








-
Salicylic acid (SA), a phenolic phytohormone, functions as a master regulator coordinating plant growth, development, and immunity[1]. It plays critical roles in basal defense, the amplification of local immune responses, and the establishment of systemic acquired resistance (SAR)[1], primarily targeting biotrophic and hemibiotrophic pathogens. Infection with these pathogens activates the SA biosynthesis pathway in plant tissues[2]. Plants have developed two major routes for SA biosynthesis: the ICS (isochorismate synthase) pathway and the PAL (phenylalanine ammonia-lyase) pathway, the latter often referred to as a peroxisomal β-oxidation-dependent route[3]. SA is perceived by receptors of the Nonexpressor of Pathogenesis-Related genes (NPR) family, which, together with TGA-type bZIP transcription factors, promote the expression of SA-responsive genes, including Pathogenesis-Related (PR) genes[4,5]. Beyond canonical immunity, SA has emerged as a pivotal molecular integrator linking environmental nutrient cues to defense reprogramming[6−8].
This integrative capacity positions SA as central to nutrient–immunity crosstalk. Plant nutritional status directly modulates SA biosynthesis, homeostasis, and signaling output. In particular, imbalances or excesses of macronutrients such as nitrogen (N), phosphorus (P), and sulfur (S) affect defense competence and SAR efficacy[9−12]. Reciprocally, SA signaling rewires hormonal networks and metabolic fluxes to influence root architecture, nutrient transporter expression, and intracellular nutrient allocation. These changes form a dynamic feedback loop that fine-tunes the growth–defense trade-off under resource constraints[13,14]. Critically, SA also extends its regulatory role to the rhizosphere. It modulates root exudation profiles and thereby shapes the microbiome. This process promotes the recruitment of beneficial microbes that enhance nutrient mobilization, such as phosphate solubilization, while also modulating immune responsiveness[14]. This tripartite SA–nutrient–microbiome axis plays a key role in enabling plants to cope with concurrent biotic challenges and nutrient fluctuations. This role is particularly important under declining soil fertility and the increasing demands for resilient, resource-efficient agriculture[15].
This review synthesizes recent advances in SA biosynthesis and the evolution of its pathway genes. We then summarize mechanistic crosstalk between SA signaling and the homeostasis of major macronutrients (P, N, S) and discuss reciprocal interactions between SA and the plant microbiome, highlighting how SA-microbe communication is coupled to plant nutrient status. Collectively, we present an integrated SA-nutrient-microbiome framework that can inform strategies to enhance crop resilience and improve nutrient-use efficiency in sustainable agriculture.
-
Numerous bacteria are capable of synthesizing SA, including several plant-associated and pathogenic genera[16,17]. Bacterial SA biosynthesis generally proceeds via two routes. In the most common pathway, ICS converts chorismate to isochorismate, which is then cleaved by isochorismate pyruvate lyase (IPL) to produce SA. Alternatively, some bacteria encode a bifunctional salicylate synthase (SAS) that directly converts chorismate into SA[16,18].
Biosynthesis of SA in plants
-
SA in plants is synthesized via two distinct pathways, namely the ICS pathway and the PAL pathway[19] (Fig. 1). The ICS pathway has been extensively characterized in Arabidopsis thaliana and other Brassicaceae species. In Arabidopsis chloroplasts, ICS1 (also known as salicylic acid induction deficient 2, SID2) catalyzes the conversion of chorismate to isochorismate (IC)[20]. IC is subsequently exported from plastids to the cytosol by enhanced disease susceptibility 5 (EDS5), a chloroplast-envelope MATE transporter. In the cytosol, PBS3 catalyzes an ATP-dependent glutamylation of IC to generate isochorismate-9-glutamate (IC-9-Glu)[20], which is highly unstable and rapidly undergoes nonenzymatic decomposition to yield SA and 2-hydroxy-acryloyl-N-glutamate. In addition to spontaneous decay, IC-9-Glu can be efficiently converted to SA by enhanced pseudomonas susceptibility 1 (EPS1), a BAHD acyltransferase-family protein[21]. It should be noted that the ICS pathway—particularly the two key components PBS3 and EPS1—appears largely restricted to Brassicaceae and is not broadly conserved across other plant lineages[20].
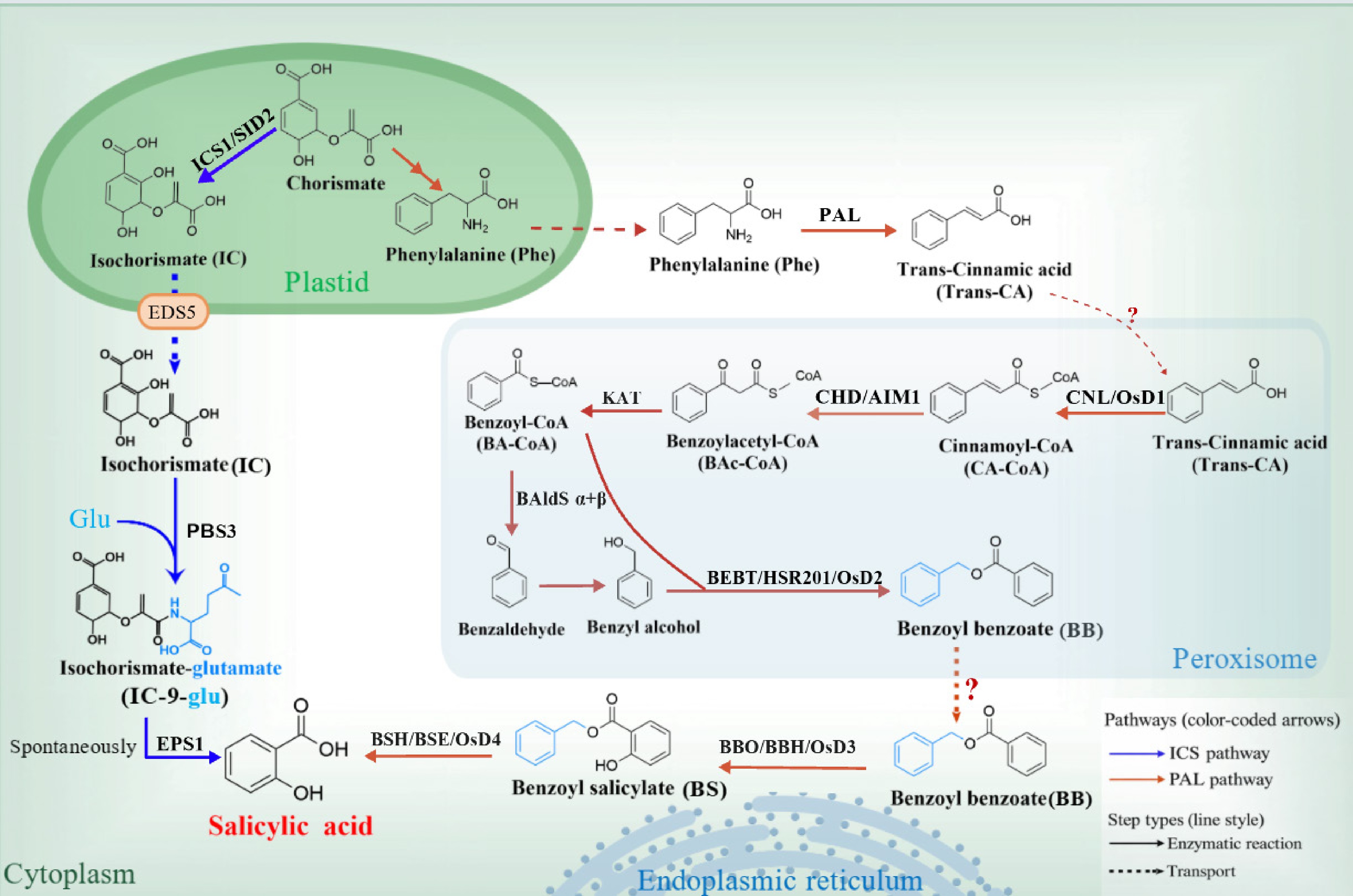
Figure 1.
Pathways for salicylic acid (SA) biosynthesis in plants. SA is produced via the plastid-derived isochorismate synthase (ICS) pathway and the phenylalanine ammonia-lyase (PAL) pathway. In plastids, chorismate is converted to isochorismate by ICS (ICS1/SID2) and exported to the cytosol for PBS3/EPS1-dependent SA formation. In the PAL route, phenylalanine is channeled through cinnamate- and benzoyl-CoA–derived intermediates, with key steps occurring in peroxisomes, followed by conversion of benzyl benzoate (BB) to benzyl salicylate (BS) at the endoplasmic reticulum (ER) by benzyl benzoate hydroxylase (BBH; OsD3 in rice, BBO in tobacco) and subsequent cytosolic hydrolysis of BS by benzyl salicylate hydrolase (BSE; OsD4 in rice, BSH in tobacco) to yield SA. Solid arrows denote enzymatic reactions and dashed arrows indicate metabolite transport; blue and orange arrows represent the ICS and PAL pathways, respectively. This schematic integrates evidence from Arabidopsis, rice, and tobacco.
The PAL pathway contributes substantially to both basal and pathogen-induced SA accumulation in several plant species, including rice and tobacco[22,23]. Multiple enzymes have been implicated in regulating this branch of SA biosynthesis[8,24]. In the phenylalanine (Phe)-derived route, PAL catalyzes the deamination of Phe to produce trans-cinnamic acid (trans-CA)[25]. The traditional view proposes that trans-CA is converted to benzoic acid (BA) via β-oxidation[26,27], and BA is subsequently hydroxylated to SA by a putative benzoic acid 2-hydroxylase (BA2H). However, the existence of BA2H has never been directly demonstrated[28], and the molecular basis of this conversion has remained poorly defined. This long-standing 'BA-to-SA hydroxylation' hypothesis has been fundamentally challenged by recent studies demonstrating that the critical hydroxylation step instead targets benzyl benzoate (BB), leading to a revised model for PAL-derived SA biosynthesis[24,29,30]. In this updated framework, trans-CA is first activated to cinnamoyl-CoA (CA-CoA) by cinnamate-CoA ligase (CNL) in peroxisomes. CA-CoA is subsequently processed by cinnamoyl-CoA hydratase/dehydrogenase (CHD/AIM1) to form benzoylacetyl-CoA (BAc-CoA), which is further converted to benzoyl-CoA (BA-CoA) by 3-ketoacyl-CoA thiolase (KAT)[24,30,31]. BA-CoA is reduced to benzaldehyde by a benzaldehyde synthase complex composed of two subunits (BAldSα+β). The newly formed benzaldehyde is rapidly reduced further to benzyl alcohol[32,33]. BA-CoA and benzyl alcohol then undergo an acyl-transfer reaction to yield benzyl benzoate (BB), catalyzed by benzyl alcohol O-benzoyltransferase HSR201 (BEBT/HSR201/OsD2)[29,30]. BB produced in peroxisomes is exported to the cytosol and subsequently trafficked to the endoplasmic reticulum (ER). At the ER, BB is hydroxylated to benzyl salicylate (BS) by benzyl benzoate hydroxylase (BBH)—encoded by OsD3 in rice and referred to as BBO in tobacco—an ER-resident cytochrome P450 monooxygenase. In the cytosol, BS is ultimately hydrolyzed to SA by benzyl salicylate hydrolase (BSE), encoded by OsD4 in rice and referred to as BSH in tobacco[24,29,30]. Overall, the PAL-derived SA pathway is a multi-compartmental metabolic program in which the chloroplast, peroxisome, ER, and cytosol act in a coordinated manner to channel Phe through CA and BB/BS intermediates toward SA production.
Conservation and evolution of the ICS and PAL pathways for SA biosynthesis
-
Comparative analyses of SA biosynthesis suggest that the ICS gene originated in chlorophyte algae and has remained highly conserved across land plants[34]. In the Brassicaceae ancestor, ICS likely underwent duplication associated with the At-α whole-genome duplication event, giving rise to two distinct clades, represented by Arabidopsis thaliana ICS1 and ICS2. ICS activity for SA biosynthesis was significantly enhanced within Brassicales species that diversified before the divergence of Capparis spinosa and likely after the Carica papaya lineage[20]. During the evolutionary transition toward terrestrial plants, EDS5 emerged in streptophyte algae[34]. Within Brassicales, the ancestral EDS5 gene (EDS5A) duplicated after divergence from the Moringa oleifera and Carica papaya lineages. This duplication generated two Brassicaceae-derived clades. One lineage includes Arabidopsis thaliana EDS5, whereas the other contains Arabidopsis thaliana EDS5H, whose biological function remains to be elucidated[20]. Arabidopsis PBS3 belongs to Clade A, an early-diverging lineage of the GH3 gene family. In Brassicales, Clade A experienced a lineage-specific local duplication following the early divergence of the Carica papaya and Moringa oleifera lineages, giving rise to the A1 and A2 subclades. Among them, the A1a sublineage—containing AtPBS3 and AtGH3.7—acquired glutamate-conjugating activity toward IC, producing IC-9-Glu and thereby promoting downstream SA formation. EPS1 belongs to a Brassicaceae-restricted subgroup of BAHD acyltransferases within the Ib clade. More broadly, the BAHD family has undergone extensive radiation during land plant evolution, generating functionally diverse acyltransferases that are widely distributed across specialized metabolite biosynthetic pathways[20].
Phylogenetic and comparative genomic analyses of enzymes in the PAL-derived SA pathway indicate that most homologues of core pathway components are absent from Rhodophyta, with the exception of AIM1[30]. Phylogenetic evidence indicates that homologues of the OsKAT1/OsKAT2 family and OSD1 originated within the Chlorophyta lineage. This implies that a conserved β-oxidation-dependent metabolic pathway had already evolved and become operational during the early phases of plant evolution[29]. In contrast, homologues of the cytochrome P450 enzyme OSD3 first emerged in the early-diverging vascular plant genus Selaginella and are highly conserved across most tracheophytes. Homologues of OSD2 (BEBT) and OSD4 (BSE) arose later, coincident with the emergence of gymnosperms, and are broadly conserved across seed plants[29]. Collectively, these observations support a stepwise evolutionary assembly of the PAL pathway, with a fully functional module established before the divergence of gymnosperms[30,35]. Notably, despite this conservation, OSD2 (BEBT) and OSD4 (BSE) are absent from Arabidopsis and other sequenced Brassicaceae species[33].
-
P is an essential macronutrient that constitutes nucleic acids, membrane phospholipids, and the universal energy carrier ATP. It plays an indispensable role in plant growth and development[36]. Increasing evidence indicates that a bidirectional regulatory interaction exists between P signaling and SA-mediated basal immunity (Fig. 2). Under P-deficient conditions, SA biosynthesis is enhanced through transcriptional upregulation of PAL genes, leading to elevated SA levels in both rice roots and shoots. This increase in SA promotes the expression of phosphate transporters such as PT2 and PT6, facilitating inorganic phosphate (Pi) translocation from roots to shoots[37]. Consistently, SA biosynthesis mutants (e.g., pal3) exhibit decreased root and shoot soluble Pi content, confirming SA's involvement in P homeostasis[37]. Conversely, exogenous application of SA or immune elicitors significantly enhances cytosolic Pi concentration in Arabidopsis roots[38]. Similarly, exogenous SA application increases rice root and shoot soluble P content under P deficiency[37]. These findings indicate that SA signaling actively regulates P uptake and distribution, while P status reciprocally modulates SA biosynthesis.
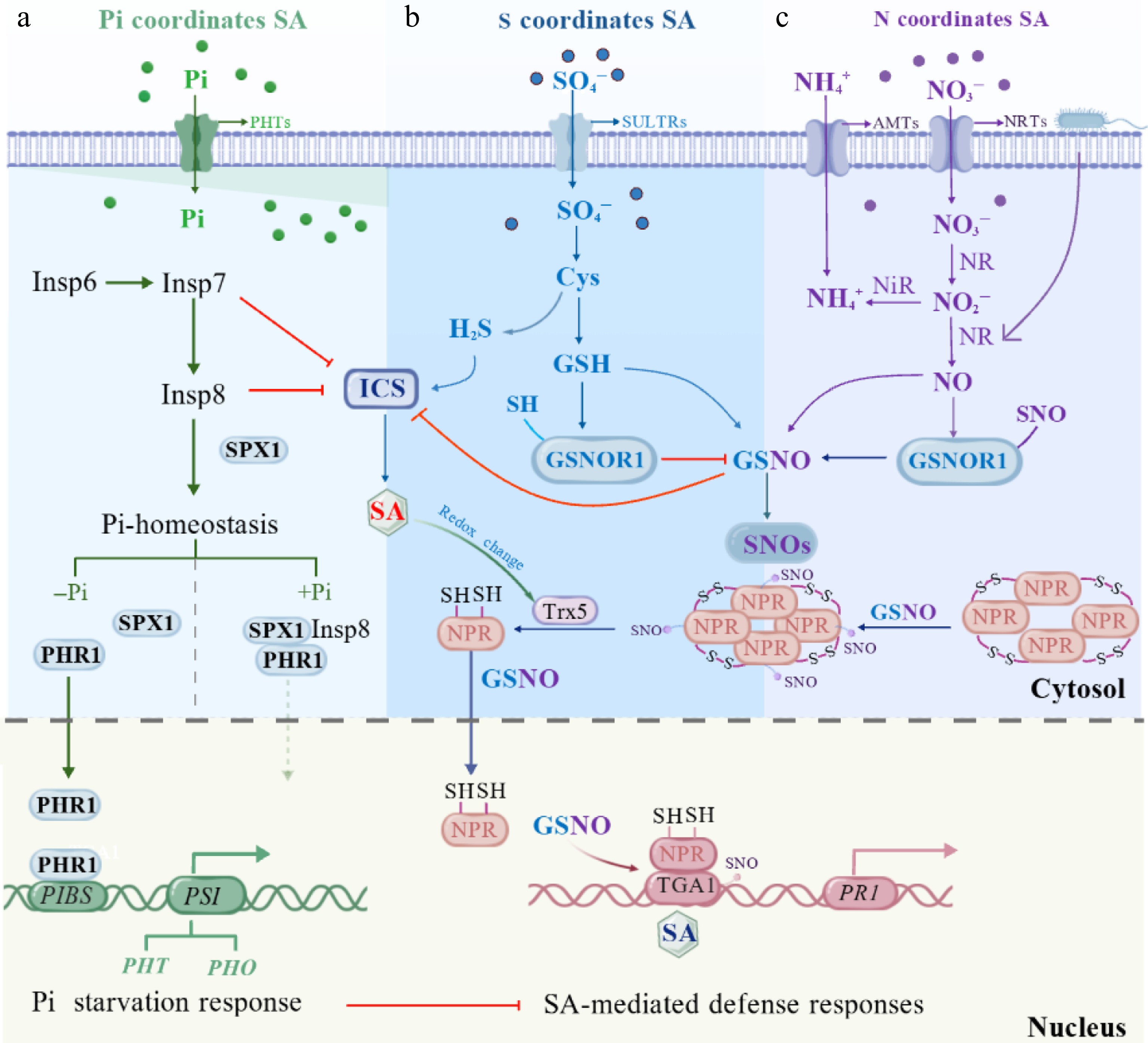
Figure 2.
Integrated model of crosstalk between phosphorus (P), sulfur (S), and nitrogen (N) and SA-mediated plant immunity. (a) Phosphorus availability regulates SA biosynthesis and defense responses through inositol polyphosphate (InsP) signaling and phosphate (Pi) homeostasis. (b) Sulfur assimilation influences SA signaling via cysteine (Cys)-derived redox regulation, controlling glutathione (GSH), hydrogen sulfide (H2S), and GSNOR1-dependent GSNO/SNO homeostasis, which modulates NPR1 activity. (c) Nitrogen-derived nitric oxide (NO) signaling intersects with SA-mediated immunity through GSNO-dependent protein S-nitrosylation, regulating NPR1–TGA–dependent activation of defense genes such as PR1.
The P-signaling network governed by phosphate starvation response (PHR) transcription factors, SPX (for Syg1/Pho81/XPR1) domain-containing proteins, and inositol pyrophosphates (InsP7/InsP8) module, serves as a central hub connecting Pi sensing with SA-mediated immunity[39]. Under Pi-sufficient conditions, InsP8 binds to SPX domains, promoting SPX-PHR complex formation and repressing PHR-mediated activation of P-related genes[36]. Under P deficiency, altered InsP7/InsP8 levels trigger SPX dissociation from PHRs and SPXs degradation. This enables PHR to bind to the P1BS cis-element in P-related gene promoters, thereby activating P uptake and metabolic reprogramming[39,40]. Notably, activation of the PHRs module under P deficiency not only enhances Pi acquisition but also concurrently represses SA-mediated immune responses[39]. This is evidenced by phr1 and phr1phl1 mutants exhibiting enhanced SA-dependent resistance to pathogens like Pseudomonas syringae pv. tomato DC3000 (Pst DC3000) and Hyaloperonospora arabidopsidis Noco2 under low-Pi conditions[41]. Besides, the inositol phosphate metabolic pathway exerts negative regulation on SA immunity. InsP6 is sequentially phosphorylated and pyrophosphorylated by IPK1 and ITPK1 to generate InsP7, which is further converted to InsP8 by VIH1/2 kinases. Under normal growth conditions, IPK1, ITPK1, and VIH2 collectively suppress SA-mediated basal immunity. Accordingly, Arabidopsis ipk1-1, itpk1-2, and vih2-4 mutants exhibit elevated SID2/ICS1 transcript levels and increased free SA accumulation compared to wild-type Col-0 plants. Additionally, ipk1-1 and itpk1-2 mutants display marked NPR1 protein accumulation. These observations collectively indicate that the InsP7/InsP8 pathway functions as a negative regulator of SA immunity[14].
Phosphate transporters (PHTs), traditionally considered as nutrient transporters, have emerged as important regulators of SA-mediated immunity[42,43]. The PHT4 family members, particularly PHT4;1 and PHT4;6, exhibit direct roles in immune signaling[44,45]. Arabidopsis pht4;1-1 mutants display increased susceptibility to Pseudomonas syringae, which can be partially rescued by SA agonist treatment, positioning PHT4;1 functionally upstream of SA signaling[44]. Its function appears independent of known SA biosynthetic genes but may be controlled by the circadian transcription factor CCA1[44]. In contrast, PHT4;6 acts as a negative regulator of SA-mediated defense responses. Loss-of-function PHT4;6 mutations enhance Arabidopsis resistance to Pst DC3000, elevate endogenous SA levels, and compromise salt tolerance[45]. Similarly, the pho2 loss-of-function mutant, which hyperaccumulates P and exhibits elevated ROS levels, shows constitutive activation of SA-associated defense genes even in the absence of infection and displays enhanced resistance to necrotrophic and hemibiotrophic fungal pathogens (e.g., Plectosphaerella cucumerina and Colletotrichum higginsianum)[46].
The relationship between Pi levels and plant immunity is complex and context-dependent. In Arabidopsis, conditions that lead to elevated Pi levels (e.g., miR399 overexpressors, pho2 mutants, or high Pi supply) display enhanced resistance to necrotrophic and hemibiotrophic pathogens[46]. However, in rice, high Pi fertilization and subsequent Pi accumulation increase susceptibility to Magnaporthe oryzae, the causal agent of rice blast disease[47]. This contrasting effect is further demonstrated by OsPT8 overexpression in rice, which suppresses resistance to Magnaporthe oryzae and Xanthomonas oryzae and reduces expression of the defense-related genes like PAL and PBZ1 following pathogen inoculation[48]. These contrasting outcomes underscore that the impact of Pi on immunity is not uniform but is shaped by the specific plant-pathogen system. Overall, how Pi homeostasis is coupled to PHT function and SA signaling, and how this coupling leads to opposite immune outcomes across different species and pathosystems, remains poorly understood; many key mechanisms await further investigation.
Relationship between nitrogen availability and SA-mediated immunity
-
N availability and SA-mediated immunity engage in a tight crosstalk in plants, collectively shaping growth-defense trade-off and pathogen resistance (Fig. 2). In general, high N supply increases plant susceptibility to biotrophic pathogens, while reducing susceptibility to necrotrophs. Tomato plants grown under N-limited conditions exhibit enhanced resistance to pathogens, as evidenced by a reduced number of lesions compared with high-N treatments[8]. Furthermore, plants grown under low N conditions display elevated SA accumulation and up-regulation of defense-related genes, including PAL1, PRs[49]. This phenomenon is commonly explained by two conceptual frameworks: the carbon–nutrient balance hypothesis and the growth–defense balance hypothesis. Together, these frameworks provide a basis for interpreting how nitrogen availability shapes plant immunity[10].
Beyond the overall N supply, the chemical form of N acquired and assimilated by plants—ammonium (NH4+) vs. nitrate (NO3−)—can markedly influence defense responses to pathogens[8,10]. Overall, nitrate-based nutrition generally enhances disease resistance, whereas ammonium-based nutrition frequently compromises it[49−51]. For instance, under low N supply, plants supplied solely with NO3− exhibited stronger resistance to rice stripe virus than those receiving a mixed NO3−/NH4+ supply[49]. Similarly, winter wheat fertilized with nitrate showed significantly lower infection rates by pathogens than plants supplied with NH4+[50]. Plants take up NO3− via nitrate transporters (NRT/NPF and NRT2 families), after which nitrate reductase (NR) reduces it to NO2−—a process that contributes to pathogen-induced nitric oxide (NO) production[52]. This NR-dependent NO production promotes SA accumulation, strengthens the hypersensitive response, and upregulates defense genes. Conversely, NH4+ uptake (via AMT1 transporters) and assimilation bypass NR-mediated steps, suppressing NO generation and attenuating SA-dependent immunity[53,54].
The interaction between SA and NO is a tightly regulated, bidirectional process[55,56]. In Arabidopsis, genetic manipulation of endogenous plant phytoglobin expression to modulate Pseudomonas syringae-induced NO generation results in increased levels of SA accumulation, placing SA downstream of NO in this context[55]. Conversely, exogenous application of SA induces endogenous production of NO and ROS, thereby enhancing innate immunity and overall vigor in tomato plants under Fusarium oxysporum stress[57]. It is well established that NO regulates the expression of the SA receptor gene NPR1 during plant defense responses[58,59]. Furthermore, NO reacts with reduced glutathione (GSH) to form S-nitrosoglutathione (GSNO), an S-nitrosothiol that mediates protein S-nitrosylation (SNO) and regulates protein function as well as gene expression[60]. A key target of GSNO is the SA signaling receptor NPR1. S-nitrosylation of NPR1 stabilizes its oligomeric form and retains it in the cytosol, while simultaneously enhancing the DNA-binding activity of TGA1 (see Section 4.3 for details)[8,60]. Conversely, GSNO-mediated protein S-nitrosylation also exerts negative regulation by inhibiting the expression and enzymatic activity of ICS1, consequently suppressing plant immune signaling[61].
Relationship between sulphur availability and SA-mediated immunity
-
The S nutritional status of plants is tightly integrated with SA signaling and plays a central regulatory role in modulating plant immune responses through the metabolism of cysteine (Cys), glutathione (GSH), and their derived signaling molecules such as H2S and GSNO (Fig. 2). Plant S nutritional status itself functions as a regulatory signal. In Arabidopsis thaliana, S deficiency induces SA-mediated defense responses and strongly activates the SA pathway marker gene PR1, enhances resistance to the hemibiotrophic bacterial pathogen Pst DC3000, but increases susceptibility to the necrotrophic fungal pathogen Botrytis cinerea[62]. In contrast, under S excess conditions, susceptibility to Pst DC3000 increases, whereas resistance to Botrytis cinerea is enhanced, indicating that S status differentially shapes immunity depending on the pathogen lifestyle[62].
S is primarily taken up in the form of sulfate (SO42−) via root sulfate transporter proteins (SULTRs) and subsequently assimilated into Cys through a series of enzymatic reactions[63,64]. As the end product of sulfate assimilation and the first reduced S organic compound, Cys is a central metabolic hub that supplies S for the biosynthesis of defense-related metabolites such as GSH. Increasing evidence indicates that Cys is not only a biosynthetic precursor but also participates in immune regulation[64,65]. Cytosol cysteine can be degraded by cysteine desulfhydrases (e.g., DES/LCD and related isoforms) to release H2S[66], a gaseous signaling molecule that interacts with SA signaling. SA treatment can enhance cysteine desulfhydrase activity, thereby promoting endogenous H2S accumulation. Genetically or pharmacologically elevating H2S levels (e.g., overexpressing the relevant enzymes or applying the H2S donor NaHS) enhances Arabidopsis resistance to P. syringae and induces SA-associated regulators (EDS1, PAD4) and SA-dependent PR genes, whereas reducing H2S levels produces the opposite effect[66−68].
GSH exhibits close functional crosstalk with the SA pathway and may contribute to plant defense against biotic stress through the NPR1-dependent SA signaling cascade. In Nicotiana tabacum, overexpression of the γ-glutamylcysteine synthetase (LeECS) gene in transgenic lines (NtGB) significantly enhanced GSH biosynthesis[69], which was accompanied by elevated SA levels and a marked upregulation of the transcription of both the core SA signaling regulator NPR1 and its downstream defense marker gene NtPR1. In addition, the expression of thioredoxin (NtTRXh) and S-nitrosoglutathione reductase 1 (NtGSNOR1) was also significantly increased. Consequently, these NtGB lines displayed enhanced resistance to infection by P. syringae pv. tabaci[69].
S-nitrosoglutathione reductase 1 (GSNOR1) functions as a critical regulatory node linking GSH metabolism with SA signaling. In the presence of H2O2, GSNOR1 undergoes post-translational activation through direct denitrosylation in a GSH-dependent manner[70]. GSNOR1 catalyzes the NADH-dependent reduction of GSNO to oxidized glutathione (GSSG) and ammonia[65]. By precisely maintaining intracellular levels of both GSNO and SNO proteins, GSNOR1 limits excessive protein S-nitrosylation, thereby positively regulating SA biosynthesis and signaling. In Arabidopsis, the gsnor1-3 loss-of-function mutant exhibits reduced GSNOR activity and elevated S-nitrosothiol levels, whereas plants with higher GSNOR1 activity show reduced SNO accumulation. Upon infection with virulent Pst DC3000, PR1 is rapidly induced in wild-type plants, but displays delayed and attenuated expression in gsnor1-3, indicating impaired SA-mediated defense gene activation. Consistently, gsnor1-3 plants exhibit enhanced susceptibility to the nonhost pathogens Bgt and Psp NPS3121, supporting a positive role for GSNOR1 in SA-mediated immunity[71,72].
In the uninduced state, NPR1 exists predominantly as an oligomer stabilized by redox-sensitive intermolecular disulfide bonds. At this stage, S-nitrosylation of NPR1 by GSNO at cysteine-156 facilitates its oligomerization, thereby contributing to the maintenance of NPR1 protein homeostasis. During pathogen-induced SAR, a change in cellular redox potential occurs[73]. Upon pathogen detection, plants activate defense signaling that upregulates ICS, increasing SA levels. Concurrently, defense-induced ROS and NO shift the GSH/GSSG ratio, elevating reduced GSH in resistant plants. Higher GSH activates GSNOR1, which degrades GSNO. Reduced GSNO lowers protein-SNO, preserving free thiol groups (protein-SH) and further promoting ICS expression and SA synthesis. Accumulated SA, together with thioredoxin (TRX)-mediated reduction of NPR1 cysteine residues, triggers NPR1 monomer release from its oligomeric complex. These active monomers are then transported into the nucleus—facilitated by GSNO—to activate defense gene expression. In the nucleus, NPR1 cooperates with TGA transcription factors to induce SA-responsive genes, including PR genes, whereas GSNO-dependent S-nitrosylation of TGA further enhances transcriptional activation[74]. Specifically, TGA1 is S-nitrosylated at Cys260 and Cys266 by GSNO, and its maximal DNA-binding activity is observed in the presence of both NPR1 and GSNO, supporting a cooperative role for NPR1 and S-nitrosylation in regulating TGA1-mediated transcriptional activation[4,59,69].
The effects of phosphorus, nitrogen, and sulfur availability on SA signaling and plant immune responses are summarized in Table 1.
Table 1. Nutrient status-dependent modulation of SA signaling and plant immune responses.
Nutrient status Key component/mechanism Modulation of SA signaling Defense response/phenotype Species Ref. Pi deficiency PHR1–SPX/PSR signaling;
P1BS elementsSA-responsive genes ↓ PTI ↓; phr1 mutant→Resistance to bacterial and fungal pathogens ↑ A. thaliana [24,41] Pi deficiency PAL3/PAL pathway; OsPT2/6 SA biosynthesis/
accumulation ↑SA→Cell wall phosphorus remobilization ↑ Rice [37] Pi sufficiency miR399–PHO2 module;
SPX–PHR networkRelief of PHR1-mediated repression on SA genes;
Defence gene induction ↓Resistance ↑ (A. thaliana); Susceptibility to Magnaporthe oryzae ↑ (rice) A. thaliana/rice [46,47] Pi transporter activity PHT4;1, CCA1; PHT4;6 Modulates SA synthesis PHT4;1-1 gain→Susceptibility ↑; PHT4;6 loss→Resistance ↑ A. thaliana [44,45] N deficiency PAL1 induction SA accumulation ↑ Resistance to Pst DC3000↑;
Trade-off with growthTomato [8,49] Nitrate (NO3−) NR-dependent NO generation SA accumulation via NO ↑ Resistance to bacterial
pathogens ↑Tomato/tobacco [49,53] Ammonium (NH4+) Reduced NR-dependent NO generation SA accumulation (low NO) ↓ Susceptibility to bacterial pathogens ↑ Tomato/A. thaliana [49,51] NO signaling NO–ROS crosstalk SA biosynthesis and signaling ↑ SAR ↑; Immune responses ↑ A. thaliana [75] S deficiency SA-mediated PR1 expression (NPR1-dependent) SA signaling ↑; SA-dependent genes (PR1) ↑ Resistance to Pst DC3000 ↑; Susceptibility to B. cinerea ↑ A. thaliana [62] S excess SA-mediated PR1 expression; SA–JA crosstalk SA biosynthesis ↓; PR1 ↓ Susceptibility to Pst DC3000 ↑; Resistance to B. cinerea ↑ A. thaliana [62] H2S signaling Cysteine desulfhydrases (LCD/DES1) SA biosynthesis and PR genes ↑ Pathogen resistance ↑; Antioxidant defense ↑ A. thaliana [64,67] Cys and GSH signaling Cys; GSH; GSNO–GSNOR1 system; S-nitrosation Regulates SA signaling & ICS1-dependent SA biosynthesis SA-mediated defense ↑; Resistance to Pst DC3000 ↑ A. thaliana [65,70] ↑ Indicates relative increases or activation, ↓ indicates decrease or suppression. PTI, PAMP-triggered immunity; SAR, systemic acquired resistance; NO, nitric oxide; ROS, reactive oxygen species; JA, jasmonic acid; GSH, glutathione; GSNO, S-nitrosoglutathione; GSNOR1, GSNO reductase 1. -
The root system represents a critical interface for nutrient acquisition and microbial exposure. The assembled root-associated microbiome contributes to both nutrient uptake and protection against pathogens, requiring the plant's innate immune system to strike a balance between permitting colonization by beneficial microbes and restricting potential pathogens[76].
Beyond its canonical role in plant immunity, SA acts as a key cue that structures the rhizosphere microbiome (Fig. 3). Research demonstrates that plants with altered SA synthesis or signaling exhibit root microbiomes with significant shifts in the relative abundance of specific bacterial families compared to wild-type plants[71,77]. These shifts are not merely quantitative changes among dominant strains but represent modulation at the taxonomic family level, with notable enrichment of Actinobacteria and Proteobacteria in SA-deficient mutants. Exogenous SA altered bacterial community composition in both bulk soil and root endophytic samples. Such shifts are consistent with SA exerting both direct and indirect effects on the physiology of a limited subset of community members, with additional changes potentially mediated via community interactions[71].
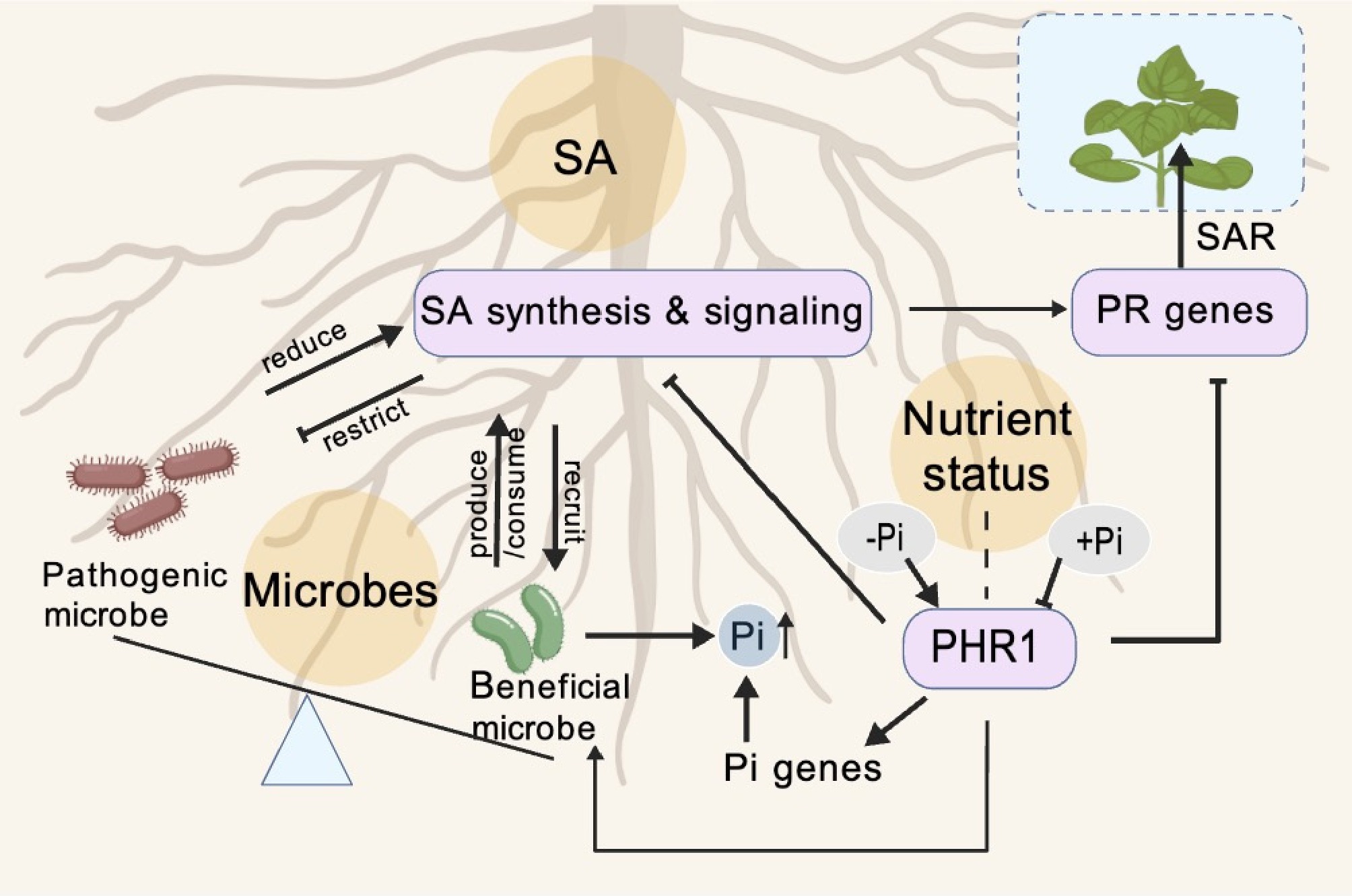
Figure 3.
Model of SA-mediated integration of plant immunity, root microbiome assembly, and phosphate status. SA produced in roots shapes the root-associated microbiome by favoring beneficial microbes and constraining pathogens, while beneficial microbes can in turn produce SA and enhance PR gene expression and SAR. Pi status regulates PHR1, which activates Pi-responsive genes and Pi uptake and influences the recruitment of beneficial, Pi-scavenging microbes, while modulating SA-dependent immune outputs, thereby linking nutrient status, microbiome composition, and plant defense.
Microbiome assembly is partly driven by differential utilization of SA among bacteria, which variably exploit it as either an extracellular signal or a carbon substrate, thereby restructuring the community. For instance, Terracoccus sp. 273 increased in abundance in SA-treated soil and root endophytic samples and exhibited SA-stimulated growth in culture, despite lacking annotated genes for SA catabolism; in contrast, Streptomyces sp. 303 grew on minimal agar with 0.5 mM SA as the sole carbon source[71]. The differential responses to SA are a key driver of community structure since the abundance of some root-colonizing bacterial families increases at the expense of others[76]. Certain plant-colonizing microorganisms promote host fitness by mobilizing nutrients and suppressing pathogens through competition, antibiosis, and lytic enzymes[78−81]. They can also indirectly benefit plants by enhancing resistance responses against pathogens[81], and SA-associated interactions represent one specific facet of such beneficial relationships. However, the ability to metabolize SA is not restricted to beneficial microorganisms. Some plant-associated bacteria and pathogens also possess SA degradation pathways, which may weaken host SA signaling and facilitate immune evasion. For example, Ralstonia solanacearum degrades plant-derived SA to protect itself from inhibitory levels of this compound and to enhance its virulence in plant hosts such as tobacco that rely on SA as a defense signaling molecule[82]. Similarly, to evade plant immunity, certain pathogens can degrade SA through metabolic enzymes such as salicylate hydroxylase, thereby interfering with host immune signaling pathways and promoting successful infection and pathogenicity[83]. Therefore, the ecological roles of SA-utilizing microorganisms in plants are likely to be highly context-dependent. Their specific effects depend on multiple factors, including the population dynamics of pathogens, the genetic backgrounds of both the pathogen and the host, biotic and abiotic conditions, and the composition and diversity of the plant microbiota[84,85].
The plant-microbe dialogue is bidirectional. Certain beneficial microbes can actively synthesize SA, thereby amplifying the host plant's defense capacity. Bacterial species such as Pseudomonas fluorescens strain CHA0, Pseudomonas aeruginosa strain 7NSK2, and Serratia marcescens strain 90–166 are well-documented SA producers[72,86,87]. Colonization by these bacteria elevates endogenous SA levels and enhances systemic resistance. Screening studies that identified SA-producing endophytes, like Pseudomonas tremae, which colonizes Nicotiana benthamiana, enhance host SA accumulation and confer resistance to tobacco wildfire disease[17]. Microbial SA production therefore, adds an additional regulatory layer to the plant SA signaling network.
Within the framework of SA-mediated microbiome modulation, plant Pi status serves as a pivotal nutritional cue that shapes plant-microbe dialogue through the regulation of SA defense signaling. Under Pi deficiency, SA-based defenses are attenuated, reflecting a strategic reallocation of resources toward Pi acquisition. This suppression facilitates associations with plant growth-promoting bacteria, thereby enhancing Pi uptake[40,88]. Conversely, excessive phosphorus supply can also suppress defense gene expression and increase susceptibility to pathogens such as Magnaporthe oryzae in rice[12], indicating a non-linear relationship where both deficiency and surplus can compromise immunity via different mechanisms. The recruited microbiome under Pi stress likely mediates this nutrient-dependent defense trade-off[88]. Down-regulation of SA signaling under Pi starvation may facilitate colonization by specific beneficial bacterial families that aid in Pi mobilization, while these microbes may in turn influence the local SA milieu[71].
A molecular nexus linking phosphate signaling with plant immunity was revealed by the identification of PHR1 as a central integrator[41]. Beyond its role in activating Pi uptake and mobilization genes, PHR1 actively suppresses components of the plant immune system. This suppression is both direct—evidenced by PHR1 binding to the promoters of defense-related genes—and functional, as shown by enhanced pathogen resistance in phr1 mutants. Transcriptomic analyses reveal that PHR1 modulates key immune hubs, including gene clusters associated with SA and JA pathways[41]. Crucially, this PHR1-mediated coordination operates at the plant-microbiome interface. Under Pi limitation, PHR1 activity fosters a root microbiome configuration that enhances Pi-scavenging capacity, while simultaneously repressing microbially triggered immune outputs[89]. Thus, PHR1 lies at the heart of a strategic adaptation that reallocates resources from defense to nutritional stress mitigation, a process inherently mediated through its dual regulation of both plant innate immunity and microbiome assembly[89].
Taken together, SA emerges as a central currency governing microbiome assembly, microbial metabolism, and interkingdom signaling. This SA-microbe dialogue is set within the overarching context of plant nutrient status (e.g., Pi starvation), which dictates the strategic prioritization of resources between immunity and nutrition. How plants precisely discriminate and integrate signals promoting mutualism versus defense at this dynamic interface between nutrition, microbiota, and immunity remains a fundamental and largely unresolved question.
-
SA represents far more than a simple defense hormone; it functions as a sophisticated signaling hub that integrates environmental cues, nutrient status, and microbial interactions to optimize plant fitness. This review has synthesized current understanding of SA biosynthesis through both the ICS and PAL pathways in plants, revealing evolutionary conservation and divergence in this critical signaling system. The reciprocal relationships between SA and essential nutrients demonstrate a sophisticated resource allocation strategy, where plants dynamically balance growth and defense. Under nutrient limitation, plants often prioritize nutrient acquisition over defense responses, mediated by transcription factors such as PHR1 that simultaneously activate phosphate starvation responses while suppressing SA signaling. Conversely, nutrient repletion could prime SA-mediated immunity as evidenced by the contrasting effects of high phosphate on resistance to different pathogen lifestyles. The tripartite interactions among nutrients, microbiome, and SA signaling further underscore the complexity of plant environmental adaptation.
Several key gaps remain in our understanding of how SA integrates nutrient signaling with immune outputs and microbiome assembly. Future research should prioritize several interconnected avenues: First, the functional divergence between the ICS and PAL biosynthetic pathways demands systematic investigation. Do these parallel pathways differentially regulate nutrient acquisition under varying immune challenges? Second, the phenotypic consequences of mutations in SA pathway components on microbiome assembly require comprehensive characterization under controlled nutrient regimes. Third, nutrient transporters and signaling molecules likely serve as recruitment signals for microbiome partners that modulate SA outputs. Future work must elucidate whether transporters for nitrogen, sulfur, or iron directly or indirectly shape microbiome composition that subsequently influences SA biosynthesis through either the ICS or PAL pathway. Finally, a fundamental challenge lies in understanding how plants discriminate between symbiotic and pathogenic microbes at the molecular interface where nutrient exchange and defense activation converge. How does SA signaling integrate with nutrient receptor systems to permit beneficial interactions while maintaining defense capacity? Resolving this question requires innovative approaches to track real-time signaling dynamics at plant-microbe contact points under varying nutrient conditions.
This work was supported by the National Natural Science Foundation of China (Grant No. 32102478).
-
The authors confirm their contributions to the paper as follows: study conception and design: Wang L, Jia X; draft manuscript preparation: Yu J, Zhao Y, Hu G, Fu J, Xu L, Wang L, Jia X, Yi K. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Jiahong Yu, Yule Zhao
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Chongqing University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yu J, Zhao Y, Hu G, Fu J, Xu L, et al. 2026. Interplay between salicylic acid and plant nutrient homeostasis. Plant Hormones 2: e013 doi: 10.48130/ph-0026-0009 

Interplay between salicylic acid and plant nutrient homeostasis
- Received: 02 February 2026
- Revised: 18 March 2026
- Accepted: 30 March 2026
- Published online: 22 May 2026
Abstract: Global agricultural intensification fosters crop yields by increasing mineral nutrient inputs; the resulting widespread nutrient imbalances can compromise plant immune responses. Salicylic acid (SA) serves as a regulator of plant defense responses against biotrophic pathogens and environmental stresses. This review summarizes current knowledge on SA biosynthetic pathways in plants, providing a detailed analysis of the distribution, function, and evolutionary history of the isochorismate synthase (ICS) and phenylalanine ammonia-lyase (PAL) pathways. This review examines the interconnections between SA and the homeostasis of phosphorus, nitrogen, and sulfur. We explore how phosphate starvation responses antagonize SA immunity and how nitrogen forms alongside sulfur metabolites to fine-tune SA-mediated resistance. Additionally, we discuss the role of SA in shaping the rhizosphere microbiome. Understanding the interplay between SA, nutrient signaling, and the microbiota is pivotal for developing sustainable agricultural strategies that simultaneously optimize crop yield, disease resistance, and nutrient use efficiency.
-
Key words:
- Salicylic acid /
- Plant nutrients /
- Microbiome /
- Plant immunity