








-
Flower coloration is a key ornamental trait, with blue hues uniquely prized for their rarity and aesthetic appeal. However, true blue flowers are rare in nature, representing less than 10% of angiosperms[1]. This scarcity contrasts with their ecological importance; short-wavelength hues, particularly blue and violet, strongly attract specific pollinators, such as bees, which possess high sensitivity to the blue spectral region[2,3].
Blue pigments are derived from delphinidin-based anthocyanins. However, delphinidin is unstable and appears purple under weakly acidic conditions (pH 4.5−5.5) of plant vacuoles[4]. Delphinidin derivatives, characterized by a tri-hydroxylated B-ring, contribute to the blue hue in species such as Delphinium and Gentiana. In addition, blue coloration can be achieved via cyanidin-based pigments such as protocyanin in the cornflower (Centaurea cyanus). Cyanidin derivatives (e.g., succinylcyanin) with flavone glycosides and metal ions form a supramolecular metal complex[5]. Trivalent metal ions (e.g., Fe3+ or Al3+) typically induce the blue hue via ligand-to-metal charge transfer (LMCT), whereas divalent metal ions (Mg2+ or Ca2+) are essential for stabilizing the rigid supramolecular assembly.
Plants induce a bathochromic shift and stabilize the chromophore to achieve a stable "true blue" color. In this review, "true blue" refers to flower colors that are perceived as predominantly blue rather than violet-blue or purple-leaning, and can be described using the blue region of the Royal Horticultural Society (RHS) color chart as a practical reference framework. By contrast, phenotypes described as violet, violet-blue, or blue-purple in the literature are not treated here as equivalent to "true blue". This transformation is facilitated via four primary mechanisms. (1) Polyacylation: In this mechanism, aromatic acyl groups (e.g., p-coumaroyl and caffeoyl) covalently attach to anthocyanins. This modification protects the chromophore via intramolecular stacking[6,7]. (2) Intermolecular copigmentation: In this mechanism, anthocyanins form supramolecular complexes with colorless copigments (typically flavones or flavonols) through noncovalent interactions[6,8]. (3) Metalloanthocyanin formation: In this mechanism, highly ordered supramolecular complexes are formed by anthocyanins, copigments, and metal ions (e.g., Mg2+, Fe3+)[9]. (4) Vacuolar pH regulation: In this mechanism, vacuolar pH is increased from acidic to neutral to promote the formation of the blue quinoidal anion form of anthocyanins[10]. These sophisticated mechanisms occur naturally in a few vivid blue flower-producing species such as Delphinium, Gentiana, and Salvia[11−13]. However, many economically important ornamental crops—including Rosa hybrida (rose), Chrysanthemum morifolium (chrysanthemum), Dianthus caryophyllus (carnation), and Phalaenopsis aphrodite (orchid)—lack the functional flavonoid 3',5'-hydroxylase (F3'5'H) gene required for delphinidin biosynthesis and the specific chromophore stabilization mechanisms described above[14,15]. The dual deficiency of F3'5'H and chromophore stabilization mechanisms makes developing blue cultivars in these species a significant challenge for traditional breeding. As a practical alternative in ornamental horticulture, some commercially available "blue" flowers are produced by postharvest artificial dyeing, reflecting the difficulty of achieving stable blue coloration through breeding or genetic engineering.
This review summarizes the biochemical strategies used for the production of blue flowers in model plants and their subsequent application in genetic engineering. The unique stabilization mechanisms in Gentiana, Lamiaceae, and Delphinium are described and assessed to understand how these natural strategies have been translated into genetic engineering breakthroughs. These advancements range from first-generation substrate engineering to third-generation synthetic biology strategies, which reconstruct autonomous stabilization mechanisms for the production of true blue color.
-
Gentians (Gentiana spp.), flowering from midsummer to early autumn, are among the most important blue-flowered ornamental crops in Japan. These flowers hold strong cultural significance and serve as offerings at Buddhist altars. Commercial cultivars are primarily derived from Gentiana triflora and G. scabra or their interspecific hybrids. The vivid blue color of gentian flowers is produced by a polyacylated anthocyanin known as gentiodelphin [delphinidin 3-O-glucosyl-5-O-(6-caffeoylglucoside)-3'-O-(6-caffeoylglucoside)] (Fig. 1a)[16]. Structurally, gentiodelphin contains delphinidin as its aglycone and is substituted with three glucose moieties at the 3-, 5-, and 3' positions. In addition, two caffeic acid residues are esterified to the glucosyl groups attached at the 5- and 3' positions.
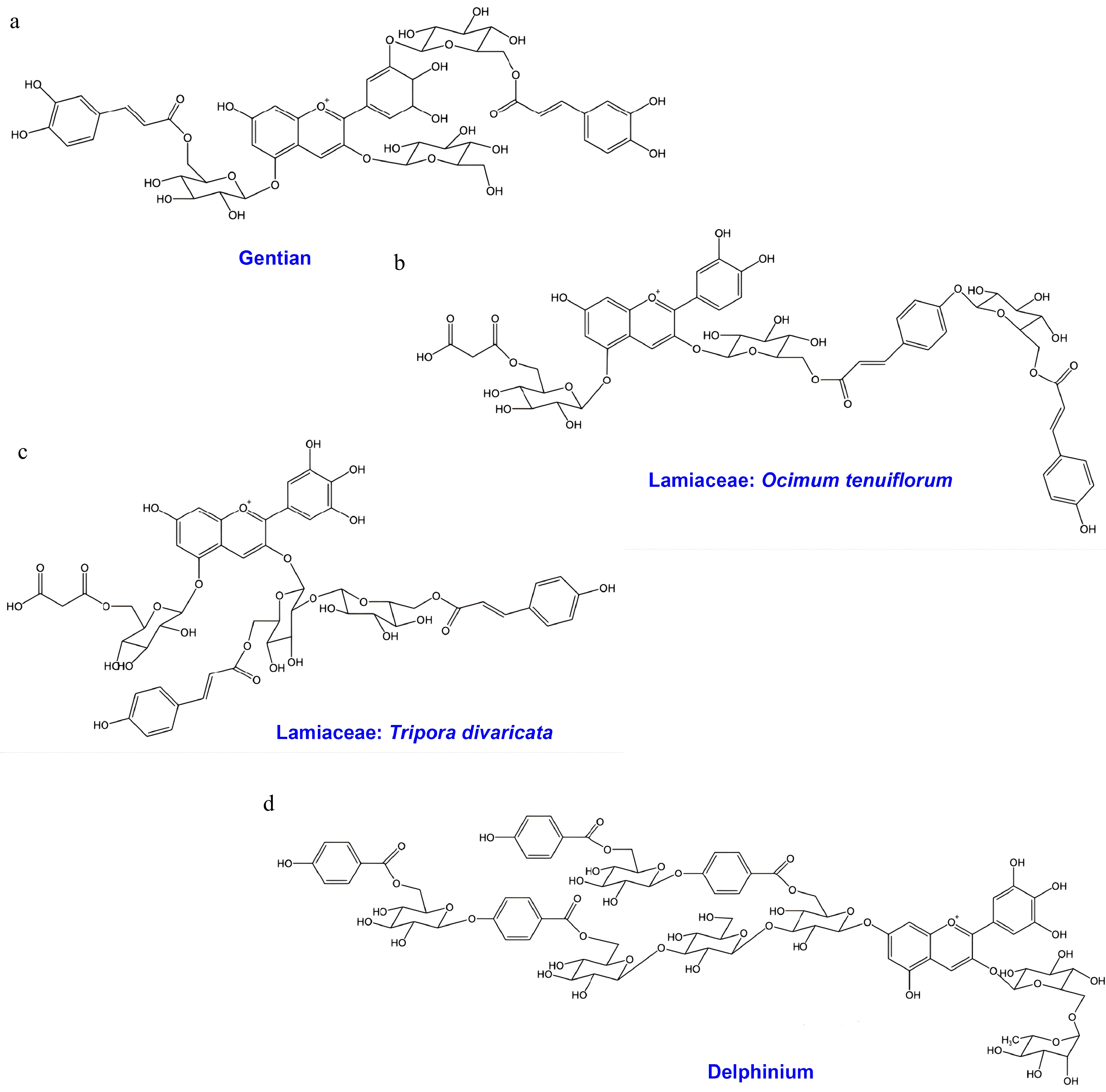
Figure 1.
Chemical structure of polyacylated anthocyanins from Gentian, Lamiaceae, and Delphinium species. (a) Gentiodelphin from Gentiana sp.; (b) polyacylated anthocyanin from Ocimum tenuiflorum; (c) polyacylated anthocyanin from Tripora divaricata; (d) cyanodelphin from Delphinium sp.
The delphinidin aglycone is essential for the blue coloration. Disruption of F3'5'H has been reported to shift the metabolic flux toward cyanidin derivatives, resulting in the production of pink flowers[17]. Beyond the aglycone itself, the stable blue hue of gentiodelphin is attributed to intramolecular copigmentation (also referred to as intramolecular stacking) between the caffeoyl residues and the anthocyanidin chromophore. Among the two acyl groups, the caffeic acid esterified to glucose at the 3′ position is particularly critical for the development of blue color[16]. This finding is supported by genome editing studies, which show that the deletion of anthocyanin 3′-glucosyltransferase leads to a loss of blue coloration[18].
The gentiodelphin biosynthetic pathway has been fully elucidated, and all the enzymes involved have been identified. The modification steps exhibit a specific order and some flexibility. Since the anthocyanin 3-glucosyltransferase (3GT) cannot utilize delphinidin 3-O-glucoside (Dp3G) as an acceptor, whereas anthocyanin 5-glucosyltransferase (5GT) accepts Dp3G efficiently[19], glucosylation at the 5 position likely follows the initial 3-glucosylation. The subsequent steps are more complex. The 3'GT enzyme can utilize Dp3,5-di-O-glucoside (Dp3,5dG) and Dp3G-5-O-caffeoylglucoside as substrates. Notably, the gentian anthocyanin acyltransferase (AT) is a unique acyl-Coenzyme A (CoA)-dependent enzyme capable of transferring caffeic acid to the 5- and 3'-glucosyl positions[16]. This dual-function 5/3'AT accepts Dp3,5dG and Dp3,5,3'G as substrates. Collectively, these findings suggest that the GT and AT reactions following 5-glucosylation are interchangeable, forming a metabolic grid rather than a single linear pathway[18].
In summary, gentiodelphin derives its stable blue color solely from covalently bound residues, without requiring intermolecular copigments or metal complexation. Since all genes involved in the stabilization mechanism have been identified, reconstructing the gentiodelphin pathway represents a promising strategy for molecular breeding—a concept that aligns perfectly with the third-generation autonomous engineering approach for creating blue flowers in other species.
-
The Lamiaceae family comprises economically important ornamental and medicinal plants. Several genera within this family, such as Dracocephalum, Glechoma, Hyssopus, Lamium, Nepeta, and Salvia, produce purple to blue flowers[20]. These species typically contain violet to violet-blue anthocyanins, including the 3,5-di-O-glucosides of cyanidin, delphinidin, and malvidin, as well as their associated p-coumaroyl, caffeoyl, and malonyl derivatives[12,21]. Several anthocyanin derivatives have been isolated from blue-flowered Salvia species, such as Salvia macrophylla, S. patens, and S. uliginosa[12,22]. The expression of blue color in these Salvia species is governed by the formation of a complex between anthocyanins and apigenin 7,4'-di-O-glucosides (functioning as copigments) in the presence of magnesium ions (Mg2+)[12]. The resulting supramolecular metal-anthocyanin complex (metalloanthocyanin) has been structurally characterized via X-ray crystallographic analysis. Two notable examples of metalloanthocyanin complexes include (1) protodelphin, composed of malonylawobanin [delphinidin 3-O-(6"-p-coumaroylglucoside)-5-O-(6'"-malonylglucoside)], isolated from S. macrophylla and S. patens flowers, and (2) cyanosalvianin, consisting of delphinidin 3-O-(6"-p-coumaroylglucoside)-5-O-(4'"-acetyl-6'"-malonylglucoside), isolated from S. uliginosa flowers[12]. Metalloanthocyanin formation is a widely recognized mechanism for blue coloration in the Lamiaceae family.
A recent study reported the isolation of a polyacylated anthocyanin, namely cyanidin 3-O-[(6-p-coumaroylglucosyl)-(1→4)-(6-p-coumaroylglucoside)]-5-O-(6-malonylglucoside) (Fig. 1b) from the aerial parts of the holy basil (Ocimum tenuiflorum, syn. O. sanctum) cultivar 'Himalayan Sunset Blue', which produces a stable sky-blue tea infusion[23]. This study represents the first record of an anthocyanin containing a glucosyl–coumaroyl–glucosyl–coumaroyl chain in the Lamiaceae family. The hydrophobic interactions between the aromatic acyl groups and the anthocyanin chromophore form a vertically stacked sandwich-type configuration, which stabilizes the blue color. The discovery of this polyacylated anthocyanin offers a novel avenue for future breeding strategies to engineer blue ornamental plants.
Furthermore, other anthocyanins with distinct glycosylation patterns, specifically the 3-O-sophoroside-5-O-glucoside of delphinidin and cyanidin acylated with p-coumaric, ferulic, and/or malonic acids, have been isolated from Ajuga and Tripora, belonging to the subfamily Ajugoideae of the Lamiaceae[24,25]. Tripora divaricata, a monotypic species distributed in Japan, Korea, and China, produces blue-violet flowers containing delphinidin 3-O-[(6-p-coumaroylglucosyl)-(1→2)-(6-p-coumaroylglucoside)]-5-O-(6-malonylglucoside) (Fig. 1c)[26]. The flower color of T. divaricata is attributed to intramolecular copigmentation, as demonstrated by in vitro reconstruction experiments[26]. Additionally, several flavones, including apigenin 7-O-glucuronide, scutellarein 4'-methyl ether 7-O-glucuronide, and acacetin 7-O-glucuronide, have been identified as accessory pigments contributing to the blue hue.
In summary, these findings indicate that, in addition to metal ions, polyacylated anthocyanins and accessory pigments play critical roles in the development of blue color within the Lamiaceae family. These findings also provide valuable insights for the future development of blue-flowered cultivars.
-
The genus Delphinium comprises approximately 650–750 species, primarily distributed in alpine regions in the mountainous regions of the Northern Hemisphere[27]. Among these, Delphinium grandiflorum and D. elatum are the primary species used as horticultural varieties and are widely used for bedding and cut flowers. Recently, their dwarf varieties, which are suitable for pot cultivation (e.g., D. grandiflorum 'Cheer Blue'), have also been developed. Characterized by their deep blue flowers, numerous cultivars have been bred within this genus.
The flower color of Delphinium is strictly determined by anthocyanins, with the characteristic blue hue resulting from the accumulation of delphinidin derivatives (Fig. 1d). The major anthocyanins in this genus are violdelphin and cyanodelphin[13]. Both pigments accumulate in blue flowers; however, the proportion of cyanodelphin positively correlates with the intensity of the blue hue, whereas violdelphin dominance results in a lavender hue[28]. Structurally, both pigments are polyacylated at the 7 position of the delphinidin aglycone[13]. Violdelphin features a structure where benzoyl groups and glucose residues are alternately linked to the 7 position glucose. In contrast, cyanodelphin possesses a branched structure at the 7 position, containing additional glucose and benzoyl attachments. It is hypothesized that these complex structures contribute to the stable deep blue coloration through intramolecular and intermolecular copigmentation (self-association)[13]. Furthermore, Delphinium anthocyanins are not uniformly dissolved in the vacuole but form filamentous aggregates known as anthocyanic vacuolar inclusions (AVIs)[29]. AVIs significantly influence flower color, enhancing intensity and blueness. The polyacylation of anthocyanins—particularly aromatic acylation—is strongly associated with AVI formation, playing a pivotal role in the expression of blue color[29].
Research into the enzymatic machinery responsible for these complex structures has progressed significantly. Currently, the genes encoding the modifying enzymes for violdelphin synthesis have been identified. The biosynthetic pathway involves a spatial separation: synthesis up to delphinidin 3-rutinoside occurs in the cytoplasm, after which the precursor is transported into the vacuole for 7 position polyacylation. Inside the vacuole, an acyl-glucose-dependent anthocyanin 7-glucosyltransferase (AA7GT) attaches a glucose moiety to the 7 position of delphinidin 3-rutinoside, utilizing p-hydroxybenzoyl-glucose (pHBG) as the glucosyl donor[30]. Subsequently, a benzoyl group is transferred to the 6 position of the 7-glucoside by the serine-carboxy-peptidase-like 2 (SCPL2) enzyme, again using pHBG as the acyl donor[30]. Next, the acyl-glucose-dependent anthocyanin 7-p-hydroxybenzoyl-glucoside glucosyltransferase (BGGT) links a glucose residue to the terminal end of the benzoyl group[30]. Finally, violdelphin synthesis is completed when SCPL2 attaches another benzoyl group to this terminal glucose. These findings suggest that complex anthocyanins are efficiently synthesized via vacuolar modification reactions utilizing acyl-glucose as a "zwitter donor" (serving as an acyl and glucosyl donor). Similar mechanisms involving zwitter donors have also been reported in Arabidopsis thaliana[31].
Delphinium is known for its blue flowers. However, pink-flowered varieties derived from D. grandiflorum also exist. These varieties accumulate pelargonidin because of a deficiency in F3'5'H, which is essential for delphinidin synthesis[32]. Additionally, wild species such as D. nudicaule and D. cardinale naturally synthesize pelargonidin derivatives, producing deep red or orange flowers. Generally, Delphinium species cannot synthesize cyanidin derivatives because F3'H is not expressed in the sepals; however, the wild species D. zalil has been found to express F3'H, leading to the creation of interspecific hybrids accumulating cyanidin derivatives[33]. Although the anthocyanins responsible for these reddish hues differ in their 3 position modification (rutinoside vs. malonyl-glucoside), they maintain the same 7 position polyacylation pattern as violdelphin. Recently, novel red Delphinium varieties possessing a pelargonidin 7 position modification similar to cyanodelphin have also been identified[13].
-
Conventional breeding has reached its biological limit for creating blue cultivars in major ornamental crops such as roses, carnations, and chrysanthemums. These species naturally lack F3'5'H, which is essential for synthesizing delphinidin (the blue precursor)[14,15]. Consequently, genetic engineering is the only viable path to overcome this barrier. Although blue color can be generated from cyanidin via complex metal coordination (as seen in Centaurea cyanus), reconstructing such intricate supramolecular complexes involving specific molar ratios of pigments and multiple metal ions (e.g., Fe3+, Mg2+, and Ca2+) has proven technically challenging[34]. Therefore, the predominant strategy for blue coloration is focused on the "delphinidin pathway", introducing F3'5'H to synthesize the blue precursor directly.
Early efforts addressed the substrate limitation through first-generation substrate engineering. Scientists successfully engineered violet carnations (D. caryophyllus) ('Moondust' series) and roses (R. hybrida) ('Applause') by introducing exogenous F3'5'H (from Petunia or Viola) to redirect metabolic flux toward delphinidin[35,36]. In roses, substrate engineering involved RNAi-mediated suppression of endogenous dihydroflavonol 4-reductase (DFR) and introduction of the Iris DFR gene to maximize delphinidin accumulation (up to 100%). However, despite high delphinidin levels, the resulting flower color was violet, indicating that delphinidin production alone is insufficient for true blue coloration. Notably, without stabilizing mechanisms such as polyacylation or copigmentation, delphinidin remains unstable and appears purple in the vacuole. Notably, a distinct strategy proved effective in the orchid P. aphrodite 'Blue Gene'. In contrast to the complex pathway engineering required in roses, the creation of blue orchids relies on precise host compatibility. Initial attempts using dicot F3'5'H genes (e.g., from Petunia) failed to produce visible blue pigment, underscoring the importance of phylogenetic compatibility. The breakthrough involved introducing CcF3'5'H from the monocot Commelina communis, which exhibited superior catalytic efficiency in the monocot host[37]. Crucially, the endogenous Phalaenopsis DFR enzyme displayed sufficient substrate promiscuity, reducing the engineered dihydromyricetin without the need for genetic replacement. The resulting blue-purple color was stabilized by endogenous copigments, demonstrating that a host with a "permissive" metabolic background enables successful blue engineering via a simplified single-gene strategy.
In contrast to the host-dependent success in orchids, the history of engineering blue chrysanthemums illustrates the transition from substrate engineering to stabilization engineering (Fig. 2). Specifically, this finding represents a shift toward second-generation strategies that activate endogenous stabilization mechanisms (Table 1). The initial attempts faced setbacks because of metabolic incompatibility. Huang et al.[38] attempted to introduce ScF3'5'H from Senecio cruentus while silencing the endogenous red pathway (CmF3′H). Unexpectedly, the exogenous enzyme functioned as F3'H in the chrysanthemum host, which led to red cyanidin accumulation rather than blue delphinidin production. Subsequently, Noda et al.[39] successfully introduced CamF3'5'H from Campanula medium and achieved high delphinidin levels (up to 95%), but the flowers exhibited only a violet hue (RHS 77C/83B). Their results showed that delphinidin is unstable in the acidic chrysanthemum vacuole and develops purple coloration. A critical advancement occurred when Noda et al.[40] upgraded the strategy by coexpressing CamF3'5'H with CtA3'5'GT (a UDP-glucose: anthocyanin 3',5'-O-glucosyltransferase from Clitoria ternatea). CtA3'5'GT modified the delphinidin B-ring, generating a novel pigment delphinidin, 3-O-(6"-O-malonyl)glucoside-3',5'-di-O-glucoside (A8, ternatin C5). The success of this approach relied on the synergy between this novel anthocyanin and the host's metabolic background. Although A8 appears violet in isolation, its specific 3',5'-diglycosylation facilitated a precise interaction with endogenous flavone copigments, specifically luteolin 7-O-(6"-O-malonyl)glucoside (C1). This strong intermolecular copigmentation induced a bathochromic shift, yielding a blue phenotype classified as RHS 100C.
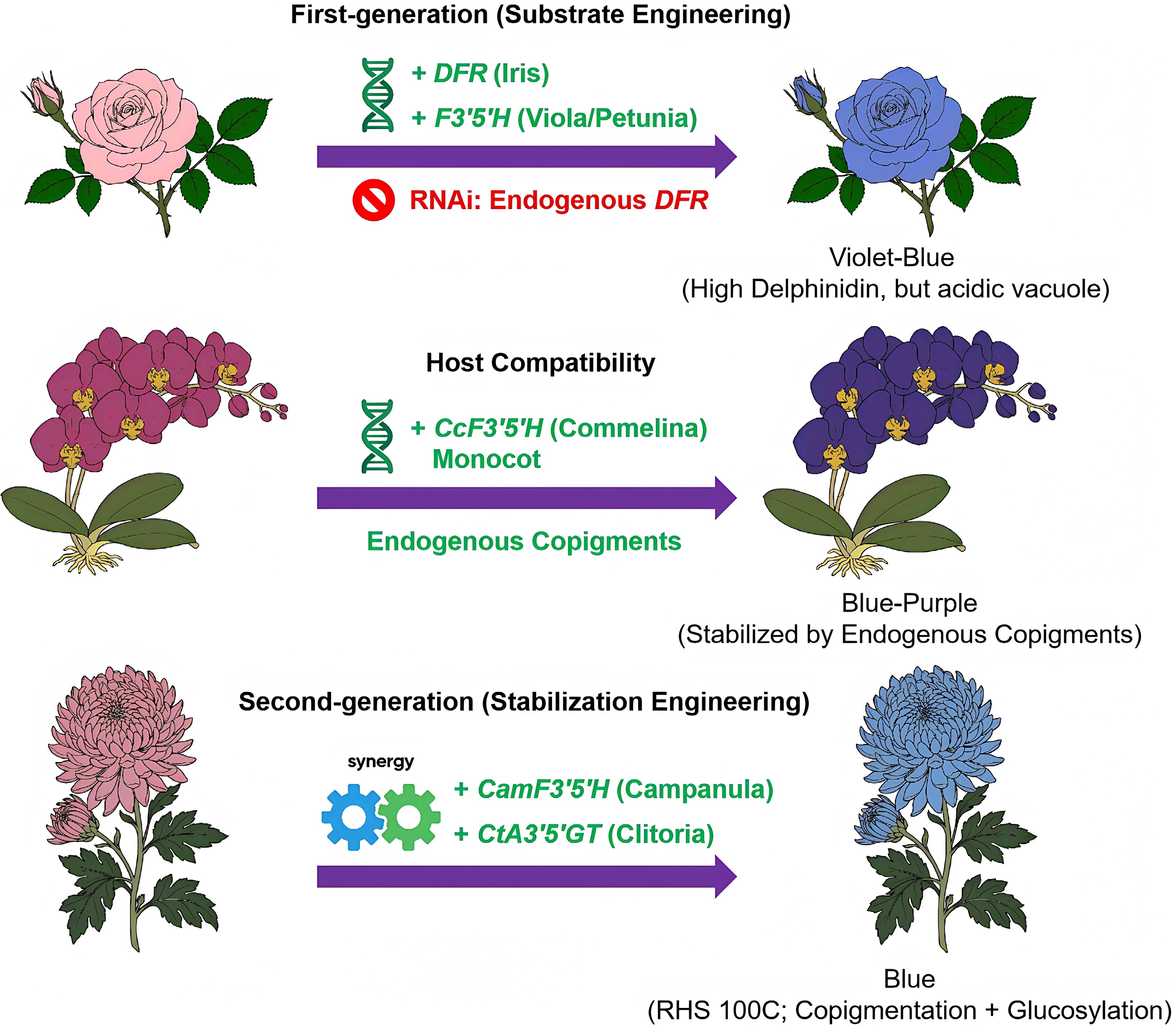
Figure 2.
Evolution of genetic engineering strategies for blue flowers. (Top) First-generation substrate engineering in rose. The introduction of heterologous F3'5'H (from Viola/Petunia) and DFR (from Iris) drives delphinidin accumulation. This strategy includes RNAi-mediated silencing of endogenous DFR. However, acidic vacuoles produce a violet-blue hue rather than a true blue. (Middle) Host compatibility in Phalaenopsis. Expression of a monocot-derived F3′5′H (from Commelina) yields blue-purple flowers. Endogenous copigments specific to orchids stabilize this color. (Bottom) Second-generation stabilization in Chrysanthemum. Co-expressing F3′5′H (from Campanula) and CtA3′5′GT (from Clitoria) produced a bluer phenotype, reported as RHS 100C in selected lines, through the combined effects of copigmentation and glucosylation. The flower illustrations are schematic and summarize the reported phenotypic trends and engineering strategies rather than reproducing the exact color intensity of the original studies.
Table 1. Comparative analysis of genetic engineering strategies for blue/violet coloration in Chrysanthemum.
Host cultivar Introduced genes Key anthocyanins produced Resulting RHS
colorCore mechanism Ref. 'LPi' ScF3'5'H (PCFH)
(Senecio cruentus) CmF3'H RNAiCyanidin Brighter red Failure: ScF3'5'H exhibited only F3'H activity; functional incompatibility [38] '94-765', 'Taihei' CamF3'5'H (Campanula medium) A5, A6 Purple (77C), violet (83B) Success (violet): Achieved high-level accumulation (up to 95%) of the delphinidin substrate in chrysanthemum for the first time [39] 'Taihei', 'Sei Arabella' CamF3'5'H (Campanula medium)
CtA3'5'GT (Clitoria ternatea)
CmF3'H RNAiA8 (ternatin C5)
A7 (preternatin C5)Blue (100C), violet-blue (94B) Success (blue, RHS 100C): B-ring glycosylation of A8; strong intermolecular copigmentation with the endogenous flavone (C1) [40] 'Nannong Fencui' OhF3'5'H (Osteospermum hybrid)
CtA3'5'GT (Clitoria ternatea)A7, A8, A9 Purple-violet (N82D), violet (84C) Success (violet): Confirmed that the gene source for the Noda strategy is replaceable [41] 'Nannong Fencui' CmF3'Hm (T485S mutation)
CtA3'5'GT (Clitoria ternatea)A3, A4 Light purple (77D), very pale purple (N76C) Failure (risk identified): The CmF3'Hm mutation was nonfunctional; Cta3'5'GT exhibited substrate promiscuity, acting on cyanidin [41] A3, cyanidin-3-O-(6"-malonyl) glucoside 3'-O-glucoside; A4, cyanidin-3-O-(3",6"-dimalonyl) glucoside 3'-O-glucoside; A5, delphinidin 3-O-(6"-O-malonyl) glucoside; A6, delphinidin 3-O-(3",6"-di-O-malonyl) glucoside; A7, delphinidin 3,3',5'-tri-O-glucoside; A8, delphinidin 3-O-(6"-O-malonyl) glucoside 3',5'-di-O-glucoside; A9, delphinidin 3-O-(3",6"-dimalonyl) glucoside 3',5'-di-O-glucoside. However, this second-generation strategy is not efficient. Han et al.[41] demonstrated that although the F3'5'H source is interchangeable (e.g., OhF3'5'H from Osteospermum hybrid), CtA3'5'GT exhibits substrate promiscuity. In the absence of efficient delphinidin synthesis, the enzyme glycosylates endogenous cyanidin, diverting metabolic flux toward unintended pink metabolites. This finding underscores the need for precise regulation when relying on the hosts' metabolic backgrounds to avoid metabolic overflow.
-
The second-generation strategy (substrate + copigmentation) has proven successful in chrysanthemum; however, reliance on the host's specific metabolic background (e.g., high flavone levels) limits its universality. To engineer blue flowers in species lacking such copigments (e.g., roses)[36], future research must advance toward a third-generation paradigm: the autonomous reconstruction of complete stabilization mechanisms that are independent of the host environment (Fig. 3).
The first strategy in autonomous engineering is precision pH engineering based on the "energy–color axis", as vacuolar pH is a critical determinant of blue coloration. A shift from acidic to weakly neutral conditions (pH 6.0−7.0) stabilizes the blue quinoidal anion form of delphinidin[10]. For instance, despite successful delphinidin accumulation, the engineered Phalaenopsis ('Blue Gene') retains a violet-blue hue because of its naturally acidic vacuolar environment[37]. This finding highlights that substrate engineering alone is insufficient without concurrent pH homeostasis. A traditional and efficient approach in pH engineering is to overexpress vacuolar proton pumps (e.g., P-ATPases like PH1/PH5) or transporters (NHX1)[42,43]. However, this strategy often alters global cellular ion homeostasis. A recent breakthrough by Yuan et al.[44] offers a more refined approach. They identified PhDC, a gene encoding a subunit of mitochondrial F-ATPase in Petunia, revealing a cross-organelle energy-color regulatory axis. Silencing PhDC decreases mitochondrial adenosine triphosphate (ATP) production, which specifically limits the activity of highly energy-consuming vacuolar H+-pumps, which, in turn, inhibits vacuolar acidification, raising the pH and shifting flower color to blue without disrupting other cellular functions. Breeders can precisely maintain vacuolar pH within the optimal window for blue coloration by fine-tuning PhDC expression (via promoter editing or CRISPR interference [CRISPRi]).
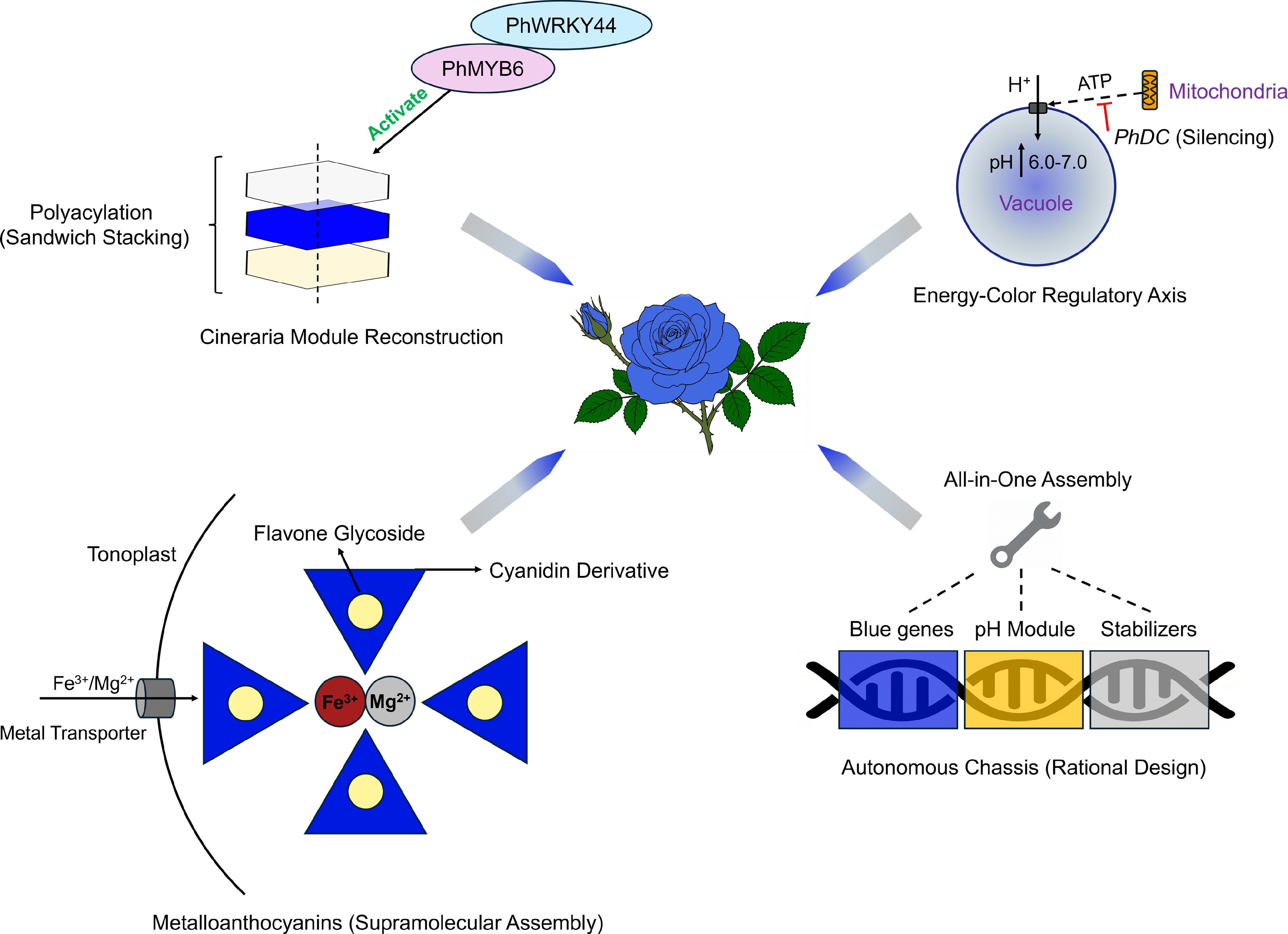
Figure 3.
Third-generation paradigm: future strategies for autonomous true blue flowers. This diagram outlines the four integrated synthetic biology modules to create an ideal true blue rose independent of the host background. (Top left) Polyacylation via the Cineraria module. The transcription factor complex PhMYB6–PhWRKY44 activates the pathway for polyacylated anthocyanins. These molecules form a stable "sandwich" stack that protects the chromophore[7]. (Top right) Engineering pH via the energy-color axis. A novel mitochondrial tuning strategy silences PhDC to limit ATP supply to the vacuolar proton pumps. This strategy raises vacuolar pH to a neutral range (6.0−7.0) and favors blue coloration[44]. (Bottom left) Metalloanthocyanin assembly. Engineered metal transporters (e.g., VIT1/MGT) move Fe3+ and Mg2+ into the vacuole. These ions drive the self-assembly of cyanidin derivatives and flavone glycosides into stable blue complexes. (Bottom right) Autonomous chassis design. This "all-in-one" approach integrates modules for blue gene expression, pH regulation, and structure stabilization. It creates a versatile chassis for universal application.
The second strategy in autonomous engineering involves targeting metal ion homeostasis. This pathway remains promising yet underexplored compared with pH regulation. The blue cornflower (Centaurea cyanus) exemplifies this strategy. Cornflower achieves its stable blue color via cyanidin-based metalloanthocyanins (protocyanin). In this complex, magnesium (Mg2+) stabilizes the self-assembly of anthocyanins and flavone copigments. Then iron (Fe3+) induces the blue color via charge transfer[9]. Notably, roses and chrysanthemums accumulate cyanidin but lack F3'5'H. Theoretically, creating a "protocyanin-like" pigment could generate blue color in these species without altering the anthocyanin backbone. This approach requires engineering metal transporters to enrich specific vacuolar ions. Concurrently, specific flavone glycosides must be introduced to facilitate supramolecular assembly.
The third strategy is reconstructing polyacylation (the ultimate stabilization). The most stable and vivid blue colors in nature (e.g., in Delphinium and Cineraria) rely on polyacylation—the covalent attachment of multiple aromatic acyl groups to the anthocyanin[45]. This attachment creates a "sandwich" stacking structure that protects the chromophore[6]. Reconstructing this complex pathway in heterologous hosts represents a major goal of engineering blue flowers. Cui et al.[7] recently elucidated the complete biosynthetic module for 3′,7-polyacylated delphinidin (cinerarin) in blue Pericallis hybrida (Cineraria). They identified two core enzymes: PhBGLU12 (a 7-position glucosidase) and PhSCPL2 (an acyltransferase). Crucially, they also mapped the upstream regulatory network, discovering that the transcription factor (TF) PhMYB6 and its partner PhWRKY44 form a positive feedback loop that synergistically activates substrate synthesis and polyacylation genes. This discovery enables a synthetic biology approach. Instead of introducing single genes, future strategies can reconstruct the entire Cineraria module (TFs + structural genes) in transgenic hosts. By introducing PhMYB6 and PhWRKY44 alongside the modification enzymes, it might be possible to convert unstable delphinidin into hyper-stable polyacylated pigments. This mechanism would enable the creation of true blue cultivars in a wide range of species, bypassing the limitations of endogenous copigments and pH constraints.
Despite this promise, the application of synthetic biology to blue flower engineering faces several practical challenges. Multigene stacking is likely to impose substantial metabolic and energetic burdens, because stable blue coloration requires the coordinated reconstruction of anthocyanin biosynthesis, structural modification, vacuolar pH regulation, ion transport, and, in some cases, copigment production. In addition, successful engineering depends not only on the presence of these components but also on their quantitative balance; otherwise, metabolic flux may be diverted toward unintended pigments or intermediate accumulation. Repeated transgene cassettes and strong constitutive expression may also increase the risk of gene silencing and phenotypic instability. Furthermore, differences among host species in endogenous flavone pools, acyl donor availability, vacuolar environment, and metal ion homeostasis are likely to affect the portability of synthetic modules. Therefore, future progress will require not only module discovery but also careful optimization of the gene dosage, expression timing, and host compatibility.
In summary, the pursuit of blue coloration in flowers is transitioning from simple gene discovery to rational metabolic design. The field has advanced from first-generation substrate engineering and second-generation host-dependent stabilization to a third generation focused on the autonomous reconstruction of color homeostasis. By using the expanding synthetic biology toolkit, ranging from mitochondrial pH regulators (PhDC) and metal transporters to complex polyacylation modules (pericallis we can now overcome the evolutionary biochemical barriers of nonblue species. The blue rose, once a symbol of the impossible, is no longer merely a poetic aspiration but an approaching scientific reality driven by precision engineering.
-
During the preparation of this work, the authors used Gemini (Google, Mountain View, CA, USA) to generate initial conceptual drafts for the flower illustrations in Figs 2 and 3. These artificial intelligence-generated drafts served solely as visual references. The final illustrations presented in this manuscript were manually redrawn, refined, and colored by the authors using Adobe Illustrator (Version CC 2025; Adobe Systems, San Jose, CA, USA) to ensure botanical accuracy.
-
The authors confirm their contributions to the paper as follows: study conception and design: Zhou LJ, Miyahara T; draft manuscript writing: Zhou LJ, Miyahara T, Sasaki N, Mizuno T; figure preparation: Miyahara T, Sasaki N, Mizuno T, Zhou LJ; manuscript discussion and suggestions: Gao X. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
-
This research was funded by the National Natural Science Foundation of China (Grant No. 32372745), the China Scholarship Council (CSC) under Grant No. 202406850091, and the Japan Society for the Promotion of Science (JSPS) KAKENHI (Grant Nos 16K18564 and 19K15829). We thank the members of the Laboratory of Biochemistry, Graduate School of Horticulture, Chiba University, for their helpful discussions and support.
-
The authors declare that they have no conflicts of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhou LJ, Sasaki N, Mizuno T, Gao X, Miyahara T. 2026. Polyacylated anthocyanins: nature's strategy for blue flower coloration and their application in genetic engineering. Ornamental Plant Research 6: e020 doi: 10.48130/opr-0026-0010 

Polyacylated anthocyanins: nature's strategy for blue flower coloration and their application in genetic engineering
- Received: 05 February 2026
- Revised: 15 April 2026
- Accepted: 22 April 2026
- Published online: 31 May 2026
Abstract: True blue flower coloration remains a rare and highly coveted trait in ornamental horticulture. Its natural occurrence is less than 10% of angiosperm species. The delphinidin derivatives provide the chromophoric basis for blue coloration; however, their structural stability is governed by intricate intramolecular and intermolecular interactions. To preserve blue hues, plants have evolved sophisticated stabilization mechanisms, including the formation of polyacylation, copigmentation, and metalloanthocyanin, as observed in Gentiana, Lamiaceae, and Delphinium. In this review, we integrate the current knowledge of biochemical mechanisms underlying natural blue pigments and evaluate the genetic engineering strategies aimed at creating blue flowers. These efforts to create blue flowers can be classified into three paradigms: (1) First-generation substrate engineering focused on introducing flavonoid 3',5'-hydroxylase (F3'5'H), which often yielded violet rather than true blue phenotypes due to vacuolar instability; (2) second-generation stabilization engineering incorporated endogenous copigments and successfully achieved stable blue coloration, as demonstrated in chrysanthemum; and (3) the emerging third-generation strategy seeks the autonomous reconstruction of complete stabilization modules. We highlight recent advances in synthetic biology that underpin this transition, including mitochondrial pH regulation via PhDC, manipulation of metal ion homeostasis, and the de novo assembly of polyacylation complexes, such as the Cineraria module. Together, these developments mark a shift from single-gene introduction toward rational metabolic design, providing a promising roadmap for overcoming evolutionary constraints in traditionally nonblue crops, such as roses and orchids.
-
Key words:
- Blue flower /
- Genetic engineering /
- Polyacylated anthocyanin /
- Synthetic biology