









-
Precise control of the plant life cycle is essential for ensuring reproductive success and ecological adaptation. In response to environmental fluctuations, plants have evolved sophisticated molecular mechanisms to coordinate transitions between developmental phases, including germination, vegetative growth, flowering, dormancy, and senescence[1]. Among these transitions, the switch from vegetative to reproductive growth, commonly referred to as flowering, represents a critical developmental decision that profoundly influences plant fitness and agricultural productivity.
Over the past two decades, significant progress has been made in elucidating the genetic and epigenetic networks that govern flowering time, where a set of 'floral pathway integrators' has been identified[1]. At the core of this pathway is the floral repressor gene FLOWERING LOCUS C (FLC), which encodes a MADS-box family transcription factor (Fig. 1a). FLC directly inhibits the expression of key floral activators, most notably FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1), which act as floral integrators in the shoot apical meristem (SAM)[2,3]. The expression of FLC is tightly controlled by both genetic and epigenetic mechanisms, with FRIGIDA (FRI) serving as a major upstream activator (Fig.1a). In addition, the photoperiod-responsive transcription factor CONSTANS (CO) functions upstream of FT as a central regulator linking circadian and light signals to flowering-time control (Fig.1a)[4].
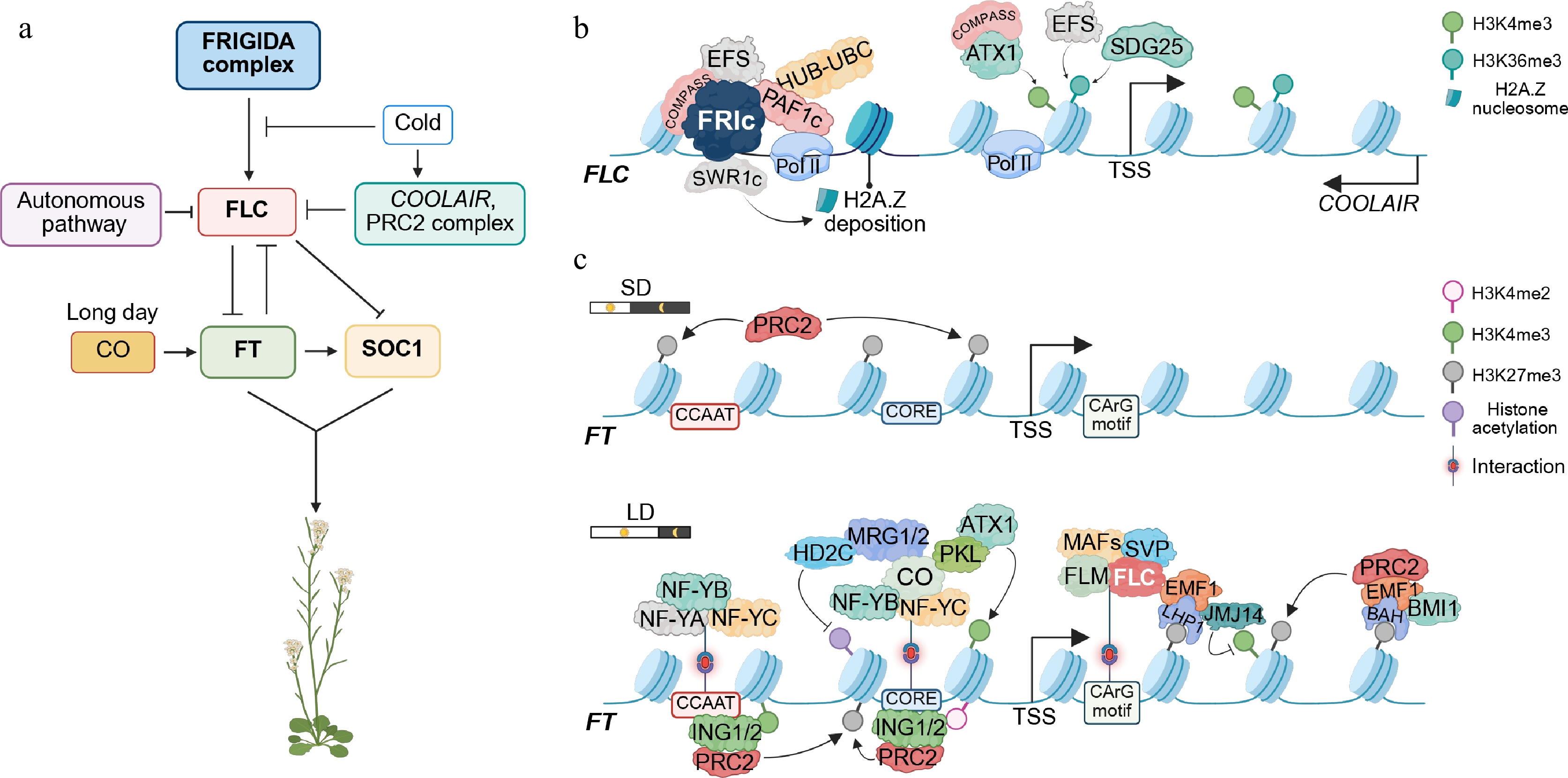
Figure 1.
Multilayered epigenetic regulation of the flowering genes FLC and FT in Arabidopsis. (a) Genetic circuitry overview. The FRIGIDA (FRI) complex delays flowering by activating the transcription of FLOWERING LOCUS C (FLC). Prolonged cold exposure (vernalization) induces COOLAIR, which silences FLC in parallel with Polycomb repressive complexes 2 (PRC2). The repression of FLC enables transcription of FLOWERING LOCUS T (FT), which subsequently upregulates the expression of SUPPRESSOR OF OVEREXPRESSION OF CO 1 (SOC1), APETALA1 (AP1), LEAFY (LFY), and SEPALLATA3 (SEP3), thereby specifying floral meristem identity and floral organ development. (b) FRIGIDA-dependent activation of FLC. The FRI complex (FRIc) recruits multiple chromatin remodelers, including the COMPASS-like complex (mediating H3K4me3, trimethylation of lysine 4 on histone H3), EARLY FLOWERING IN SHORT DAYS (EFS, also known as SDG25, a histone lysine methyltransferase responsible for H3K36me3), PAF1 complex-HUB-UBC module (PAF1c-HUB-UBC, mediating H2B mono-ubiquitylation), and the SWR1 chromatin remodeling complex (SWR1c, responsible for H2A.Z deposition), all of which establish a chromatin environment conducive to the activation of FLC expression. (c) Dynamic regulation of FT. Under short-day (SD) conditions, FT expression is constitutively repressed by Polycomb group (PcG) complexes, and thus, the floral transition is inhibited. Under long-day (LD) conditions, a trimeric NUCLEAR FACTOR Y (NF-Y) complex (composed of NF-YA, NF-YB, and NF-YC subunits) first binds to the CCAAT motif to increase chromatin accessibility. CONSTANS (CO) together with NF-Y binds the CORE element and recruits ARABIDOPSIS TRITHORAX 1 (ATX1), MORF-RELATED GENE 1/2 (MRG1, MRG2), and the chromatin remodeler PICKLE (PKL, a CHD3-type chromatin remodeler), depositing the activating mark H3K4me3 and inducing FT expression. Following FT activation at dusk, INHIBITOR OF GROWTH 1 (ING1) and ING2 recognize H3K4me2/me3 modifications on FT chromatin and recruit PRC2 to re-establish repression at night and into the early afternoon the next day. Conversely, the MADS-box proteins SHORT VEGETATIVE PHASE (SVP) and FLC occupy the CArG box, and Polycomb repressive complexes catalyze H3K27me3 deposition, repressing FT transcription. The key histone modifications are indicated on the right. Arrows denote positive effects; blunt lines denote repressive roles. Images were created by BioRender,
https://BioRender.com/6r3d71u .Beyond their canonical roles in flowering, these regulators integrate environmental cues, including temperature, photoperiod, vernalization, and gibberellin (GA), to control a wide range of developmental and physiological processes, such as seed dormancy, bud dormancy, leaf morphogenesis, branching architecture, and stress responses. Through these pleiotropic functions, flowering-time genes enable plants to adapt to seasonal changes, coordinate life-history strategies, and balance growth with survival under adverse conditions[5−7].
Importantly, the pleiotropic nature of these genes not only contributes to ecological adaptability, helping plants cope with complex environmental changes, but also holds substantial promise for agricultural improvement. Their influence on plant morphology, reproductive organ formation, and stress resistance precisely aligns with the core target traits for crop improvement[8,9]. Advances in genome editing technologies, such as CRISPR/Cas9 (clustered regularly interspaced short palindromic repeats/CRISPR-associated protein 9), now enable precise manipulation of these regulatory nodes[10,11], opening new avenues for breeding climate-resilient and high-yielding crops.
In this review, we synthesize the current knowledge on the floral integrators, FT and SOC1, and the repressors, FRI and FLC, with particular focus on their non-floral functions and potential applications in plant breeding. Other essential components of the flowering pathway, such as CO, also have increasingly recognized roles beyond flowering and have been comprehensively reviewed elsewhere[12]. Therefore, they are not a major focus of this article. We draw on studies of both model species, Arabidopsis (Arabidopsis thaliana), and major crops, including wheat, rice, maize, soybean, potato, and tomato, to provide a conceptual framework for understanding how conserved flowering-time regulators have been repurposed to control diverse physiological processes across species.
-
Genetic studies have established a well-defined regulatory network controlling the flowering time (Fig. 1a). The key components, including FRI, FLC, FT, SOC1 and CO, coordinate environmental and endogenous cues to precisely control the transition from vegetative to reproductive growth.
FRI promotes high FLC expression by associating with FLC chromatin through the FRI complex (FRIc), which includes FRIGIDA-LIKE 1 (FRL1), SUPPRESSOR OF FRIGIDA4 (SUF4), FRIGIDA-ESSENTIAL 1 (FES1), and FLC EXPRESSOR (FLX) (Fig. 1b)[13]. This complex establishes a transcriptionally active chromatin state at the FLC locus by facilitating the deposition of activating histone marks, including H3K4me3 and H3K36me3, and by promoting productive transcription and efficient RNA processing[14−16]. FRI-mediated activation of FLC is temperature dependent. During prolonged cold exposure, FRI is sequestered into nuclear condensates, a process promoted by the antisense transcript COOLAIR, leading to FLC repression. The rapid reversion of this process by warm temperatures prevents premature flowering and helps align reproductive development with favorable seasonal conditions, thus improving their survival and fitness[16,17]. In addition, FLC expression is supported by the histone H2B monoubiquitylation mediated by the E2 enzymes UBIQUITIN CARRIER PROTEIN 1 (UBC1) and UBC2, as well as the E3 ligases HISTONE MONOUBIQUITINATION 1 (HUB1) and HUB2, which enhances chromatin accessibility at the FLC locus (Fig. 1b)[18,19].
FLC encodes a MADS-box transcription factor that represses flowering by inhibiting the expression of key floral activators. In association with SHORT VEGETATIVE PHASE (SVP), FLOWERING LOCUS M (FLM), and MADS AFFECTING FLOWERING (MAF) proteins, FLC binds to CArG-box [CC(A/T)6GG] elements in the promoters of FT and SOC1, thereby delaying flowering (Fig. 1c)[1,3]. Vernalization counteracts this repression by inducing stable epigenetic silencing of FLC, allowing flowering to proceed after plants experience prolonged cold[20]. This process has been extensively studied, and we refer the readers to several comprehensive reviews that provide detailed mechanistic models[21−23].
FT functions as a mobile florigen that conveys photoperiodic information from leaves to the SAM[24−26]. FT expression is tightly regulated by day length and the circadian clock. Under short-day (SD) conditions, FT is transcriptionally silenced by Polycomb-mediated repression, whereas under inductive long-day (LD) conditions, FT is transiently activated around dusk[26,27]. This activation depends on the accumulation of CO in leaf vasculature cells and its binding to the FT promoter in cooperation with nuclear factor Y (NF-Y) transcription factors (Fig. 1c)[28]. In this context, CO serves as a key molecular link between circadian/light signals and FT transcription, a role that is broadly conserved across flowering plants. On the other hand, FT activation is rapidly attenuated during the night through the re-establishment of Polycomb-mediated chromatin repression, ensuring precise temporal control of FT expression[27,29−31] (Fig. 1c). Spatial specificity of FT expression is conferred by an unusually long promoter containing multiple regulatory elements that integrate photoperiodic and chromatin-based inputs[32−35].
The regulatory mechanisms governing FT are broadly conserved across flowering plants. In rice (Oryza sativa L.), multiple FT homologs have been identified, among which Heading date 3a (Hd3a) and Rice Flowering Locus T1 (RFT1) act as florigen genes under SD and LD conditions, respectively[36,37]. Divergence in the epigenetic regulation of these homologs enables fine-tuning of flowering responses under distinct photoperiodic environments[38].
SOC1 is an MIKCc-type MADS-box transcription factor that integrates signals from multiple flowering pathways downstream of FT and CO protein[2,39]. SOC1 is activated by the FT-FD-14-3-3 florigen activation complex under LD conditions, whereas it is repressed by FLC through direct binding to CArG-box elements in the SOC1 promoter[2]. Through interactions with other MADS-box proteins, including FRUITFULL (FUL), AGAMOUS-LIKE 24 (AGL24), and SVP, SOC1 contributes to the precise timing of the floral transition and the establishment of the annual life cycle[40].
Together, FRI, FLC, FT, and SOC1 constitute a conserved regulatory module that ensures flowering occurs at an appropriate time in response to environmental conditions (Fig. 1). In the following sections, we move beyond this canonical flowering pathway to examine how these regulators exert pleiotropic functions in diverse non-floral developmental processes and stress responses[41,42].
-
Seed dormancy is a critical adaptive trait that ensures germination occurs only under favorable environmental conditions. Beyond their well-established roles in the floral transition, FRI, FLC, and FT also exert pleiotropic influences on seed dormancy and germination (Fig. 2).
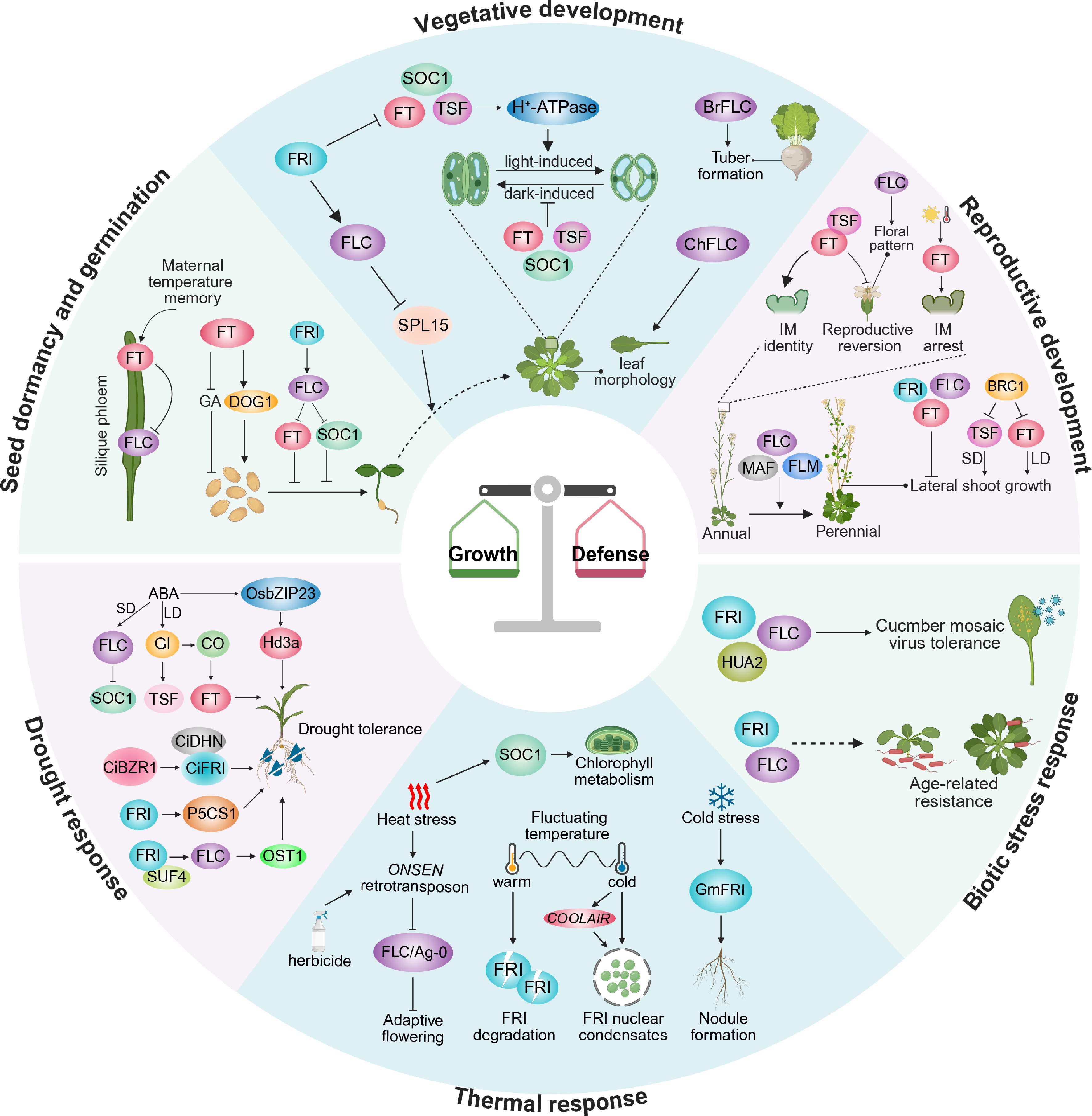
Figure 2.
Multifaceted roles of FRI, FLC, FT, and SOC1 in plant development and defense. The upper half focuses on their roles in growth and developmental transitions, and the lower half summarizes their regulation in response to abiotic environmental stresses and biotic challenges. Seed dormancy and germination: FT positively regulates seed dormancy by suppressing the GA pathway and activating the DELAY OF GERMINATION 1 (DOG1) pathway. Within the silique, FT mediates the control of offspring dormancy in response to maternal temperature signals through feedback inhibition of FLC. Conversely, FRI promotes the germination of non-vernalized maternal seeds by upregulating FLC. Vegetative development: FLC retards the transition from the juvenile to the adult phase by repressing SPL15 expression. FT, TSF, and SOC1 positively regulate stomatal opening. Furthermore, BrFLC correlates with tuber formation in turnips, whereas ChFLC regulates leaf size and complexity in Cardamine hirsuta. Reproductive development: FRI, FLC, and FT participate in regulating lateral shoot growth. BRC1 inhibits lateral shoot growth by repressing TSF or FT expression in a photoperiod-dependent manner. FT and TSF function redundantly to determine inflorescence meristem identity. Additionally, FT stabilizes the inflorescence and inhibits reproductive reversion, while also mediating the photo-thermal timing of inflorescence meristem (IM) arrest at the end of flowering. FLC, MAF and FLM are key factors maintaining perennial growth habit in perennial species. Drought response: FRI, FLC, FT enhance drought tolerance. In rice, the FT homolog Hd3a regulates drought escape, partially via an abscisic acid (ABA)-dependent pathway. Thermal response: SOC1 overexpression contributes to chlorophyll metabolism under heat stress. Heat shock activates the ONSEN retrotransposon located within the FLC intron 1, impairing FLC activity, and thereby promoting rapid adaptive flowering. Under fluctuating temperature conditions, warm destabilizes FRI, whereas cold promotes FRI nuclear condensation. In soybean, GmFRI positively regulates nodulation under cold stress. Biotic stress response: FRI, FLC and the flowering-time regulator HUA2 enhance plant tolerance to viral infection. Additionally, FRI and FLC promote age-related resistance (ARR) independently of the floral transition. Arrows denote positive effects; blunt lines denote repressive roles. Images were created by BioRender,
https://BioRender.com/cgkf6sf .Like flowering, germination represents an environmentally responsive developmental transition, and FLC plays a temperature and dormancy-dependent role in this process. Warmer conditions, such as 22 °C, enhance the regulatory impact of FLC on germination and dormancy[43,44]. However, the direction of this effect varies with the seed's primary dormancy status. Elevated FLC expression during seed maturation generally promotes dormancy[44], yet, under conditions of reduced primary dormancy, FLC can instead facilitate germination[45]. This bidirectional behavior reflects FLC's integration of environmental temperature cues with the physiological state of the seed. Mechanistically, FLC influences the key pathways controlling dormancy depth, including the balance between abscisic acid (ABA) and GA, as well as chromatin and epigenetic regulation[44−48]. Thus, understanding the environmental control of germination requires linking flowering-time regulators that sense temperature signals with hormonal and chromatin-based dormancy pathways. Collectively, these findings position FLC as a temperature-responsive hub whose developmental outcomes emerge from the interaction between external environments and internal dormancy status.
In Arabis alpina, the FLC homolog PERPETUAL FLOWERING 1 (PEP1) (also known as AaFLC) similarly regulates seed dormancy and longevity. pep1 mutants exhibit reduced dormancy and shorter seed longevity[49]. This conservation suggests that FLC-mediated regulation of germination and flowering may represent an evolutionarily conserved strategy for coordinating key life history transitions.
Given FLC's role in seed dormancy, its upregulation by FRI during seed maturation is crucial for fine-tuning seed dormancy responses[46,50]. The effects of FLC on germination depend on the presence of functional FRI: loss of FLC reduces germination only when FRI is active[45]. Functional FRI alleles typically maintain higher FLC expression, which enhances dormancy in highly dormant seeds but promotes germination when dormancy is low[45,46]. Conversely, plants with non-functional or weak FRI alleles show reduced FLC expression and earlier germination, which may confer adaptive advantages in milder climates with less stringent seasonal dormancy requirements[45,51]. The natural genetic variation on FRI thus contributes to population-level differences in FLC expression and seed behavior, forming an important axis of adaptation to diverse thermal environments. These context-dependent interactions between FRI and FLC therefore underpin population-level differences in germination strategies across thermal environments. Given the temperature-sensitive response of FRI during vernalization[16], it would be interesting to investigate whether cold-induced nuclear condensate formation contributes to seed dormancy regulation.
In addition to the zygotic function, FLC also exerts a maternal effect on progeny dormancy, mediating the influence of the maternal thermal environment during seed development[44,46]. This maternal influence underscores FLC's role as a transgenerational integrator of temperature signals, aligning offspring behavior with seasonal climatic patterns[52].
Also, FT plays a pivotal role in transgenerational dormancy memory by integrating maternal temperature cues in silique phloem, thereby aligning progeny behavior with seasonal cycles (Fig. 2). FT has a > 100-fold higher expression in siliques compared to leaves, where it processes maternal environmental signals to modulate dormancy[28,43]. Although FT mRNA is undetectable in seeds, FT-GFP fusion proteins translocate from silique phloem to accumulate at the chalazal pole of the seed coat, potentially through inter-tissue protein movement[43].
Parallel to FT, its ortholog MOTHER OF FT AND TFL1 (MFT) transduces oxylipin signals into dormancy responses[48]. In wheat (Triticum aestivum), MFT is expressed in the embryo and acts as a germination inhibitor, with its expression peaking during seed maturation under low temperatures. The natural variation at the MFT locus accounts for differences in seed dormancy among East Asian cultivars[53]. Loss-of-function mutations in MFT consistently reduce dormancy, confirming its repressive role. MFT functions within a conserved regulatory network involving ABSCISIC ACID INSENSITIVE 5 (ABI5), DELLAs, and hormone metabolism[48], though the mechanistic details, particularly in crops with divergent temperature responses, remain to be fully resolved. Its cross-species conservation positions MFT as a prime target for dormancy manipulation, suggesting that compartmentalized signaling cascades generate tissue-specific outcomes from shared genetic components.
Although FLC and FT coregulate flowering time, they exhibit functional divergence in regulating seed traits. FT modulates seed dormancy largely independent of FLC, indicating distinct regulatory pathways for flowering and germination[43,54]. A few studies indicate some feedback regulation from FT on FLC: FT suppresses FLC expression via a thermosensitive feedback loop through COOLAIR[44], and FT facilitates epigenetic silencing of FLC by influencing levels of H3K27me3 and of H3K36me3 at the FLC nucleation region in siliques and leaves[55]. These works indicate that an FT-FLC feedback axis integrates temperature cues to coordinately regulate both flowering time and seed dormancy. Moreover, components of both the autonomous and vernalization pathways influence seed dormancy through FLC-dependent and -independent mechanisms, confirming the shared genetic pathways between germination and flowering and providing insight into the genetic basis of adaptive life-history strategies.
-
FLC plays a crucial role in vegetative development by directly repressing SQUAMOSA PROMOTER-BINDING PROTEIN-LIKE 15 (SPL15), a key factor that promotes the juvenile-to-adult phase transition, thereby maintaining juvenile characteristics in plants (Fig. 2)[3,56]. In addition to this role, FLC has been reported to influence various aspects of vegetative growth independently of flowering regulation, including the timing and progression of the vegetative phase change and leaf morphology in Arabidopsis (Fig. 2)[3,56]. However, whether FLC plays a direct role in leaf shape and size remains unclear. Several studies propose that FLC may exert indirect effects on these traits through modulation of hormonal signalling and interactions with other MADS-box proteins, such as FLOWERING LOCUS M (FLM), MADS AFFECTING FLOWERINGs (MAFs), and SVP, particularly under temperature-sensitive conditions[57]. In contrast, in Cardamine hirsuta, an annual member of the Brassicaceae family, FLC has been shown to directly regulate leaf size and complexity. Accessions with reduced ChFLC expression exhibit early flowering and produce leaves with increased leaflet number (Fig. 2), indicating a close association between FLC expression levels, leaf morphology, and flowering time[58]. These observations suggest that while the role of FLC in developmental processes such as leaf patterning may vary across species, it is likely modulated by environmental cues, particularly temperature.
The florigen FT also significantly influences leaf development and overall plant architecture. Recent evidence indicates that the balance between indeterminate and determinate growth is governed by the activities of FT-like and TFL1-like genes, which profoundly affect floral transition and architectural patterning[59]. For instance, ectopic overexpression of GhFT1, a FT homolog from cotton (Gossypium hirsutum), in tobacco (Nicotiana tabacum) leads to altered leaf morphology, increased chlorophyll content, and enhanced shoot branching[60]. Similarly, the tomato (Solanum lycopersicum L.) FT homolog SFT regulates leaf development (Fig. 3a)[61], representing a significant evolutionary adaptation in flowering plants. In contrast, the maize FT ortholog Zea mays CENTRORADIALIS 8 (ZCN8) plays an additional role in restricting leaf growth and initiating axillary meristems (Fig. 3b)[41]. In Arabidopsis, FD functions as the central cofactor of FT in promoting flowering. Overexpression of its rice ortholog, OsFD2, results in smaller leaves and shortened plastochrons (the time interval between successive leaf primordia), indicating its involvement in leaf formation and phyllotactic timing[62−64]. This suggests that FT, like FLC, participates in complex regulatory networks that modulate plant growth and development, offering potential avenues for improving crop yield through architectural optimization.
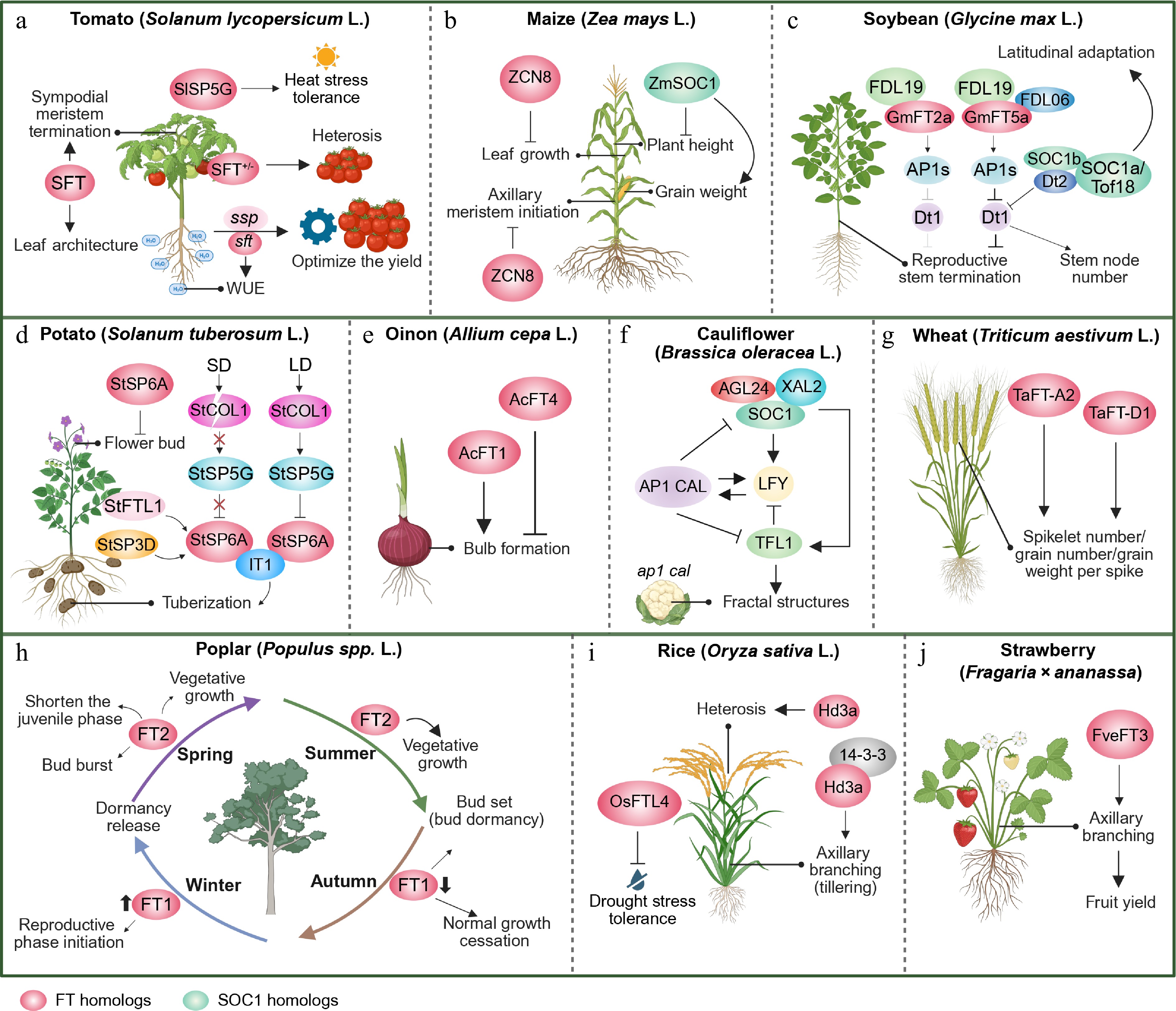
Figure 3.
Non-floral roles of FT and SOC1 family genes in major crops. (a) The tomato FT homologue SINGLE-FLOWER TRUSS (SFT) modulates sympodial meristem termination, leaf architecture, and water-use efficiency (WUE). SELF PRUNING 5G (SlSP5G) homologous to the TERMINAL FLOWER 1 (TFL1) promotes heat-stress tolerance; The combination of heterozygous sft and suppressor of sp (ssp) optimizes the fruit yield. (b) The maize FT homologue ZCN8 inhibits leaf growth and axillary meristem initiation, whereas the SOC1 homologue ZmSOC1 reduces plant height and enhances grain weight. (c) The soybean FT homologue GmFT5a rather than GmFT2a promotes stem termination through a specific interaction with FDL06 to inhibit Dt1-mediated reproductive stem growth. Tof18/SOC1a enhances latitudinal adaptation, and the SOC1s-Dt2 complex regulates stem node number by inhibiting Dt1. (d) Under long days (LD), CONSTANS-LIKE1 (StCOL1) activates the repressor StSP5G, which inhibits FT homologue StSP6A expression in potato; Under short days (SD), StSP6A triggers tuberization when the repressor is destabilized. Additionally, the florigen SELF PRUNING 3D (StSP3D) and FLOWERING LOCUS T-like 1 (StFTL1) are the novel tuberization signals. StSP6A also inhibits flowering buds. (e) Two onion FT homologs coordinate bulb formation with opposing functions: AcFT4 suppresses the formation of bulbs, while AcFT1 promotes it. (f) In cauliflower, the SOC1-centered floral gene network drives curd morphogenesis: loss of AP1 and CAL disrupts LFY expression maintenance and derepresses SOC1/AGL24, which induces ectopic TFL1 expression in floral primordia. TFL1 then further suppresses LFY, causing the primordia to lose floral identity, revert to inflorescence meristems, and ultimately form the characteristic cauliflower curds. (g) Two wheat FT homologs, TaFT-A2 and TaFT-D1, enhance spikelet number, grain number, and grain weight per spike, directly contributing to yield. (h) Two FT homologs act seasonally to coordinate poplar’s perennial growth cycle: FT2 promotes spring/summer vegetative growth, shortens the juvenile phase, and drives bud burst and bud set, whereas FT1 mediates autumn growth cessation, winter dormancy release, and the transition to reproduction. (i) The rice FT homolog Hd3a drives heterosis and regulates tillering (axillary branching) via 14-3-3 interaction, while OsFTL4 confers drought stress tolerance. (j) In strawberry, the FT homolog FveFT3 positively controls axillary branching, thereby increasing fruit yield. FT homologues are shown as pink circles and SOC1 homologues as light-green circles. Solid arrows indicate promotion/activation, while flat-ended (truncated) arrows denote repression/inhibition. Images were created by BioRender,
https://BioRender.com/xr70vik .Similarly, the flowering pathway integrator SOC1 coordinates developmental transitions at the shoot apex through multilayered regulatory interactions. Recent studies have demonstrated that SOC1 cooperates with FUL to activate the SPL9 and SPL15 transcription factors, establishing a regulatory module that integrates photoperiod, GA signaling, and endogenous age cues[65−67]. In addition to its role in development, SOC1 acts as a key repressor of leaf senescence. Loss-of-function mutations in SOC1 accelerate leaf yellowing in Arabidopsis, whereas transgenic overexpression lines exhibit delayed senescence[68]. This anti-senescence function is mediated by the direct binding of SOC1 to the CArG box in the promoter of PHEOPHYTINASE (PPH), a critical enzyme in chlorophyll degradation, thereby suppressing its expression and preserving photosynthetic capacity.
Stomatal regulation
-
A new cell-autonomous function for FT has been identified, independent of its mobile form, wherein FT is specifically expressed in guard cells and regulates stomatal aperture by activating the plasma membrane H+-ATPase[69]. Similarly, TSF is expressed in guard cells and mediates light-induced stomatal opening[70]. Additionally, TSF negatively regulates water-use efficiency (WUE) in tomato through an ABA-independent mechanism that involves alterations in stomatal conductance and leaf anatomy (Fig. 2)[71]. These findings indicate that FT and TSF play direct roles in gas exchange and WUE, thereby extending their regulatory functions beyond flowering to include stomatal regulation and environmental adaptation.
FLC expression levels in guard cells are negatively correlated with those of FT, TSF, and SOC1. Plants carrying an active FRI allele exhibit reduced light-induced stomatal opening, coinciding with elevated FLC expression and decreased expression of FT, TSF, and SOC1 in guard cells (Fig. 2)[72,73]. Moreover, FT, TSF, and SOC1 negatively regulate the dark-induced stomatal closure mediated by CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1)[73].
Stem elongation and termination
-
Stems are integral components of the plant shoot system, influencing the development of reproductive structures and branching patterns. Several genes involved in flowering regulation have been shown to concurrently affect stem development, primarily by regulating floral transition and reproductive organ formation. Notably, the FT homolog StSP6A functions as a key tuberigen that promotes tuber (underground stem) development in potato (Solanum tuberosum L.)[74], while FT and SOC1 homologs have been clearly implicated in regulating aboveground stem growth in various species[41,75].
The systemic growth-regulatory function of FT is evolutionarily conserved. In temperate cereals, stem elongation is closely linked to plant height. Functional diversification of FT-like genes enables fine-scale regulation of stem development and fertility beyond flowering time control. In wheat (Triticum aestivum), the FT paralog FT1 primarily modulates the pace and duration of stem elongation, with RNA interference (RNAi) mutants higher than wildtype which is likely caused by the delayed heading date[76]. In barley (Hordeum vulgare), FT-like genes have diversified into functionally antagonistic regulators, exemplified by HvFT3, which promotes spikelet initiation[77], and HvFT4, which represses reproductive development and floret fertility[78]. In tomato, the FT ortholog SFT acts not only as a mobile floral inducer but also as a general systemic growth regulator. Ectopic overexpression of this gene (35S:SFT) results in reduced leaf complexity, shorter internodes, thinner stems, and apical meristem arrest (Fig. 3a)[61,79]. Together, these studies indicate that FT-like genes play diverse roles in coordinating plant height and reproductive development.
In contrast, heterozygosity for loss-of-function sft alleles leads to heterosis in tomato, resulting in a yield increase of up to 60%[80], likely through modulation of meristem activity and growth termination[81]. This role of FT homologs in regulating stem growth termination is also evident in soybean (Glycine max). Among the two FT homologs, GmFT5a rather than GmFT2a plays a central role in terminating reproductive stem growth, by interacting with FD-LIKE 19 (FDL19) and FDL06 to induce high-level expression of APETALA1 (AP1) homologous genes (Fig. 3c)[75]. This regulatory cascade suppresses Dt1, a key gene promoting indeterminate stem growth, thereby facilitating stem termination. In parallel, SOC1 homologs also play crucial roles in fine-tuning stem development in crops. Studies in soybean have shown that Tof18/SOC1a enhances latitudinal adaptation, and the SOC1s-Dt2 complex directly binds to the Dt1 promoter to regulate stem node number (Fig. 3c)[10]. Importantly, this mechanism complements the FT-mediated pathway: whereas FT homologs such as GmFT5a indirectly inhibit Dt1 via AP1 homologs, SOC1 homologs directly target Dt1, forming a synergistic regulatory network that governs stem termination. The introgression of the natural Tof18A allele into modern soybean cultivars may therefore offer a promising strategy for enhancing the yield[10].
Storage organ development
-
Many plants have evolved specialized organs to survive adverse conditions, such as underground rhizomes and stem structures that enable them to withstand severe cold. The FLC and FT gene families play important roles in the formation of underground storage organs, including tubers and bulbs.
In turnips (Brassica campestris L. ssp. rapifera Matag syn. B. rapa L.), BrFLC1 expression in the hypocotyl correlates with tuber formation, and cold vernalization treatment decreases BrFLC1 transcript levels, which is associated with repression of tuberization (Fig. 2)[82]. A hypocotyl micrografting system showed that late-flowering turnip rootstocks delayed flowering in both early-flowering turnip and rapeseed scions. This delay was linked to elevated BrFLC1 transcript levels and H3K4me3 enrichment in scion leaves, together with reduced FT expression[83]. Conversely, grafting onto vernalized rootstocks promoted early flowering in non-vernalized scions with a corresponding increase in FT homolog expression. Collectively, these findings suggest that BrFLC1 coordinates flowering and tuberization in turnip, possibly through mobile signals modulating FT pathways, and highlight how FLC expression levels influence environmental adaptation and yield potential.
In contrast to the inhibitory effects of FLC, the FT gene family generally promotes storage organ formation, and this function is conserved across multiple organ types. The potato serves as a classical model of underground storage organ formation where the FT homolog StSP6A functions as a key tuberigen. Similar to FT in Arabidopsis, StSP6A functions as a phloem-mobile signal integrating environmental cues, particularly photoperiod, to initiate tuber formation through the tuberigen activation complex (TAC) (Fig. 3d)[8]. In potato, photoreceptor PHYTOCHROME B (StPHYB) stabilizes CONSTANS-LIKE1 (StCOL1) to activate the repressor StSP5G, which inhibits StSP6A expression under LD conditions[9]. Under SDs, StCOL1 is destabilized, repression is released, and StSP6A is activated[9]. Additionally, the florigen SELF PRUNING 3D (StSP3D) and FLOWERING LOCUS T-like 1 (StFTL1) were identified as novel long-range signals that act as tuber organogenesis stimuli, and StSP3D and StFTL1 are responsible for the secondary activation of StSP6A in stolon tips to amplify the tuberigen signal[74]. Intriguingly, StSP6A also represses floral bud development[84], thereby redirecting assimilates such as sucrose toward tuber enlargement. This dual role highlights the evolutionary repurposing of FT-like proteins in balancing reproductive and vegetative growth, with direct consequences for yield.
Similar regulatory patterns are observed in above-ground storage organs. In biennial onion (Allium cepa), distinct FT homologs control flowering and bulb formation: AcFT1 promotes bulbing, whereas AcFT4 acts as an inhibitor (Fig. 3e)[85]. Thus, onions and potatoes exemplify how FT family members have diverged to regulate both above- and below-ground storage organs, further emphasizing the widespread role of FT in storage organ development. In orchids, FT-like proteins are implicated in pseudobulb formation[86,87], providing additional evidence for the broad functional diversification of this gene family.
FT-like genes have also been identified in root crops such as sweet potato and cassava (Manihot esculenta)[88,89]. In cassava, MeFT1 is expressed in leaves without a strong photoperiod response, while MeFT2 shows photoperiod-dependent expression. Overexpression of MeFT1 induces early flowering but reduces storage root formation in grafting studies, suggesting a preferential allocation of sucrose to reproductive organs[89]. However, most of these results are based on gene expression correlations or indirect phenotypic evidence, and direct functional validation of FT/FT-like genes in root crop storage organ formation through gene editing or transgenesis is still lacking.
Potato, nevertheless, remains the best-studied system, not only because of its well-characterized StSP6A mechanism but also due to its hybrid origin, which provides an evolutionary context for functional diversification[90]. Comparative analyses have shown that tuberization-related genes often derive from hybridization events. For example, StSP6A and StGIGANTEA (GI) were inherited from tomato, whereas PHYB and certain epigenetic regulators originated from wild potato relatives such as S. etuberosum[90]. The tomato-derived SP6A homolog may primarily function in floral regulation, but it acts together with wild-derived PHYB to acquire a novel role in promoting tuberization in potato. This mosaic inheritance pattern reflects the hybrid origin of cultivated potato and illustrates how interspecific hybridization can generate new gene combinations that drive trait innovation.
-
One of the first steps in flowering initiation is the conversion of SAM into either an inflorescence meristem (IM) or an FM. In certain plant species, SAM directly differentiates into an FM without generating lateral IMs. In many others, IM produces FMs on its flanks while maintaining its own identity, resulting in an indeterminate inflorescence. The precise specification of meristem identity, along with the establishment of correct floral organ patterns and the coordinated development of the inflorescence structure, constitutes a fundamental aspect of reproductive success in flowering plants, significantly influencing the crop yield.
In Arabidopsis, FT and its close homolog TSF function redundantly to determine and maintain IM identity (Fig. 2)[91]. This is clearly demonstrated by the ft-10 mutant phenotype, where the inflorescence apex reverts to vegetative growth, producing rosette-like leaves[92]. This phenotype is markedly enhanced in ft-10 tsf-1 double mutants[92−94], highlighting the essential role of FT and TSF in sustaining reproductive fate after flowering. Additionally, FT promotes the photo-thermal timing of IM arrest at the end of the reproductive phase[95]. This role of FT in meristem termination is evolutionarily conserved.
In winter wheat, flower bud formation, commonly referred to as spike development, occurs in day-neutral conditions around the spring equinox. Under natural photoperiods, spike development is governed by a stepwise increase in FT1 expression. Inflorescence initiation is triggered under day lengths exceeding 11.5 h, which induces a modest rise in FT1 transcript levels. Longer photoperiods promote advanced developmental stages by further elevating FT1 expression and activating a second wheat FT orthologue, FT2. These findings reveal the presence of a photoperiod-responsive developmental checkpoint during floral development in cereals, functionally analogous to the mechanism by which long days alleviate ecodormancy during bud break in perennial species[96]. On the other hand, a natural variant of the A-genome copy of FT2 (FT-A2), named as FT-A2 A10 allele, has been recently associated with increases in spikelet number per spike (SNS), grain number per spike, and grain weight per spike (Fig. 3g)[97]. Further study found that the interaction of basic leucine zipper transcription factor C1 with FT2 contributes to the regulation of SNS[98]. A recent work found that TaFT-D1 increased grain weight by promoting cell proliferation and starch synthesis, and the TaFT-D1(G) allele correlates with a greater grain weight and earlier heading[99].
Floral organ formation is governed by the 'ABC' model of floral organ identity, which involves several MADS-box transcription factors, including AP1, AP3, AGAMOUS (AG), and SEPALLATA3 (SEP3) (Fig. 1). Although the complex regulatory interactions among MADS-box genes throughout the flowering pathway remain incompletely understood, FLC has been shown to repress the expression of floral patterning genes, such as SEP3, a central regulator of floral organ identity[100]. In turn, SEP3 activates the expression of class B and C genes, the core components of the 'ABC' model that specify petal and stamen identities, and stamen and carpel identities, respectively[100]. This regulatory hierarchy is essential for proper floral patterning, as ectopic expression of SEP3 in Arabidopsis leads to abnormal inflorescence development[3,101].
Following the initial formation of the floral structure, the specification of FM identity becomes a critical prerequisite for the normal development of floral organs, a process tightly regulated by SOC1[2]. Loss-of-function mutations in SOC1 result in delayed flowering and defects in floral organ development, including abnormal petal and stamen formation[39]. Recent studies in Chinese walnut (Juglans cathayensis) have identified a distinctive in-frame deletion in the SOC1 gene that may contribute to early maturation differences between J. cathayensis and other Juglans species, offering the potential for leveraging SOC1 in crop improvement and genetic resource utilization[102].
In addition to these functions, SOC1 also contributes to morphological formation. In cauliflower (Brassica oleracea var. botrytis), studies on the molecular mechanisms underlying the formation of its helical, fractal structures have revealed that an SOC1-centered floral gene network plays a pivotal role in determining curd architecture (Fig. 3f)[103]. The loss of AP1 and CAULIFLOWER (CAL) abolishes the AP1/CAL-mediated positive-feedback loop, preventing the maintenance of LEAFY (LFY) expression. Meanwhile, the loss of AP1 and CAL relieves repression on SOC1 and AGL24, resulting in ectopic TFL1 expression in floral primordia. Thus, TFL1 further suppresses LFY, causing the primordia to lose their floral identity and revert to inflorescence meristems, ultimately giving rise to cauliflower-like curds[103].
In summary, FLC regulates the floral patterning genes to establish the molecular foundation for floral development. FT primarily governs the maintenance and timely termination of inflorescence meristem identity, thereby stabilizing overall inflorescence architecture. SOC1 acts at the level of individual FMs, linking flowering time with floral organogenesis. Together, these three factors form a coordinated regulatory network that orchestrates reproductive development across multiple scales: from global inflorescence patterning to the precise development of individual flowers, ensuring the structural fidelity and reproductive integrity of flowering plants.
Annual−perennial life histories
-
An active SAM continuously produces leaves and sustains vegetative growth in plants. In annual species, the SAM transitions irreversibly to an FM or IM, followed by senescence and plant death. In contrast, perennial plants undergo repeated cycles between vegetative and reproductive phases. Vegetative growth in perennials can be maintained either by retaining some meristems in a vegetative state after flower initiation or by reverting to vegetative development after flowering.
In perennial Brassicaceae, orthologues of FLC are repressed by prolonged winter cold and subsequently reactivated in spring, thereby conferring seasonal flowering patterns, while in annuals, FLC is stably silenced by vernalization (Fig. 2)[104,105]. Zhai et al. demonstrated that the expression level of FLC-like MADS-box genes governs the reciprocal conversion between annual and polycarpic perennial flowering behavior within the Brassicaceae[106]. Knockout of FLC homologs in perennial species resulted in loss of typical perennial traits, leading to an annual-like flowering behavior. Conversely, overexpression of FLC homologs in annual species induced perennial characteristics, including prolonged vegetative growth and repeated flowering cycles[106]. This functional evidence highlights the critical, dosage-dependent role of FLC in determining life history strategies. It provides direct genetic support for FLC's role in maintaining vegetative growth and explains the association between spring reactivation of FLC and the perennial habit.
Stable repression of FLC in annuals is mediated by both trans-acting regulators and cis-regulatory elements, such as the vernalization response element (VRE). However, it remains unclear whether perennial plants are similarly controlled by cis-regulatory variation. Kiefer and colleagues identified two regulatory regions, located at the 5'-end of the VRE and at the 5'-end of FLC intron 1, that may contribute to the divergence between annual and perennial species. One of these elements is hypothesized to enable reactivation of FLC orthologs after vernalization. The annual flowering pattern may have evolved multiple times independently through recurrent loss of this perennial-specific regulatory element[107]. Future studies should elucidate the molecular mechanisms underlying FLC reactivation in perennials and examine how FLC regulatory networks have co-evolved with shifts in life history strategies.
Bud dormancy
-
Bud dormancy, a temporary suspension of meristematic activity, is a key adaptive strategy in many perennial plants. In these species, floral initiation typically occurs during spring or summer, resulting in the formation of dormant flowers above- or below-ground that remain quiescent until the following spring. A study on winter oilseed rape (Brassica napus) demonstrated that vernalized plants undergoing floral initiation exhibit delayed bolting and flowering when exposed to warm SD conditions, compared to those receiving chilling treatments[108]. These findings indicate that the combination of SDs and insufficient winter chilling induce flower bud dormancy in B. napus. This provides compelling evidence that bolting and flowering in annual species can be temporally uncoupled, suggesting a broader role for flower bud dormancy in regulating flowering time across annual plants[96].
In winter oilseed rape, dormant flower bud formation occurs in late autumn under SD conditions, where floral integrator FT activity is low. This process requires GA to promote the expression of both LFY and SOC1 at the shoot apex. Subsequent warming increases ABA levels in flower buds and upregulates genes associated with ABA-mediated repression of axillary bud outgrowth, as characterized in Arabidopsis. During dormancy, expression of FLC is elevated; its downregulation coincides with dormancy release, indicating that FLC, in concert with ABA signalling, suppresses meristematic activity during dormancy[109−111]. FLC thus plays a critical role in regulating meristem fate, shaping plant architecture in response to seasonal cues. As vernalization proceeds, axillary meristems destined for reproductive development remain in a dormant state, while others retain vegetative potential. This regulatory mechanism allows the plant to sustain vegetative growth and produce new branches even after flowering has commenced[112].
In woody perennial plants, which have extended growth cycles and must endure winter dormancy, FT paralogs have undergone substantial functional divergence to accommodate the annual rhythm of growth and dormancy. In Populus species, FT paralogues have acquired specialized roles: FT1 promotes the release from winter dormancy[113], whereas FT2 supports vegetative growth during the growing season (Fig. 3h)[42]. In Populus trichocarpa, the CO-FT regulatory module also governs SD-induced growth cessation and bud set in autumn, processes that require the downregulation of PtFT1[114]. Prolonged chilling during bud dormancy can activate FT independently of CO, thereby triggering bud burst in Populus[115]. In perennial Norway spruce, PaFT4 serves as a key integrator of growth rhythm control, including bud set and bud burst[116]. Notably, across diverse woody perennials and Arabidopsis, elevated FT expression is consistently linked to the suppression of bud dormancy, underscoring its conserved role in promoting a low-dormancy state.
Collectively, bud dormancy represents a widespread adaptive strategy employed by both perennial and annual plants to survive in seasonal environments. It allows plants to withstand adverse conditions and ensures the coordinated resumption of growth, a process largely regulated by flowering-time genes that also control the initiation of flowering. A deeper understanding of these regulatory mechanisms is crucial for predicting the impacts of climate change on agricultural productivity and natural ecosystems.
Branching
-
Branching architecture, determined by the number, position, and developmental fate of lateral branches, shapes plant morphology and influences reproductive output in both annual and perennial species. Although traditionally viewed as developmental traits distinct from flowering time, emerging evidence indicates that core flowering-time regulators also contribute to the control of branching patterns across diverse plant lineages.
In annual species such as Arabidopsis, genetic studies have revealed specific interactions between flowering-time genes and branching traits. Quantitative trait locus (QTL) analysis has identified FRI (REDUCED STEM BRANCHING7, RSB7) and FLC (RSB6) as epistatically interacting loci that suppress stem branching. Furthermore, FT (RSB8) exerts pleiotropic effects, both modulating the flowering time and reducing stem branching, particularly in late-flowering genotypes with active FRI and FLC. This demonstrates context-dependent roles for canonical flowering genes in shaping shoot architecture[117]. In A. alpina, loss-of-function flc mutants exhibit abnormal meristem behavior, as all axillary meristems transition directly to reproductive development, thereby compromising the maintenance of vegetative growth after flowering[112] and resulting in a disrupted branching pattern[112,118]. In cereal crops such as rice, FT homologs primarily regulate inflorescence and branch number. The florigen Hd3a, which shares approximately 70% amino acid sequence identity with FT[63,64], promotes lateral bud outgrowth and branching via interaction with 14-3-3 proteins (Fig. 3i)[64]. This regulatory function operates independently of OsFD1[64], indicating that Hd3a can engage alternative signaling pathways to influence plant architecture. Similarly, in the SD species strawberry, FveFT3 is not a florigen but promotes plant branching by changing the axillary meristem fate when it is overexpressed, resulting in a 3.5-fold increase in fruit yield (Fig. 3j)[119].
In contrast, the role of SOC1 in regulating branching exhibits species-specific variation, particularly in polyploid and Solanaceous crops. In the allotetraploid Brassica juncea, SOC1 regulation is closely linked to branching patterns and agronomic traits[120]. Suppression of BjuSOC1 genes in transgenic lines results in reduced lateral branch numbers and delayed flowering, ultimately decreasing the silique number and total seed yield. Conversely, enhanced expression of BjuSOC1 in Arabidopsis offers a promising strategy for optimizing branch architecture and improving yield potential. In tomato, SOC1 homologs regulate inflorescence branching through interactions with other transcription factors[121]. Among four dynamically expressed SOC1 homologs identified in Solanaceae, TOMATO MADS 3 (TM3) and SISTER OF TM3 (STM3) interact with FUL homologs (FUL2) to co-regulate downstream targets. Notably, although TM3/STM3 and FUL2 synergistically promote flowering, they exhibit antagonistic interactions during inflorescence development[121]. In addition, SOC1 acts redundantly with other MADS-box transcription factors, including SVP, AGL24, and SEP4, to regulate inflorescence architecture by controlling meristem identity and branch patterning in both Arabidopsis and rice[122]. These observations highlight the conserved yet flexible role of SOC1 homologs in maintaining inflorescence branching across species.
-
Plants are constantly challenged by diverse environmental stresses. This section explores the pivotal roles of FRI, FLC, FT, and SOC1 in mediating plant responses to both abiotic and biotic factors. Acting as integrators of environmental signals, these genes coordinate developmental programs and stress adaptation, ensuring plant survival under fluctuating conditions.
Temperature stress
-
Temperature stress represents a major abiotic challenge that influences plant distribution and development, encompassing three primary conditions: extreme cold, extreme heat, and temperature fluctuations. FLC, FRI, FT, and SOC1 exhibit functional specialization in thermal adaptation.
FRI and FLC are key regulators of vernalization response, modulating the requirement for prolonged cold exposure to induce flowering. Low-temperature stress significantly affects plant geographic distribution, yet the ecological significance of FRI-mediated vernalization requirements in Arabidopsis accessions varies with latitude[17]. In soybean, GmFRI positively regulates root hair deformation and nodulation under cold stress through the nodulation factor signaling pathway (Fig. 2)[123], suggesting a role for FRI in cold acclimation. Further investigation into the roles of FRI and FLC in cold adaptation, particularly in relation to their function in promoting a winter-annual life history, could reveal the mechanisms by which plants achieve long-term protection under cold conditions.
High temperatures disrupt photosynthetic efficiency and metabolic homeostasis. In response, members of the FT gene family and SOC1 act in a complementary manner to enhance stress resilience through both damage mitigation and growth maintenance. Recent evidence indicates that SlSP5G, a tomato FT-like gene, enhances thermotolerance by reducing leaf damage via reactive oxygen species (ROS) scavenging and activation of antioxidant systems[124]. Overexpression of SOC1 or SOC1-like genes contributes to chlorophyll metabolism under heat stress, thereby enhancing photosynthesis[125].
Compared to extreme temperatures, temperature fluctuations have a broader impact on plant development. Both FLC and its antisense transcript COOLAIR exhibit transcriptional plasticity in response to fluctuating temperatures during vernalization. Notably, field experiments in Sweden revealed that COOLAIR is strongly induced by the first freezing event, functioning as a thermal indicator that enables FLC to monitor seasonal progression[126]. This induction leads to a stronger FLC repression than observed under constant low temperatures (Fig. 2). FLC's temperature-dependent expression also involves the thermosensitive protein FRI, which forms nuclear condensates in response to cold, facilitating FLC downregulation, a process rapidly reversed upon warming[16,127]. Moreover, research shows that temperature sensing within the FLC regulatory network is distributed across multiple components, including COOLAIR, FRI condensates, and chromatin regulators, allowing plants to register transient warm spells and buffer premature vernalization (Fig. 2)[20,128,129]. This transcriptional plasticity is critical during early autumn vernalization, preventing premature flowering. Extensive nucleotide diversity has been documented at the FRI locus across Arabidopsis accessions, resulting in truncated or mutated proteins[130]. However, it remains unclear whether these amino acid variations correlate with altered thermo-sensitivity.
FLC dynamically maintains 'temperature memory' through epigenetic modifications, ensuring flowering time is aligned with seasonal cues. However, this memory can be disrupted by heat stress. When vernalized seedlings are exposed to 30 °C, the repressive histone mark H3K27me3 is reduced at the FLC locus, leading to a late-flowering phenotype, unless the seedlings first experience a stabilizing period at 20 °C[131]. Recent studies have identified small molecules termed devernalizing agents that can reactivate silenced FLC alleles following vernalization, particularly when combined with heat treatment[132]. Given the ongoing climate change and the increasing frequency of extreme weather events, further research is needed to understand the impact of temperature fluctuations and the interactions between extreme climatic conditions on plant developmental and stress response pathways.
Drought stress
-
In Arabidopsis, WUE is positively correlated with flowering time. Late flowering in accessions with putatively functional FRI is associated with reduced precipitation in January at the site of origin. FRI overexpression enhances drought tolerance by upregulating DELTA1-PYRROLINE-5-CARBOXYLATE SYNTHASE 1 (P5CS1) expression and thereby promoting proline accumulation[5,133] (Fig. 2). In citrus, FRI is involved in direct protein interactions, where it partners with the dehydration-associated protein dehydrin (DHN) and the transcription factor BRASSINAZOLE-RESISTANT 1 (BZR1) to collectively enhance drought tolerance by upregulating stress-responsive and hormone biosynthesis genes[134] (Fig. 2). Furthermore, winter annuals with a functional FRI allele exhibit enhanced drought tolerance by reducing stomatal aperture and water loss, a response mediated by an FLC-OPEN STOMATA 1 (OST1) regulatory module crucial for adapting to seasonal transitions[6,72] (Fig. 2).
SOC1 is a direct executor of the drought escape response. In Arabidopsis, the ABA-responsive element (ABRE)-binding factors ABSCISIC ACID INSENSITIVE 3 (ABI3)-BINDING FACTOR 3 (ABF3) and ABF4 regulate this process by forming a transcriptional complex with NF-YC, which directly binds to the SOC1 promoter and activates its expression[135] (Fig. 2). Moreover, it was reported that the loss of OsFTL4 enhances drought tolerance by decreasing stomatal conductance and water loss[136]. In contrast, Hd3a, a key floral promoter in rice, plays a positive regulatory role in the drought escape process (Fig. 2). Under low water-deficit treatment conditions (LWT), the ABA signaling pathway upregulates Hd3a expression. Notably, in ABA-hypersensitive OsbZIP23-overexpressing plants, the transcript level of Hd3a is significantly enhanced, thereby accelerating the floral transition and heading time, allowing rice to complete its reproductive development before severe stress occurs. Quantitative reverse transcription polymerase chain reaction analysis further reveals that the drought-induced upregulation of Hd3a is markedly suppressed in ABA-deficient mutants such as photosensitivity 3-1 (phs3-1) and OsPDS-RNAi (RNA interference targeting PHYTOENE DESATURASE), leading to delayed flowering[137]. These results strongly support that Hd3a acts as a central executor in the ABA-dependent drought escape pathway.
Salt stress
-
Soil salinization is another common type of abiotic stress, and FLC has been preliminarily linked to plant responses to this stress. Ma et al. (2015) found that Arabidopsis CYCLIN-DEPENDENT PROTEIN KINASE G2 (CDKG2) acts as a negative regulator of salt tolerance: its loss enhances salt tolerance by up-regulating stress-responsive genes like SALT OVERLY SENSITIVE 1 (SOS1), SOS3, and P5CS1, while also accelerating flowering[138]. Notably, this dual effect of CDKG2 is mediated through FLC. CDKG2 promotes FLC expression, while reduced FLC levels (in cdkg2 mutants) not only drive early flowering but also align with the enhanced salt tolerance phenotype[138]. This indicates FLC's involvement in the crosstalk between salt stress response and flowering regulation, though further studies are needed to clarify FLC's specific molecular role in modulating salt stress adaptation.
Chemical stress
-
Another notable example of FLC's versatility in adapting to abiotic stress is its involvement in herbicide resistance, a common form of chemical stress. A recent study showed that in Arabidopsis natural accession Ag-0, a heat shock inducible ONSEN retrotransposon insertion (a heritable genomic variation that affects gene function by altering gene expression or structure) in FLC's first intron facilitates an adaptive response to the herbicide isoxaben (Fig. 2). This modification alters FLC expression, enhancing the plant's ability to resist cell wall-targeting agents and demonstrating FLC responds to chemical stressors[139]. Currently, only the retrotransposon insertion in the first intron of FLC has been found to enhance isoxaben resistance. However, the specific mechanism by which this variation alters FLC's expression pattern (e.g., promoter binding sites, transcript splicing) and its interaction with proteins such as SVP remain to be further investigated.
Biotic stress
-
Based on the response to abiotic stress, the functions of FLC and FRI have further expanded into the field of biotic stress, constructing an integrated regulatory network for 'development defence' in plants.
FRI exhibits pleiotropic effects in plant stress defense, but its core mechanism centers on regulating age-related resistance (ARR) and mediating the growth−defence trade-off. FRI primarily functions by regulating ARR, a process dependent on the salicylic acid (SA) signaling pathway (Fig. 2). FRI promotes the maturation of the plant immune system, which is independent of the floral transition[7]. However, it still needs further investigation whether FRI confers ARR competence through the SA signalling pathway or FRI generally regulates immune responses in a broader species regime.
Recent studies on plant virus tolerance mechanisms have further revealed that FLC and FRI do not act alone in biotic stress responses; instead, they synergize with other flowering-time regulators (e.g. HUA2) to enhance plant tolerance to viral infections. For example, one recent study has explored the genetic basis of Arabidopsis's tolerance to cucumber mosaic virus (CMV), highlighting the collaborative role of these flowering repressors (Fig. 2)[7,140]. Using a recombinant inbred line (RIL), the study identified three major QTLs for tolerance, which co-locate with these flowering repressor genes. Functional alleles of FLC, together with FRI and/or HUA2, were found to be necessary for both CMV tolerance and resource reallocation from vegetative growth to reproduction[140]. Interestingly, FLC alleles from wild accessions modulated tolerance differently, depending on their effects on the flowering time[140]. These findings reveal that FLC plays a novel role in plant defense, suggesting that flowering-time regulators can influence both tolerance and developmental processes through distinct mechanisms. This discovery opens new avenues for studying the balance between plant defense and developmental regulation.
-
The expanding functional repertoire of flowering-time regulators such as FRI, FLC, FT, and SOC1 underscores their significance as integrators of developmental and environmental pathways. While these genes were originally characterized for their roles in floral transition, accumulating evidence highlights their involvement in processes ranging from branching and storage organ formation to abiotic and biotic stress responses. These findings illustrate that the flowering genes function as versatile hubs shaping plant architecture, developmental plasticity, and environmental resilience. Thus, understanding these non-floral roles is essential for elucidating how global growth and developmental networks are coordinated, and it provides promising avenues for targeted crop improvement.
Despite these advances, several critical gaps still remain. Most functional evidence comes from Arabidopsis and a few model crops, leaving the conservation of non-floral functions across diverse plant lineages largely unexplored, and the molecular mechanisms by which flowering genes mediate stress adaptation, hormonal crosstalk, or chromatin-based regulation remain poorly characterized. Most studies rely on phenotypic observations, leaving the underlying mechanisms and their evolutionary conservation unclear. From a practical perspective, traits directly related to architecture and yield may warrant higher priority for functional validation due to their immediate agricultural relevance. Furthermore, some reported non-floral effects differ between species or experimental contexts, emphasizing the need to resolve potential inconsistencies and to identify species-specific versus conserved functions. Distinguishing direct regulatory effects from indirect pleiotropic consequences remains a major challenge.
Methodological limitations further restrict our current understanding. Conventional genetic approaches such as knockouts or overexpression may produce indirect effects, and analyses often lack tissue- or cell-type-specific resolution. Whole-organ transcriptomic studies can obscure subtle regulatory patterns within specific cell populations. Addressing these limitations will require spatially resolved, multi-omics strategies, including single-cell transcriptomics, epigenomics, and hormone profiling.
To advance the field, we propose several concrete directions for future research: (1) tissue- and cell-type-specific manipulations of flowering genes, coupled with gene expression titration and transcriptomic, epigenomic, and hormone profiling, to distinguish direct from indirect effects; (2) comparative analysis across species to define conserved versus species-specific non-floral roles, revealing evolutionary trajectories; and (3) integration of multi-omics and single-cell approaches to resolve the spatiotemporal dynamics of flowering gene activity in non-reproductive tissues.
Following these directions, researchers can clarify how flowering regulators are co-opted into non-floral contexts, uncover their contributions to plant adaptation, and generate knowledge directly translatable to crop breeding. Ultimately, a deeper understanding of the non-floral functions of FRI, FLC, FT, SOC1, and other key flowering genes will illuminate the evolutionary logic underlying gene multifunctionality, advance our comprehension of plant developmental networks, and inform strategies to enhance crop yield stability and stress resilience under changing environmental conditions.
-
The authors confirm their contributions to the paper as follows: draft manuscript preparation: Zhou HY, Guo P, Liu HM, Zhu P; manuscript and figure revision: Zhou HY, Guo P, Zhu P. All authors approved the final version of the manuscript.
-
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
-
We apologize for not being able to discuss and cite certain relevant studies here due to space restrictions. We thank members of the Zhu and Dean laboratories for insightful discussions. This work is supported by the National Natural Science Foundation of China (Project number: 32570395, awarded to Zhu P), Zhejiang Natural Science Foundation (Project number: LR26C020001, awarded to Zhu P), and the Westlake Education Foundation (awarded to Zhu P).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Huiyan Zhou, Peng Guo, Huimin Liu
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhou H, Guo P, Liu H, Zhu P. 2026. Beyond flowering: the pleiotropic functions of key flowering-time genes. Seed Biology 5: e013 doi: 10.48130/seedbio-0026-0007 

Beyond flowering: the pleiotropic functions of key flowering-time genes
- Received: 09 December 2025
- Revised: 25 January 2026
- Accepted: 02 February 2026
- Published online: 15 April 2026
Abstract: Genetic pathways regulating flowering time have been intensively studied for decades, yet an intriguing paradox has emerged: genes classically defined as floral regulators frequently function in biological processes far beyond flowering. Despite growing evidence, the broader roles of these genes in plant development and stress responses have not been systematically evaluated. In this review, we synthesize recent findings indicating that core flowering regulators serve as central integrators throughout the plant life cycle: influencing processes ranging from seed dormancy and germination to organ development and stress adaptation. The key floral integrators, FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1), have been evolutionarily co-opted to regulate the formation of diverse storage organs, reproductive development, and tolerance to heat stress. Similarly, the floral repressors FRIGIDA (FRI) and FLOWERING LOCUS C (FLC) coordinate vernalization, drought tolerance, and pathogen defense through transcriptional and epigenetic regulatory mechanisms. We propose that the recurrent involvement of these genes across diverse physiological processes reflects their strategic position as regulatory hubs that link environmental sensing to developmental reprogramming. By redefining FRI, FLC, FT and SOC1 as multifunctional integrators rather than dedicated flowering-time regulators, this review presents an updated conceptual framework for understanding how plants coordinate growth, reproduction and resilience in fluctuating environments. Furthermore, it provides valuable insights for developing breeding strategies to simultaneously enhance crop resilience and productivity in the context of climate change.
-
Key words:
- Flowering time /
- FLC /
- FRI /
- FT /
- SOC1 /
- Crops /
- Arabidopsis /
- Environment /
- Growth /
- Development /
- Defense /
- Stress response