










-
Genetic transformation underpins research in plant genetics and molecular breeding. Exogenous gene delivery is mainly achieved through Agrobacterium tumefaciens-mediated transformation and particle bombardment[1]. However, these conventional approaches typically require labor-intensive and time-consuming tissue-culture procedures and often show limited efficiency. Given the growing concerns over food security, there is a need to develop transformation strategies that are simpler, faster, and more efficient to accelerate progress in molecular breeding of plants.
Previous studies have demonstrated that specific in vitro tissues from various plant species exhibit high regeneration potential. For example, isolated roots of Mongolian dandelion (Taraxacum mongolicum) and Rehmannia glutinosa display a robust capacity to regenerate into complete plants[2]. This regeneration potential underpins the development of the cut–dip–budding (CDB) genetic transformation method, which bypasses conventional sterile tissue culture in these species[3]. Ectopic expression of developmental regulators (DRs) genes can further enhance regeneration. In maize, overexpression of WUSCHEL2 (WUS2) promotes somatic embryogenesis and improves transformation-positive frequency[4]. In soybean (Glycine max), expressing chimeric growth-regulating factors (GRFs) fused with GRF-interacting factors (GIFs) markedly increases the efficiency of regeneration and transformation[5]. Isopentenyl transferase (ipt), a key enzyme in cytokinin biosynthesis, has also emerged as a research focus: Its overexpression elevates endogenous cytokinin levels, thereby stimulating axillary bud proliferation and promoting adventitious bud formation in multiple plant species[6]. However, most of these strategies still rely on, or partially involve, tissue culture procedures that are complex and require sterile conditions.
Suaeda glauca is an annual halophytic herbaceous plant with both forage and medicinal value[7,8]. As a pioneer species for ecological restoration of saline-alkaline soils in northern China, it contributes to soil improvement, habitat provision, and biological carbon sequestration[9]. Moreover, S. glauca represents an important reservoir of salt-tolerance genes, offering valuable genetic resources for enhancing salt tolerance in conventional crops[10]. Nevertheless, reference protocols for its genetic transformation remain extremely limited. Soybean is one of the most important crops worldwide and is a major source of plant protein for food and feed. Although multiple Agrobacterium-mediated transformation systems have been developed for soybean[11], tissue culture steps still constitute the major bottleneck. Transformation-positive frequency is strongly affected by the genotype, Agrobacterium density, duration of co-cultivation, and the medium's composition[11−13], and the tissue culture process is highly susceptible to microbial contamination. Consequently, the development of innovative genetic transformation strategies has become a pressing priority.
Building on the direct delivery genetic transformation method established by Maher et al.[6], we implemented several improvements, including optimization of the co-cultivation medium's composition, expansion of the vector's payload capacity, redesign of the multiple cloning site (MCS), and incorporation of epitope tags. On this basis, we established a streamlined transformation system characterized by simple operation, a high proportion of positive events, and the ability to generate heritable transgenic lines with transmission to T1 progeny from the T0 founders. We demonstrate the utility of this approach in S. glauca and soybean, providing a practical route to reduce reliance on tissue culture.
-
The soybean germplasm used in this study comprised ChiDou, Williams 82, Qihuang 34 (QH34), Jidou 12 (JD12), Cuishan 2 (CS2), Qindou 2018 (QD2018), and Zhonghuang 13 (ZH13). The halophytic species used in this study was Suaeda glauca.
Preparation of Agrobacterium suspensions
-
Plasmid constructs were introduced into A. tumefaciens strains GV3101-P19 and EHA105 by chemical transformation. Transformants were selected on Yeast Extract Peptone (YEP) agar plates containing the appropriate antibiotics. A single colony was inoculated into 20 mL of the YEP liquid medium with antibiotics and grown overnight at 28 °C with shaking at 200 rpm. Cells were harvested by centrifugation at 4,000×g for 10 min at room temperature and resuspended in the co-cultivation medium (YEP, and antibiotics) to an optical density (OD600) of 0.3, followed by incubation at 28°C with shaking at 200 rpm overnight. The culture was centrifuged again at 4,000×g for 10 min, the supernatant was discarded, and the pellet was resuspended in an infection solution (10 mM 2-[N-morpholino] ethanesulfonic acid [MES], 10 mM MgCl2, pH 5.6) to an OD600 of 0.1–0.3. The suspension was incubated at room temperature in the dark for 2–4 h before use.
Agrobacterium-mediated transformation
-
The apical meristem of soybean seedlings was inoculated with Agrobacterium suspension using a syringe. The plants were then transferred to a growth chamber and maintained at 26 °C under a 16/8 h light/dark photoperiod. For S. glauca, the apical meristems of the plants were inoculated with Agrobacterium suspension using a syringe following the same method, and the plants were subsequently cultured in a growth chamber at 24 °C under a 13/11 h light/dark photoperiod.
Identification of positive plants
-
For the T0 generation plants, two or three of the uppermost leaves were sampled for detection. For the T1 generation plants, the first true leaf was collected from the aboveground parts and one random root was selected from the underground parts for analysis. Leaf tissue was ground, and genomic DNA from the T0 and T1 plants was extracted using the cetrimonium bromide (CTAB) method[14]. Polymerase chain reaction (PCR) was performed with P222 DNA polymerase using genomic DNA as the template and the primers listed in Supplementary Table S1, following the manufacturer's instructions (Nanjing Vazyme Biotechnology Co., Ltd.).
For luciferase complementation imaging, infiltrated leaves were sprayed with 1 mM luciferin (Yeasen Biotechnology, Shanghai, China) in the dark and incubated for 10 min. Luminescence signals were recorded using a multispectral imaging system (PlantView100; Biolight Co., Ltd.).
-
To establish a direct delivery transformation system applicable to soybean and S. glauca, we adapted and optimized the co-cultivation medium and infection procedure described by Maher et al.[6]. The apical buds of soybean and S. glauca seedlings were selected as the infection sites. (Fig. 1). The apical meristem of the plants was infected with the Agrobacterium suspension using a syringe. (Fig. 1b, c). After inoculation, plants were subjected to dark co-cultivation and then grown under species-specific conditions. During cultivation, axillary buds emerging from nonwounded sites were removed.
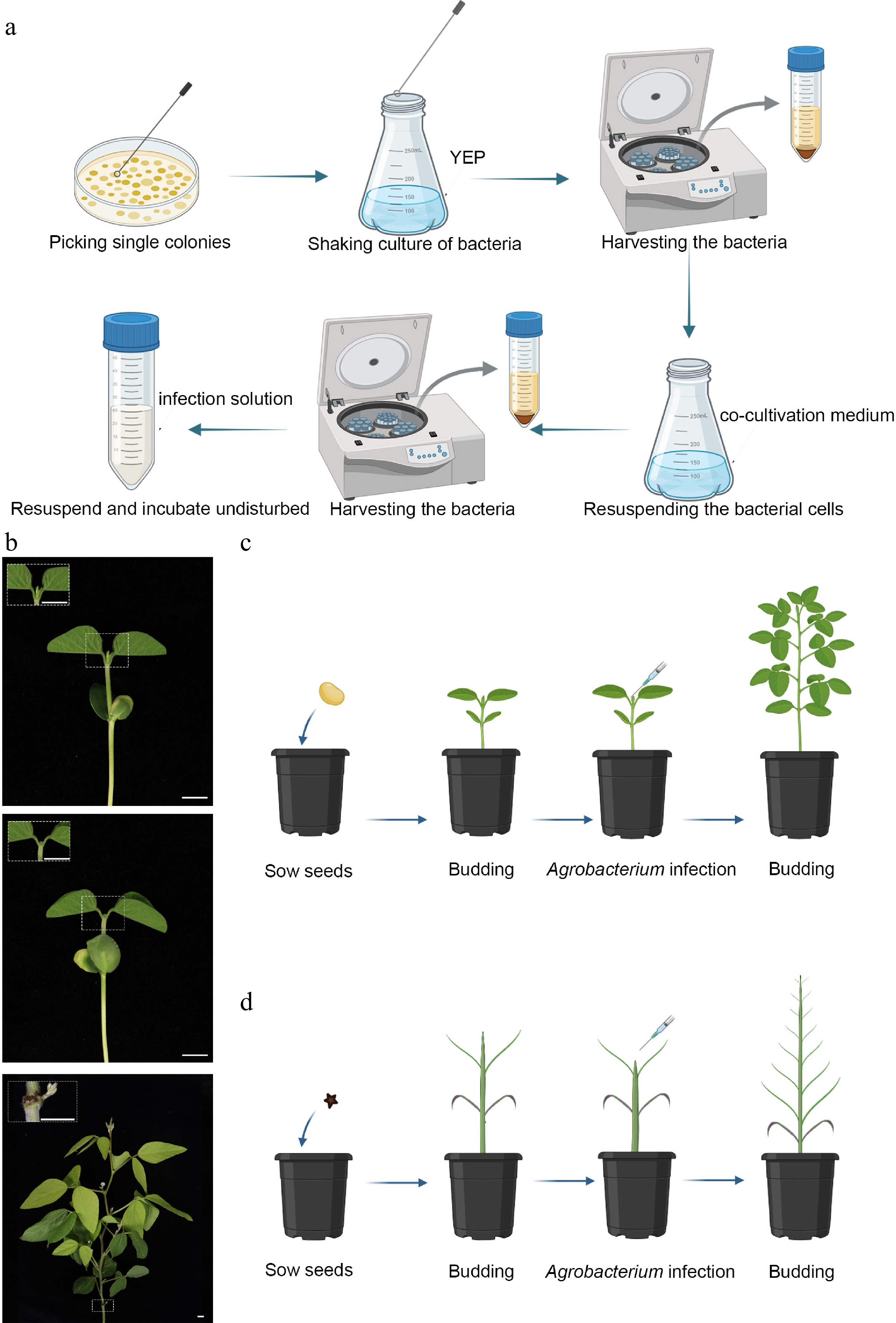
Figure 1.
Gene transformation methods for soybean and S. glauca. (a) Preparation of the A. tumefaciens suspension. A single colony was picked and inoculated into YEP medium, followed by incubation with overnight shaking under optimal conditions. The bacterial cells were harvested, the OD600 value was adjusted, and the culture was transferred to the co-cultivation medium for an additional overnight incubation. The cells were then collected again, resuspended in the infection solution, and incubated in the dark until use. Figure was created with BioRender;
https://app.biorender.com/illustrations/canvas-beta/6945203f781eec89c8bf494e . (b) Tissue culture-free organ regeneration of soybean. After sowing, the plants were grown to the seedling stage; positive shoots appear 1–2 weeks after inoculation, and transgenic seeds can be harvested after about 4 months. Scale bar = 1 cm. (c) Schematic representation of soybean transformation. Figure was created with BioRender;https://app.biorender.com/illustrations/canvas-beta/6945203f781eec89c8bf494e . (d) Schematic representation of S. glauca transformation. The time from sowing to the first pair of cotyledons takes about 1 week, positive shoots appear within 1–2 weeks after infection, and transgenic seeds can be harvested after about 5 months. Figure was created with BioRender;https://app.biorender.com/illustrations/canvas-beta/6945203f781eec89c8bf494e .Using the optimized protocol, the pMKV057 vector harboring LUC, WUS2, and ipt[6] was introduced into multiple soybean cultivars via A. tumefaciens strain EHA105. Transgenic plants were identified by ipt-specific PCR, yielding an overall positive transformation frequency of 73.7% (PCR-positive plants/total inoculated plants) (Fig. 2b). In addition, luciferase fluorescence signals were detected in the majority of leaves (Fig. 2c). Compared with the nontransgenic control plants, positive transformants displayed normal growth and development and were able to flower and set seeds successfully (Fig. 2d, e).
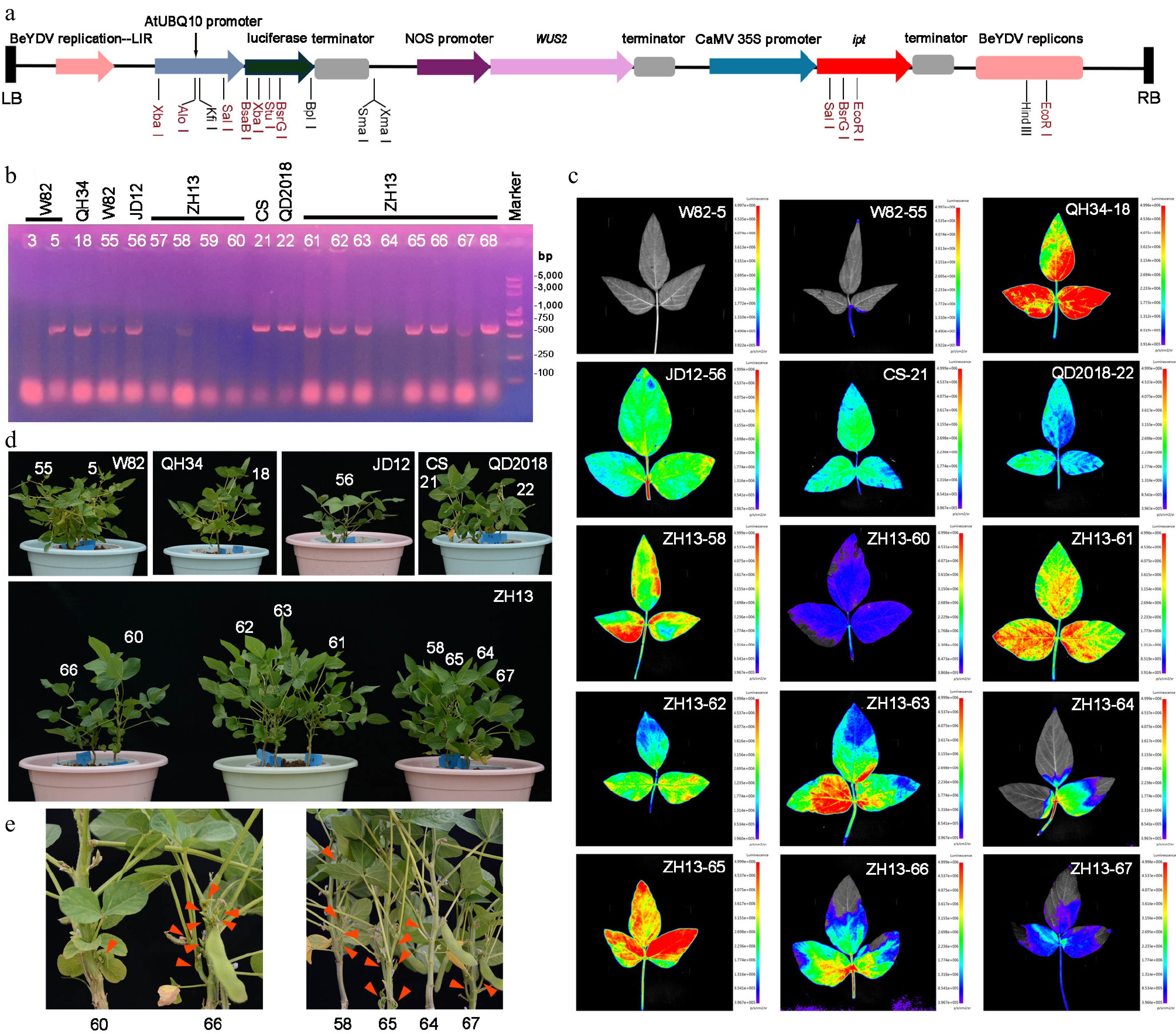
Figure 2.
Identification of transformed soybean seedlings across different cultivars. (a) Expression cassette of the vector pMKV057. Black labels indicate single-restriction enzyme sites, whereas red labels indicate double-restriction enzyme sites. (b) PCR-based identification of the ipt gene in transformed seedlings from different cultivars. Transformation-positive frequency = number of PCR-positive plants (14)/total number of surviving inoculated plants (19) = 73.7%. This experiment was independently repeated twice, and representative experimental data are presented in this study. (c) Detection of luciferase fluorescence in transformed seedlings across different cultivars. (d) Growth performance of transformed seedlings. (e) Flowering and pod development in transformed seedlings and negative controls (red arrows indicate flowers).
Because the original pMKV057 backbone limited modular cloning and lacked unique sites suitable for straightforward replacement of the LUC cassette, we generated derivative vectors to increase cloning flexibility and cargo capacity. Specifically, two constructs separately expressing WUS2 and ipt were generated. The LUC expression cassette was removed and replaced with modular expression cassettes incorporating epitope tags (FLAG and MYC) to facilitate detection of heterologous proteins, whereas the original promoter was replaced with a legume-active promoter. In addition, the multiple cloning site (MCS) was redesigned to include unique, high-efficiency restriction sites, and an optimal Kozak sequence was inserted immediately upstream of the MCS to support efficient translation, multigene assembly, and co-transformation. Redundant restriction sites outside the MCS were eliminated without altering the encoded amino acid sequence, thereby facilitating the subsequent co-transformation of exogenous genes (Fig. 2a, 3a). When the modified vectors were introduced into the soybean germplasm 'ChiDou' via A. tumefaciens GV3101-mediated transformation, the transformation-positive frequency reached 57.6%–65.4%, and the ipt-positive shoots exhibited normal development and seed set (Fig. 3b, c, Supplementary Fig. S2, Supplementary Table S2). Furthermore, application of this system to S. glauca resulted in a ipt-positive transformation rate of 73.1% (Fig. 4a).
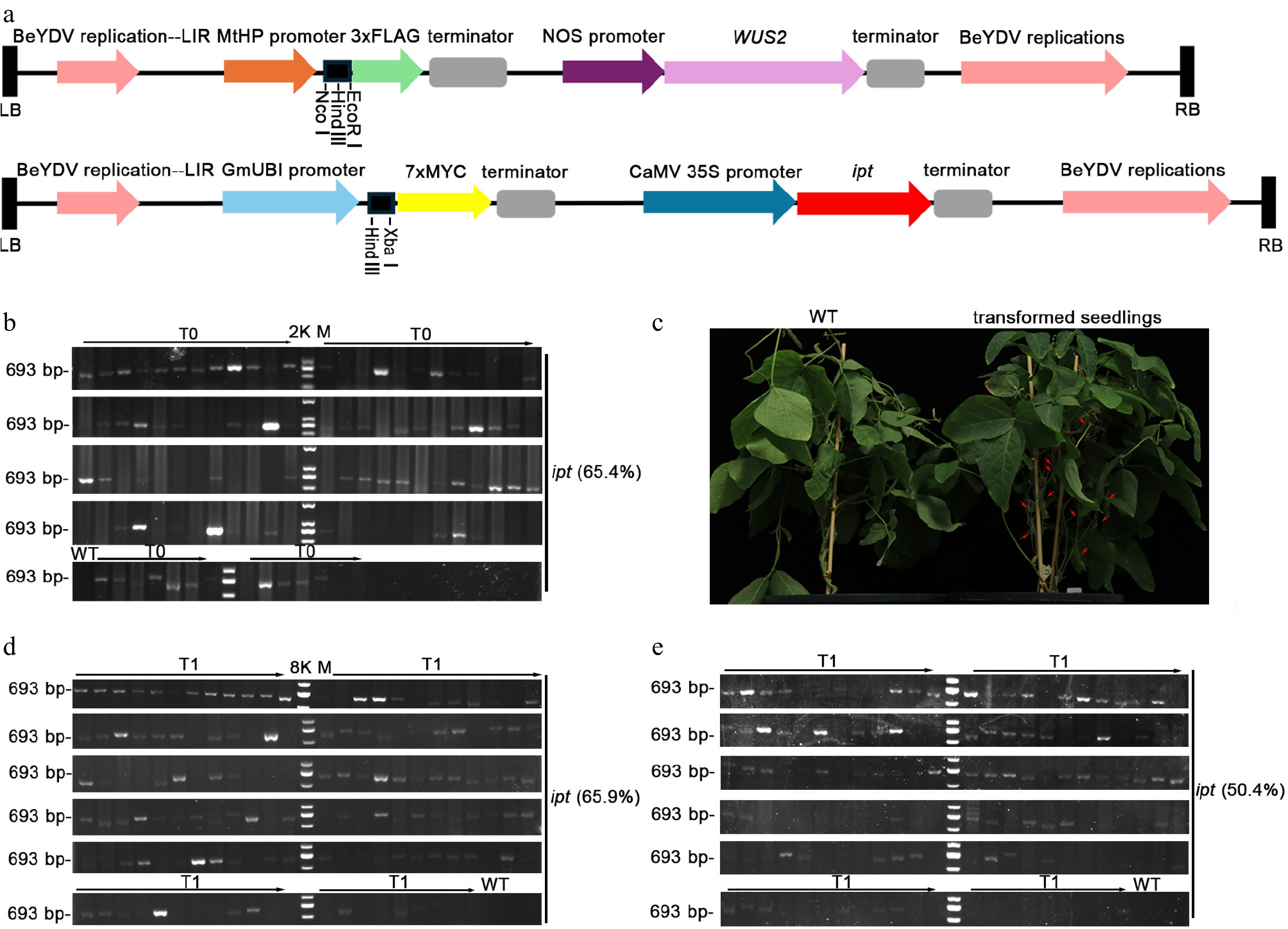
Figure 3.
Vector schematic and transformation-positive frequency. (a) Schematic representation of the vector's expression cassette (single-restriction enzyme sites are indicated in black). (b) Transformation-positive frequency in the soybean germplasm ChiDou. Transformation-positive frequency = number of PCR-positive plants (72)/total number of surviving inoculated plants (110) = 65.4%. This experiment was independently repeated twice, and representative experimental data are presented in this study. (c) Phenotype of transformed seedlings. Scale bar = 1 cm. (d) Transformation-positive frequency in the aboveground parts of T1 generation of the soybean germplasm ChiDou. Transformation-positive frequency = number of PCR-positive plants (93)/total number of surviving inoculated plants (141) = 65.9%. (e) Transformation-positive frequency in the underground parts of the T1 generation of the soybean germplasm ChiDou. Transformation-positive frequency = number of PCR-positive plants (71)/total number of surviving inoculated plants (141) = 50.4%.

Figure 4.
Identification of T1 transgenic S. glauca seedlings. (a) Identification of T0 transgenic S. glauca. Transformation-positive frequency = number of PCR-positive plants (68)/total number of surviving inoculated plants (93) = 73.1%. Transformation-positive frequency = number of PCR-positive plants (19)/total number of surviving inoculated plants (24) = 79.2%. This experiment was independently repeated twice, and representative experimental data are presented in this study. (b) Phenotype of T1 transgenic S. glauca. Scale bar = 1 cm. (c) Detection of the ipt gene in T1 transgenic S. glauca.
To assess heritable transmission, the T1 progeny of S. glauca and ChiDou derived from independent T0 events were identified (Fig. 4). Because we were unable to obtain sufficient numbers of healthy, highly viable seeds from either wild-type or transgenic S. glauca grown in the growth chamber, T1 analyses were performed using a single T0 event that produced adequately filled seeds with acceptable germination. Compared with the wild-type plants, the T1 progeny of S. glauca displayed slightly altered leaf morphology but no apparent developmental abnormalities, and they flowered and set seeds normally (Fig. 4b). The ipt gene, which was absent in the wild-type, remained detectable in T1 plants (Figs. 3d, e and 4a, c). Through separate identification of the aboveground and underground parts of the T1 generation of ChiDou soybean plants, we found that some chimeras still existed among the harvested T1 plants (Fig. 3d, e). In this study, 16 independent T0 transformation events were randomly selected, and 6–9 T1 plants from each event were subjected to detection. Based on the combined identification results of the aboveground and underground parts, the positive rate was calculated using plants with target bands amplified in both tissues, which was approximately 45.4%. (Supplementary Table S3). Together, these results indicate that the exogenous genes introduced using this transformation strategy can be stably transmitted to the next generation.
-
By optimizing a previously reported direct delivery genetic transformation approach[6], we established a rapid and efficient plant transformation system that can be operated under nonsterile conditions without reliance on sterile in vitro tissue culture. First, this system eliminates the need for sterile facilities and complex media, whereas conventional transformation methods relying on adventitious bud differentiation and rooting typically depend on tissue culture techniques[6,11,12]. In recent years, several tissue culture-free transformation strategies for soybean have been developed; however, they mainly use germinating seeds or hypocotyls after embryo excision as the recipients and require highly precise technical skills[15−17]. In contrast, our method involves inoculation at the wound site after removal of the apical bud at the unifoliolate stage, uses a simple medium formulation, and substantially lowers the technical threshold. Moreover, because the wound site is spatially distant from the soil, the risk of fungal contamination is effectively minimized. Second, by exploiting the functions of the developmental regulators WUS2 and ipt, our approach does not require supplementation with exogenous hormones. Inoculated plants regenerated shoots at a high frequency, achieving a transformation-positive frequency exceeding 57.6%, and the introduced traits were stably inherited by the progeny. Moreover, the positive rate of T1 generation plants derived from multiple independent T0 transformation events was approximately 45.4%. Recently, Cao et al. demonstrated that soybean exhibits strong in situ tissue and organ regeneration capacity, and that vectors co-expressing ipt with other developmental regulators markedly enhance regeneration and transformation-positive frequency. Although the transformation-positive frequency was only 0.42% in the absence of developmental regulators, it achieved 4.68% following their introduction[16]. Alok et al. likewise successfully applied ipt and GmWUS2 for transforming soybean. However, their system remains confined to sterile environments, with a maximum transformation-positive frequency of 22.3%[17]. Third, T0 founder plants can be recovered without in vitro rooting and can transmit the transgene to their T1 progeny. Only 2–3 weeks are required from preparation to the appearance of positive shoots, with 4–5 months to harvest transgenic seeds. Compared with traditional protocols that involve independent stages of adventitious bud induction and rooting, our system omits these time-consuming steps and shortens the experimental cycle by approximately 6–11 weeks[12,18]. Fourth, this strategy shows a degree of general applicability: In addition to soybean, it can also be applied to species that lack well-established tissue culture systems, such as S. glauca. Collectively, these improvements, together with their successful application across multiple species, suggest that this method may accelerate the deployment of genetic engineering technologies in basic research and agricultural production pending validation across broader genotypes and taxa.
In an early study, Maher et al. showed that co-expression of ipt and WUS2 in tobacco (Nicotiana benthamiana) very effectively promoted de novo meristem formation but also led to abnormal phenotypes and sterility[6]. Similarly, Cao et al. and Kshetry et al. observed aberrant development in regenerated soybean seedlings expressing ipt[16,19]. By contrast, neither our study nor that of Alok et al. detected abnormal shoot development, and fertile transgenic progeny were obtained[17]. These differences may be related to variation in the developmental stage used for inoculation, as well as the constructs' architecture (e.g., promoter choice), the expression level, the genotype, and the tissue context.
Nevertheless, in S. glauca, we observed leaf phenotypes that differed from those of the wild-type. Similarly, Debernardi et al. showed that transformation with GRF4–GIF1 increased spikelet number per inflorescence in wheat[20]. Together, these observations suggest that the introduction of exogenous developmental regulators may, to some extent, perturb intrinsic growth traits. Although empty-vector controls can help disentangle these effects, future work should aim to minimize the physiological influence of WUS2 and ipt (e.g., by using inducible or tissue-specific expression, transient delivery, or excisable regulator cassettes), thereby enabling a more accurate assessment of the consequences of exogenous gene expression or endogenous gene editing mediated by this system. This issue represents a broader challenge for transformation platforms that rely on developmental regulators. Mei et al. successfully obtained stably heritable sweet potato (Ipomoea batatas) plants using the regenerative activity–dependent in planta injection delivery (RAPID) method without introducing any developmental regulatory factors[21]. However, the progeny seedlings obtained by RNA virus-mediated gene editing are virus-free and harbor biallelic or homozygous mutations. Nevertheless, the applicability of these approaches in soybean remains to be further explored[22].
-
This study establishes a direct-delivery transformation system applicable to both soybean and S. glauca. The procedure can be carried out under nonsterile, tissue culture-free conditions with a streamlined workflow, a short turnaround time, and a high transformation-positive frequency. Transgene-positive T0 plants were obtained without an in vitro rooting step, and the inheritance of the transgene was confirmed in the T1 offspring.
-
The authors confirm their contributions to the paper as follows: designed the research: Liu J, Yang W; performed the research: Lin X, Zhang X, Lin L, Cao Y; analyzed the data: Lin X, Liu J; wrote the manuscript: Lin X, Liu J, Liu H, Yang W, Wang H. All authors reviewed the results and approved the final version of the manuscript.
-
The pMKV057 vector is available from Addgene, and the corresponding plasmid map can be obtained from the repository. Sequence information for the developmental regulators (DRs) is provided in Supplementary Fig. S1. The nucleotide sequences of the oligonucleotide primers are listed in Supplementary Table S1.
-
This work was supported by the National Natural Science Foundation of China (32573581, 32071871), the Key Research and Development Projects in Shaanxi Province of China (2024NC-YBXM-005), the Interdisciplinary Frontier Innovation Team Program of Northwest A&F University (A1080524001), the National Key R&D Program of China (2025YFE0200400), and the Biological Breeding-National Science and Technology Major Project (No: 2024ZD04079).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xizhen Lin, Xinfei Zhang, Lin Lin
- Supplementary Table S1 List of oligo sequences.
- Supplementary Table S2 Transformation‑positive frequency of ChiDou and S. glauca.
- Supplementary Table S3 Transgene segregation ratio of ChiDou T1 plants.
- Supplementary Fig. S1 Sequences of developmental regulators cloned in T-DNA constructs.
- Supplementary Fig. S2 Transformation‑positive frequency in the germplasm ChiDou.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Lin X, Zhang X, Lin L, Liu H, Cao Y, et al. 2026. A rapid and efficient plant genetic transformation system based on direct delivery. Seed Biology 5: e007 doi: 10.48130/seedbio-0026-0012 

A rapid and efficient plant genetic transformation system based on direct delivery
- Received: 29 December 2025
- Revised: 09 February 2026
- Accepted: 22 February 2026
- Published online: 25 March 2026
Abstract: Genetic transformation of plants underpins functional genomics and molecular breeding, but many plants are still constrained by slow, labor-intensive, tissue culture-dependent methods, including Glycine max (soybean), and Suaeda glauca. Here we develop a rapid, tissue culture-free Agrobacterium-mediated transformation approach based on targeted wounding of the seedlings' shoot apical region, followed by Agrobacterium inoculation under nonsterile conditions. Using this direct delivery strategy, we obtained transgenic plants in the same generation, with a transformation-positive frequency exceeding 57.6%. Transgenic lines transmitted the introduced gene to their progeny, supporting stable inheritance across generations. This streamlined method substantially lowers technical barriers and accelerates transgenic production in recalcitrant crops and halophyte species, offering a practical platform for rapid trait engineering and functional gene validation.
-
Key words:
- Direct delivery /
- Developmental regulators /
- Soybean /
- Suaeda glauca