









-
Crataegi Fructus (Hawthorn, CF) is derived from the dried mature fruits of Crataegus pinnatifida Bge. var. major N.E. and Crataegus pinnatifida Bge, as recommended by the Chinese Pharmacopoeia[1]. Pharmacological studies have demonstrated its protective effect on myocardial ischaemia[2]. Wang et al. studied the mechanism of the decoction with the odor of charred CF products on rat models with high-calorie-diet-induced dyspepsia (HC-DID) and observed that it effectively alleviated HC-DID in the studied animals[3]. Dahmer & Scott reported the application of CF in the treatment of dyslipidemia, hypotension, angina, arrhythmias, and indigestion[4]. CF is not only used in clinics, but is also made into jam, wine, and canned due to its taste. Therefore, the National Health Commission of the People's Republic of China has recommended CF as a medicinal and edible herb. In addition to China, CF has a long medical and edible history in Europe and North America. Given its widespread use, the safe utilization of CF is a global concern. In recent years, numerous cases of fungal contamination in herbs have been reported globally. In India, Singh et al. isolated 858 strains from six medicinal plant samples and observed that Aspergillus was the dominant genus[5]. Furthermore, 366 isolates were identified as Aspergillus flavus, Aspergillus niger, and Aspergillus terreus, which were toxigenic[6,7]. de Sousa Lima et al. reported that fungal contamination was observed in 35.6% of 132 herbal medicine samples, and the contamination level in 31.0% of samples exceeded the legal limits set by the World Health Organization[8]. In China, a report by Yu et al. studied the fungal contamination in Platycladi Semen, and the results showed that fungal contamination was detected in all samples. Four potential mycotoxin-producing fungi, including A. flavus, A. fumigatus, Fusarium poae, and Penicillium steckii, were detected in all 11 samples[9]. Storage is an essential procedure for herbs in practical production chains. However, herbs are susceptible to fungal contamination during storage under proper conditions. Wang et al. assessed the effect of storage condition on the fungal community in Ophiopogonis Radix. The results showed that A. flavus, A. tubingensis, and Gibberella avenacea were the main contaminating fungi. Furthermore, the fungal biomass increased as the storage temperature and humidity increased[10]. Another study performed by Kulshrestha et al. also showed that A. flavus contamination was observed in ten medicinal herbs under storage temperatures at 25, 30, and 40 °C[11]. Therefore, it is essential to evaluate the effect of storage conditions on the changes of fungal communities in CF to provide valuable information for preventing and controlling fungal and mycotoxin contamination.
With the development of next-generation platforms, Taberlet et al. proposed the concept of DNA metabarcoding[12]. Compared to the traditional Sanger sequencing method, DNA metabarcoding can identify multiple species from a single environmental sample. Moreover, it enables the comparison across studies and within time-series simultaneously[13]. This method has been applied in various fields, including food, environment, and clinics[14−16]. In recent years, DNA metabarcoding has been used to study fungal communities in some herbal materials[17−20]. The utilization of DNA metabarcoding provides an efficient method for monitoring the changes of fungal communities in CF during storage.
In this study, we analyzed the changes of fungal communities in CF at different storage time points, and studied the differences of fungal communities in CF stored at room temperature (RT), and 4 °C. To our knowledge, this research is the first to assess the effect of storage on fungal contamination in CF using DNA metabarcoding. Thus, this research provides a basis for the safe utilization of CF in clinical settings.
-
Approximately 500 g of CF materials were collected from Hebei province in China, which is a main producing area for CF. The materials were placed into sterile bags, and stored at the Institute of Medicinal Plant Development, Chinese Academy of Medical Sciences (Beijing, China). The CF materials were divided into two parts, and stored at room temperature (RT, ranging from 18.0−27.7 °C with an average temperature at 20.9 °C, Supplementary Table S1), and 4 °C. The storage temperature was recorded three times daily (8:00, 14:00, and 20:00). CF samples stored under two temperature conditions were collected at four time points during the storage period (at 0, 8, 16, and 24 weeks). The initial sample stored for 0 week was designated as CF0W. The samples stored at RT for 8, 16, and 24 weeks were designated as CFR8W, CFR16W, and CFR24W respectively. The samples stored at 4 °C for 8, 16, and 24 weeks were designated as CFF8W, CFF16W, and CFF24W. Detailed information is listed in Table 1.
Table 1. Sample information for the CF investigated in this study.
Storage time Storage temperature Room temperature 4 °C 0-Week CF0W CF0W 8-Week CFR8W CFF8W 16-Week CFR16W CFF16W 24-Week CFR24W CFF24W DNA extraction and polymerase chain reaction (PCR) amplification
-
Approximately 3.30 g of CF sample was transferred into a 50 mL sterilized centrifuge tube. The elution procedure was performed as described by Yu et al.[21]. DNA extraction was performed according to the instructions of the EZNA® soil DNA kit (Omega Bio-tek., Inc., Norcross, GA, USA). The DNA products were frozen immediately at −20 °C until PCR amplification. The primers of ITS3 (5'-GCATCGATGAAGAACGCAGC-3'), and ITS4 (5'-TCCTCCGCTTATTGATATGC-3') were designed to amplify the internal transcribed spacer 2 (ITS2) region as described by White et al.[22]. The PCR conditions were as follows: initial denaturation at 95 °C for 3 min; 35 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, elongation at 72 °C for 45 s, and a final extension at 72 °C for 10 min. The quality of PCR products was analyzed on 1% agarose gel, and the desired fragment was purified. The purified ITS2 amplicons were sequenced through the Illumina Miseq PE300 platform (Illumina, San Diego, CA, USA).
Bioinformatic analysis
-
The raw sequences were uploaded to the National Center for Biotechnology Information Sequence Read Archive database with the accession numbers (SAMN18864584, SAMN20297324−SAMN20297329). The raw FASTAQ files were demultiplexed and quality filtered by Fastp. Reads were truncated at any site when received an average quality score of at least 20 over a 50 bp sliding window. The valid sequences were then clustered into OTUs (97% similarity) through UPARSE software in the UNITE database in Quantitative Insights into Microbial Ecology (QIIME, V. 1.9.1,
http://qiime.org/install/index.html )[23,24]. Chimeric sequences were removed using USEARCH. Each OTU was annotated against the UNITE (version 8.0) database at the phylum, class, order, family, genus, and species level. To increase the accuracy of the OTU annotation, we verified the taxonomical classification of each OTU through manual BLAST search in the International Nucleotide Sequence Database Collaboration. Venn analysis and rarefaction curves were constructed using R software (version 3.3.1). Four alpha indices (Shannon, Simpson, Chao, and Coverage) were calculated using Mothur to evaluate the fungal diversity in the CF samples. Fungal community analysis was performed using Bar plot and heatmap via Vegan packages in R software (version 3.3.1). Statistical differences in the fungal community between two samples were analyzed using Fisher's exact test by Stats packages in R software (version 3.3.1) and SciPy packages in Python. -
A total of 415,534 valid sequences (280−400 bp) were obtained from all seven CF samples. The raw sequences were uploaded to the National Center for Biotechnology Information Sequence Read Archive database and were assigned accession numbers. All sequences were clustered into 329 operational taxonomic units (OTUs) with 97% similarity using the UNITE database. The rarefaction curves showed that all seven curves were homogeneously parallel to the x-axis, indicating that the normalization of data to the sequencing depth of each sample was sufficient to assess fungal diversity (Supplementary Fig. S1). The distribution of OTUs in seven CF samples is listed in Supplementary Table S2. The Venn analysis result revealed the numbers of shared and unique OTUs between the two groups (Fig. 1). A total of 108 shared OTUs were obtained between the two groups. The CFR group had 54 unique OTUs, while the CFF group had 151 unique OTUs. CFR8W (126 OTUs), and CFF16W (196 OTUs) had the highest number of OTUs in the CFR and CFF groups, and the CFR16W (51 OTUs), and CFF8W (128 OTUs) had the lowest number of OTUs. Fungal diversity was estimated using four alpha indices (Table 2). The coverage results showed that all seven CF samples yielded over 99.9%. The Chao1 index was used to evaluate the fungal community richness at different storage stages. It was the highest in the CFR8W, indicating the highest community richness in these samples during storage. The Shannon and Simpson indices were used to assess the fungal community diversity and observed that CFR24W and CFF24W exhibited the highest community diversity. The results showed that the community diversity of samples during storage at 4 °C increased with storage time, whereas the community diversity of samples during storage at RT showed an increase-decrease-increase trend and reached the peak at storage at 24-Week. Similarly, the community richness of samples stored under RT and 4 °C showed an increase-decrease trend, with peaks observed at 8-Week and 16-Week for samples stored at RT and 4 °C, respectively.
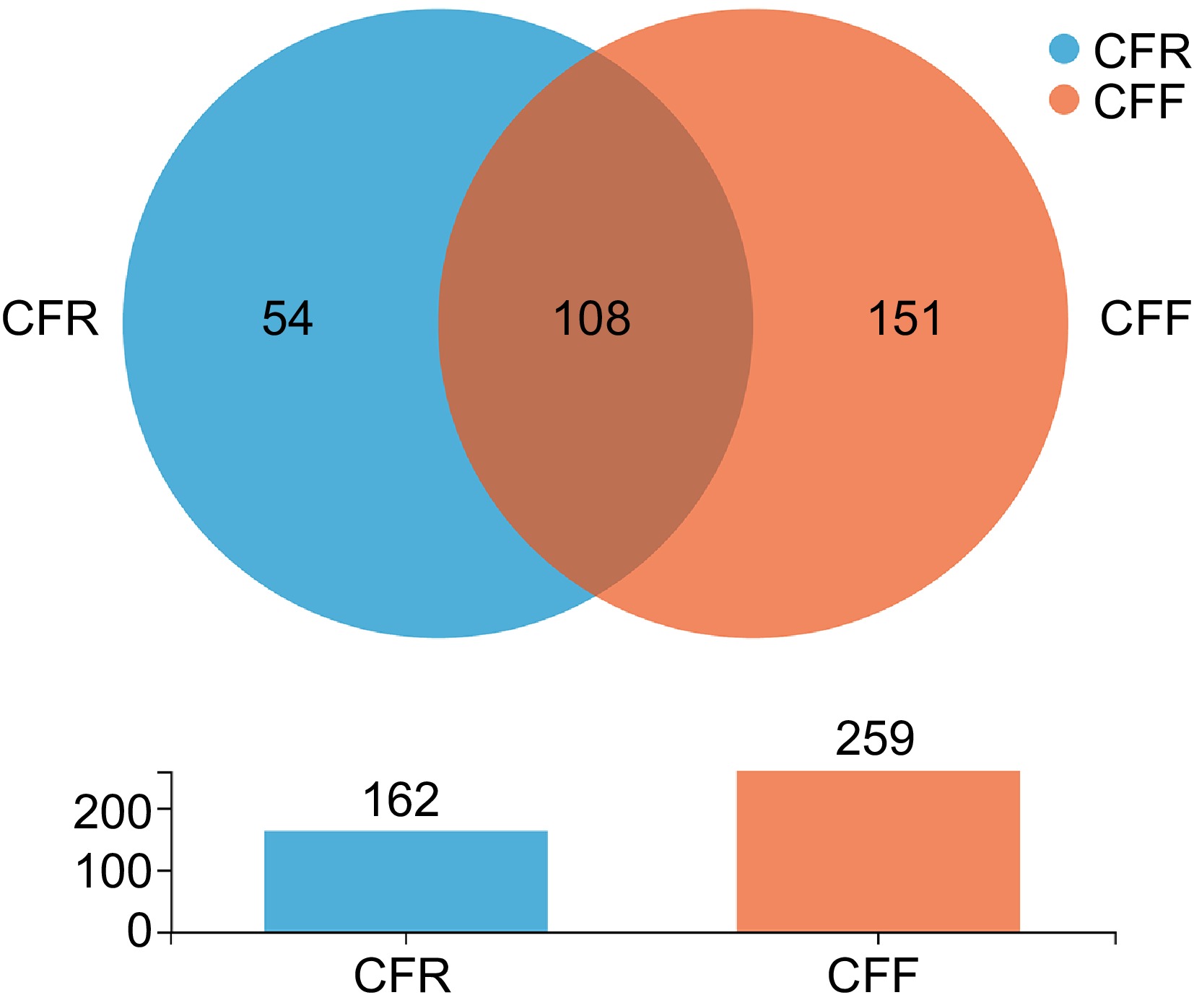
Figure 1.
Venn diagram of OTUs in CFF and CFR groups.
Table 2. Observed OTU numbers and alpha diversity indices of CF samples.
Sample Observed
OTU numbersShannon Simpson Chao Coverage Genbank
accession No.CF0W 81 2.45 0.18 81.00 1.000000 SAMN18864584 CFR8W 126 2.63 0.12 129.33 0.999928 SAMN20297327 CFR16W 51 1.01 0.67 51.00 0.999980 SAMN20297328 CFR24W 38 2.95 0.08 38.00 1.000000 SAMN20297329 CFF8W 128 1.02 0.62 129.62 0.999902 SAMN20297324 CFF16W 196 2.00 0.25 208.00 0.999627 SAMN20297325 CFF24W 131 3.29 0.08 136.00 0.999859 SAMN20297326 Fungal community composition in CF samples
-
All 329 OTUs were annotated against the UNITE database at various taxonomic levels. At the phylum level, Ascomycota was dominant with a relative abundance of 82.88%−99.39%, followed by Basidiomycota (0−16.68%) and others (Fig. 2a). Among these samples, the relative abundance of Ascomycota was the highest in CFF8W and lowest in CFF24W, respectively. At the class level, Dothideomycetes (40.43%−94.18%) and Eurotiomycetes (0.82%−27.85%) were the most abundant classes in all seven CF samples (Fig. 2b). Further taxonomic analysis at the order level showed that Pleosporales and Capnodiales predominated among all orders (Fig. 2c). At the family level, Didmosphaeriaceae and Pleosporaceae had the highest relative abundance in the CF samples (Fig. 2d). In addition, the fungal community composition was analyzed at the genus level, with Paraconiothyrium, Alternaria, and Nothophoma being the three most abundant genera among all genera (Fig. 2e). To ensure the accuracy of identification, the taxonomical classification of all OTUs was verified at the species level through manual BLAST search in the International Nucleotide Sequence Database Collaboration. A total of 67 species were identified, including seven potentially toxigenic species: Rhizopus microsporus, Penicillium oxalicum, Trichothecium roseum, Candida tropicalis, Penicillium brevicompactum, Beauveria bassiana, and Talaromyces rugulosus.
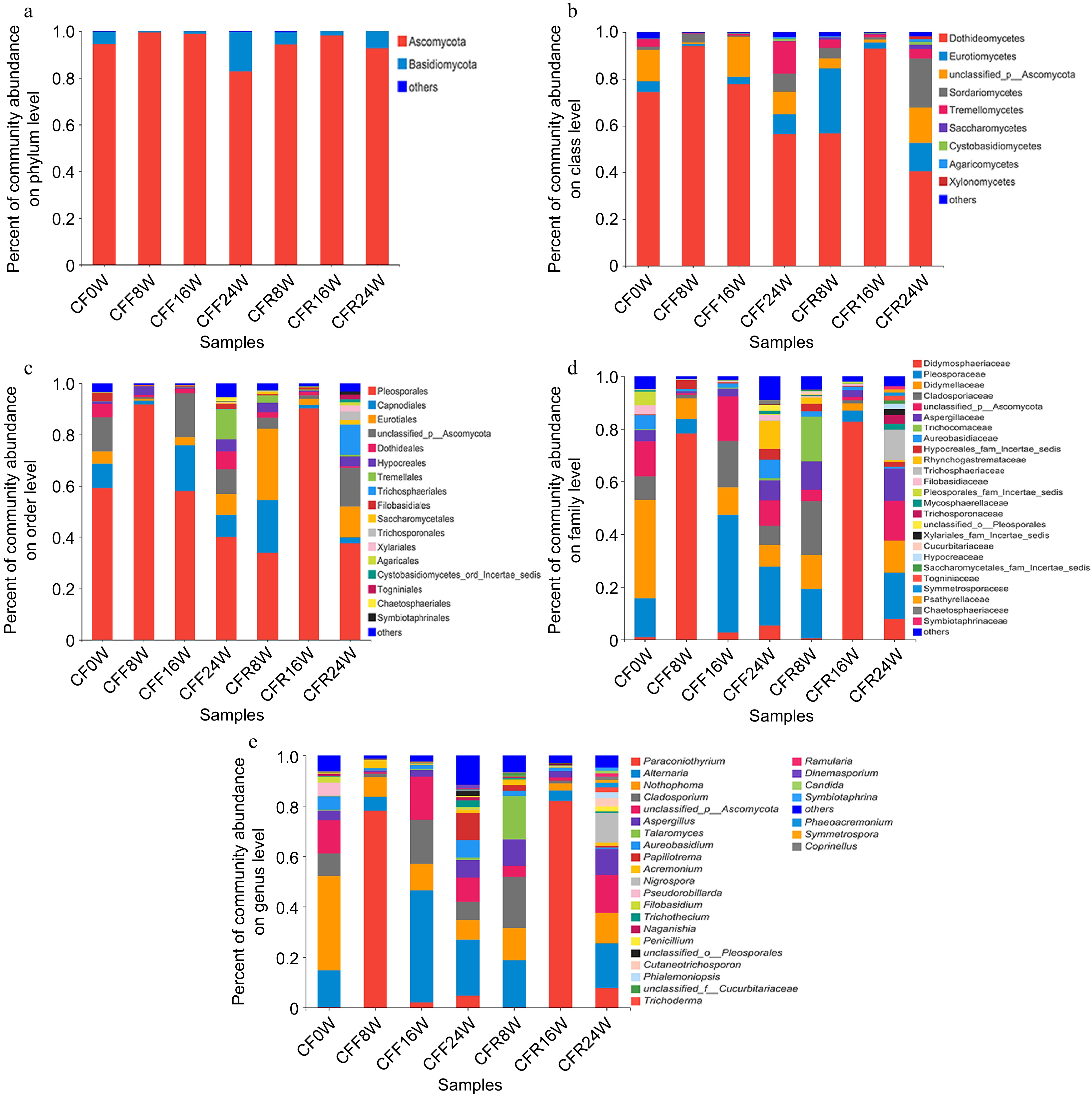
Figure 2.
Analysis of fungal community composition of seven CF samples. (a) Phylum level; (b) class level; (c) order level; (d) family level; (e) genus level.
Changes of fungal community in CF samples of different storage times stored at RT
-
We used DNA metabarcoding to monitor the changes in the fungal community in samples stored at RT. The relative abundance of Ascomycota was the highest in CFR16W (98.16%), followed by CF0W (94.46%), CFF8W (94.29%), and CFF24W (92.69%). Meanwhile, Basidiomycota exhibited the highest abundance in CFR24W (7.31%), followed by CF0W (5.28%), CFR8W (5.16%), and CFR16W (1.84%, Fig. 3a). At the class level (Fig. 3b), compared to other samples (40.43%−74.29%), the relative abundance of Dothideomycetes was higher in CFR16W (93.05%). Pleosporales was the most abundant order in the CFR samples, with the highest level of Pleosporales was detected in CFR16W (90.28%), followed by CF0W (59.27%), CFR24W (37.58%), and CFR8W (33.85%, Fig. 3c). At the family level, the relative abundance of Didmosphaeriaceae was detected highest in CFR16W (82.74%), followed by CFR24W (7.75%), CF0W (0.87%), and CFR8W (0.52%, Fig. 3d). We analyzed the changes in the fungal community composition at the genus level (Fig. 4a). Nothophoma predominated at the initial stage. The relative abundances of Talaromyces, Alternaria, Cladosporium, and Aspergillus were the highest at the 8-Week, whereas Paraconiothyrium predominated at the 16-Week. Furthermore, Nigrospora and Cutaneotrichosporon were dominant at the 24-Week. At the species level, we observed that the relative abundances of potential mycotoxin fungi increased with storage time, except 16-Week (Table 3).
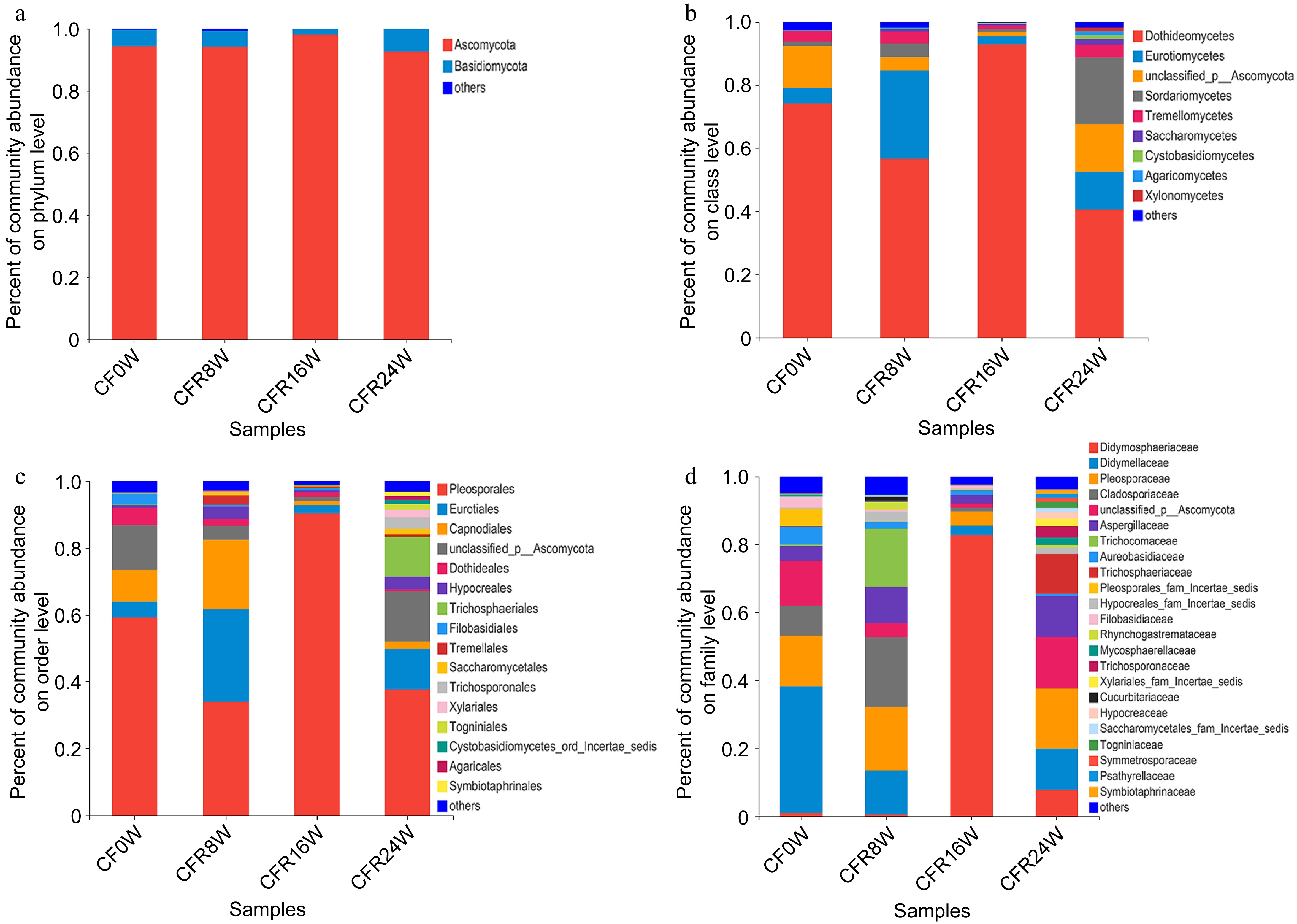
Figure 3.
Analysis of fungal community composition of the CFR samples. (a) Phylum level; (b) class level; (c) order level; (d) family level.
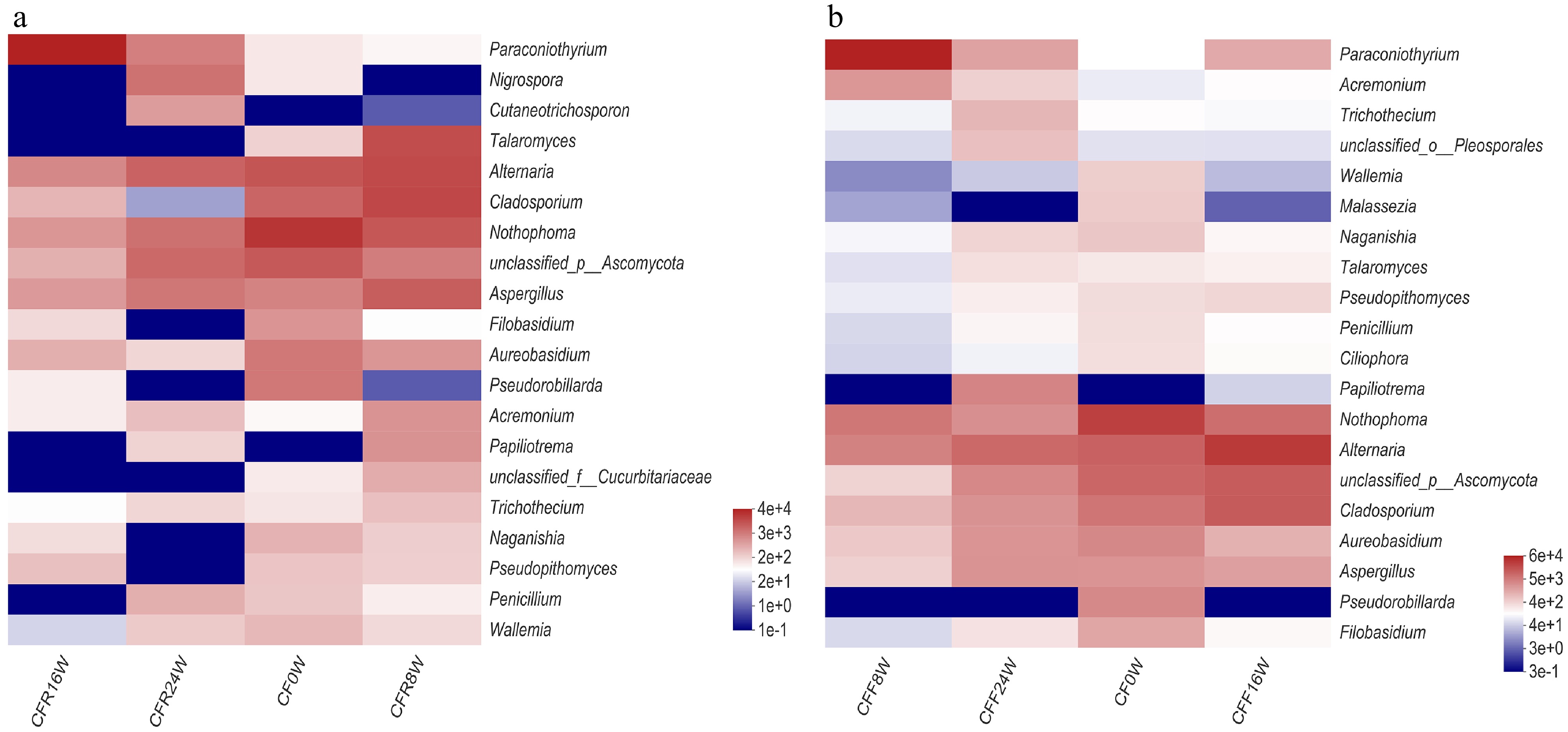
Figure 4.
Differences of fungal community at the genus level in (a) the CFR samples and (b) the CFF samples.
Table 3. Proportion of potentially toxigenic fungi in seven CF samples.
Sample No. The relative abundances of potentially toxigenic fungi in CF samples Rhizopus microsporus Penicillium oxalicum Trichothecium roseum Candida tropicalis Penicillium brevicompactum Beauveria bassiana Talaromyces rugulosus CF0W — — 47 18.29% 139 6.86% — — — — — — — — CFR8W — — 59 22.96% 422 20.82% — — 29 46.03% 16 66.67% 164 99.39% CFR16W — — — — 68 3.35% — — — — — — — — CFR24W — — — — 218 10.75% 448 100.00% — — — — — — CFF8W — — 12 4.67% 96 4.74% — — 26 41.27% — — — — CFF16W 3 100.00% 89 34.63% 112 5.53% — — 8 12.70% 8 33.33% 1 0.61% CFF24W — — 50 19.46% 972 47.95% — — — — — — — — Total 3 100.00% 257 100.00% 2027 100.00% 448 100.00% 63 100.00% 24 100.00% 165 100.00% Changes of fungal composition in CF samples of different storage times stored at 4 °C
-
The fungal community in CF samples stored at different storage times at 4 °C was analyzed over different storage times. As the storage time increased, the fungal richness in the CF samples increased except for CFF8W. The Shannon index in the CFF8W sample was the lowest, representing the lowest fungal richness. We also monitored the changes in the fungal community during storage. The relative abundance of Ascomycota remained at a high level at 8-Week stage (99.39%) and 16-Week stage (98.85%, Fig. 5a). At the class level (Fig. 5b), the relative abundance of Dothideomycetes was the highest and lowest in CFF8W (94.18%) and CFF24W (56.42%), respectively. Moreover, Pleosporales feathered the highest level in CFF8W (91.76%), followed by CF0W (59.27%), CFF16W (58.13%), and CFF24W (39.91%, Fig. 5c). At the family level, the relative abundance of Didmosphaeriaceae was significantly higher in CFF8W than that in the other three samples (Fig. 5d). Paraconiothyrium was the dominant genus among samples, particularly in CFF8W, followed by Alternaria and Nothophoma. At the initial stage (0-Week), Nothophoma was the most abundant genus. At the 8-Week stage, the dominant genus changed from Nothophoma to Paraconiothyrium. At the later period (16-Week and 24-Week), the relative abundance of Alternaria increased (Fig. 4b). At the species level, the relative abundance of potentially toxigenic fungi was highest in CFF24W, followed by CFF0W, CFF8W, and CFF16W (Table 3).
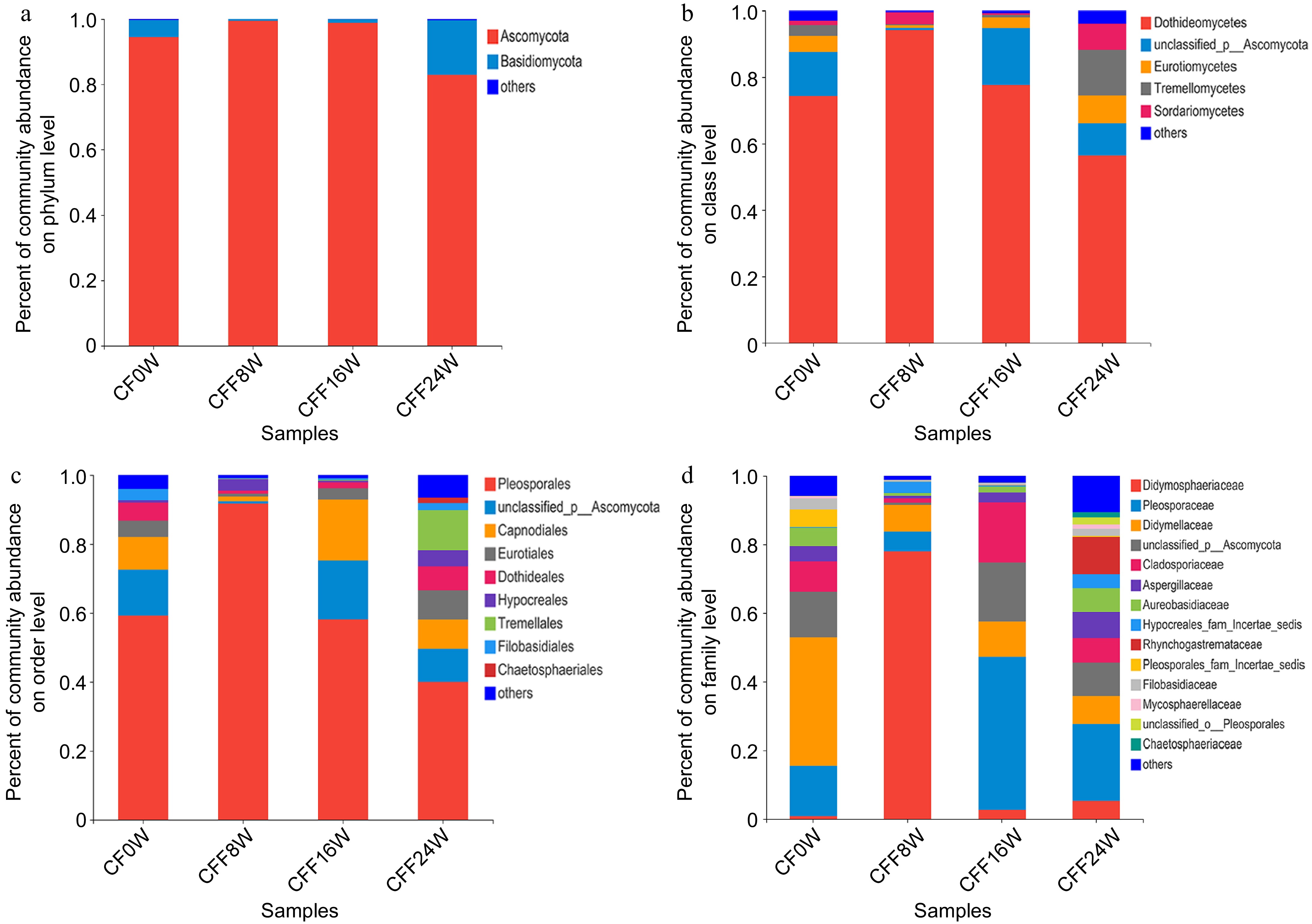
Figure 5.
Analysis of fungal community composition in the CFF samples. (a) Phylum level; (b) class level; (c) order level; (d) family level.
Comparison of the differences of fungal community between samples stored at RT and 4 °C
-
To study the effect of storage temperature on the fungal community in CF, we compared the differences in the fungal community between the samples stored at RT and 4 °C at each time point. At the 8-Week stage (Fig. 6a), the relative abundances of Paraconiothyrium and Acremonium in CFR8W were significantly higher than those in CFF8W, whereas Alternaria, Cladosporium, Nothophoma, Talaromyces, and Aspergillus had higher numbers in CFF8W than in CFR8W. At the 16-Week stage (Fig. 6b), the differences of Paraconiothyrium, Alternaria, Cladosporium, Nothophoma, Aspergillus, Fusarium, and Trichothecium were remarkably distinguishable. At the 24-Week stage (Fig. 6c), compared with those in CFF24W, the relative abundances of Nothophoma, Aspergillus, Paraconiothyrium, Nigrospora, and Penicillium were significantly higher in CFR24W, while Alternaria, Papiliotrema, Aureobasidium, Cladosporium, and Trichothecium had lower relative abundance. At the species level, the relative abundances of seven potentially toxigenic fungi in the CFF samples were higher than those in the CFR samples (Table 3).
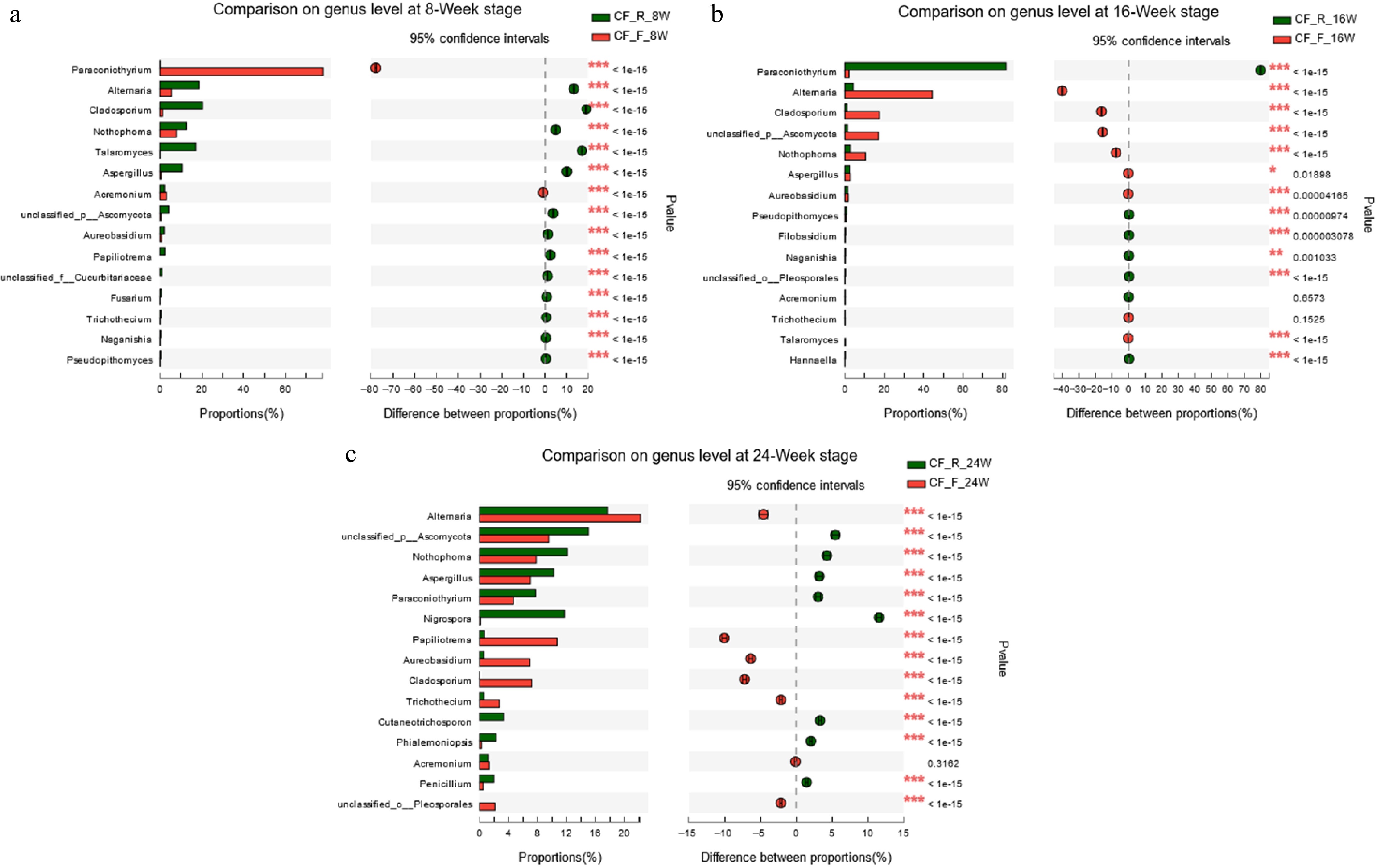
Figure 6.
Comparison of the differences of fungal community at the genus level between samples stored at RT and 4 °C for (a) 8-Week; (b) 16-Week; (c) 24-Week.
-
Fungal contamination, which may originate from various stages (harvest, processing, storage, and transport), affects the quality and safety of food and herbs. Numerous studies have reported the occurrence of fungal and mycotoxin contamination in food and herbs during storage. Storage time is a crucial factor that influences the changes in fungal community. Yang et al. analyzed the dynamics of fungal community in Citri Reticulatae Pericarpium stored for three years, indicating that Penicillium predominated at the initial stage, and the relative abundance of Aspergillus increased at the late stage[25]. Based on the report by Krijgsheld et al., Aspergillus spp. were common fungi that occurred prevalently at the late stage during storage[26]. In Uganda, Kaaya et al. studied the effect of storage time on fungal and aflatoxin contamination in maize, observing that the fungal counts and aflatoxin levels increased with storage time, especially the percentage of Aspergillus and Penicillium species in Mid-Altitude maize samples[27]. Manthou et al. assessed the changes in the fungal community in pineapple and indicated that the dominant genus was Candida during storage, with Fusarium predominating at the late stage[28]. Therefore, the fungal community may change at different storage times. In the present study, the changes of fungal community in CF during storage were monitored through DNA metabarcoding. At the genus level, the changes in the fungal community in CFR samples at various stages were analyzed. We observed that the dominant genera changed from Nothophoma and Pseudorobillarda to Alternaria, Trichothecium, and Papiliotrema during storage. Significant differences in the fungal community in CFF samples at different times were also observed, especially the relative abundances of Paraconiothyrium, Alternaria, and Nothophoma. Furthermore, Pseudorobillarda and Filobasidium exhibited high numbers in the initial period, while Trichothecium and Papiliotrema showed high numbers in the later period. Aspergillus remained at relatively stable low levels during storage. In conclusion, it is significant to study the effect of the storage time on the fungal community to provide references for the quality improvement of CF. At the same time, DNA metabarcoding also displayed its advantages in dynamically monitoring the fungal contamination in CF during storage.
Relationship between storage temperature and fungal community in food and herbs
-
Fungal and mycotoxin contamination occurs under proper storage conditions, and significantly affects the quality and safety of food and herbs. In addition to storage time, storage temperature is another crucial factor influencing the fungal community during storage. In recent years, numerous studies have reported the effect of storage temperature on the fungal community in herbs and food. For instance, Lu et al. evaluated the effect of storage temperature on the growth and mycotoxin production of Aspergillus versicolor in Ilex asprella, observing that A. versicolor grew rapidly, and produced large quantities of sterigmatocystin at temperatures above 26 °C[29]. Similarly, Abdelfattah et al. also demonstrated that low-temperature storage could inhibit fungal contamination in apples[30]. Zhang et al. compared the differences in the fungal community in harvested wheat stored at four temperature conditions (15, 20, 25, and 30 °C). The result showed that the relative abundances of Aspergillus and Penicillium increased at 25 and 30 °C, respectively, while the relative abundance of Papiliotrema decreased at high temperatures[31]. In the present study, we compared the differences in the fungal community of CF stored at two temperature conditions (RT and 4 °C). In the CFF samples stored at 4 °C, the three most abundant genera were Paraconiothyrium, Alternaria, and Nothophoma. Meanwhile, in CFR samples stored at RT, Paraconiothyrium, Alternaria, and Cladosporium predominated. Although the storage temperature did not affect the types of dominant fungal genera in CF, with the exception of Nothophoma and Cladosporium, it affected the relative abundances of dominant fungal genera. The average relative abundance of Paraconiothyrium was higher in CFR samples than that in CFF samples, and the average relative abundance of Alternaria samples was lower in the CFR than that in CFF samples. Additionally, several interesting results were observed. Paraconiothyrium was the most abundant genus in all CF samples. Its highest abundance of Paraconiothyrium in the CFF samples was observed at the 8-Week stage, while its highest level in the CFR samples was observed at the 16-Week stage. This indicates that the lower storage temperature slowed the growth of Paraconiothyrium. It has been reported that this genus is usually distributed in multiple hosts including plant, soil, and marine environments[32]. Therefore, the presence of this genus in CF might be attributed to the surrounding environment during storage.
This work was supported by the CAMS Innovation Fund for Medical Sciences (CIFMS) (No. 2021-I2M-1-071).
-
The authors confirm contribution to the paper as follows: data curation: Yu J, Dao y, Han J; methodology: Yu J, Yang M, Pang X; formal analysis, writing—original draft, review & editing: Yu J, Pang X; conceptualization, funding acquisition, resources, validation: Pang X. All authors reviewed the results and approved the final version of the manuscript.
-
The data underlying this article are available in the GenBank Nucleotide Database, and can be accessed with SAMN18864584, SAMN20297324−SAMN20297329.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Temperature recording for CF samples that stored at RT for 24 weeks.
- Supplementary Table S2 Distribution of OTU for seven CF samples.
- Supplementary Fig. S1 Rarefaction curves for OTU in seven CF samples.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yu J, Dao Y, Yang M, Han J, Pang X. 2025. Effect of storage conditions on fungal community in Crataegi Fructus through DNA metabarcoding. Medicinal Plant Biology 4: e013 doi: 10.48130/mpb-0025-0008 

Effect of storage conditions on fungal community in Crataegi Fructus through DNA metabarcoding
- Received: 07 January 2025
- Revised: 28 February 2025
- Accepted: 10 March 2025
- Published online: 18 April 2025
Abstract: Herbs are susceptible to fungal contamination during storage under proper conditions, which affects their quality and safety. Storage time and temperature are two crucial factors that influence the fungal community. In this study, we investigated the impact of both factors on the dynamics of the fungal community in Crataegi Fructus (CF), a medicinal and edible herb in China. The CF materials were stored under two different temperatures (room temperature and 4 °C) and collected at four time points (0, 8, 16, and 24 weeks). DNA metabarcoding was employed to monitor the changes of fungal communities in CF. Results showed that the fungal community in CF samples remarkably changed over the storage period. Significant differences in the relative abundances of dominant genera were observed in CF samples stored at different temperatures. A total of 67 species were identified, including seven potentially toxigenic fungal species, namely Rhizopus microsporus, Penicillium oxalicum, Trichothecium roseum, Candida tropicalis, Penicillium brevicompactum, Beauveria bassiana, and Talaromyces rugulosus. Both storage time and temperature significantly influenced the relative abundances of certain fungi in CF samples, particularly the dominant fungi. This study demonstrates that DNA metabarcoding is an effective method for assessing fungal contamination in CF. It also discusses the effects of storage time and temperature on the fungal community in CF, providing references for the proposal of prevention strategies in practical CF production chains.
-
Key words:
- Hawthorn /
- Storage time /
- Storage temperature /
- Fungal contamination /
- DNA metabarcoding