










-
Seed germination represents the first and most critical stage of the plant life cycle, playing a decisive role in crop yield[1,2]. However, soybean (Glycine max L.) seeds are short-lived and highly susceptible to deterioration because of their high protein and oil content, leading to significantly reduced seed vigor and germination rates[3]. Previous studies have found a strong positive correlation between soybean's germination rate and yield[4]. In practical production, high-germination soybean seeds could enhance the uniformity of field emergence, reduce the amount of seed sown, lower agricultural planting costs, and ultimately increase yields[5]. Therefore, identifying the key genes regulating soybean's germination and elucidating their molecular mechanisms is vital for advancing high-yield soybean breeding programs.
The transition of crop seeds from dormancy to germination is determined by the balance between the levels of abscisic acid (ABA) and gibberellin (GA) within the seeds. A lower ABA/GA ratio is required for seed germination[6]. These hormones act antagonistically, mutually and negatively regulating each other's biosynthetic pathways[7]. Reducing the expression levels of ABA biosynthesis genes (such as 9-cis-epoxycarotenoid dioxygenases [NCEDs]) and GA metabolic genes (such as GA2OXs, gibberellin 2-oxidases), while increasing the expression levels of the ABA metabolic gene CYP707As (cytochrome P450 707As) and GA biosynthetic genes (such as GA20OXs, gibberellin 20-oxidases), can lower the ABA/GA ratio, thereby enhancing the rate of seed germination[8,9]. ABI5 (abscisic acid-insensitive 5) is one of the master regulators in ABA signaling that negatively regulates seed germination, which regulates ABA and GA metabolic and signaling genes to control ABA and GA levels, thereby repressing seed germination[10]. The DOG1 (Delay of Germination 1) protein is another major regulator of seed dormancy, which interacts with ABA signaling and delays seed germination[11,12].
Jasmonate-Zinc-finger expressed in inflorescence meristem (ZIM) domain (JAZ) proteins, as components of jasmonic acid (JA) signaling, participate in various growth and developmental processes of plants[13−15]. Studies have demonstrated that JAZ proteins interact with ABI3/ABI5 and suppress their transcriptional activity, thereby affecting seed germination in wheat (Triticum aestivum), rice (Oryza sativa), and Arabidopsis thaliana[16−19]. The soybean genome encodes 28 JAZ proteins (Wm82.a4.v1), with some JAZ repressors involved in plant development and abiotic stress responses. For instance, JAZ1/2 interacts with Avh94 to negatively regulate soybean's resistance to Phytophthora infection through the jasmonate signaling pathway[20]. GmJAZ3 directly interacts with the jasmonate signaling transcription factor GmMYC2a, suppressing its transcriptional activation of GmCKX3-4. Simultaneously, GmJAZ3 interacts with both GmRR18a and GmMYC2a to inhibit their activation of the cytokinin oxidase gene GmCKX for controlling seed size and weight[21]. GmJAZ8 may mediate soybean's resistance to soybean cyst nematodes through jasmonate signaling processes[22]. However, the role of JAZs in soybean seed germination remains to be elucidated.
This study provides important insights into the role of GmJAZ8 in soybean seed germination.
-
A set of 277 soybean accessions (with 164 improved varieties and 113 landraces)[23] used for a genome-wide association study (GWAS) was planted at the Yangdu experimental station of Zhejiang Academy of Agricultural Sciences during autumn in 2023. Detailed information regarding these soybean accessions is provided in Supplementary Table S1. Field planting was performed according to a randomized complete block design, which used a plot with a single row and three replicates. Seeds were collected at the fully ripe stage. Arabidopsis (Col0 and overexpression-GmJAZ8) was grown under uniform conditions in a climate-controlled growth chamber with a 16-h light/8-h dark cycle at 22 °C.
Seed germination
-
Before germination, chlorine gas (5 mL HCl + 95 mL sodium hypochlorite) was used to sterilize the seeds and reduce the risk of microbial contamination. Twenty healthy soybean seeds were sown on a 0.9% agar medium, and 6 mL of sterile water was then added. The soybean seeds were incubated at 25 °C under dark conditions for 48 h after imbibition (HAI). Arabidopsis seeds were incubated for 2 d at 22 °C under a 12-h light/12-h dark cycle after 3 d of cold treatment. The germinated seeds (embryonic axis penetrating the seed coat) were counted to obtain the germination rate (GR) from 0 to 48 HAI. Three biological replications were performed.
Genome-wide association study
-
Single-nucleotide polymorphisms (SNPs) derived by resequencing 277 soybean accessions were used to conduct a GWAS for seed germination. In total, 89,125,094 high-quality SNPs with a missing rate of < 0.1 or a minor allele frequency of > 0.05 were used for the GWAS. The threshold for a significant association was set to 1/n (n is the number of SNPs, p < 1.122 × 10−8 or −log10(p) > 7.95). The general linear model (GLM) and the mixed linear model (MLM) were used to analyze the association with seed germination. Manhattan plots were generated by the R package rMVP.
RNA extraction, cDNA, and quantitative real-time polymerase chain reaction
-
Total RNA was extracted from dry seeds utilizing Vazyme's FastPure® Universal Plant Total RNA lsolation Kit (RC411-01) following the manufacturer's protocol, and three biological replicates were performed for each experiment. Reverse transcription was carried out using a cDNA synthesis kit. The cDNA sequence was subsequently used as the template for quantitative real-time polymerase chain reaction (qRT-PCR). qRT-PCR was performed using a LightCycler 480 with SYBR Green I Master Mix. Gene expression was normalized to the expression of AtTUBLIN. The relevant primers used are listed in Supplementary Table S2.
Phylogenetic analysis
-
Duplicates and orthologs with high similarity to GmJAZ8 from soybean and other representative species (rice and Arabidopsis) were obtained from Phytozome 13 (
https://phytozome-next.jgi.doe.gov/blast-search ). The aligned sequences were used to construct phylogenetic trees in MEGA5 software. The bootstrap value was set to 1,000 replications.Vector construction and transformation
-
To construct the GmJAZ8 (Glyma.08G096500) overexpression vector, the coding DNA sequences (CDSs) of GmJAZ8 were amplified from Willimas 82 (Wm82) and ligated into the pBinGFP4 vector. The overexpression vector was subsequently introduced into Agrobacterium tumefaciens strain EHA105 and then transformed into Arabidopsis. All primers used to construct the vectors are listed in Supplementary Table S2.
Overexpression of GmJAZ8 in Arabidopsis
-
To obtain the stable expression of GmJAZ8 in Arabidopsis, we transformed the recombinant vector into Agrobacterium tumefaciens(EHA105 strain) and then transformed Arabidopsis via the foral dip method[24]. The transformants of the primary plants were placed on a Murashige and Skoog (MS) medium using kanamycin (0.05 mg/mL) for testing resistance. After 10 d, the resistant seedlings were transplanted into the soil. We collected the resulting seeds and tested them for resistance to kanamycin, selecting from three different generations to produce transformed homozygous plants. All primers used to construct the vectors are listed in Supplementary Table S2.
Transcriptome sequencing and analysis
-
For the RNA-seq analysis, total RNA was extracted from dry seeds for six soybean accessions, GmJAZ8-overexpressing Arabidopsis lines, and wild-type Arabidopsis, and seeds germinated at 0 and 24 HAI of GmJAZ8-overexpressing and wild-type Arabidopsis using Vazyme's FastPure® Universal Plant Total RNA lsolation Kit (RC411-01), following the manufacturer's guidelines. Both the GmJAZ8-overexpressing lines and the wild-type were cultivated under identical conditions. Three biological replicates were performed for each sample. The RNA libraries were sequenced on the Illumina NovaseqTM 6000 platform. and a bioinformatic analysis was conducted by LC Bio Technology Co., Ltd. (Hangzhou, China).
The sequence quality was verified using FastQC (including the Q20, Q30, and GC content of the clean data), and mapped to the soybean reference genome (Wm82.a4.v1) using the HISAT2 package. The mapped reads of each sample were assembled using StringTie with the default parameters. All transcriptomes from all samples were merged to reconstruct a comprehensive transcriptome using gffcompare software. After the final transcriptome was generated, StringTie and ballgown were used to estimate the expression levels of all transcripts and the expression abundance for mRNAs by calculating the FPKM (fragment per kilobase of transcript per million mapped reads) value.
DESeq2 was used to calculate differentially expressed genes (DEGs), using the threshold of a false discovery rate of < 0.05 and an absolute fold change of ≥ 2. Principal component analysis (PCA) was performed with the princomp function of R. Gene Ontology (GO) enrichment analyses of the DEGs were performed using the cloud platform provided by Shanghai OEbiotech with the default parameters.
Data analysis
-
The experimental data were analyzed using GraphPad Prism software (version 10.1.2). The significant differences among samples were compared using Student's t-test.
-
To investigate the key genes affecting variation in the rate of seed germination in soybean, we analyzed the rate of seed germination at 36 and 48 HAI in 277 soybean accessions included in a previous resequencing population[23] (Supplementary Table S1). Subsequently, association analysis of the germination rate at 36 and 48 HAI was performed using the GLM and MLM. Finally, 59 significant SNPs were identified on chromosome 8 in 36HAI_GLM, whereas 36HAI_MLM identified 14 significant SNPs on chromosome 8. For 48HAI_GLM, 68 significant SNPs were identified on four chromosomes (1 SNP on chromosome 4, 62 SNPs on chromosome 8, 1 SNP on chromosome 11, and 4 SNPs on chromosome 20). Meanwhile, 60 significant SNPs were located on chromosome 8 (59 SNPs) and chromosome 20 (1 SNP) for 48HAI_MLM (Supplementary Table S3). Notably, across all four GWAS results, 14 significant SNPs on chromosome 8 were consistently identified (Fig. 1a–h and Table 1). These SNPs are physically close on the chromosome, forming a continuous cluster within a 113.5-kb region (7,394,400–7,507,912 bp). We focused further on the genes within this 113.5-kb interval block.
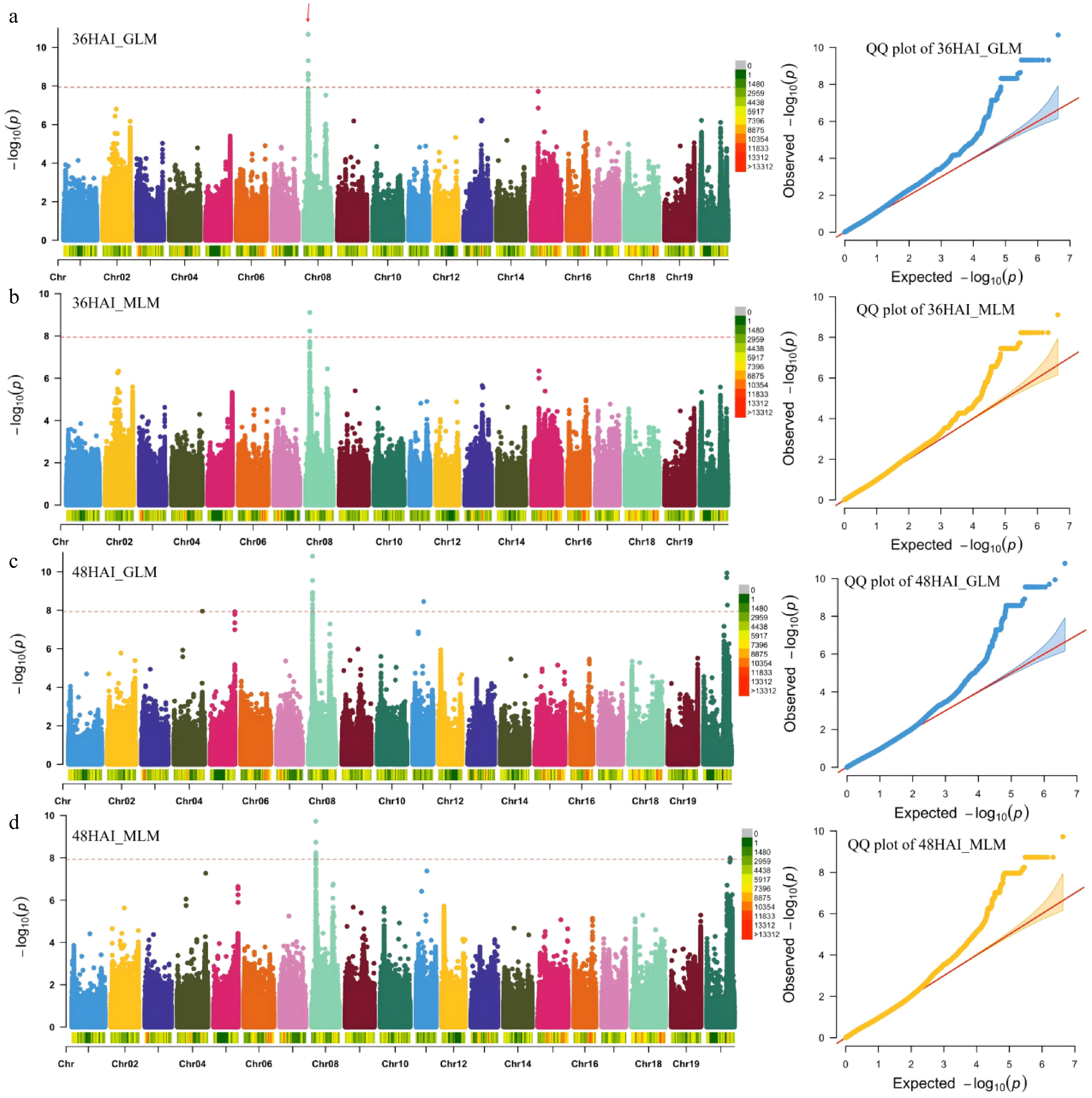
Figure 1.
GWAS of the rate of seed germination in soybean. (a)−(d) Manhattan plots and quantile–quantile (Q–Q) plots for the whole population of soybean accessions. The red arrows indicate the quantitative trait loci (QTLs) identified. GLM, general linear model; MLM, mixed linear model; HAI, hours after imbibition.
Table 1. Information of 14 significant SNPs within the 113.5-kb interval block (7,394,400–7,507,912 bp).
SNP Alleles 36HAI_GLM
p-value36HAI_MLM
p-value48HAI_GLM
p-value48HAI_MLM
p-valueChr08_7394400 C/A 2.12E-11 7.74E-10 2.72E-09 1.90E-10 Chr08_7400080 C/T 4.86E-10 5.82E-09 1.57E-11 1.86E-09 Chr08_7416347 C/T 4.86E-10 5.82E-09 2.82E-10 1.86E-09 Chr08_7427722 G/T 4.86E-10 5.82E-09 2.82E-10 1.86E-09 Chr08_7459314 T/C 4.86E-10 5.82E-09 2.82E-10 1.86E-09 Chr08_7464017 G/A 4.86E-10 5.82E-09 2.82E-10 1.86E-09 Chr08_7465749 G/A 4.86E-10 5.82E-09 2.82E-10 1.86E-09 Chr08_7470131 A/G 4.86E-10 5.82E-09 2.82E-10 1.86E-09 Chr08_7476997 C/A 4.86E-10 5.82E-09 2.82E-10 1.86E-09 Chr08_7490743 T/A 4.86E-10 5.82E-09 2.82E-10 1.86E-09 Chr08_7490778 T/C 4.86E-10 5.82E-09 2.82E-10 1.86E-09 Chr08_7500442 T/C 4.86E-10 5.82E-09 2.82E-10 1.86E-09 Chr08_7500556 T/G 4.86E-10 5.82E-09 2.82E-10 1.86E-09 Chr08_7507912 A/T 4.86E-10 5.82E-09 2.82E-10 1.86E-09 HAI, hours after imbibition; GLM, general linear model; MLM, mixed linear model. Identification of potential candidate genes for seed germination
-
The potential candidate genes within the 113.5-kb interval block were analyzed, and a total of 15 genes were annotated according to the reference genome Wm82 (Wm82.a2.v1) (Table 2). We obtained the expression levels of 15 genes in flowers and at different seed developmental stages from SoyOmics (
https://yanglab.hzau.edu.cn/SoyMD/#/tools/blast ), and found that Glyma.08G096500 was highly expressed in 10-week-old seeds. Further analysis of the expression patterns of these 15 genes during seed germination revealed that Glyma.08G096500, Glyma.08G096800, and Glyma.08G097700 were expressed throughout all stages of seed germination and were highly expressed in dry seeds (Supplementary Fig. S1a, S1b).Table 2. Fifteen potential candidate genes within the 113.5-kb interval block (7,394,400–7,507,912 bp).
Gene ID Position (bp) Functional annotation Homologous Arabidopsis genes Glyma.08G096500 7,396,887–7,399,006 Jasmonate–ZIM domain protein 8 AT1G30135 Glyma.08G096600 7,404,774–7,409,459 (S)-2-hydroxy-acid oxidase AT4G18360 Glyma.08G096700 7,412,769–7,417,800 (S)-2-hydroxy-acid oxidase AT4G18360 Glyma.08G096800 7,418,970–7,424,007 (S)-2-hydroxy-acid oxidase AT3G14420 Glyma.08G096900 7,424,892–7,426,785 Tetratricopeptide repeat (TPR)-like superfamily protein AT4G21065 Glyma.08G097000 7,429,593–7,429,941 AA_trans domain-containing protein AT5G19875 Glyma.08G097100 7,432,417–7,434,453 (S)-2-hydroxy-acid oxidase AT3G14420 Glyma.08G097200 7,435,704–7,438,009 (S)-2-hydroxy-acid oxidase AT4G18360 Glyma.08G097300 7,438,009–7,444,035 (S)-2-hydroxy-acid oxidase AT4G18360 Glyma.08G097400 7,447,072–7,448,398 Leucine-rich repeat receptor-like protein kinase AT2G42800 Glyma.08G097500 7,457,157–7,457,877 Cotton fiber expressed protein AT2G34610 Glyma.08G097600 7,460,358–7,463,786 BED zinc finger AT3G42170 Glyma.08G097700 7,468,455–7,476,125 RNA-binding KH domain-containing protein AT5G46190 Glyma.08G097800 7,478,544–7,486,117 Ornithine aminotransferase AT5G46180 Glyma.08G097900 7,489,719–7,494,791 TCP2 family transcription factor AT4G18390 To identify the key genes involved in the rate of seed germination, transcriptomic sequencing of dry seeds was conducted in three soybean accessions with low germination rates (GDC198 with 37.5%, GDL060 with 57.5%, and GDL062 with 57.5%) and three soybean accessions with high germination rates (GDC018 with 90%, GDL023 with 100%, and GDL075 with 100%) (Fig. 2a). Among these 15 genes, the transcriptomic data revealed that 13 genes were successfully detected but two genes (Glyma_08G097100 and Glyma_08G097200) remained undetected. In these 13 genes,the expression level of four genes was zero (Supplementary Table S4). Eight genes showed no significant differences between soybean accessions with low or high germination rates (Fig. 2b and Supplementary Table S4). Specifically, only Glyma.08G096500 demonstrated a statistically significant difference and exhibited significantly higher expression levels in high-germination accessions compared with low-germination accessions (Fig. 2b). Further validation using qRT-PCR to assess the expression levels of the 15 genes in the high-germination and low-germination materials revealed that also only Glyma.08G096500 showed a significant difference in its expression levels, which was consistent with the transcriptomic data (Supplementary Fig. S1c, S1d). According to the functional annotation of the soybean genome, Glyma.08G096500 encodes JAZ protein 8 (Table 2) and is considered to be a candidate gene named GmJAZ8. In a comparative analysis of the amino acid sequence similarity between GmJAZ8 and JAZ proteins from Arabidopsis thaliana and rice, GmJAZ8 contains a ZIM and a Jas domain and shares higher homology with AtJAZ7 and AtJAZ8 in Arabidopsis (Fig. 2c, d). In Arabidopsis, AtJAZ7 interacts with AtARF10 and AtARF16, integrating hormone signaling to suppress the rate of seed germination[18]. AtJAZ1/5/8 interact with AtABI3/5, integrating JA and ABA signaling during seed germination[25].
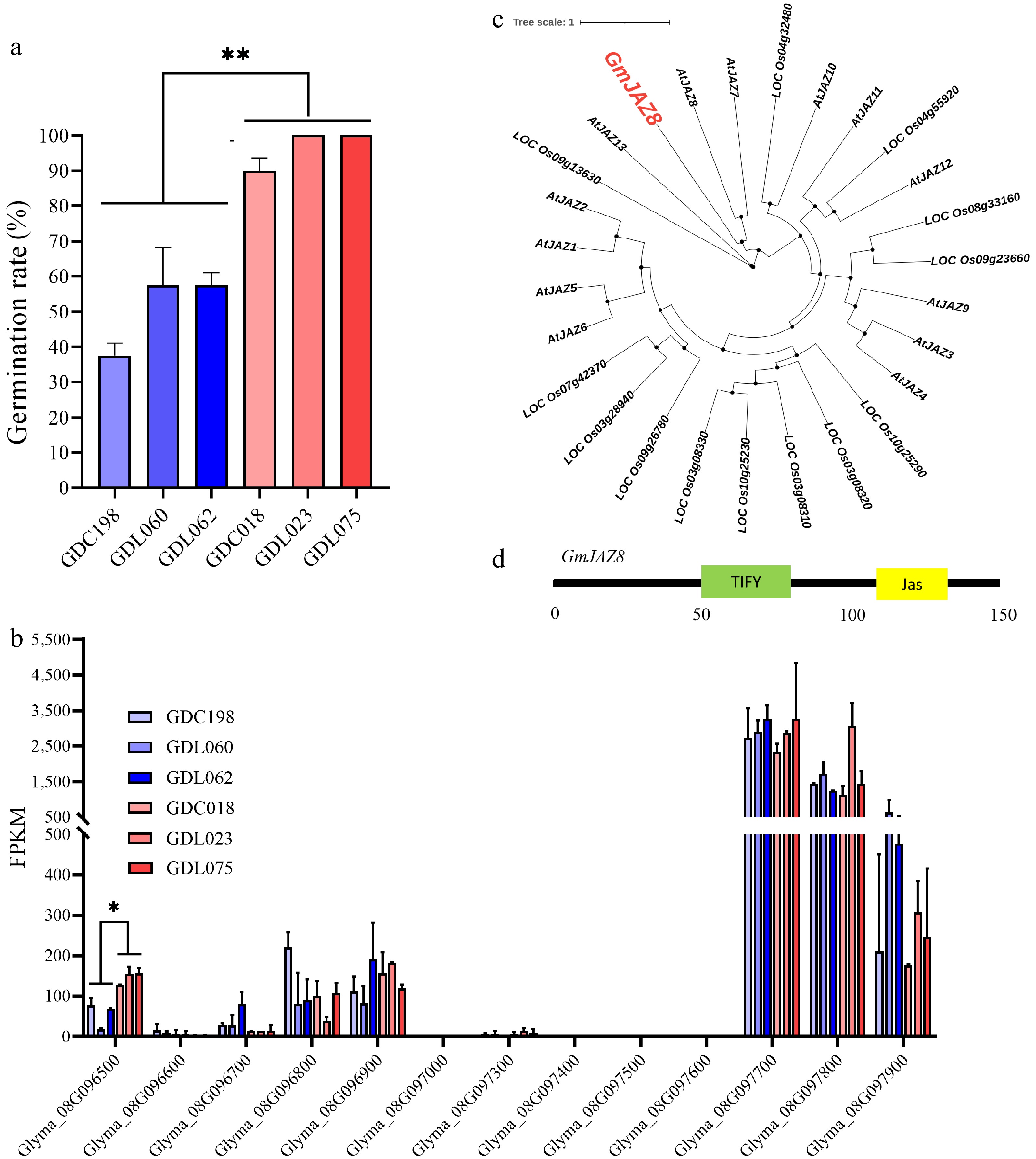
Figure 2.
Identification of GmJAZ8 for soybean the rate of seed germination. (a) Germination rate of six soybean accessions. **, p < 0.01. (b) FPKM of nine genes. *, p < 0.05. ns, no significance. Only genes with FPKM values greater than 0 are displayed. (c) Phylogenetic analysis of the GmJAZ8 and JAZ family from Arabidopsis and rice. (d) Domain organization of GmJAZ8. The numbers under the black line indicate the position of the amino acids.
Allelic variations within the promoter and intragenic regions of GmJAZ8 were detected in a resequenced population. Eight SNPs were identified within the promoter region (Supplementary Table S5). On the basis of these SNPs, the population was divided into three haplotype groups (hap_1 to hap_3) (Fig. 3a). The population primarily consisted of hap_1 (n = 215), hap_2 (n = 36), and hap_3 (n = 18). The rate of seed germination showed no significant differences between haplotype groups hap_1 and hap_2. However, the hap_3 haplotype group exhibited a significantly lower rate of seed germination than all other haplotype groups (Fig. 3b). According to these results, we propose that GmJAZ8 (Glyma.08G096500) is likely to be a key gene regulating seed germination in soybean.
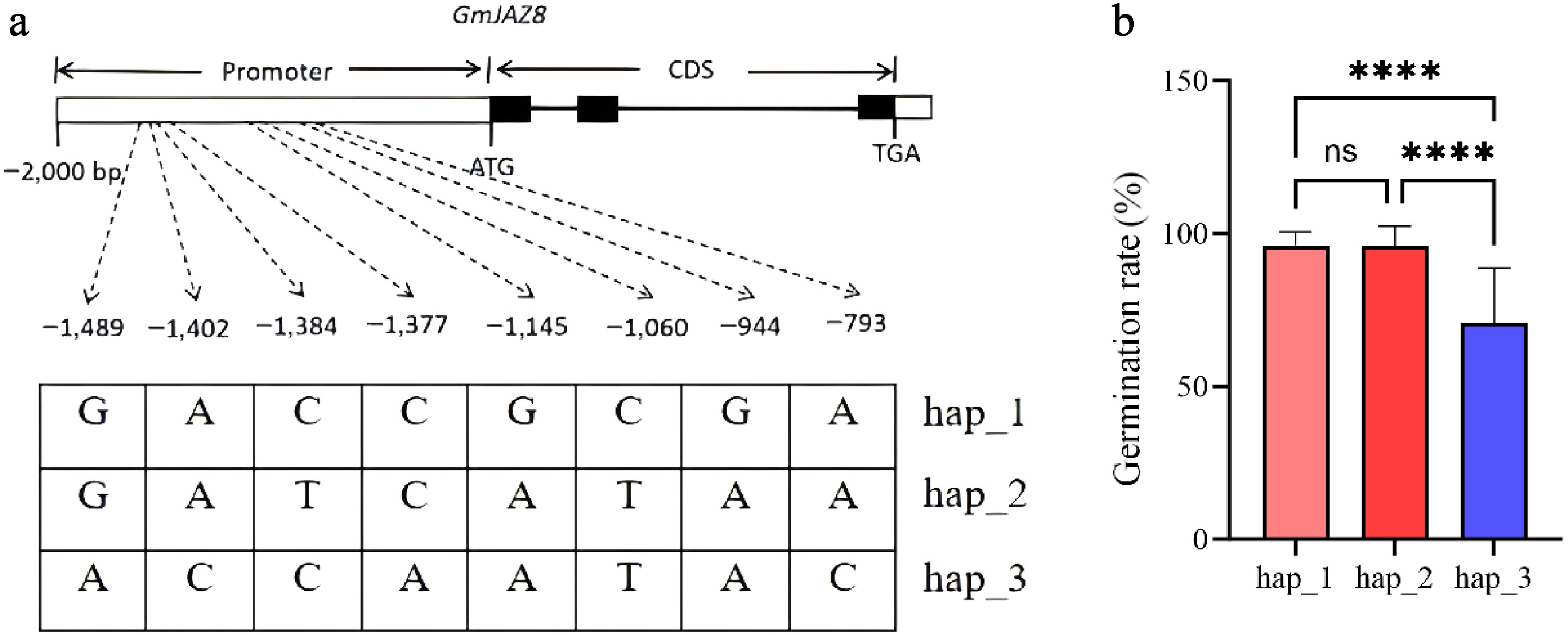
Figure 3.
Allelic variations in the GmJAZ8 gene (promoter and intragenic regions). (a) Haplotypes of GmJAZ8. (b) Box plots of the germination rate of the accessions of different haplotypes. Different colors indicate the different haplotypes. ****, p < 0.0001; ns, no significance.
GmJAZ8 promotes seed germination
-
The candidate gene Glyma.08G096500 encodes the JAZ protein named JASMONATE ZIMDOMAIN 8 (GmJAZ8). To validate the function of GmJAZ8 in seed germination, GmJAZ8 (Glyma.08G096500) overexpression vectors were constructed and then transformed into Arabidopsis, and two highly expressed lines (OE_6 and OE_28) were chosen by qRT-PCR analyses for subsequent experiments (Fig. 4a). Seeds of sterilized wild-type (Co10), OE_6, and OE_28 were first stratified at 4 °C for 3 d to break seed dormancy and were then sown in culture dishes with filter papers. The rate of seed germination was recorded at 12, 24, and 48 HAI. The results showed that the germination rate of all GmJAZ8 overexpression lines was ssignificantly higher than that of the wild-type line at 24 HAI (Fig. 4b). The seeds from all lines eventually germinated at 48 HAI. These indicated that the overexpression of GmJAZ8 promoted the speed of seed germination in OE_6 and OE_28 compared with the wild-type in Arabidopsis (Fig. 4b, c, and Supplementary Table S5).
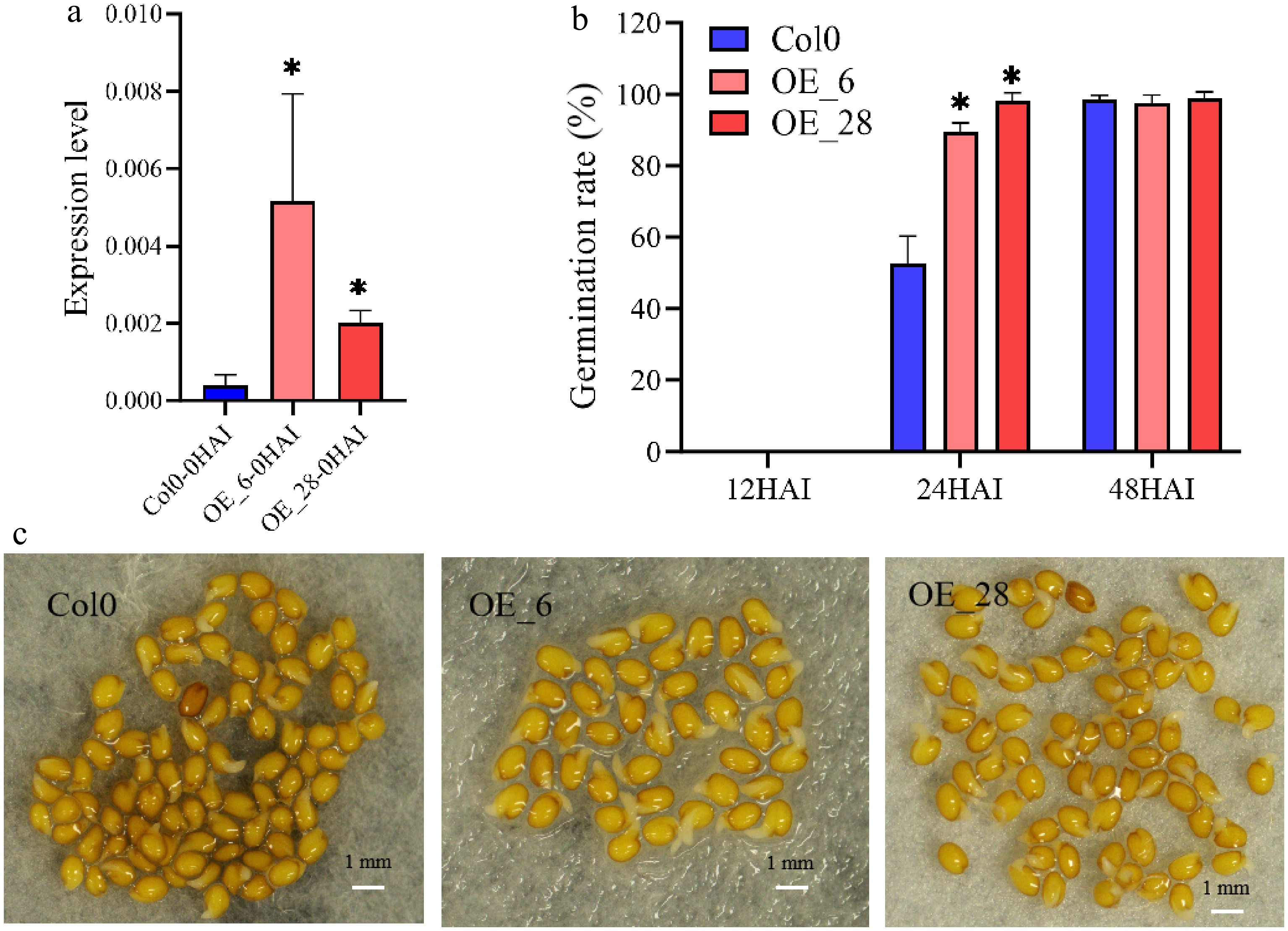
Figure 4.
GmJAZ8 promotes seed germination. (a) Relative expression of GmJAZ8 in Col0 and transgenic lines (OE_6, and OE_28) at 0 HAI. (b) The rate of germination of Col0 and the transgenic lines (OE_6, and OE_28). (c) Seedlings of the wild-type, OE_6, and OE_28 lines at 24 HAI. *, p < 0.05.
GmJAZ8 altered expression of hormone signaling genes
-
To investigate the molecular mechanisms underlying the regulation of GmJAZ8 in seed germination, RNA sequencing (RNA-seq) was performed on seeds of the wild-type (Col0) and GmJAZ8 overexpressing lines (OE_6 and OE_28) at the 0 and 24 HAI timepoints. The PCA results showed that biological replicates clustered well (Fig. 5a). Notably, Col0 and the overexpression lines (OE_6 and OE_28) were separated into two distinct clusters at 0 HAI, whereas Col0 and the overexpression lines (OE_6 and OE_28) grouped together within one cluster at 24 HAI (Fig. 5a), indicating that the differences in germination between Col0 and the overexpression lines primarily originated from the values at 0 HAI.
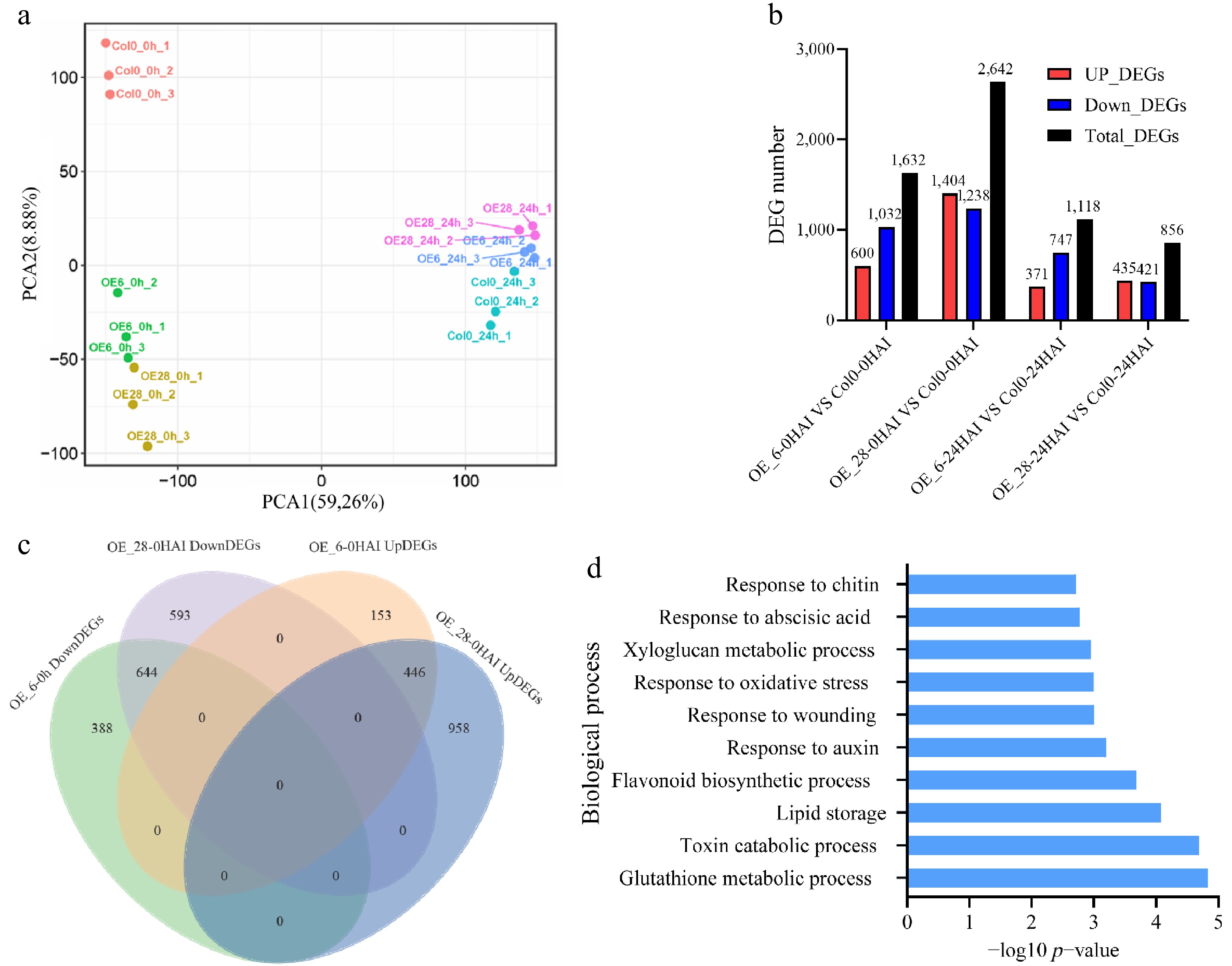
Figure 5.
Differentially expressed genes (DEGs) with p < 0.05 between the wild-type (Col0) and GmJAZ8 overexpressing lines. (a) PCA of all samples. (b) The number of DEGs. (c) Overlapping DEGs between OE_6 and OE_28 at 0 HAI. (d) GO assay of downregulated overlapping DEGs at 0 HAI.
Compared with Col0, 1,632 significant DEGs were detected in OE_6, including 600 upregulated and 1,032 downregulated genes at 0 HAI, whereas 2,642 DEGs were identified in OE_28, comprising 1,404 upregulated and 1,238 downregulated genes (Fig. 5b and Supplementary Table S6). OE_6 exhibited 1,118 DEGs (371 upregulated and 747 downregulated), whereas OE_28 showed 856 DEGs (435 upregulated, 421 downregulated) at 24 HAI (Fig. 5b and Supplementary Table S6).
Further accurate analysis of DEGs in the GmJAZ8 overexpressing lines was performed. We examined overlapping DEGs between OE_6 and OE_28 at 0 and 24 HAI. A cross-comparison of GmJAZ8 overexpressing lines revealed 1,090 overlapping DEGs at 0 HAI (664 downregulated and 446 upregulated), decreasing to 497 overlapping DEGs at 24 HAI (300 downregulated and 197 upregulated) (Fig. 5c, Supplementary Fig. S2a, and Supplementary Table S6).
GO analyses revealed that upregulated DEGs at 0 HAI were enriched in biological processes including RNA modification, recognition of pollen, the innate immune response, glycolytic processes, microtubule-based movement, the response to zinc ions, pentose-phosphate shunt, oxidative branch, the response to light stimulus, intercellular transport, and cellular carbohydrate metabolic process, whereas the upregulated DEGs at 24 HAI were enriched in the regulation of lignin biosynthetic processes, the response to phenylpropanoid, phloem transport, seed coat development, xylem development, regulation of secondary cell wall biogenesis, cytokinin-activated signaling, defense response, L-phenylalanine biosynthetic processes, and lipid transport (Fig. 5d, Supplementary Figs S2c, S2d, and Supplementary Table S6). Downregulated DEGs at 0 HAI were primarily enriched in biological processes including glutathione metabolic processes, toxin catabolic process, lipid storage, flavonoid biosynthetic processes, response to auxin, response to wounding, response to oxidative stress, xyloglucan metabolic process, response to ABA, and response to chitin. However, downregulated DEGs at 24 HAI were mainly enriched in the chitin catabolic process, the polysaccharide catabolic process, leaf senescence, response to gibberellin, programmed cell death involved in cell development, the toxin catabolic process, transmembrane transport, the response to phenylpropanoid, cell wall macromolecule catabolic process, and aging. Notably, downregulated DEGs at 0 and 24 HAI showed enrichment in hormone signaling (Fig. 5d, Supplementary Fig. S2b, and Supplementary Table S6).
ABA and GA play important roles in controlling seed germination. Notably the ABA biosynthetic gene NCED4 was significantly downregulated in OE_6 and OE_28, whereas the catabolic gene CYP707A2 was upregulated (Fig. 6). In contrast, overexpression of GmJAZ8 increased the expression of the GA biosynthetic gene GA20OX3 and repressed the expression of the GA catabolic gene GA2OX6 (Fig. 6). These suggested that a lower ABA/GA ratio was induced by the overexpression ofGmJAZ8. Additionally, the overexpression of GmJAZ8 also inhibited the expression of AtDOG1, which is another key component controlling seed germination (Fig. 6). Thus GmJAZ8 controls seed germination by repression of the ABA/GA ratio and DOG1 transcription.
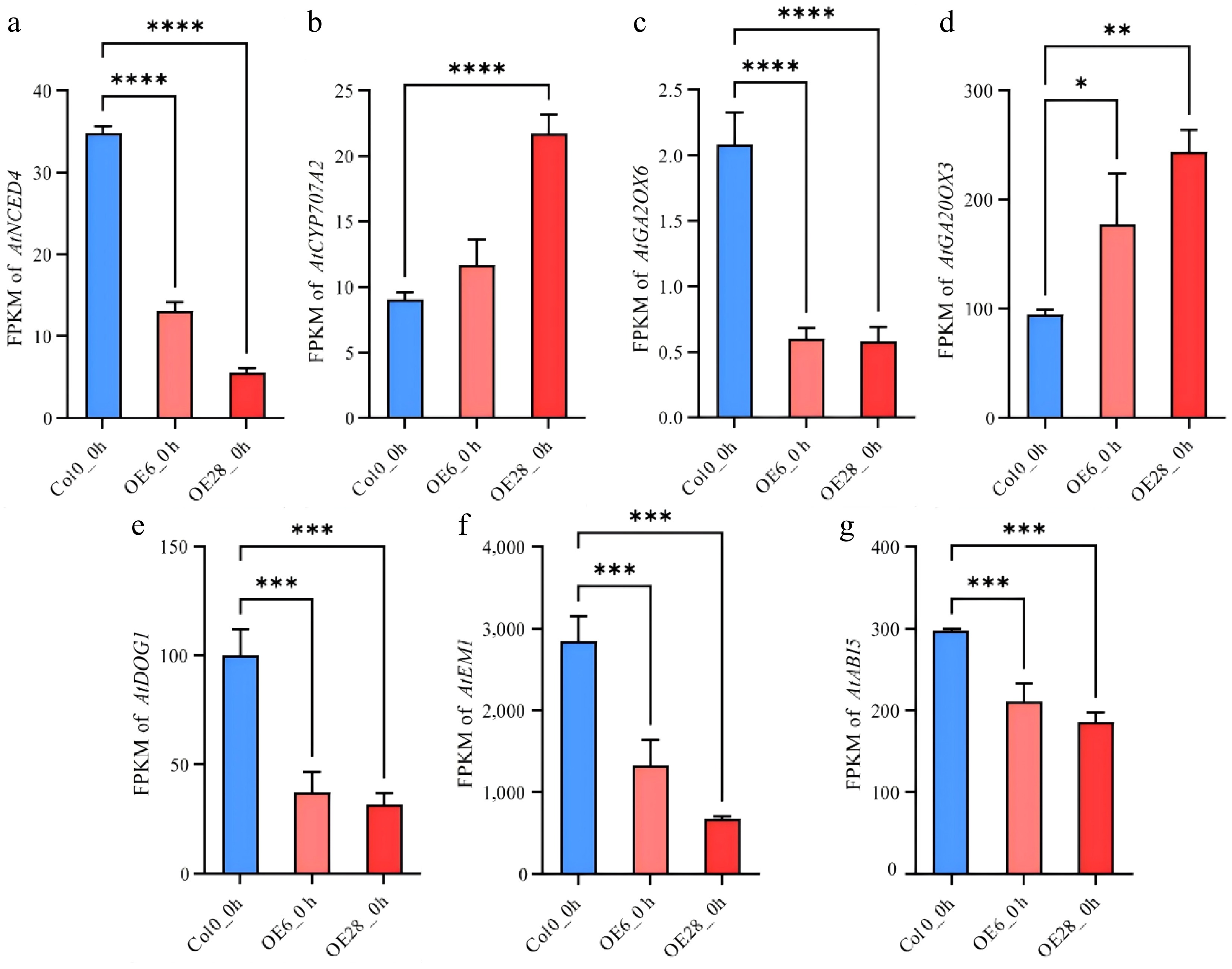
Figure 6.
GmJAZ8 altered expression of hormonal signaling genes. the FPKM values of (a) AtNCED4, (b) AtCYP707A2, (c) AtGA2OX6, (d) AtGA20OX3, (e) AtDOG1, (f) AtEM1 and (g) AtABI5. *, p < 0.05; **, p < 0.01; ***, p < 0.001; ****, p < 0.0001.
ABI5, as a master regulator in ABA signaling, negatively regulates seed germination[19,26]. JAZ proteins interact with ABI5 and suppress its transcriptional activity, thereby modulating EM1 expression levels to ultimately regulate seed germination[16,27,28]. In our results, the expression levels of AtEM1 and AtABI5 were also significantly suppressed in both the OE_6 and OE_28 lines (Fig. 6). As a result, overexpression of GmJAZ8 altered the expression of genes involved in ABA and GA biosynthesis and signaling. Taken together, these findings demonstrate that GmJAZ8-mediated regulation of seed germination may involve coordinated ABA/GA pathway interactions (Fig. 6 and Supplementary Table S7). The expression levels of these genes were validated using qRT-PCR, and the results were consistent with the transcriptomic data (Supplementary Fig. S3).
-
Seed germination is the process by which a mature, dry seed begins with imbibition and progresses to the gradual elongation of the hypocotyl until it breaks through the seed coat. As the first and most critical stage in a plant's growth cycle, seed germination plays a crucial role in determining crop yield[1]. Studies have shown a significantly positive correlation between the rate of seed germination and crop yield in soybean[4]. Soybean seeds are short-lived seeds that are prone to deterioration, leading to a significant decline in seed vigor and, consequently, a reduced germination rate. In this study, we combined GWAS and transcriptomic analyses to identify a major QTL on chromosome 8 associated with the rate of seed germination. Within this locus, GmJAZ8 was pinpointed as a key candidate gene. Functional validation in Arabidopsis demonstrated that the overexpression of GmJAZ8 accelerates seed germination. Further transcriptomic profiling revealed that GmJAZ8 likely exerts its function by modulating the expression of genes involved in ABA and GA biosynthesis and signaling, thereby lowering the ABA/GA ratio.
To increase soybean production, it is essential to dissect and understand the mechanisms involved in the regulation of seed germination. GWAS has been widely used in plant research to identify loci that are significantly associated with seed germination[29−34]. At present, several genetic loci for seed germination in soybean have been detected via linkage mapping and GWAS[3,34,35]. Sharmin et al.[35] found a major SNP, Gm_08_46239716, for the germination rate and identified a candidate gene Glyma.08G348500 encoding uridine diphosphate glycosyltransferase. Zhang et al.[3] detected a GWAS signal on chromosome 18 which was significantly associated with seed germination in soybean. In this study, GWAS revealed that 14 significant SNPs were co-localized in close proximity across multiple environments on chromosome 8, distributed within a 113.5-kb interval block (7,394,400–7,507,912 bp) (Fig. 1a–h and Supplementary Table S3).
A 113.5-kb interval block (7,394,400–7,507,912 bp) on chromosome 8 for soybean germination was identified, which contained 14 SNPs. Moreover, the key candidate gene Glyma.08G096500, which encodes a JAZ protein, was pinpointed by integrating transcriptomic analysis, homology analysis, and haplotype analysis (Figs 2, 3, and Table 2), which might be responsible for seed germination in soybean.
JAZ proteins constitute a plant-specific subfamily of the TIF[F/Y]XG domain (TIFY) superfamily characterized by TIFY and jasmonate domains. Studies have demonstrated that JAZ domains exhibit relatively conserved functional mechanisms in Arabidopsis[36,37]. The number of JAZ proteins exhibits substantial variation across plant species, with Arabidopsis thaliana possessing 12[38], rice having 15[39], Zea mays (maize) having 16[40], and soybean having 28 members (Wm82.a4.v1). Research indicates that different JAZ proteins play diverse and versatile roles in regulating distinct phenotypic traits, with JAZ1 inducing flowering[41], whereas JAZ11 regulates seed width and weight[42]. Although JAZ protein families in Arabidopsis and rice have been extensively characterized, considerably less attention has been devoted to their soybean orthologs. ASMONATE ZIM-DOMAIN 5 (OsJAZ5) involving an OsABI3-mediated ABA pathway was verified, which positively regulates seed germination in rice[18]. JAZ proteins interact with ABI3/ABI5 and suppress their transcriptional activity, thereby affecting seed germination in wheat and Arabidopsis[15,17]. Similar to previous studies[15,17,18], our GmJAZ8 overexpression lines of Arabidopsis exhibited significantly higher germination rates than the wild-type plants (Fig. 4), providing evidence that GmJAZ8 functions to promote seed germination.
ABA and GA levels in seeds are significantly correlated with seed germination[43,44]. ABA inhibits seed germination, whereas ABI5 is a key component of ABA signaling[18]. In wheat, the JAZ1 protein can directly interact with ABI5, inhibiting its transcriptional activation activity, thereby negatively regulating the ABA response and promoting seed germination[16]. Simultaneously, Arabidopsis JAZ proteins can also interfere with the activation of AtABI5 by the Indole-3-acetic acid (IAA)-dependent transcription factors AtARF10 and AtARF16, thus affecting seed germination[18]. In rice, OsJAZ6/7 can interact with OsABI3/5, influencing the transcriptional activity of OsABI3/5, and OsABI3/5 can participate in the process of regulating seed germination mediated by the UDP glucosyltransferase OsUGT75A[45]. Overall, these findings reveal that JAZ proteins can participate in ABA hormone signaling in coordination to regulate seed germination.
ABA is biosynthesized by NCED and ABA genes[8,46,47], and catalyzed by CYP707As[48]. GA is mainly biosynthesized by GA3OXs[49,50] and GA20OXs[51,52], and catalyzed by GA2OXs[53,54]. JAZ proteins regulate seed germination through a dual mechanism. By suppressing the activity of the transcription factor MYC2, they modulate the stability of Asp-Glu-Leu-Leu-Ala (DELLA) proteins, thereby indirectly but effectively influencing the expression of GA biosynthesis/metabolism-related genes[36]. This results in the suppression of GA biosynthetic genes (e.g., GA20OX) and the activation of GA catabolic genes (e.g., GA2OX). Simultaneously, JAZ proteins regulate ABA biosynthesis/metabolism genes by inhibiting transcription factors such as MYC2, which can bind to the promoters of ABA biosynthesis genes (e.g., NCED). This leads to downregulation of NCED transcription and reduced ABA levels. Seed germination is ultimately controlled through by coordinated regulation[36]. When the expression levels of ABA synthesis and GA metabolic genes decrease, or the expression levels of GA synthesis and ABA metabolic genes increase, the ABA/GA content ratio decreases, which promotes seed germination[55,56].
In our results, GmJAZ8-overexpressing lines exhibited upregulated expression of the ABA catabolic gene AtCYP707A2 but downregulated expression of the ABA biosynthesis gene AtNCED4 (Fig. 6). GA–ABA antagonism fundamentally operates through bidirectional suppression of hormone biosynthesis, with GA inhibiting ABA production and ABA repressing GA synthesis, thus collectively gate-keeping the initiation of germination[5,57,58]. By upregulating a GA biosynthetic gene (GA20OX) while suppressing a GA catabolic gene (GA2OX), seed germination can be effectively promoted[59]. Consistent with this established mechanism, our findings demonstrate that in GmJAZ8-overexpressing lines, AtGA20OX3 expression was significantly upregulated, whereas AtGA2OX6 transcript levels were markedly downregulated (Fig. 6). Transcription factors such as ABI3 and ABI5 function as master regulators in ABA signaling and negatively regulate seed germination[25]. Studies have shown that JAZ proteins interact with ABI3/ABI5, suppress their transcriptional activity, and modulate ABA signaling, thereby affecting seed germination in species such as wheat, rice, and Arabidopsis[15,18]. JAZ proteins interact with ABI5 and suppress its transcriptional activity, thereby modulating EM1 expression levels to ultimately regulate seed germination[15,26,27]. In the GmJAZ8-overexpressing lines (OE_6 and OE_28), transcript levels of ABI5 and EM1 were significantly downregulated, whereas the germination rate markedly increased (Fig. 6). We therefore propose that the accelerated germination in GmJAZ8-overexpressing lines may result from the altered expression of hormone signaling components.
Collectively, this GWAS pinpointed GmJAZ8 as a pivotal regulator of the rate of seed germination. Functional validation demonstrated that GmJAZ8 overexpression enhances the germination rate. Mechanistically, GmJAZ8 likely modulates the germination rate by integrating ABA and GA signaling cascades.
-
In this study, we performed a GWAS and identified a QTL for the rate of seed germination in soybean on chromosome 8. By using transcriptomic analysis, cross-species homology alignment, and haplotype-based selection analysis, GmJAZ8 was identified as a key candidate gene which regulates soybean's germination rate. GmJAZ8 was then confirmed to accelerate the rate of seed germination by regulating the transcriptional levels of genes involved in ABA and GA signaling. Thus GmJAZ8 might be useful for improving the rate of seed germination in soybean breeding programs.
This study was supported by the grants from Xianghu Laboratory for Special Research Project (2023C2S02001).
-
The authors confirm their contributions to the paper as follows: funding support, study conception and design: Yuan F, Chen X, Zhao J; material treatment, data analysis, and manuscript writing: Xu W, Yang Y, Liu B, Chen Y; material preparation: Deng P, Yu H. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Fig. S1 Expression analysis of 15 candidate genes in soybean.
- Supplementary Fig. S2 Differentially expressed genes(DEGs) with p < 0.05 between WT(Col0) and GmJAZ8 over-expressing lines.
- Supplementary Fig. S3 The expression of all genes in Figure 6 at 0 HAI and 24 HAI.
- Supplementary Table S1 The information and germination rate of soybean re-sequence population.
- Supplementary Table S2 The name and sequence of primers.
- Supplementary Table S3 The information of SNPs from GWAS.
- Supplementary Table S4 The FPKM of 13 genes and germination rate between soybean accessions.
- Supplementary Table S5 The germination rate of GmJAZ8-overexpression.
- Supplementary Table S6 The DEGs in GmJAZ8-overexpression lines.
- Supplementary Table S7 Transcriptional levels of hormone related genes in GmJAZ8-overexpression lines.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xu W, Yang Y, Liu B, Chen Y, Deng P, et al. 2026. Genome-wide association study reveals that GmJAZ8 is involved in the germination of soybean seeds. Seed Biology 5: e006 doi: 10.48130/seedbio-0026-0003 

Genome-wide association study reveals that GmJAZ8 is involved in the germination of soybean seeds
- Received: 09 October 2025
- Revised: 27 November 2025
- Accepted: 12 January 2026
- Published online: 16 March 2026
Abstract: Soybean (Glycine max L.) is one of the major oil and economic crops for human beings. Seed germination is a critical stage in soybean's growth cycle and plays a crucial role in determining soybean yield, yet only few genes controlling the rate of seed germination have been characterized in soybean. Here, we performed a genome-wide association study (GWAS) and identified a quantitative trait locus (QTL) for the rate of seed germination on chromosome 8. By using transcriptomic analysis, cross-species homology alignment, and haplotype-based selection analysis, we identified GmJAZ8 (Glyma.08G096500) as a key candidate gene regulating soybean's germination rate. We further confirmed that GmJAZ8 accelerated the rate of seed germination by regulating the transcriptional levels of genes involved in abscisic acid and gibberellin signaling. Thus GmJAZ8 might be useful for improving the rate of seed germination in breeding soybean.
-
Key words:
- Soybean /
- The rate of seed germination /
- Genome-wide association study /
- JAZ protein