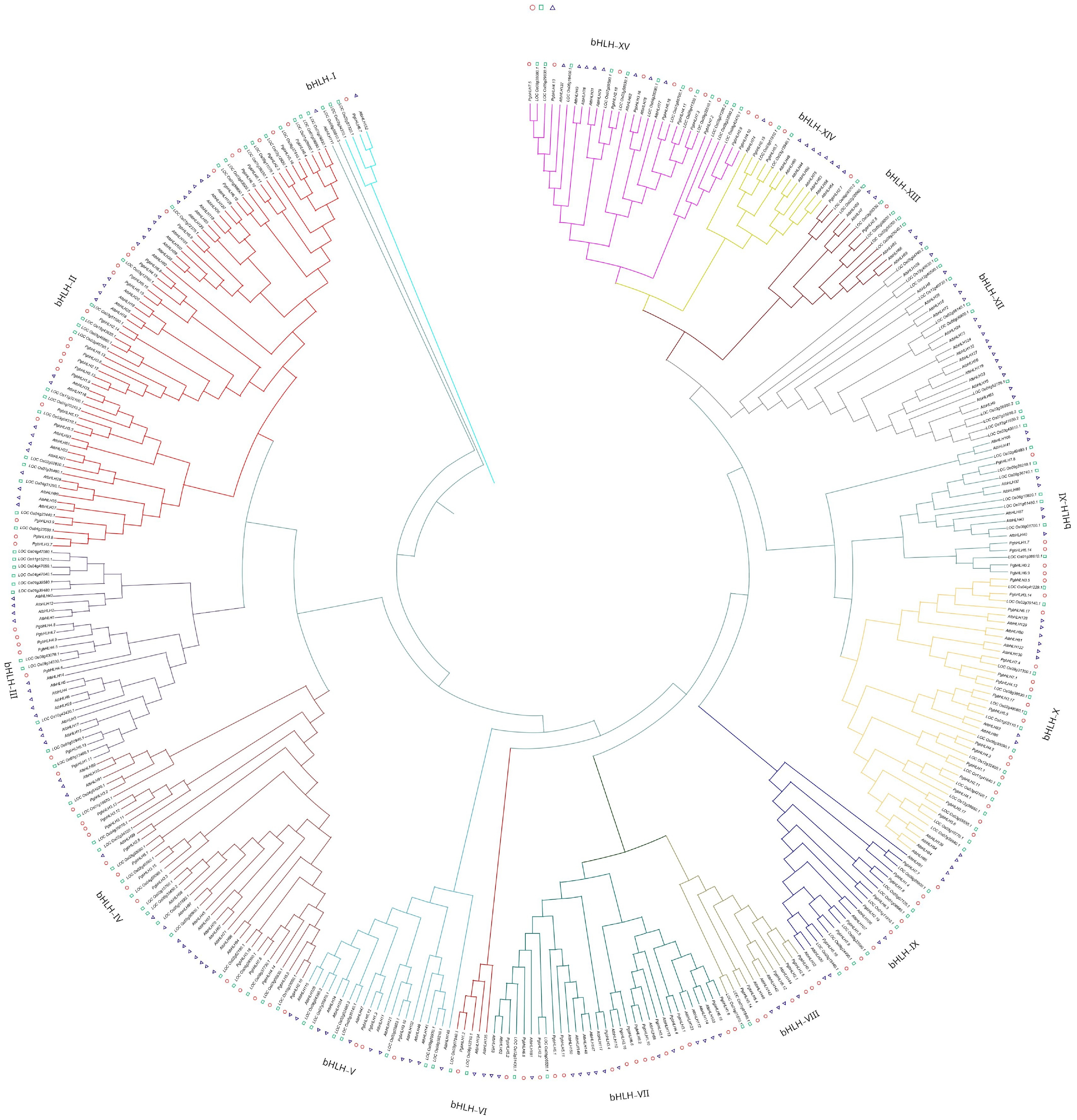
Figure 5.
Multi-species neighbor-joining phylogenetic tree was constructed using Clustal Omega, 1,000 bootstrap replicates to evaluate the validity of the tree, containing bHLH genes of Pennisetum glaucum (110 PgbHLH), Oryza sativa (139 OsbHLH), and Arabidopsis thaliana (145 AtbHLH). All of the bHLH genes were classified into 15 subgroups (bHLH-I to bHLH-XV) highlighted by the different colors in the tree. Circles represent Pennisetum glaucum, triangles represent Arabidopsis thaliana, and squares represent Oryza Sativa.
Collinearity of bHLH genes between pearl millet and other plant species
-
Comprehensive syntenic analysis was performed to define the collinearity relationship of PgbHLH with five representative species, such as Arabidopsis thaliana, Solanum lycopersicum, Setaria italica, Oryza sativa, and Zea mays. Dual synteny plot MCScanX (
https://github.com/wyp1125/MCScanX )[69] was used for this analysis (Fig. 6). These species included two dicotyledonous species (Arabidopsis thaliana, Solanum lycopersicum) and three monocotyledonous species (Setaria italica, Oryza sativa, and Zea mays). One hundred and ten PgbHLH genes displayed syntenic relationships; pearl millet showed 460 orthologous pairs with all mono and dicotyledonous species (Supplementary Table S6). Zea mays showed the highest collinearity among all the species, with 150 (33%) collinear pairs, followed by Setaria italica with 137 (30%) collinear pairs and Oryza sativa with 125 (27%) collinear pairs. Similarly, Solanum lycopersicum had 38 (9%) collinear gene pairs, and the least pairs were found between A. thaliana and P. glaucum with only ten (3%) collinear genes (Supplementary Table S6). Interestingly, P. glaucum formed strong collinearity with the monocotyledonous species (Zea mays, Setaria italica, and Oryza sativa), but had the least collinearity with the dicotyledonous species (Solanum lycopersicum and Arabidopsis thaliana) (Supplementary Table S6, Fig. 6). Moreover, the collinearity analysis of PgbHLH genes with these plant species suggested that genes present within homologous regions might be inherited from the monocotyledonous origins.
Figure 6.
Collinearity analysis of bHLH genes between Pennisetum glaucum, Setaria italica, Oryza sativa, Zea mays, Solanum lycopersicum, and Arabidopsis thaliana. The green color shows the chromosomes of pearl millet, and their number is mentioned above the boxes. Different colored boxes show the different chromosomes within each species. Red colored lines show the orthologous pairs between the species, and grey lines represent collinearity blocks.
Putative promoter cis-acting regulatory elements of PgbHLHs
-
Analysis of cis-acting elements in the PgbHLH gene promoters revealed the distribution and various types of regulatory elements present in their promoter regions. Each horizontal line represents a domain name having the putative cis-acting elements depicted as different colored boxes along these lines. For this analysis, 1 Kb upstream promoter regions from the genomic DNA of the PgbHLH genes were searched from the NCBI (
www.ncbi.nlm.nih.gov ) (Supplementary Table S3), and then submitted to the online database PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )[71] (Fig. 7). In the PgbHLH genes' promoters' regions, putative CREs were found to be responsible for the root, seed (RY-E), meristem, and endosperm development. Notably, defense and stress-responsive elements crucial for abiotic responses, such as low temperature and TC-rich cold-responsive element, high light intensity responsive elements (G-box), light responsive elements (LRE), for biotic stress (fungal and bacterial responsive elements, (W-box). Hormone-responsive-elements, gibberellin responsive element, auxin-responsive element, methyl jasmonic acid responsive element, abscisic acid responsive element, salicylic acid responsive element. Putative promoter, CREs responsible for plant physiological and developmental processes such as circadian cycle, cell cycle regulation (CCR), meristem response element (MRE), unfolded protein response elements, meristem maintenance, and stem cell regulation responsive elements were identified (Supplementary Table S3). Moreover, the PgbHLH3.7 and PgbHLH4.10 contained the maximum number of putative CREs (20), while PgbHLH2.14 had the least number (five). The proportion of promoter putative cis-elements identified in this study was as follows: hormone-responsive elements (17%), and physiological and plant developmental process response elements (24%). The majority of the promoter putative CREs were defense and stress-response elements, comprising 34% (Fig. 7), indicating that bHLH genes might be involved in numerous abiotic and biotic stress responses, thereby improving stress resilience in pearl millet.
Figure 7.
The promoter putative CREs of the PgbHLH, gene which is represented by different colored boxes along the horizontal line, with the gene names listed on the left side of the figure. The top right corner represents the key that explains the different color boxes used to represent various types of putative CREs.
In silico expression analysis of PgbHLHs
-
To analyze the in silico expression of PgbHLH genes in leaves and roots of pearl millet, the expression datasets were downloaded from 'milletdb' (
http://milletdb.novogene.com )[62] and the expression of 110 PgbHLH genes were analyzed under three abiotic stresses (drought, salinity, and heat) at various time points (1, 3, 5, 7, 24, 48, 96, and 144 h) compared with the control (CK) (Supplementary Table S4). All 110 PgbHLH genes were selected for the heat map visualization (Fig. 8a–c). Expression levels of the 110 PgbHLH genes revealed their widespread expression across both tissues. Different colors indicate distinct patterns: red for increased expression and blue for decreased expression in response to varying stress levels (Fig. 8a–c). Analysis showed that five genes exhibited significant up-regulation under high temperature stress, including PgbHLH1.1 in root tissue at 96 and 144 h, and also 144 h in leaves tissue, PgbHLH5.1 in leaves at 1 h, PgbHLH1.2 in root tissue at 1 h, and PgbHLH6.4 in leaves at 1 h. Heat stress also induced the expression of PgbHLH4.1 in leaves at 5 and 96 h (Fig. 8a). Conversely, three genes showed down-regulation, including PgbHLH2.10 in the root at 24 and 48 h, PgbHLH3.8 in root at 24 and 96 h, and PgbHLH4.10 in leaves at 144 h and in root at 48 and 144 h.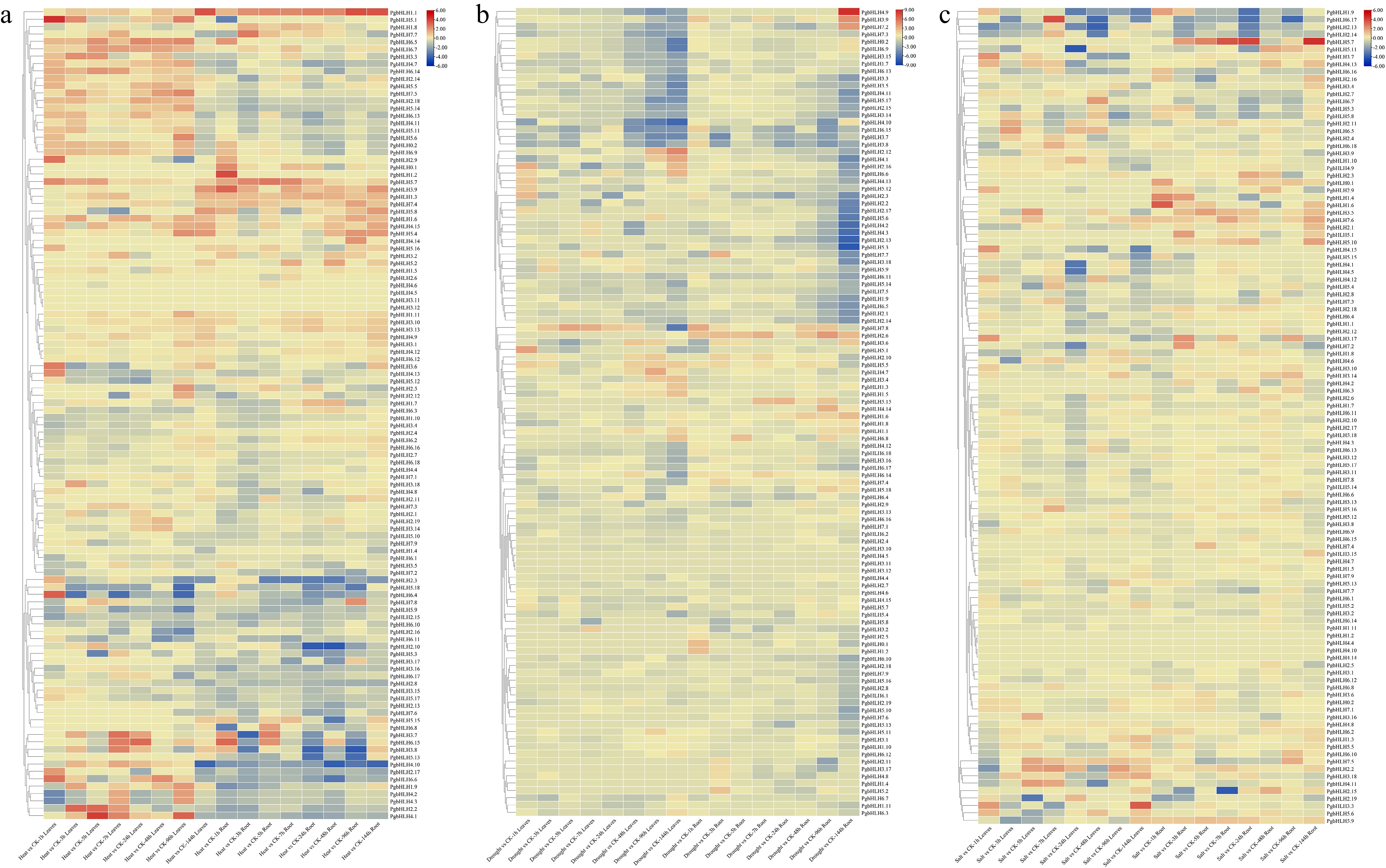
Figure 8.
Expression analysis of 110 PgbHLH genes at various time points (1, 3, 5, 7, 24, 48, 96, and 144 h) compared with control (CK), heat maps as per the k-means clustering. Clustering of PgbHLH genes in the rows under different abiotic stresses (a) heat stress, (b) drought stress, and (c) salinity stress. Scale on the upper right corner represents the value of expressions, which shows the value of decreased expression level in blue color and increased expression level in red color.
Under drought stress, one gene (PgbHLH4.9) showed significant up-regulation in the root at 144 h. However, three genes show down-regulation, including PgbHLH4.10 in leaves at 144 h, PgbHLH5.3 in root tissue at 144 h, and PgbHLH7.8 in leaves at 144 h. (Fig. 8b). Under salt stress, two genes showed significantly higher expression, including PgbHLH5.7 in the root at 24 and 144 h and PgbHLH3.3 in leaves at 144 h. In contrast, PgbHLH5.11 in leaves at 24 h and PgbHLH2.15 in roots at 7 h were significantly down-regulated (Fig. 8c).
Comparatively, the expression profile of PgbHLH genes under heat, drought, and salt stress revealed a unique pattern of up-regulation and down-regulation at various time points, compared to the control. In contrast to salt and drought stress, numerous PgbHLH genes showed more pronounced responses to heat stress, with notable up-regulation and down-regulation. Moreover, the expression profile of PgbHLH4.10 suggested its functional role in responding to both drought and high temperature stress (Supplementary Table S4).
-
Pearl millet (Pennisetum glaucum L.) is an important warm-season, C4-plant species of Panicoideae subfamily, cultivated extensively for food, feed, and biofuel production in arid and semi-arid regions worldwide[73]. Despite its potential, the genomic resources of pearl millet, such as transcription factor families regulating genes under abiotic conditions, remain unexplored. Among the key transcription factor families, bHLH has been reported to perform crucial physiological and biological functions in many plant species[57,74]. bHLH TFs have been found to have a significant impact on the development, adaptation, and evolution of various plant species, including apple[60], peanut[75], common bean[58], tomato[76], Brachypodium distachyan[18], Chinese cabbage[77], wheat[78], and Carthamus tinctorius[79].
Previous studies have shown the potential involvement of bHLH transcription factors in abiotic stress regulation in various plant species[26,33]. It was found that despite the high degree of variation among PgbHLH genes, they might exhibit notable conservation in their structural characteristics, particularly in the bHLH domain. Studying the bHLH gene family in multiple plant species and identifying its members can provide insights into understanding their putative roles in stress signal transduction[19]. In this study, 110 putative PgbHLH genes were identified from pearl millet, and PgbHLH0.1 to PgbHLH7.9 were designated, based on their specific positions on chromosomes 1 to 7, respectively (Fig. 1). This conservation suggests functional importance, and the variation may indicate functional diversity or specialization among the different PgbHLH genes.
Evolutionary phylogenetic analysis was performed based on the study of Riechmann et al.[80] to identify orthologous and paralogous relationships that offer functional insights into PgbHLH genes. According to the previous classification of Toledo-Ortiz et al. and Li et al.[27,57], a phylogenetic tree was constructed comprising 394 bHLH genes from Pennisetum glaucum (110 PgbHLHs), Arabidopsis thaliana (145 AtbHLH genes) (Supplementary Table S1), and Oryza sativa (139 OsbHLH genes) (Supplementary Table S2), which were subdivided into 15 subgroups (bHLH-I to bHLH-XV) and one unclassified group (orphan group) (Fig. 5). The unclassified group (UC) contained one AtbHLH (AtbHLH111) and one OsbHLH (LOC Os06g50910.3) gene. Most clades (bHLH-IV, bHLH-V, bHLH-VI, bHLH-VII, bHLH-X, bHLH-XI) contained ortholog pairs between Pennisetum glaucum and Oryza sativa, indicating a closer evolutionary relationship between these two species[57]. Only two clades were found to contain ortholog pairs between Pennisetum glaucum and Arabidopsis thaliana, which might suggest a distant evolutionary relationship[27] (Fig. 5). The largest clades, bHLH-IV and bHLH-X, contained the maximum number of orthologous gene pairs between Pennisetum glaucum and Oryza sativa, suggesting that these proteins might have been conserved during the long-term evolution of P. glaucum[27].
The genetic structure of the PgbHLH genes among the various subgroups (clusters) illustrated the differences among gene structures within these clusters. The findings of this study defined the classification of the subgroups; moreover, about 12.72% (14) PgbHLHs did not contain any intron, while 6.36% (seven) PgbHLHs contained only one intron, which might result in family expansion and evolution of new functions. Furthermore, the evolutionary development of the species might be due to the lack of some specific introns[81−83]. The analysis of exon–intron revealed that the lengths of the PgbHLH genes were mainly distributed within the range of 0–26 Kb, and the total number of introns ranged from one to ten. The maximum number of introns (nine) was usually observed in PgbHLH2.15 from cluster-I and PgbHLH3.4 from cluster-XII, showing its abundance among all 110 PgbHLH genes. The observed heterogeneity in terms of gene length, intron number, and exon number among PgbHLH genes indicated that the bHLH gene family has undergone a complex evolutionary process[84]. Moreover, genome contraction and expansion are largely influenced by repetitive elements located within intron regions, and gene function is determined by domain architecture[78].
The non-uniform dispersal of PgbHLH genes across the pearl millet (Pennisetum glaucum) genome, characterized by varying densities, might suggest that chromosomal rearrangements and duplications have likely occurred during the evolutionary history of this species, contributing to the dynamic reorganization of pearl millet's genome[85]. Hence, the disproportionate abundance of PgbHLH genes on chromosomes 2, 3, 5, and 6 indicated a suitable hotspot region for the distribution of the PgbHLH family members; moreover, the blue lines represented the paralogous or duplicated PgbHLH genes on the same or different chromosomes[86]. These duplications suggested that some PgbHLH genes might have retained ancestral functions, while others could have undergone neofunctionalization (new functions) or pseudogenization (lost the original functions), contributing to functional divergence within the gene family[87]. This pattern underscores the potential role of gene duplication events in expanding the repertoire of bHLH functions relevant to stress responses and developmental processes in pearl millet (Fig. 1)[87].
Phylogenetic and motif conservation analysis related to the previous findings of Zhou et al.[88] correlates with the study's investigation, where a diverse array of motif types among bHLH family members, with the canonical bHLH motif being ubiquitously present in all PgbHLH sequences. Moreover, the study of Qin et al.[89] found that the additional structurally conserved motifs were identified in a subset of PgbHLH sequences, suggesting potential roles under various abiotic and biotic stress conditions, which is in accordance with the present findings. A total of twenty (one to 20) conserved motifs were found. By identifying the motifs, putative paralogs were found within the clusters, showing that the same motif arrangements belong to the same phylogenetic group, with only slight differences being shown within the groups (Fig. 3b)[53]. These analyses revealed that E-box (CANNTG) binding sites were partially conserved among PgbHLHs, although variations in neighboring residues might influence binding specificity (Fig. 2)[53]. It has been found from this study that closely related genes within the same phylogenetic group (clade) might have identical motif arrangements. These clades showed less differentiation from each other, hence were named as putative paralogs to each other (Fig. 3a, b)[90].
Previous study on putative cis-acting regulatory analysis[91] is consistent with the study's findings, which identified PgbHLHs genes possessing putative CREs in the promoter regions, thus suggesting the involvement of bHLH TFs in various physiological and developmental processes of the plants, including circadian cycle, cell cycle regulation (CCR), meristem response element (MRE), unfolded protein response elements, meristem maintenance and stem cell regulation responsive elements (Fig. 7)[92,93]. The presence of defense and stress-responsive elements crucial for abiotic responses, such as low temperature and TC-rich cold-responsive element, high light intensity responsive elements (G-box), light responsive elements (LRE), for biotic stress (fungal and bacterial responsive elements, W-box), in the promoter regions of PgbHLH genes was also observed, thereby contributing to the inherent stress tolerance of pearl millet (Pennisetum glaucum) via regulation of stress-responsive genes expression[94,95]. Moreover, the putative CREs responsible for the phyto-hormones like abscisic acid (ABA)[96], methyl jasmonate (MeJA)[97], and salicylic acid (SA)[98], indicated that PgbHLHs might be involved in regulating hormone signaling pathways related to the biotic and abiotic stresses, thus associated with stress adaptation, plant defense responses, and hormone crosstalk[98].
The dynamic appearance and disappearance of collinear relationships provide insight into the syntenic trajectory of PgbHLH genes in pearl millet. The collinearity relationships might suggest expansion and contraction of the bHLH TFs family in pearl millet across different species[99]. Among all the species, Zea mays has been reported to show the major co-relationship percentage as 33% with 150 collinear pairs, followed by Setaria italica with 137 (30%) collinear pairs, Oryza sativa with 125 (27%) collinear pairs, Solanum lycopersicum with 38 (9%) collinear pairs and least pairs with only ten (3%) were found between the A. thaliana and P. glaucum (Fig. 5). Relevant to a previous study of Feng et al.,[100] collinearity analysis revealed a robust clustering of pearl millet with monocot species (Zea mayz, Setaria italic and Oryza sativa), instead of dicotyledonous species (Solanum lycopersicum and Arabidopsis thaliana), showing that bHLH pairs might have appeared concomitantly with the pearl millet's collinearity history. Furthermore, this study's results correlate with the findings of Chanwala et al.[53] that bHLH family members might have been inherited from monocotyledonous origins and have undergone significant expansion across different plant species, remarkably showing dynamic development.
The bHLH genes have been shown to be involved in the regulation of plant growth and development[101,102]. 110 PgbHLHs were selected and assessed for their expression levels under three abiotic stresses, including heat, drought, and salt stress, at various time points (1, 3, 5, 7, 24, 48, 96, and 144 h) compared with control (CK), enabling the elucidation of their dynamic expression patterns and potential roles in stress adaptations. Compared with the findings of Hao et al. and Altschul et al. [103,104], the expression profile of the PgbHLH gene family when exposed to heat stress, drought stress, and salt stress showed a unique pattern of up-regulation and down-regulation at various time points, with comparable expression levels relative to the control. Notably, plants under heat stress exhibited significantly more pronounced up-regulation and down-regulation when compared with salt and drought stress[105]. Moreover, the expression profile of PgbHLH4.10 suggested its functional role in responding to both drought and high temperature stress (Supplementary Table S4), which is consistent with previous findings where individual transcription factors could participate in multiple signaling pathways as exemplified by one of the gene in rice OsWRKY11, has also been implicated in multiple abiotic stress responses[106], and AtWRKY39, which is responsible for multiple pathways like stress regulations responsible such as heat stress and hormone signaling cascades[107]. These observations suggested that the bHLH TFs might be involved in many plant stress activities, underscoring the complexity as well as interlinked connections of stress signaling networks.
-
This study presents the genome-wide analysis and in silico characterization of the bHLH transcription factor (TF) family in pearl millet (Pennisetum glaucum). Through phylogenetic comparison with Arabidopsis thaliana and Oryza sativa, putative orthologous and paralogous relationships that offered functional insights into PgbHLH genes were identified. Chromosomal localization, gene structure, conserved motifs, and collinearity analyses revealed both evolutionary conservation and divergence of this TF family. Promoter putative cis-regulatory element analysis highlighted an enrichment of stress-responsive elements, and in silico expression profiling under heat, drought, and salt stresses demonstrated that many PgbHLHs were transcriptionally active under abiotic stress conditions. These findings underscore the critical role of bHLH TFs in stress adaptation and provide a foundation for future functional validation studies aimed at improving abiotic stress resilience in pearl millet through molecular breeding.
-
Not applicable.
This work was supported by the National Key R&D Program of China (Grant No. 2024YFF1001300), 'Sichuan International Cooperation Project' (Grant No. 2024YFHZ0251), 'Modern Agricultural Industry System Sichuan Forage Innovation Team' (Grant No. SCCXTD-2020–16), and Sichuan Province's Science Fund for Distinguished Young Scholars with the grant (Grant No. 2021JDJQ001).
-
The authors confirm contribution to the paper as follows: study conception and design: Xie Z, Huang L; data collection: Najeeb A; analysis and interpretation of results: Hussain A, Khalid U, Hassan MJ, Tariq R; draft manuscript preparation: Najeeb A, Hassan MJ, Tariq R, Yun Z, Khalid U, Hussain A; manuscript review: Tariq R, Hassan MJ, Xie Z, Yan H, Huang L. All authors reviewed the results and approved the final version of the manuscript.
-
The data used and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Protein sequences of Arabidopsis thaliana.
- Supplementary Table S2 Protein sequences of Oryza sativa.
- Supplementary Table S3 Putative cis-elements.
- Supplementary Table S4 In-silico expression analysis.
- Supplementary Table S5 Properties of PgbHLHs.
- Supplementary Table S6 Collinearity data of Pearl millet and Arabidopsis.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Najeeb A, Tariq R, Hussain A, Hassan MJ, Khalid U, et al. 2025. Genome-wide identification, evolution, and expression profiling of bHLH transcription factor family in response to abiotic stresses in pearl millet (Pennisetum glaucum L.). Grass Research 5: e029 doi: 10.48130/grares-0025-0026 

Genome-wide identification, evolution, and expression profiling of bHLH transcription factor family in response to abiotic stresses in pearl millet (Pennisetum glaucum L.)
- Received: 04 January 2025
- Revised: 04 September 2025
- Accepted: 17 September 2025
- Published online: 13 November 2025
Abstract: Pearl millet (Pennisetum glaucum L.) is a highly valuable crop for food, fodder, and biofuel production; however, its productivity is significantly hindered by various abiotic stresses. It is one of the most functionally versatile and expansive transcription factor (TF) families in plants. The basic helix-loop-helix (bHLH) plays an imperative role in regulating responses under environmental stresses. Therefore, by using phylogenetic analysis, this study identified 110 PgbHLH TFs and broadly classified them into 12 subfamilies with 20 conserved motifs. The structural analysis illustrated that PgbHLHs proteins within each subgroup were relatively conserved. Chromosomal mapping revealed the distribution of PgbHLH genes across all seven chromosomes of pearl millet. Evolutionary analysis of pearl millet with Arabidopsis thaliana (145 AtbHLH genes) and Oryza sativa (139 OsbHLH genes) revealed distinct orthologous and paralogous genes, facilitating gene functions prediction in reference species. Collinearity analysis showed both the conservation of PgbHLH genes across multiple plant species and their collinear divergence based on gene order and similarity metrics, providing valuable insights into the functional evolution of this gene family. Additionally, putative cis-regulatory element (CRE) analysis of the PgbHLH promoters revealed a diverse range of regulatory elements, with stress-responsive CREs accounting for 34% of all identified elements. Moreover, in-silico expression profiling of PgbHLH genes in leaf and root tissues under heat, drought, and salt stress conditions demonstrated their diverse regulatory roles in abiotic stress responses. This study provides a valuable resource for future functional validation studies aimed at improving abiotic stress resilience in pearl millet through molecular breeding.
-
Key words:
- Genome wide /
- Pearl millet /
- bHLH /
- Phylogenetic tree /
- Collinearity /
- Evolution /
- Abiotic stress.