










-
Foxtail millet (Setaria italica) is a diploid plant belonging to the genus Setaria within the Poaceae family, and it ranks among the world’s most important grain crops. Foxtail millet and its wild species, Setaria viridis, have the characteristics of a smaller diploid genome, a short growth cycle, easy transformation, and convenient laboratory operation compared with the closely related C4 cereal crops, such as maize (Zea mays), sugarcane (Saccharum officinarum), sorghum (Sorghum bicolor), and switchgrass (Panicum virgatum). It is increasingly emerging as a model crop for research on C4 photosynthesis, as well as cereal crops' resistance and nutrient use efficiency[1]. Double haploid (DH) technology involves the generation of haploid plants followed by chromosome doubling to restore the normal chromosome number of the plant. This approach is an important way to accelerate genetic gains by significantly shortening the breeding cycle[2]. Compared with traditional breeding, the haploid induction and doubling technique can make genetic material homozygous in one generation, which greatly improves the progress and efficiency of breeding.
The identification of haploid inducer genes in maize has spurred extensive genome-editing efforts in related crop species. Among the many haploid-induced genes, PLA (Phospholipase A) was first identified in maize[3]. Subsequent studies have identified and characterized PLA homologs in several species[4], with consistent findings that these genes are highly expressed in the pollen. Multiple plant-like phospholipase homologs exist in foxtail millet, but it is not clear which one triggers haploid induction. Cheng et al.[5] used clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein 9 (Cas9) technology combined with an efficient genetic transformation platform to edit the SiMTL gene and create a haploid induction line. Huang et al.[6] successfully generated haploid plants through seeds by using the egg cell expression of PARTHENOGENESIS (PAR) in dandelion (Taraxacum officinale), which provided the possibility of further promoting the application of DH technology to breeding foxtail millet. According to our preliminary research[7], we screened PLA and found that SiPLA (Seita.9G114100) was highly expressed in the pollen, with the expression levels 500 times higher than those of SiMTL (Seita.9G376800) (Fig. 1a; Supplementary Table S1). We also performed reverse transcription–quantitative polymerase chain reaction (RT-qPCR) on different organs of the cultivar Ci846, and the results showed that the expression level of SiPLA was higher in the pollen than in other tissues (Fig. 1b). Therefore, we designed a guide RNA sequence for the first exon with the use of the CRISPR/Cas9 editing system (Fig. 1c). The guide RNA expression was driven by the OsU3 promoter, whereas Cas9 was under control of the maize Ubiquitin-1 promoter. Cultivar Ci846 was transformed by the Agrobacterium-mediated method, and 104 plants were obtained. Sequence analysis identified 48 allelic editing events at the target site. We generated T1 plants from eight T0 events carrying frameshift mutations in SiPLA (Fig. 1d; Supplementary Fig. S1) and examined their seed set rates. These sipla mutants exhibited an average seed setting rate of 50.1%, ranging from 11.63% to 77.56% (Fig. 1e). Starch accumulation in the individual pollen grains appeared to be normal (Supplementary Fig. S2), consistent with observations in rice (Oryza sativa) and wheat (Triticum aestivum)[8,9]. Next, we selfed three homozygous lines to produce T2 progeny, from which 10 haploids were identified via flow cytometry, yielding an average haploid induction rate (HIR) of 6.33%. To assess the inducer line's capacity to generate haploid embryos in diverse maternal backgrounds, we used artificially emasculated wild-type Ci846 as the maternal parent and pollinated it with pollen from Line 5 (Fig. 1g; Supplementary Fig. S3). Flow cytometry analysis of 441 hybrid plants identified 26 haploids, whereas when wild-type Ci846 was the paternal parent, no typical haploids were detected among 188 F1 plants. The haploid plants exhibited characteristic reductions in the size of all organs, including the leaves, spikes, and anthers, compared with wild-type plants (Fig. 1h).
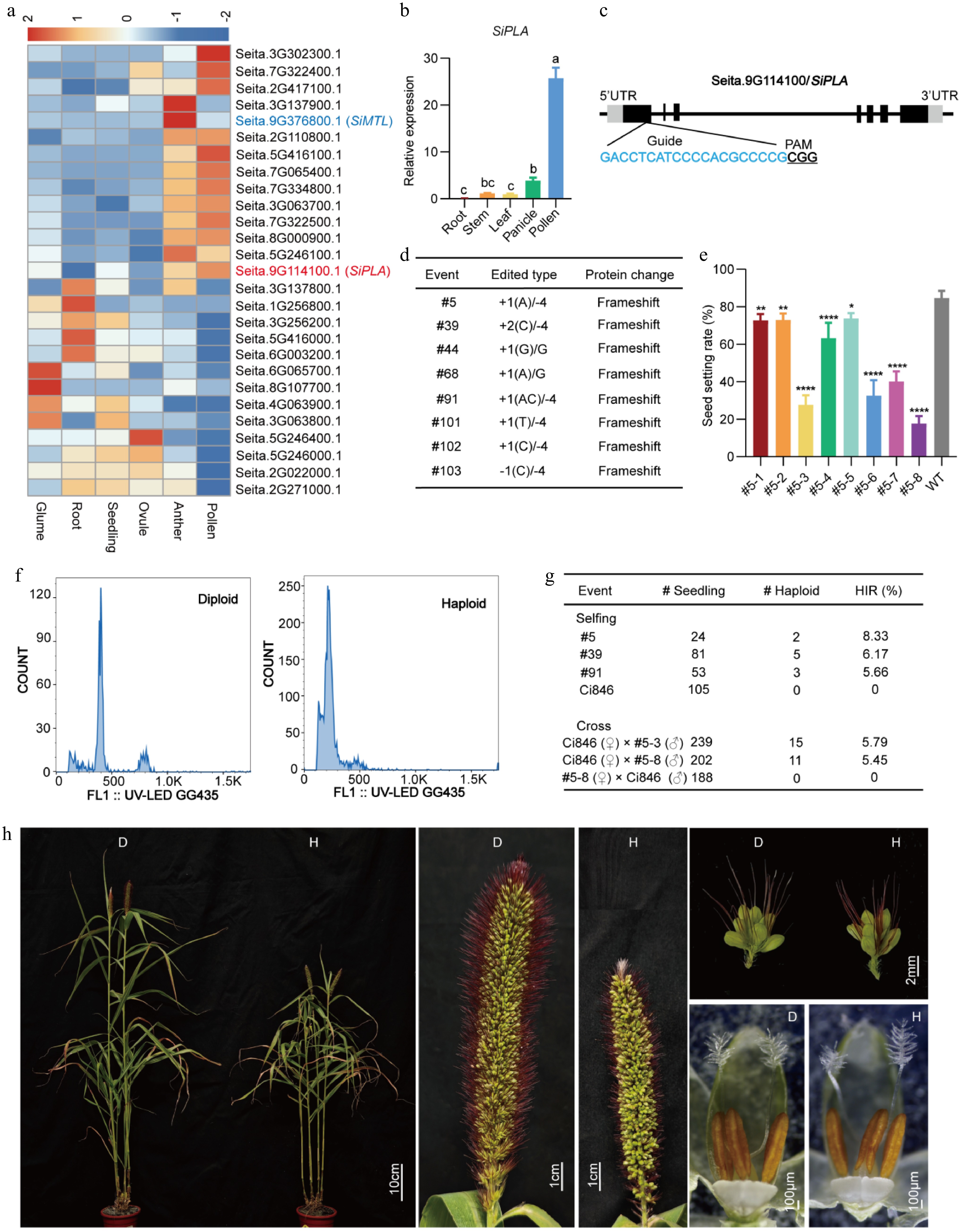
Figure 1.
Haploid induction in foxtail millet. (a) Heatmap of PLA expression identified from our transcriptome data of foxtail millet. Colored boxes represent the log2 fold change. The numbers represent the fragments per kilobase of transcript per million mapped reads (FPKM) value in relative expression levels. Red represents upregulated expression and blue represents downregulated expression. (b) RT-qPCR showed that the relative expression of SiPLA in pollen was high. Significant differences were analyzed using ordinary one-way analysis of variance (ANOVA) by Tukey's method; lowercase letters indicate statistical significance at p < 0.05. (c) Gene structure of SiPLA and CRISPR/Cas9-targeted sites. PAM is underlined and the guide RNA sequence is highlighted in blue. (d) Editing sites and nucleic acid sequence changes in T0 plants. (e) The seed-setting rate of self-pollinated T1 plants derived from Mutant #5. Each error bar represents the mean of three biological replicates with their standard deviations (±SD) (n = 10). Significant differences were analyzed by one-way ANOVA followed by Tukey's multiple comparison test (* p < 0.05; ** p < 0.01; **** p < 0.0001). (f) Verification of haploid plants by flow cytometry analysis. The x-axis indicates the signal peak for the nuclei and the y-axis indicates the number of nuclei. (g) Haploid induction efficiency (HIR) determined by self-pollination or crossing. (h) Phenotype of haploid and diploid plants. "D" represents the diploid control, and "H" represents the haploid.
Our results suggest that haploid induction in foxtail millet can be achieved through knocking out SiPLA. SiPLA is the second homologous phospholipase proven to induce haploids in foxtail millet. Its HIR (6.3%) is significantly higher than that of the previously reported SiMTL (2.8%), providing a more efficient candidate gene for breeding haploid foxtail millet. This finding offers valuable theoretical guidance for the efficient synthesis of apomixis in foxtail millet. Given that different inducer lines acquire different HIRs from maize, we expect that the HIR can be improved by generating monolines with diverse genetic backgrounds using CRISPR/Cas9 methods in foxtail millet, or through combination with mutations in other relevant genes[10].
HTML
-
The transformation was performed using the foxtail millet variety Ci846. Foxtail millet materials were cultivated in the greenhouse of the College of Agriculture, Shanxi Agricultural University, China. The greenhouse's environmental conditions during cultivation were strictly regulated, with the temperature maintained at 25 °C and a photoperiod configured as a 16 h light/8 h dark cycle.
RNA extraction, cDNA synthesis, and RT-qPCR
-
RNA was extracted from the plant materials using a FlaPure Plant Total RNA Extraction Kit (Genesand Biotech, Beijing, China), and subsequently reverse-transcribed into cDNA with the Union Script First-strand cDNA Synthesis kit (Genesand Biotech, Beijing, China). Gene expression levels were assessed using gene-specific primers (Supplementary Table S1) and SYBR Green Super Mix (Mei5bio, Beijing, China) on a Bio-Rad CFX Duet (Bio-Rad CFX Duet, BIORAD, USA). A 20-µL RT-qPCR reaction mixture was prepared, using 2 µL of cDNA obtained by reverse transcription as the template, which included 10 µL of 2× Real-time PCR Super Mix (SYBR Green, with anti-Taq), 0.5 µL each of 10 µM forward and reverse primers, and double distilled H2O to adjust the volume. The procedure was as follows: 95 °C for 1 min, then 40 cycles of 95 °C for 15 s, 60 °C for 15 s, and 72 °C for 30 s. The foxtail millet actin gene SiActin (Seita.8G043100) was used as the internal reference[11]. The relative expression levels of the target genes were calculated by the 2−ΔΔCᴛ method.
Construction of plasmids and generation of transgenic plants
-
The CRISPR-GE online website (
http://skl.scau.edu.cn/home/ ) was used to design sgRNA. Primers were designed (Supplementary Table S1), cloned using pCBC-MT1T2 as a template, and connected to the pHUE411[12] plasmid cut by the BsaI enzyme. The plasmid was sent to the Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing, China, and Ci846 was used as the receptor for genetic transformation.Statistical analysis
-
Analysis of the data was conducted using one-way analysis of variance (ANOVA), followed by Tukey's multiple comparisons test. The significance levels were indicated as follows: * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001 or by lowercase letters at p < 0.05. The software used for image acquisition and processing was Image J software.
Pollen grain staining
-
When the culture material had headed, we took the spiky flower of the grain ear the next day and put it in a centrifuge tube with water. We took an anther of a spiky flower and put it on a slide, added one or two drops of a 5% I2-KI solution, covered the cover glass with forcepsm and gently pressed it so that the pollen grains were fully released, and let it stand for 2–3 minutes. They were observed under a microscope (Olympus BX51, Japan).
Flow cytometry analysis
-
Flow cytometry analysis was performed by Guangzhou Ruiyi Biotechnology Co., Ltd.
Plant materials and growth conditions
-
The authors confirm their contributions to the paper as follows: study design, manuscript writing: Wang JG, Chu X, Chen DY; performing experiments and data analysis: Chen DY, Su M, Sun M, Wang H, Yuan X, Dong S, Li X, Gao L, Yang G. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files. Raw data files in alternative formats may be obtained from the corresponding author upon reasonable request.
-
This work was supported by the National Key Research and Development Program of China (2023YFD1202702-6) and the National Natural Science Foundation (32400217 and 32200222).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Dan-Ying Chen, Min Su, Mengmeng Sun
- Supplementary Table S1 The FPKM values of SiPLA transcriptome data in different tissues and organs of foxtail millet.
- Supplementary Fig. S1 The sequence changes of SiPLA knockout lines on the target site.
- Supplementary Fig. S2 Representative pollens from a sipla plant, showing normal starch accumulation relative to Ci846.
- Supplementary Fig. S3 Phenotypic diagram of wild-type and haploids generated in hybrid progen.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
Chen DY, Su M, Sun M, Wang H, Yuan X, et al. 2026. Establishing a haploid inducer line by edited pollen high expressed gene SiPLA in foxtail millet (Setaria italica). Seed Biology 5: e010 doi: 10.48130/seedbio-0026-0011


|