








-
Blueberries, one of the world's most economically significant berry crops, are loved by consumers for their unique palatability and excellent nutritional attributes[1,2]. Nevertheless, postharvest softening, mechanical injury, and nutritional degradation frequently occur during storage, thereby reducing the economic benefits and hindering sustainable growth of the blueberry supply chain[3,4]. Resolving blueberry softening has always been a focus of attention in the industry, as it impedes the postharvest storage, distribution, and consumer acceptance of blueberries[5,6]. Accordingly, it is requisite to study the potential mechanism of postharvest softening of blueberries, to extend the storage life and to promote the industrial advancement of blueberries. Recent times have witnessed the exploration and utilization of many new preservation technologies, such as the combination of cryogenic stockpiling and irradiation maintenance[7], and edible coatings[8]. However, these technologies not only require precise control of various conditions but also have shortcomings in terms of stability and timeliness. Thus, the development of safe and reliable preservation techniques is a prerequisite for extending the storage life of blueberry fruit.
MT (melatonin), as a safe, efficient, natural, and easily obtainable bioactive substance, has been increasingly used in the field of preservation. Meanwhile, a growing body of research indicates that exogenous MT treatment exerts multifaceted functions throughout postharvest handling for fruit freshness retention by activating antioxidant defense systems, mitigating cold damage, suppressing pathogenic infections, hindering softening progression, and quality decline[9−11]. MT can crosstalk with multiple plant hormones to jointly affect the development, maturation, and aging processes of fruits[12]. MT treatment can slow down the decrease in postharvest firmness of jujube fruits, activate the AsA-GSH cycle, inhibit the enzyme activity, thereby improving the quality traits of jujube fruits and delaying the softening performance[10]. Postharvest MT treatment restrained the enzymatic activities of PG, β-gal, and PE, delayed pectin degradation, and thus suppressed postharvest texture loss in mango fruit[11]. Research on blueberry fruits has indicated that postharvest MT treatment can maintain nutrient content, reduce ROS accumulation and lipid peroxidation of membranes, and thereby improve blueberry postharvest storability[13].
As an essential plant hormone, abscisic acid (ABA) is indispensable to the plant expansion regulation and ontogenic programs[14,15]. In climacteric fruits, the presence of ABA can increase the cell wall-degrading capacity, promote mature fruit softening, and indirectly regulate the fruit softening process by accelerating ethylene biosynthesis. Accumulating studies suggest that ABA is instrumental in modulating postharvest senescence and softening in non-climacteric fruits. An increase in ABA content accelerates the progression of fruit ripening, whereas a decline in ABA content exerts the opposite effect[16]. Scholarly investigations have revealed that exogenous ABA treatment in blueberries promotes softening and coloration during ripening[17]. Previous studies from our research group have indicated that the decrease in firmness of postharvest Bluecrop blueberries is significantly correlated with the accumulation of endogenous ABA. Exogenous ABA treatment accelerates cell wall degradation and fruit softening, whereas treatment with an ABA inhibitor delays this process[14]. IAA is a primary endogenous auxin, modulating fruit ripening and senescence through interactions with other hormones, including ethylene and ABA[18]. Research has demonstrated that IAA may participate in ripening and senescence-associated fruit firmness loss by affecting the accumulation of ABA in fruit. During grape ripening, exogenous IAA treatment can inhibit ABA accumulation, downregulating the expression of cell wall-degrading genes, thereby delaying the ripening process[19]. Besides, several studies have established that key factors in the IAA signaling pathway affect ABA homeostasis in plants by mediating ABA biosynthesis or signaling, and modulate plant physiological and developmental processes. Existing studies have demonstrated that the auxin-responsive factor PavARF8, whose transcription level is inhibited by ABA, can bind to the promoter of ABA synthesis gene PavNCED1, regulate its expression, and participate in ABA-regulated fruit softening[20].
Tan et al.[21] uncovered that MT inhibits ABA accumulation, suppressing postharvest senescence in fresh produce by coordinating ABA biosynthesis, degradation, and signal transduction. Although there have been numerous studies on the roles of ABA and IAA in fruit ripening and softening, covering fruits with climacteric fruits (such as tomatoes[22] and bananas[23]) as well as non-climacteric fruits (such as grapes[24]), there is currently no in-depth exploration of how melatonin regulates both ABA metabolism and IAA signaling to delay softening in harvested blueberries. On this basis, this study intends to ascertain the function of exogenous MT in ABA biosynthesis, degradation, signaling, as well as IAA signaling, in postharvest blueberries, and to elucidate the mechanism by which it suppresses postharvest softening.
-
The blueberry cultivar used in the experiment was Bluecrop, which was obtained directly from the Shisheng Blueberry Base in Shenyang, Liaoning Province, China. Blueberries of homogeneous size, optimal maturity, and unblemished appearance were selected and quickly transported back to the laboratory under low temperature conditions after harvest. They were pre-cooled for 5 h at 4 °C to eliminate field heat, and then randomly divided into four groups, with three parallel replicates per group. Fruits were treated with 50, 100, and 150 µM melatonin (MT), with 2% ethanol in each solution. The control group (CK) was treated with distilled water plus 2% ethanol. The four solutions were evenly sprayed on blueberry surfaces at room temperature (20 ± 0.5) °C in the dark, followed by 12 h of natural air-drying. After drying, the blueberries were placed in a plastic box, then put into a polyethylene (PE) fresh-keeping bag with a thickness of 0.02 mm, and subsequently placed in a biological fresh-keeping cabinet at (20 ± 0.5) °C with a relative humidity of 80% to 85%. Various indicators were measured every other day. Sufficient blueberry fruits were taken, instantly snap-frozen in liquid nitrogen, and stored at −80 °C until further analysis of relevant physiological parameters. The experiment was performed with three biological replicates.
Measurement of firmness
-
Use a texture analyzer to measure the firmness of blueberries, randomly select blueberry fruits, and determine two symmetrical parts of each fruit. Measure the firmness of its pericarp and flesh. Data are presented in Newtons (N).
Pectin content
-
According to the method of Sun et al.[25], the sample was boiled and washed with 95% ethanol to remove soluble sugars. The precipitate was then extracted with distilled water in a 50 °C water bath to obtain soluble pectin. After centrifugation, the supernatant was diluted to a certain volume. The remaining precipitate was hydrolyzed in a 0.5 mol/L sulfuric acid boiling water bath for 1 h to extract the protopectin. After centrifugation, the supernatant was diluted to a certain volume for testing. Aliquots of the test solution and the standard solutions of galacturonic acid (0–100 μg) were taken separately. Concentrated sulfuric acid and carbazole ethanol solution were added for color development, and the absorbance was measured at 530 nm. The mass of galacturonic acid was calculated based on the standard curve y = 0.0031x + 0.0662 (R2 = 0.9996). The result is expressed as the mass fraction of galacturonic acid (%).
Cell wall-degrading enzyme activity
-
A 10 g portion of fruit tissue was ground in the presence of 20 mL ice-cold 95% ethanol. The mixture was first kept at 4 °C for 10 min, then centrifuged at 12,000 g for 20 min. The supernatant was discarded, and the pellet was rinsed with 10 mL of 80% ethanol by shaking; this rinsing step was repeated once more. The cleaned pellet was then resuspended in cold extraction buffer and incubated at 4 °C for 20 min. After centrifugation, the supernatant was obtained and applied to assess enzyme activity. The determination methods for polygalacturonase (PG), endocellulase (Cx), β-glucosidase (β-Glu), and β-galactosidase (β-Gal) activities were based on Chen et al.[26], Sun et al.[25], Tang et al.[10], and Song et al.[27], respectively, each with minor modifications.
Endogenous ABA and IAA contents
-
About 1 g of blueberry fruit tissue was disrupted in 9 mL of ice-cold PBS (pH 7.4). After centrifugation at 5,000 rpm for 15 min, the resulting supernatant was harvested for further evaluation. ABA and IAA levels were quantified using a commercial kit, with absorbance measured at 450 nm. Hormone contents (μg·L−1) were calculated by plugging the measured absorbance values into the corresponding calibration curves.
Extraction of RNA, synthesis of cDNA, and analysis of RT qPCR
-
The RNA of blueberry fruit was purified utilizing an OminiPlant RNA Kit. Its quality was checked by agarose gel electrophoresis. Subsequently, cDNA was synthesized using RNA as a template and used for RT-qPCR analysis. The primer sequences for cell wall metabolism genes were based on Song et al.[27]. Primers targeting key genes implicated in ABA biosynthesis, degradation, and signal transduction were designed based on Li et al.[28]. According to the transcriptome sequencing results of blueberry softening at room temperature, differentially expressed VcTIR1, VcARF1, VcIAA9, VcIAA27, VcGH3.1, and VcSAUR gene sequences were identified. Primers for qRT-PCR were constructed employing Primer Premier 5.0 software (Table 1). The expression level was computed via the 2−ΔΔCᴛ method.
Table 1. Primers for real-time q-PCR assays.
Gene Forward primer (5'–3') Reverse primer (5'–3') Actin ACTACCATCCACTCTATCACCG AACACCTTACCAACAGCCTTG VcPG1 ACCACCAACCGCATTA AAGCGTCAGGTGAGTAAG VcPG3 GCTATGGCATTACAAAGACAA CCATGAATGGTACTGGTGGA Vcβ-Gal CTTCTCTCTCTTCTCGCCGC CGAATGCCTTTGCCCTCAAC Vcβ-Glu TCGACCGAAGCGTCGCTACT GTCCACCATGTCCGCCCAA VcCx1 CGCGTTCAGCCTTTACCTTA TTCGCCAGTCGCTATCCCCT VcPE CCGACCTCTCCGCCTTCT CGCCTTCACCGGTTTCAA VcPL CGATGGGCTGATTGATGC TGCCTTGGCTGTTGATGGT VcNCED1 GATTTCCCTCCAACAGCCTACT CGGACATCTTGAACACCACCT VcNCED6 CATCAGGTGGTGTTCAA GACTCGTCCGTATCATTT VcZEP1 CGGCCCGATTCAACTCCT TCCGGTCTCCGGTAACAC VcAAO3 TGGACAAGAGGGCTACCG GCTGGCTGGACATTTGAG VcCYP707A TATGAATTGGGCGGTAGG AAAGCGATTATGGGAACG VcPYL TTGATGACGAGGAACA CAACCGCTCAGATACA VcSnRK2 CACGCTGTTGGATGGA TGGTTGGTGAGCCTTG VcABF CGGTGACTGCCATTAG CCCTCCATTCCCATAC VcABI5 ATTGGGCGTGTTCGTC TTTGCTCCCTTTCTCG VcPP2C TGGTGGAGGAGTCAGGGAT TAGGGTGATTGCGAAT VcTIR1 GCTTCGTTCCTCTATCTCCCTG GCCCTCCAATCCCTTGTCTT VcARF1 ATGCTGAATCCGATGGGTAT CCGCAGAGTTCGCCTTTA VcIAA9 ATGTCTCCACCACGGATAGG GGGAACCAGGTAGACCAAGC VcIAA27 AAAGGCACAGGTAGTGGGATG GGGCTGATGAAAGTTGGAC VcGH3.1 TTATCGGCTCATCCCATCT GGTCTTTGTCTCGGACTTCAC VcSAUR GCCAAGTCCCTCTGCTGTTC AGTCGCCGTCTTTCTGTCCC Experimental design and statistical methods
-
All experimental data underwent one-way analysis of variance (ANOVA) using SPSS version 26.0. Independent sample t-tests were conducted at each time point, with * indicating significant differences (p < 0.05) and ** indicating highly significant differences (p < 0.01). All charts were plotted using Origin 2021 software.
-
Fruit firmness is a key parameter for evaluating postharvest qualitative traits and intuitively reveals the degree of fruit softening. As shown in Fig. 1a, b, both fruit and flesh firmness of all four groups declined continuously during storage. Blueberries treated with 50 or 100 μM MT maintained significantly higher firmness than the control (p < 0.05), with the 100 μM treatment being the most effective. Additionally, from day 4 through the end of storage, flesh firmness in all MT-treated groups remained consistently higher than in the control, and again the 100 μM MT treatment had the strongest effect (p < 0.05).
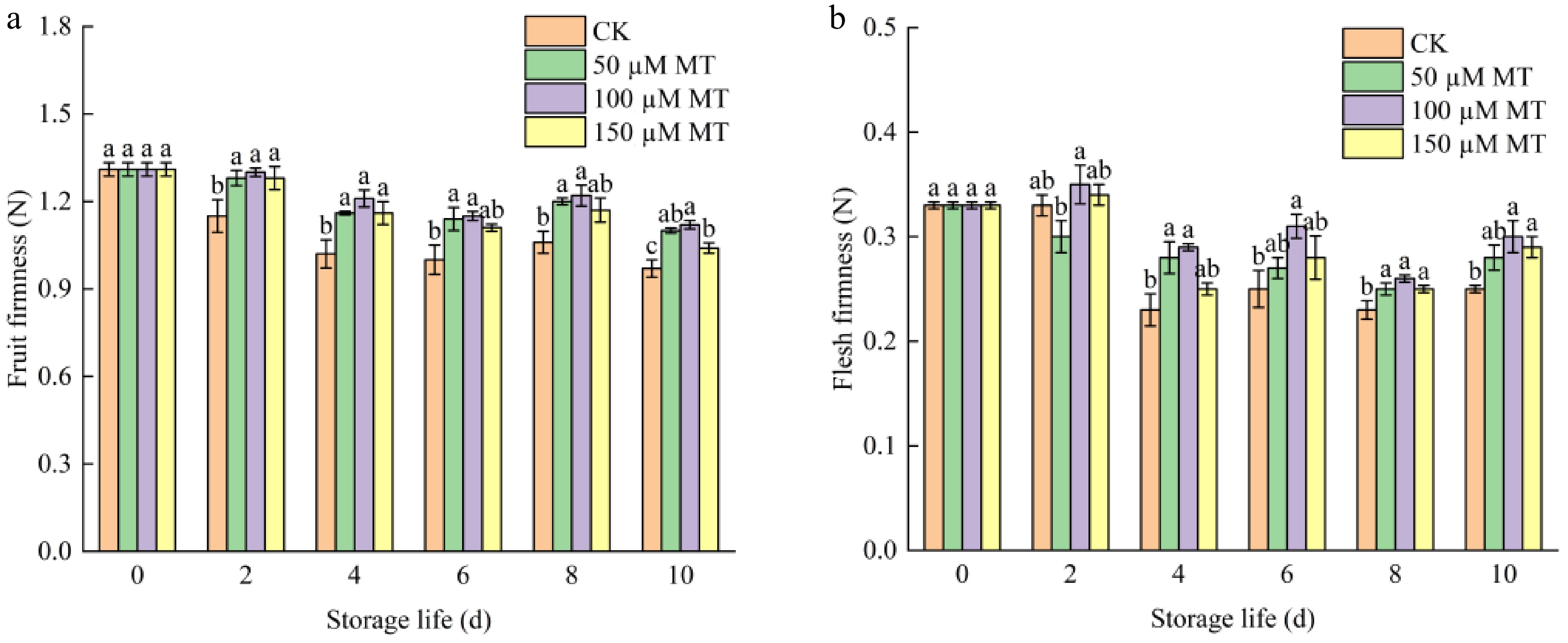
Figure 1.
Effects of MT treatments at different concentrations on (a) fruit firmness and (b) flesh firmness of blueberry fruit during room temperature storage. Significant differences among treatments are marked with * (p < 0.05) and ** (p < 0.01).
Effects of exogenous MT on pectin content of blueberries
-
The content of protopectin (Fig. 2a) decreased gradually in all blueberry fruits during storage, particularly within the initial 6 d. MT treatment significantly retarded this decrease in contrast to the control (p < 0.05), with highly marked differences detected on days 2, 4, and 10 (p < 0.01). Conversely, the soluble pectin content (Fig. 2b) underwent a marked rise; MT-treated fruits maintained markedly lower soluble pectin levels than the untreated fruits throughout the shelf life, except on day 8 (p < 0.05).
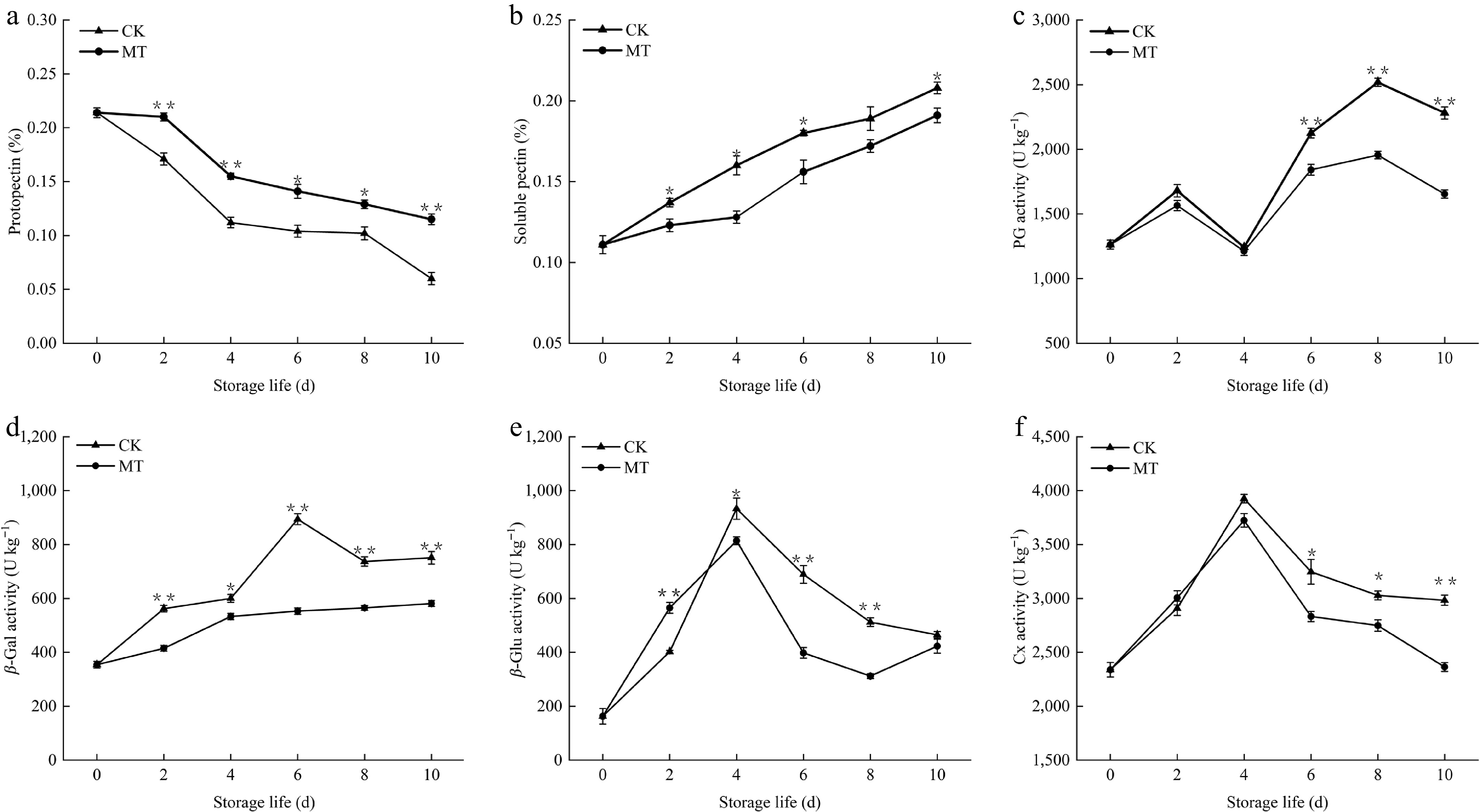
Figure 2.
Effects of exogenous MT on (a) protopectin, (b) soluble pectin, and (c) activities of PG, (d) β-Gal, (e) β-Glu, and (f) Cx of blueberry fruit during room temperature storage. Significant differences among treatments are marked with * (p < 0.05) and ** (p < 0.01).
Effects of exogenous MT on cell wall metabolism-related enzyme activities and gene expression in blueberries
-
PG activity in both groups followed a similar pattern, gradually increasing from day 4 to 8 and then decreasing rapidly (Fig. 2c). From day 6 to 10, PG activity in the MT-treated group was markedly reduced vs that in the control group (p < 0.01).
The β-Gal activity in the control group increased rapidly to a peak on day 4 and then declined gradually throughout the remaining storage period (Fig. 2d). MT treatment greatly suppressed β-Gal activity in comparison with untreated fruit (p < 0.05). At the termination of the storage life, the activity in the control increased to 751 µmol h−1 g−1; the treatment group showed a much lower activity, at 581 µmol h−1 g−1, representing a highly significant difference (p < 0.01).
Interestingly, within the first 8 d of storage, β-Glu activity in both groups of blueberries gradually increased to a peak and then began to decrease (Fig. 2e). By day 4, the activity levels of both groups reached their maximum values, with the control group at 933 µg h−1 g−1 and the treatment group at 814 µg h−1 g−1, which revealed a distinct variation between the two groups (p < 0.05). Moreover, on the 6th and 8th days, the β-Glu activity of the MT-treated group was pronouncedly attenuated vs the control (p < 0.01).
In the course of storage, the Cx activity of the two groups gradually rose to a peak on day 4, and then began to decrease (Fig. 2f). After a shelf life of 4 d, blueberry fruits in the treated group still exhibited lower activity than the untreated fruits, and the difference between the two groups was highly significant by the conclusion of the shelf life (p < 0.01).
To elucidate the molecular mechanisms underlying MT-regulated fruit softening, cell wall metabolism-related genes (VcPG1, VcPG3, Vcβ-Gal, Vcβ-Glu, VcCx1, VcPE, and VcPL) were investigated. The findings illustrated in Fig. 3 clearly demonstrate that MT treatment significantly downregulated the transcript levels of these genes, thereby effectively retarding the softening process.
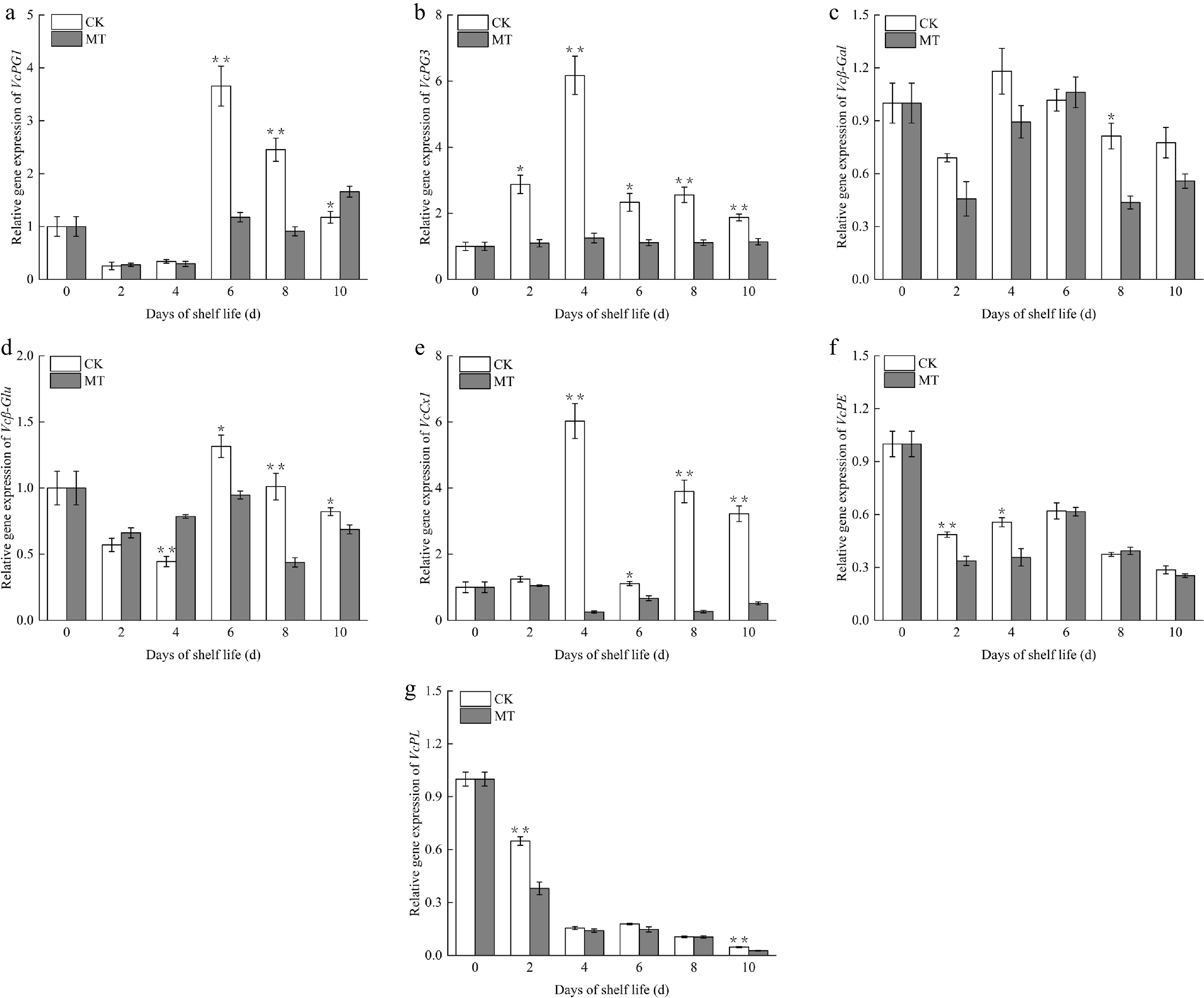
Figure 3.
(a)–(g) Expression of cell wall metabolism-associated genes in blueberry fruit during ambient storage. Significant differences among treatments are marked with * (p < 0.05) and ** (p < 0.01).
Effects of exogenous MT on the endogenous ABA content of blueberries
-
It is evident from Fig. 4a that the ABA content in the control fruits sharply increased from day 0 to day 4 of shelf life, slightly decreased from 4–6 d, and increased again at the ultimate stage of shelf life. In contrast, the ABA content in the MT-treated group decreased during the first 2 d and then continued to increase thereafter. The ABA content in the treatment group was appreciably lower than that in the control (p < 0.05), and reached a highly significant difference toward the end of the storage period (p < 0.01).
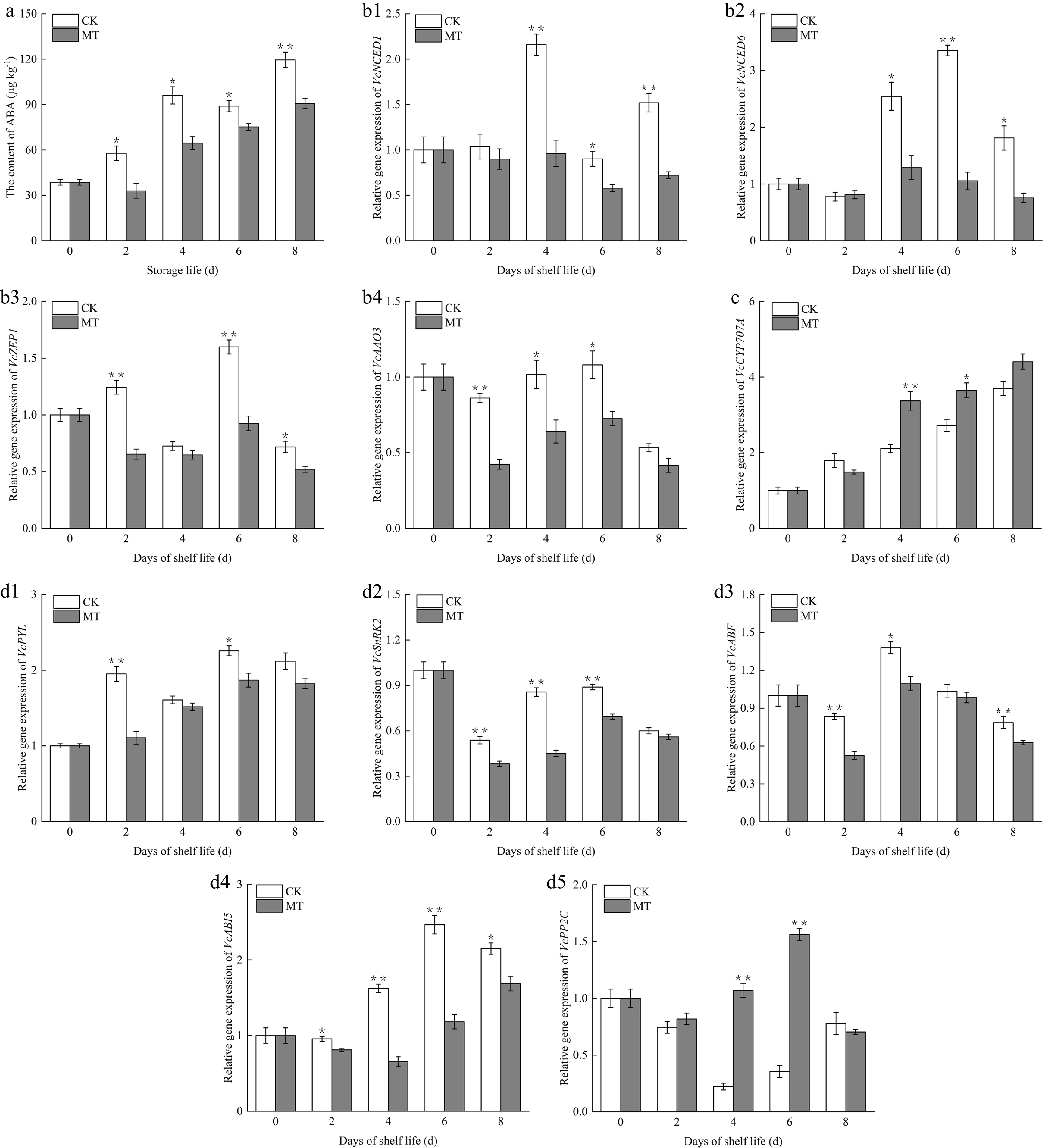
Figure 4.
Effects of exogenous MT on (a) endogenous ABA content, (b1)–(b4) ABA biosynthesis-related genes, (c) catabolism-related genes, and (d1)–(d5) ABA signal transduction genes of blueberry fruit during room temperature storage. Significant differences among treatments are marked with * (p < 0.05) and ** (p < 0.01).
Correlation analysis between endogenous ABA content, firmness, and cell wall metabolism-related enzymes
-
During room temperature storage, blueberry fruit firmness decreased continuously, while ABA content continued to increase. In order to explore the relationship between the two during the softening process of blueberries, a correlation analysis was conducted on ABA content, firmness, the activities of cell wall metabolic enzymes that are central to the firmness decrease process (Fig. 5). Analysis of correlations revealed a strong inverse correlation between ABA content and fruit firmness (p < 0.01), whereas a distinct parallel association between ABA content and the activities of PG, β-Gal, and β-Glu (p < 0.01), as well as a notable concomitant linkage between ABA content and Cx activity, were recorded (p < 0.05). The above data imply that the accumulation of ABA significantly affects blueberry firmness and may exert its regulatory effect by modulating the activities of cell wall-degrading enzymes.
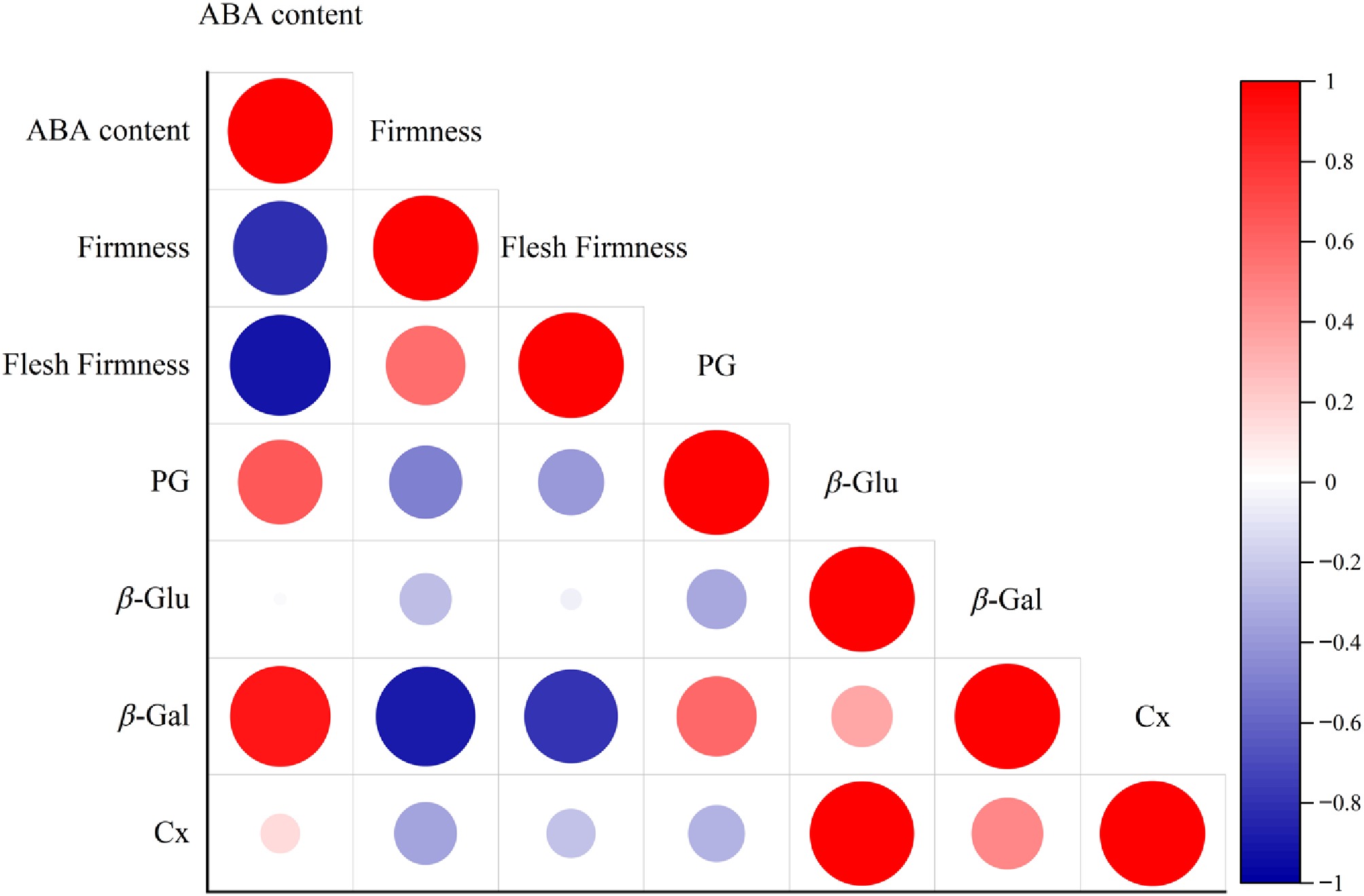
Figure 5.
Correlation analysis between ABA, firmness, and cell wall metabolic enzymes in blueberries.
Effects of exogenous MT on gene expression related to ABA biosynthesis, degradation, and signal transduction in blueberries
-
The relative level of the VcNCED1 gene in the control gradually increased from day 2 to 4 of shelf life, peaking on day 4. Afterward, it decreased between days 6 and 8, followed by a subsequent increase (Fig. 4b1). For the MT-treated group, the relative transcription level of VcNCED1 did not change significantly throughout the shelf life, only decreasing slightly on day 6. Moreover, its expression remained below the control level over the whole shelf life, with a significant difference detected on days 4, 6, and 8 (p < 0.05).
Regarding VcNCED6, its transcript level in the two blueberry groups declined marginally over the initial 2 d of shelf life, and surged rapidly on day 4 (Fig. 4b2). The expression level in the MT-treated group rose to the highest expression level at this time and continued to decrease thereafter. In contrast, VcNCED6 expression in the untreated fruit kept rising, peaking on day 6 at 3.18 times the initial level, with a highly significant difference from the MT-treated group (p < 0.01). Except on day 2, the relative transcript level of VcNCED6 exhibited significantly lower values than those in the control (p < 0.05).
In untreated fruit, VcZEP1 expression first increased, then decreased, peaking on day 6 of shelf life (Fig. 4b3). The expression level in MT-treated fruits decreased at the start of storage, slightly increased from day 6 to 8, and then decreased again. Across the whole storage duration, except on day 4, the relative expression level of VcZEP1 was markedly lower than that in the control (p < 0.05).
VcAAO3 expression rapidly decreased in the first 2 d of storage, then increased, followed by a further decline on day 8 (Fig. 4b4). Differing significantly from the untreated fruit, the expression level in the MT-treated group was strikingly downregulated (p < 0.05). Specifically, on day 2, the VcAAO3 gene expression level was 49% of that in the control (p < 0.01).
The expression of VcCYP707A gradually increased in blueberry fruits throughout the shelf life (Fig. 4c). Except on day 2, VcCYP707A expression in MT-treated fruits was greater than that in the control between days 4 and 6, with a marked difference (p < 0.05). Specifically, on day 4, the expression level of VcCYP707A in the untreated fruit was 1.6 times lower than that in the MT-treated group (p < 0.01).
The VcPYL expression level in the untreated fruits rose rapidly in the initial two days of storage, followed by a downward trend; it slightly rebounded on day 6 and dropped again on day 8 of shelf life (Fig. 4d1). The expression of VcPYL in the MT-treated group gradually rose in the first 6 d and slightly declined on day 8. Notably, the expression level of VcPYL was lower than that in the control throughout the shelf life, with significant differences observed between the two groups on days 2 and 6 (p < 0.05).
The expression levels of VcSnRK2 in both fruit groups exhibited a similar trend during shelf life, initially decreasing, then increasing, and finally declining again (Fig. 4d2). The disparity between the two groups was most pronounced on day 4 of storage. Apart from the 8th d, these contrasts were highly significant (p < 0.01) at all other time points.
As depicted in Fig. 4d3, the expression of VcABF in blueberry fruits declined over the first couple of days, which was significantly lower than that in the control (p < 0.01). Subsequently, VcABF expression rose rapidly from day 4 to 8 before gradually decreasing. Across the whole duration, the expression level in the control remained steadily higher than that in the MT-treated group, with significant differences observed at all time points except day 6 (p < 0.05).
VcABI5 expression in the untreated fruit dropped first, then rose over the storage period, reaching its peak on day 6 and then decreasing again (Fig. 4d4). The fruit treated with MT also followed a similar trend of first declining, then rising. VcABI5 expression in the treatment group was considerably lower than that in the control (p < 0.05), and the difference was highly significant on day 4 and day 6 (p < 0.01).
PP2C acts as a negative regulator within the ABA signaling pathway. The expression level of the VcPP2C gene in blueberry fruits declined initially, then increased during the first 6 d (Fig. 4d5). Specifically, the transcriptional levels of VcPP2C in the control were significantly lower than those in the MT-treated group on the 4th and 6th days (p < 0.01).
Effects of exogenous MT on endogenous IAA content of blueberries
-
IAA content in the untreated fruit dropped significantly during the initial 2 d, followed by a slight upward trend on day 4 and a subsequent decline (Fig. 6a). The IAA content declined gradually throughout storage in the treatment group, with a minor increase observed on the 8th day. Notably, MT-treated fruits sustained consistently higher IAA levels than the control, with significant differences detected except on the 4th d (p < 0.05). Postharvest MT application retarded the depletion of IAA content, thereby preserving the endogenous IAA level in blueberry fruits.
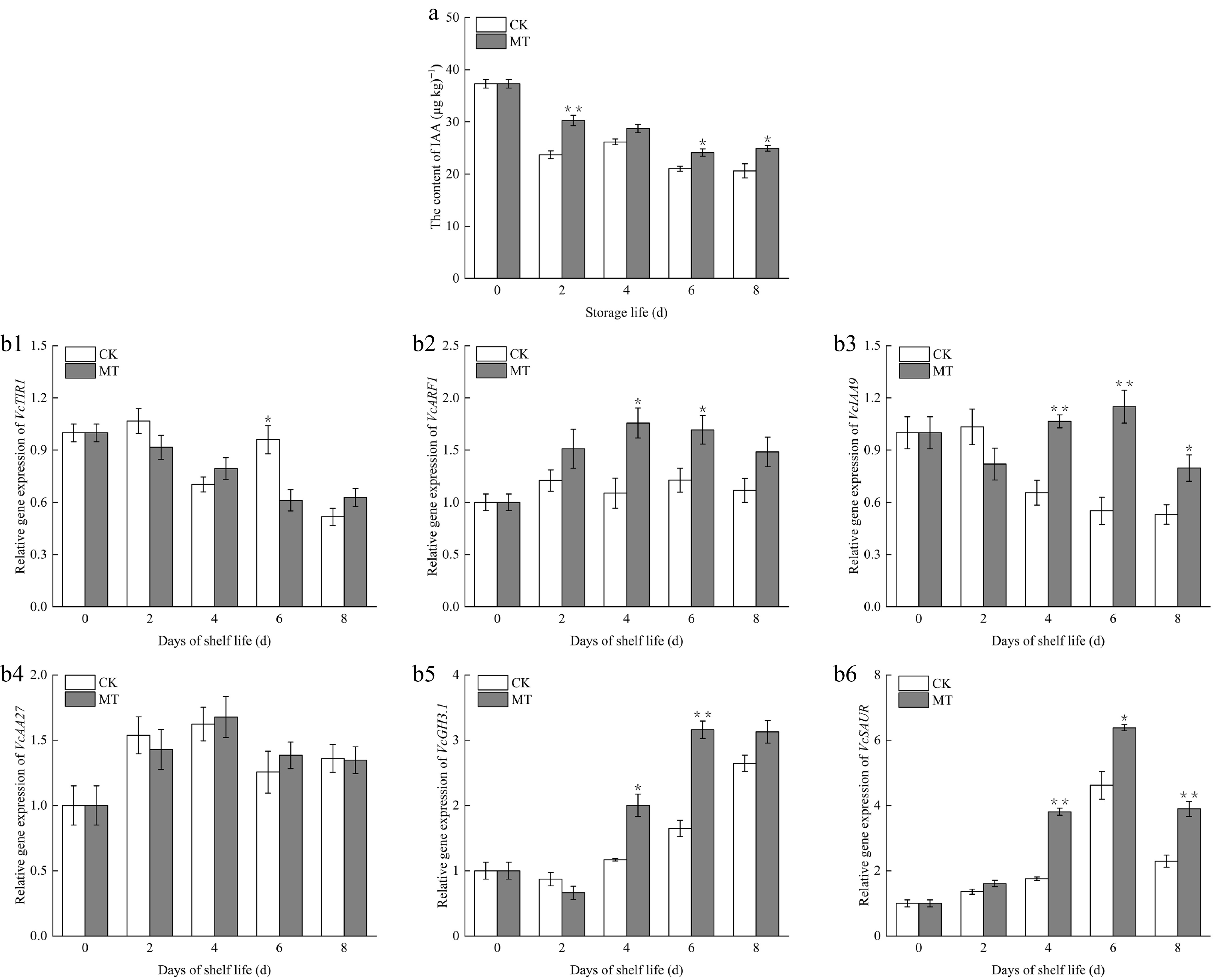
Figure 6.
Effects of exogenous MT on (a) endogenous IAA content, and (b1)–(b6) IAA signal transduction genes in blueberry fruit during room temperature storage. Significant differences among treatments are marked with * (p < 0.05) and ** (p < 0.01).
Effects of exogenous MT on IAA signal transduction-related gene expression in blueberries
-
VcTIR1 expression in the untreated fruit first rose and then fell during the first 4 d of postharvest storage, then rose again on day 6 (Fig. 6b1). Over the shelf life, the expression level of VcTIR1 in the MT-treatment group gradually declined, but was slightly elevated on day 8. The gene expression in the treatment group on the 4th and 8th days was higher than that in the control group.
The overall change in the VcARF1 gene in the untreated fruit was not significant. After MT treatment, the expression level in fruit gradually increased and then continued to decrease (Fig. 6b2). Throughout the shelf life, the MT treatment group stayed higher than that in the control, with the differences on days 4 and 6 reaching a significant level (p < 0.05).
VcIAA9 expression in the control rose slightly in the first 2 d of storage, then declined continuously (Fig. 6b3). In the MT treatment group, VcIAA9 expression declined slightly at the onset of shelf life, but rose gradually from day 4 to 6, reaching levels statistically greater than those in the control (p < 0.01). The gene expression level of the MT-treated group decreased again, but still showed significant differences versus the control (p < 0.05).
During the shelf life, VcIAA27 expression in both groups of fruits first peaked on day 4, followed by a gradual decline (Fig. 6b4). No significant difference occurred in gene expression levels throughout the shelf life, indicating that VcIAA27 expression was not apparently affected by MT treatment.
Importantly, the expression of early auxin response genes VcGH3.1 and VcSAUR was analyzed. The overall expression level of VcGH3.1 in the two groups of fruits during shelf life displayed a progressive upward trend (Fig. 6b5). Against the control, MT treatment significantly enhanced VcGH3.1 expression, with the exception of day 8 (p < 0.05). For the VcSAUR gene (Fig. 6b6), both groups manifested a trend of a gradual rise followed by a modest fall at the late stage of shelf life. The highest expression levels were observed on day 6, with the treatment group attaining a 1.38-fold increase versus the control. Notably, significant differences in VcSAUR expression were identified between the two groups from day 4 to 8 (p < 0.05).
-
Postharvest softening of blueberries is a common physiological phenomenon and a core challenge that limits the shelf life, commercial value, and storage safety of the fruit. MT, an endogenous signaling molecule ubiquitously distributed across the plant kingdom, has arisen as a prospective candidate for postharvest preservation owing to its exceptional safety profile, superior biocompatibility, and potent regulatory effects[29]. Research has confirmed that MT treatment is capable of preserving the firmness of harvested berry fruits such as strawberries and raspberries, delaying fruit softening[30]. Liu et al.[31] identified that MT is involved in the postharvest wax metabolism of blueberries, slowing down the deterioration of cuticular wax, inhibiting cell wall architecture disassembly, thereby maintaining the postharvest firmness of blueberries.
The treatment effect was best at 100 μM among the three concentration; thus, 100 μM was adopted for subsequent trials (Fig. 1). Pectin dissolution is a major feature of fruit softening, during which protopectin is converted to soluble pectin and further hydrolyzed. The dissolution and depolymerization of cell wall polysaccharides modify their composition and viscosity, causing the softening of fruit[32]. A 100 μM MT treatment can effectively suppress softening of blueberry fruits and preserve fruit quality. Pectin, cellulose, and hemicellulose are the core structural components of fruit cell walls. During storage, these components progressively degrade via the action of cell wall-modifying enzymes, compromising cell wall integrity. Concomitantly, the solubilization of bound pectin into soluble forms further contributes to the loss of fruit firmness[33]. Experiments have found that as the shelf life is prolonged, the protopectin of blueberry fruits gradually degrades, soluble pectin increases, bringing about fruit softening, which is similar to the changes in pectin during strawberry softening[34]. MT treatment sustained an elevated content of insoluble pectin over storage, whilst soluble pectin levels were lower than the control group, apart from day 8 (Fig. 2). MT treatment considerably retarded the solubilization of pectin by preserving insoluble pectin levels and limiting the formation of soluble pectin, which collectively orchestrated the maintenance of fruit firmness. This observation corroborates earlier research on jujube[10].
The integrity of the cell wall structure is essential for sustaining cellular function, and fruit softening after harvest is often attributed to an array of cell wall-degrading enzymes. The activities of critical cell wall-degrading enzymes were significantly diminished by MT treatment, including PG, Cx, β-Glu, and β-Gal. This result accords with findings reported in tomatoes[35] and blueberries[26]. ABA promotes pectin breakdown and contributes to fruit softening in postharvest European pears by upregulating the activities of PG and PL enzymes, as well as the expression of genes linked to them[36]. Further analysis revealed that MT treatment also brought about a significant decrease in the expression of cell wall metabolism genes, in agreement with previous research by Liu et al.[31]. This study did not measure the endogenous MT content, but previous studies have shown that exogenous MT can induce the accumulation of endogenous MT in fruits such as broccoli[37] and bananas[38]. Thus, exogenous MT may also upregulate the endogenous MT level in blueberries, thereby synergistically regulating softening with ABA. Softening of harvested fruits triggers changes in cell wall structure, regulated by various plant hormones, including ABA. As Bluecrop blueberries are non-climacteric[14,39], ethylene was not considered a key regulator, while ABA has been proven to be closely associated with fruit softening. In addition, MT may partially delay fruit softening by regulating ethylene metabolism, which provides a new entry point for subsequent research. Existing research has demonstrated that MT inhibits postharvest senescence and softening of fruits and vegetables by suppressing ABA accumulation[21,40], which corresponds to the results in Fig. 4. Exogenous MT treatment was observed to delay the continuous accumulation of ABA content during sample storage. Yin et al.[41] confirmed that PMTR1, as a melatonin receptor, affects ABA metabolism by regulating NCEDs and CYP707As, thereby regulating Arabidopsis seed germination. Based on this, we speculate that MT in blueberries may participate in postharvest softening regulation through a receptor-mediated ABA signaling pathway.
The correlation analysis in Fig. 5 reflects the correlation between variables, but combined with previous studies, we can further infer the causal relationship whereby ABA and MT mediate the delayed softening of blueberries. Inhibiting ABA synthesis in grape fruits significantly weakens the promoting effect of MT on anthocyanin accumulation, indicating that ABA plays a mediating role in MT-induced fruit ripening[42]. It is speculated that MT inhibits ABA accumulation, so that cell wall degradation is lessened and blueberry softening is retarded. Endogenous ABA content in fruit is closely linked to its biosynthesis, degradation, and signal transduction. Specifically, the ABA biosynthetic pathway in higher plants involves three major enzyme families: ZEP, NCE, and AAO, among which NCED is considered the predominant rate-limiting enzyme[43]. Our data revealed that MT slowed the onset of fruit softening by curbing endogenous ABA accumulation in blueberries. MT treatment suppressed the increase in endogenous ABA content in postharvest fruits, downregulated the expression of ABA biosynthesis genes VcNCED1, VcNCED6, VcZEP1, and VcAAO3, and reduced the expression of ABA-degrading gene VcCYP707A. Dai et al.[44] demonstrated that anthocyanin treatment suppresses ABA biosynthesis by downregulating VcNCED1 expression, thereby retarding senescence and softening in blueberries. Core elements of the ABA signal transduction pathway comprise the PYR/PYL receptors, PP2C phosphatases, SnRK2 kinases, ABF transcription factors[45−48]. MT inhibits the postharvest senescence process of leaves by downregulating the expression of ABFs in cabbage hearts, thereby blocking their ABA biosynthesis[21]. The same results were obtained in this experiment, where MT treatment substantially dampened the expression levels of VcABF, VcABI5, VcPYL, and VcSnRK2 genes in the ABA signal transduction pathway. PP2Cs act as core negative regulators that repress ABA signaling. Studies have shown that MT treatment markedly decreases ABA content and increases PP2C expression, indicating that ABA signal transduction mediates MT’s inhibition of postharvest senescence in litchi[49]. MT treatment boosted the expression level of the VcPP2C gene during the postharvest shelf life in blueberry fruits, indicating that MT inhibits ABA signal transduction by activating VcPP2C transcription. Li et al.[50] showed that tomato SlPP2C2 interacts with IAA synthase FZY and IAA signaling protein SAUR, and that IAA treatment induces SlPP2C2 expression. Modulating SlPP2C2 levels bidirectionally alters endogenous IAA content. We therefore speculate that MT-activated IAA signaling may suppress ABA accumulation by regulating negative regulators such as VcPP2C. The ABI5-like transcription factor in bananas can directly bind to the promoter of cell wall degradation genes and activate transcription. The interaction between EBF1 protein and ABI5-like further enhances gene expression, thereby promoting the ripening and softening of banana fruits[51]. Xiang et al.[52] confirmed in Arabidopsis seeds that ABI5 can directly regulate the transcription of and inhibit the cell wall modification gene PME31, and participate in ABA-mediated seed germination by regulating pectin metabolism, indicating that ABI5 can directly target and regulate cell wall-related genes. This study confirmed the close association between VcABF/VcABI5 and cell wall degradation-related genes from the perspectives of physiological phenotypes, gene expression, and cell wall degradation enzyme activity, thereby proposing this regulatory mechanism and constructing a molecular interaction model diagram.
Involved in numerous physiological processes in fruits, IAA is an important plant hormone. The relationship network between plant hormones in fruits is vast and complex, and studies have shown that IAA may regulate endogenous ABA content through signal transduction pathways[19]. MT maintains cherry tomato quality by regulating ABA synthesis and degradation genes, reducing ABA content, upregulating SlYUCCA, and increasing IAA levels[22]. During banana fruit ripening, ABA promotes fruit softening, while IAA inhibits fruit softening[23]. In grapes, MT regulates ABA and IAA through the H2O2 signaling pathway[24]. Based on this, this study explores whether MT can regulate ABA accumulation in blueberries by affecting IAA content. The study found that the IAA content in blueberries decreased continuously after harvesting, while exogenous MT treatment inhibited the decrease in IAA content and maintained IAA levels (Fig. 6). The changes in IAA levels are tightly linked to downstream transduction of IAA signaling. Therefore, we determined the IAA signaling-related genes (VcTIR1, VcARF1, VcIAA9, VcGH3.1, and VcSAUR). In strawberries, AUXIN RESPONSE FACTOR 2 (FaARF2) can directly bind to the promoter of the key ABA synthesis gene FaNCED1 and inhibit its transcriptional activity, thereby reducing ABA levels and inhibiting receptacle maturation. It is speculated that the IAA signal activated by MT in blueberries may inhibit ABA accumulation through a similar mechanism, which is worth further exploration[53]. Our results confirm that blueberry fruits treated with 100 µM MT exhibit elevated IAA content as storage proceeds. The expression of IAA signaling genes was upregulated by MT treatment, indicating that MT enhances IAA signaling in blueberry fruits by activating these pathways, sharing an analogy with the observations of Li et al.[54].
The critical role of MT treatment on ABA and IAA signaling in fruit ripening suggests that there may be an interaction between these pathways. Figure 7 depicts the effects of exogenous MT treatment on ABA biosynthesis, degradation, signal transduction, and IAA signal transduction levels. The results indicate that MT treatment can suppress ABA accumulation and delay blueberry softening, and MT may inhibit the increase in ABA content by regulating IAA levels. Nonetheless, the complex mechanism of the interaction between IAA and ABA signal transduction during blueberry ripening needs further investigation.
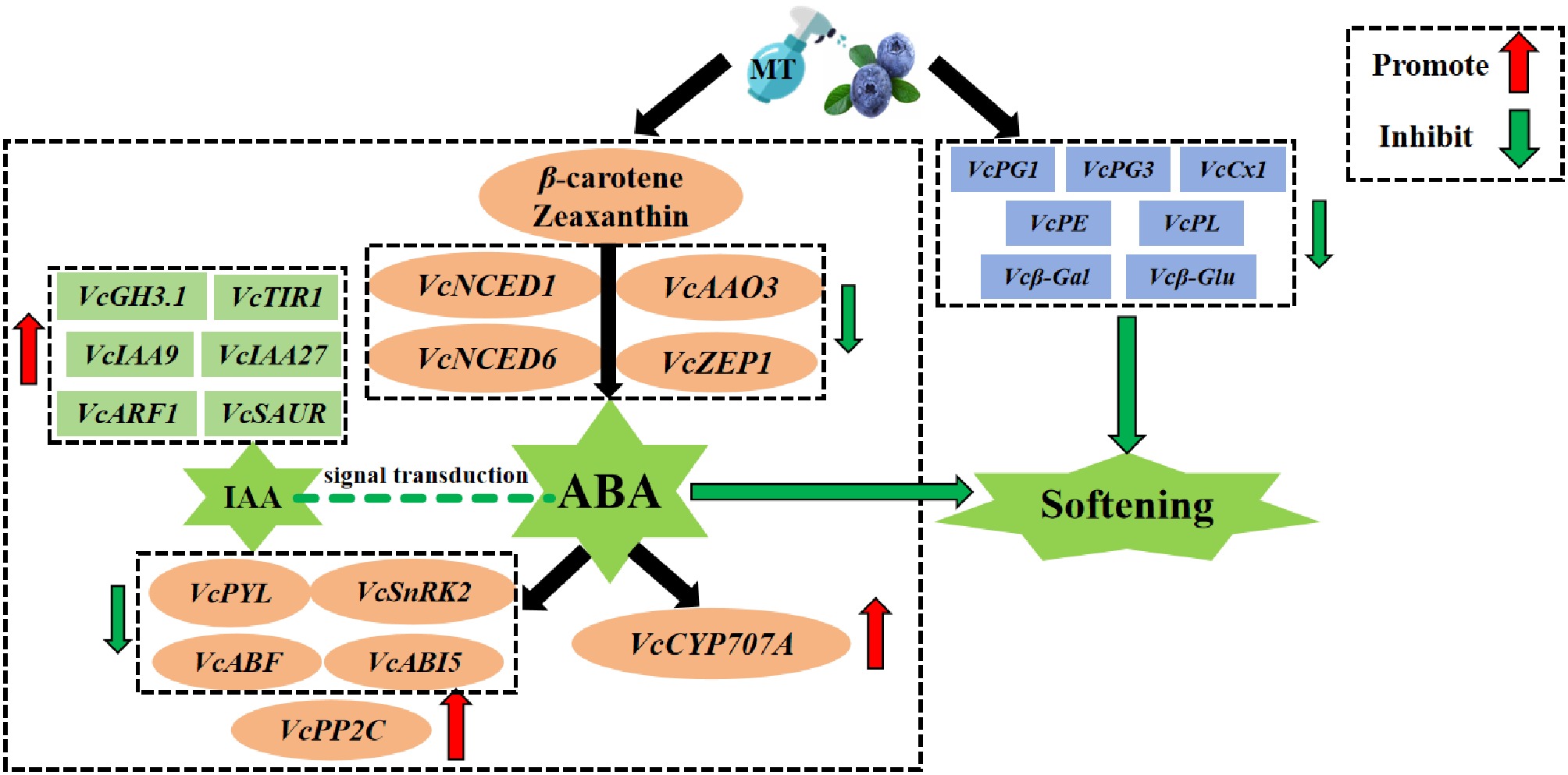
Figure 7.
Interaction between exogenous MT and abscisic acid.
-
In summary, research has found that treatment with 100 μM MT can maintain the original pectin content of blueberry fruit, inhibit the increase of soluble pectin, and regulate the expression of genes related to cell wall metabolism; Simultaneously regulate ABA metabolism and signal transduction related genes (upregulate VcCYP707A, VcPP2C, inhibit synthesis genes such as VcNCED1, and signal genes such as VcABF), slow down the decrease in endogenous IAA content, and upregulate signaling pathway related genes. Postharvest MT treatment inhibits the increase of ABA content by regulating ABA synthesis, degradation, and signal transduction, while maintaining its level by regulating IAA signal transduction and inhibiting endogenous ABA accumulation, thereby delaying postharvest softening of blueberries. This study explored the mechanism of blueberry shelf life softening at room temperature from the perspective of inhibiting ABA accumulation, providing a theoretical basis for exogenous MT to inhibit blueberry softening.
-
The authors confirm their contributions to the paper as follows: study conception and design: Lin MF, Quintana R, Sun L; methodology: Wang LL, Quintana R, Song ZC; software, investigation: Song ZC; formal analysis: Sun L, Zhang J; validation: Wang LL, Quintana R, Zhu YX; data curation: Lin MF; visualization: Zhu YX; resources: Wei BD, Zhou Q; draft manuscript preparation: Wang LL; writing − review and editing: Wang LL, Lin MF; supervision: Zhang J, Wei BD; project administration, funding acquisition: Zhou Q. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated and analyzed in this study are presented in the published article and its supplementary material files.
-
This study was funded by the Liao Ning Revitalization Talents Program (Grant No. XLYC2403139), and the China-Chile Belt and Road Joint Laboratory on Blueberry (Grant No. 2024021198-JH2/1021).
-
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Chongqing University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang LL, Lin MF, Quintana R, Song ZC, Zhu YX, et al. 2026. Exogenous melatonin delays ABA-mediated postharvest softening of blueberry fruits by regulating IAA homeostasis. Plant Hormones 2: e014 doi: 10.48130/ph-0026-0012 

Exogenous melatonin delays ABA-mediated postharvest softening of blueberry fruits by regulating IAA homeostasis
- Received: 17 March 2026
- Revised: 09 May 2026
- Accepted: 11 May 2026
- Published online: 08 June 2026
Abstract: Fruit softening is a key factor affecting the postharvest quality of blueberries. Prior research by our team has demonstrated that 100 μM melatonin (MT) treatment effectively delays softening and preserves quality. The core of this article lies in the consequences of exogenous MT treatment on abscisic acid (ABA) biosynthesis, degradation, and signal transduction, as well as auxin (IAA) signaling, in relation to postharvest blueberry softening. The results demonstrated that MT treatment delayed pectin degradation and significantly reduced cell wall degradation-related gene expression, downregulated ABA biosynthetic genes (VcNCED1, VcNCED6, VcZEP1, and VcAAO3) while upregulating the expression of the ABA catabolism gene VcCYP707A. MT treatment also downregulated ABA signal transduction genes (VcABF, VcABI5, VcPYL, and VcSnRK2) while upregulating the negative regulator VcPP2C, collectively suppressing ABA accumulation. Furthermore, MT treatment delayed the reduction of endogenous IAA levels, upregulated the expression levels of IAA signal transduction genes, including VcTIR1, VcARF1, VcIAA9, VcGH3.1, and VcSAUR. Overall, postharvest MT treatment inhibited ABA accumulation by modulating its biosynthesis, degradation, and signal transduction while maintaining IAA levels through regulating IAA signal transduction. This dual regulatory mechanism ultimately delays postharvest blueberry softening.