









-
Sexual reproduction is fundamental for the propagation, genetic diversity, and long-term adaptability of plant species. It ensures the formation of viable seeds, supports crops' productivity, and enables plants to respond evolutionarily to changing environments. Because reproductive development is highly sensitive to external cues, understanding its regulatory mechanisms is crucial for both basic plant biology and agricultural improvement. In plants, spore mother cells arise from somatic cells in the reproductive tissues and undergo meiosis to produce megaspores and microspores. These further develop through mitosis into female and male gametophytes. The female gametophyte comprises the egg cell, the central cell, synergids, and antipodals, whereas the mature male gametophyte (pollen grain) consists of two sperm cells and a vegetative cell. Upon pollination, pollen germinates on the stigma, and the sperm cells are delivered to the ovule through the pollen tube[1,2]. Double fertilization occurs, giving rise to the embryo and endosperm, the latter providing nutrients for the embryo's development[2]. Recent studies indicate that environmental stresses such as heat, cold, drought, and salinity threaten plants' sexual reproduction, reducing fertility, seed yield, and quality, especially in major crops like rice (Oryza sativa) and wheat (Triticum aestivum)[3−7]. Being sessile, plants have evolved sophisticated regulatory mechanisms to sense and respond to these challenges, thereby safeguarding reproductive success under stress[8,9].
Epigenetic regulation is essential for controlling gene expression, silencing transposable elements (TEs), and managing developmental plasticity in plants. The main mechanisms include DNA methylation (5mC), small RNA-mediated regulation, histone modifications, and chromatin remodeling[10−13]. In the model plant Arabidopsis thaliana, DNA methylation occurs at the cytosine residues in the context of CG, CHG, and CHH (where H represents A, C, or T) sequences [10,14]. CG and CHG methylation are mainly maintained by METHYLTRANSFERASE 1 (MET1) and CHROMOMETHYLASE 3 (CMT3), respectively. The maintenance of asymmetric CHH methylation is managed by DOMAIN REARRANGED METHYLTRANSFERASE 2 (DRM2) and CMT2, depending on the genomic region[15−18]. DNA demethylation happens through passive and active processes[19]. Passive demethylation occurs when DNA methyltransferase activity is suppressed during DNA replication, leading to dilution of the methylation marks in newly synthesized DNA strands[10]. In contrast, active demethylation removes 5mC through the base-excision repair pathway and is mainly mediated by DNA glycosylases such as REPRESSOR OF SILENCING 1 (ROS1), DEMETER (DME), DEMETER-LIKE 2 (DML2), and DML3[20]. This dynamic regulation is essential for plants' development and ensures precise epigenetic control during growth and reproduction. Small interfering RNAs (siRNAs) are produced through the coordinated action of RNA POLYMERASE IV (Pol IV), RNA-DEPENDENT RNA POLYMERASE 2 (RDR2), and DICER-LIKE 3 (DCL3). These siRNAs are loaded onto ARGONAUTE 4 (AGO4) to guide RNA-directed DNA methylation (RdDM), which directs DRM2-dependent de novo methylation at homologous loci[20,21]. In parallel, histone modifications influence the chromatin structure and transcriptional activity[22]. Activating marks such as the trimethylation of Histone H3 at Lysine 4 (H3K4me3) and histone acetylation (HAc) promote gene expression[23,24], wherea repressive marks such as the dimethylation of Histone H3 at Lysine 9 (H3K9me2) and trimethylation of Histone H3 at Lysine 27 (H3K27me3) establish heterochromatin and silence transcription[25,26]. Notably, the Polycomb Repressive Complex 2 (PRC2) mediates the deposition of H3K27me3, which is crucial for regulating developmental transitions[27,28]. Together, these epigenetic pathways form an integrated regulatory framework that enables plants to safeguard their reproductive development. Under environmental stresses, this framework becomes particularly vital, providing the molecular flexibility needed to maintain fertility and ensure successful seed set.
This review summarizes the epigenetic regulatory mechanisms underlying sexual reproduction in plants in response to environmental stresses, with a focus on DNA methylation, histone modifications, noncoding RNAs, and chromatin remodeling. Integrating recent advances in epigenetics, we discuss how novel technologies can further illuminate these regulatory mechanisms under stress and explore their potential applications for engineering crops with enhanced "smart" traits and improved resilience to environmental challenges.
-
Environmental stresses have a great effect on plants' growth, development, and reproduction. To maintain fitness and ensure reproductive success, plants have evolved intricate epigenetic regulatory networks that allow them to sense, respond to, and "remember" environmental cues[29,30]. Among these mechanisms, DNA methylation plays a key role in coordinating gene expression, genomic stability, and transposon silencing during stress adaptation[31].
Dynamic regulation of DNA methylation in plant reproduction under abiotic stress
-
Abiotic stresses such as heat, cold, salinity, and drought can cause rapid changes in DNA methylation at specific loci, reflecting an adaptive plasticity of the plant epigenome. Heat stress has been widely studied and is known to cause significant genome-wide changes in DNA methylation patterns (Fig. 1a)[32,33]. For instance, heat stress decreases global DNA methylation in Arabidopsis[34]. In addition, high temperature causes genome-wide hypomethylation in cotton (Gossypium hirsutum) anthers during the tetrad and tapetal degradation stages[12]. In Brassica napus, short-term heat shock promotes demethylation and decreases global methylation in cultured microspores[35]. Thus, there is no consistent trend in heat-induced changes in methylation across different species or cell types.
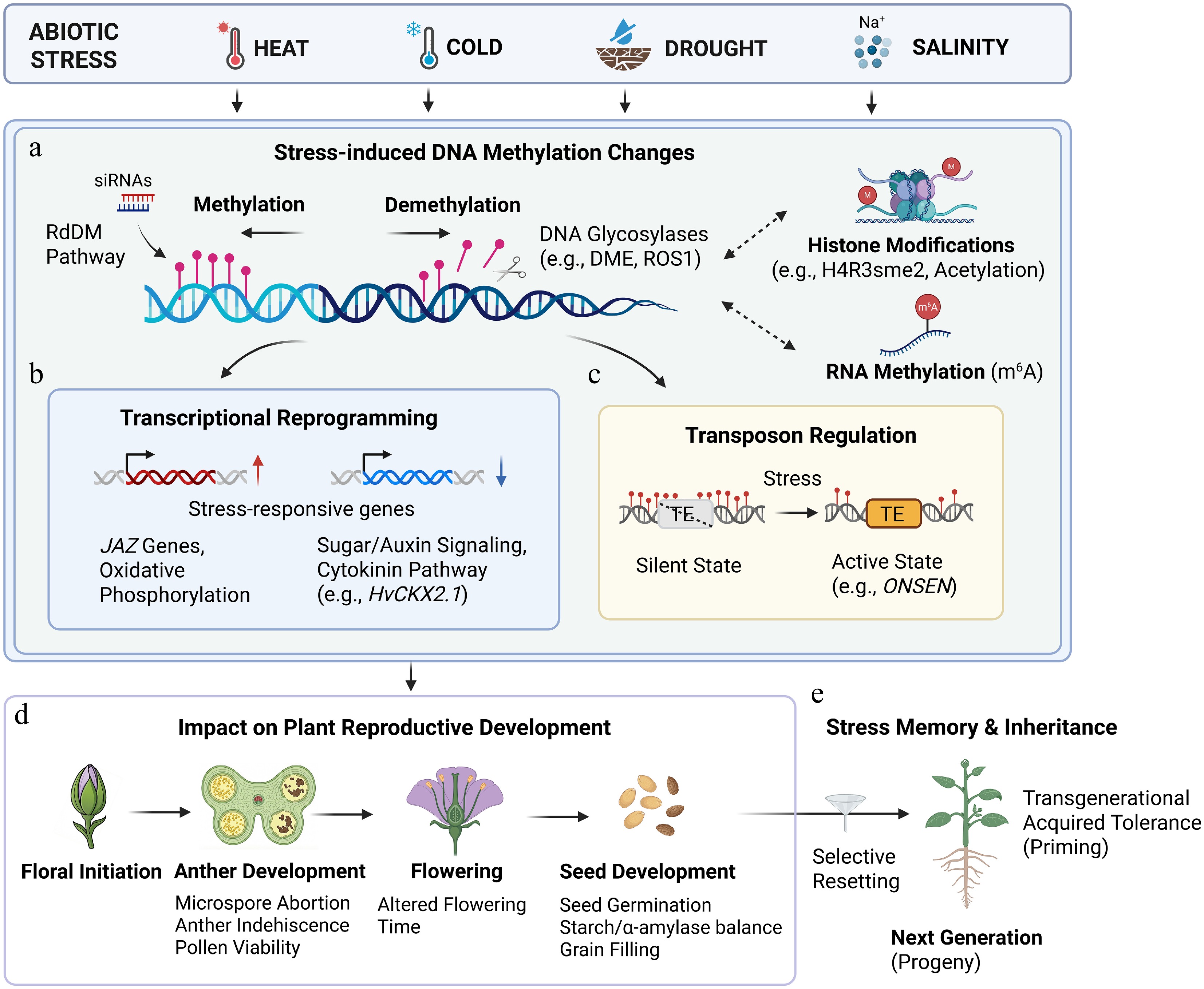
Figure 1.
Abiotic stress-induced DNA methylation dynamics and their roles in plants' reproductive development and stress memory. (a) Abiotic stresses including heat, cold, drought, and salinity trigger dynamic changes in DNA methylation patterns. The regulation involves a balance between the RdDM pathway and active demethylation catalyzed by DNA glycosylases (e.g., DME and ROS1). These dynamic DNA methylation changes interact with histone modifications (e.g., H4R3 symmetric dimethylation and histone acetylation) and RNA methylation (m6A) to modulate the state of chromatin and gene regulation. (b) Stress-induced methylation changes lead to transcriptional reprogramming, including the upregulation of stress-responsive genes (e.g., JAZ genes, oxidative phosphorylation-related genes) and the downregulation of developmental pathways (e.g., sugar/auxin signaling, cytokinin pathway). (c) Altered methylation states also affect transposon regulation, enabling transitions from silenced to active states under stress conditions, exemplified by the activation of the heat-responsive retrotransposon ONSEN. (d) Stress-induced epigenetic alterations affect reproductive development, causing defects such as microspore abortion, anther indehiscence, pollen viability, altered flowering time, and impaired seed germination and grain filling. (e) Epigenetic modifications can form a somatic memory within the same generation or be transmitted to the next generation. Although some stress-induced epigenetic marks contribute to somatic memory or transgenerational acquired tolerance (priming) in the progeny, others undergo selective resetting to maintain genomic integrity. Created in BioRender.
https://BioRender.com/zj6e1eq .High temperature affects seed germination, pollen viability, and anther development, leading to significant yield reductions in crops (Figs 1d and 2a)[36−38]. During seed development, the regulation of DNA (de)methylation positively influences seed germination and thermotolerance[36]. In cotton, heat stress disrupts genome methylation, leading to microspore abortion and anther indehiscence. This disruption alters the expression of genes in the sugar and auxin signaling pathways, which are essential for pollen maturation (Fig. 1b, d)[35−39]. Furthermore, single-base resolution methylome analysis of cotton's cytoplasmic male sterility (CMS) systems showed extensive epigenomic changes under heat stress. DNA methylation-mediated transcriptional changes in genes related to oxidative phosphorylation may modulate the balance of mitochondrial adenosine triphosphate (ATP) synthesis and reactive oxygen species (ROS) bursts under heat stress during anther development (Fig. 1b)[33]. In rice, an analysis of spikelets from 14 varieties revealed that heat-tolerant varieties exhibited DNA demethylation during flowering, whereas heat-sensitive varieties showed increased methylation. Their flowering time differences are associated with their methylation status (Fig. 1d)[40]. Additionally, in the heat-tolerant rice variety Nagina 22, JASMONATE ZIM-DOMAIN (JAZ) genes are specifically upregulated under heat stress, with demethylation and increased histone acetylation at their promoters (Fig. 1b)[41]. These findings show that DNA methylation status is closely linked to heat tolerance and yield stability. Loss of methylation can activate TEs. For example, defects in the siRNA biosynthesis pathway lead to heat-induced activation of ONSEN retrotransposon[42]. ONSEN couples its activation to a stress-defense pathway and amplifies preferentially in dividing tissues. This ensures efficient propagation while avoiding stable remethylation (Fig. 1c). These findings demonstrate that heat-induced demethylation can reactivate retrotransposons, which exploit the plant's stress pathways for mobilization[43].
Plants' responses to cold stress are also regulated by DNA methylation. Adaptive stress tolerance in plants can be transmitted across generations via epigenetic mechanisms. In rice, repeated low-temperature exposure confers heritable cold tolerance mediated by DNA methylation. Hypomethylation of the ACQUIRED COLD TOLERANCE 1 (ACT1) promoter sustains its expression under cold stress, directly affecting tolerance, and natural variation in ACT1 methylation aligns with geographic distribution and cold adaptation[44]. Importantly, this epigenetic change improves reproductive performance under cold stress, as evidenced by increased seed-setting ratios, whereas key agronomic traits such as panicle number, grains per panicle, 1,000-grain weight, and overall grain yield are maintained or enhanced[44]. These findings highlight DNA methylation as a molecular basis for inherited adaptive traits and underscore the potential of epigenetic regulation for improving stress resilience and crops' productivity.
Salinity stress also triggers epigenetic modifications to help plants cope with high salt conditions. In Arabidopsis, the floral initiator Shk1 KINASE BINDING PROTEIN 1 (SKB1) and Histone H4 Arginine 3 symmetric dimethylation (H4R3sme2) integrate the plant's developmental progression with responses to salt stress. Loss of SKB1 function results in salt hypersensitivity, delayed flowering, and retarded growth. SKB1 is associated with chromatin and promotes the deposition of H4R3sme2, thereby repressing the transcription of FLOWERING LOCUS C (FLC) and multiple stress-responsive genes (Fig. 1a)[45]. In response to drought stress, plants used a variety of mechanisms to enhance their tolerance. For example, the rice bZIP transcription factor OsbZIP46 contributes to drought resistance by directly or indirectly activating the expression of multiple abscisic acid (ABA)- and stress-responsive genes[46]. At the same time, drought stress can also trigger epigenetic mechanisms, which help fine-tune the expression of stress-responsive genes and facilitate adaptive responses. Active DNA demethylation, mediated by DNA glycosylases like DME and ROS1, balances methyltransferase activity to maintain epigenomic flexibility (Fig. 1a). In barley (Hordeum vulgare), the DME homolog HvDME is essential for seed development and participates in drought-responsive demethylation[47]. In addition, terminal drought stress induces hypermethylation of the promoter of the cytokinin oxidase gene HvCKX2.1, reducing its expression and helping maintain cytokinin levels for grain-filling (Fig. 1b)[48]. Together, these findings highlight that DNA methylation is a highly dynamic regulator of stress-responsive transcriptional networks, rather than a static silencing mechanism.
DNA methylation and transgenerational stress memory
-
Epigenetic states triggered by environmental fluctuations can be inherited across generations in both mammals and seed plants[49,50]. These stress-induced changes in DNA methylation often persist even after the stress is removed, establishing an epigenetic memory that pre-adapts plants to subsequent environmental challenges (Fig. 1e)[29,51]. For instance, in Arabidopsis, parental exposure to stress alters global DNA methylation in the offspring and enhances their stress tolerance, suggesting that stress signals can be transmitted through the germline[52]. Similarly, in rice, acquired cold tolerance can be inherited across generations via heritable changes in DNA methylation[44]. Moreover, heat stress during grain filling leads to hypomethylation of starch biosynthesis gene promoters and hypermethylation of α-amylase gene promoters in rice. These epigenetic modifications establish an adaptive transgenerational memory, enabling subsequent generations to better withstand heat stress[53]. However, the transmission of stress-induced epialleles is tightly regulated to preserve genomic integrity. In Arabidopsis, the memory of hyperosmotic stress is embedded at epigenetically unstable loci. During pollen development, the male germline utilizes DNA glycosylases to erase many stress-induced epigenetic marks, thereby preventing uncontrolled inheritance of such epialleles (Fig. 1e)[54]. Collectively, these studies demonstrate that DNA methylation-linked stress memory is not simply passed on passively but undergoes strict regulation during reproduction, balancing environmental adaptation with epigenomic stability.
-
Although DNA methylation plays a key role in stress-responsive reproductive regulation, histone modifications act in parallel to further modulate flowering and seed development under changing environments.
Histone methylation as a crucial epigenetic regulator of plant reproduction under environmental stresses
-
In addition to DNA methylation, diverse histone and RNA modifications constitute key epigenetic mechanisms that precisely regulate plants' reproductive development under environmental stresses (Fig. 2a). Beyond DNA methylation, histones are also subject to methylation, which play indispensable roles in the epigenetic regulation of flowering in plants. In Arabidopsis thaliana, the floral repressor FLC modulates flowering time by regulating the expression of its downstream targets FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1)[55]. Elevated temperature induces the JUMONJI-C domain-containing demethylases JMJ30 and JMJ32, which remove the repressive H3K27me3 mark from the FLC locus, thereby maintaining the expression of FLC and preventing premature flowering[56]. Similarly, JMJ13, another H3K27me3 demethylase, functions as a temperature- and photoperiod-dependent floral repressor, and precisely regulates flowering time under varying environmental cues[57]. Under high-temperature conditions, jmj13 mutant plants exhibit early flowering irrespective of daylength, whereas under low temperature, early flowering occurs only under short days. In all cases, FT transcript levels are elevated, suggesting that JMJ13 modulates flowering primarily through activating FT[57]. Another H3K27me3 demethylase, RELATIVE OF EARLY FLOWERING 6 (REF6), forms a positive regulatory loop with the heat-induced transcription factor HEAT SHOCK TRANSCRIPTION FACTOR A2 (HSFA2) (Fig. 2b)[58]. This feedback loop activates SUPPRESSOR OF GENE SILENCING 3 (SGS3)-INTERACTING PROTEIN 1 (SGIP1), promoting SGS3 degradation and thereby releasing the trans-acting siRNA (tasiRNA) target HEAT-INDUCED TAS1 TARGET 5 (HTT5), ultimately facilitating heat-induced flowering (Fig. 2b)[58]. Histone methylation also exerts critical regulatory effects on seed development under thermal stress. In rice, the PRC2 component FERTILIZATION-INDEPENDENT ENDOSPERM 1 (OsFIE1) is essential for the transition of the endosperm from the syncytial to the cellularized state during seed development[59]. The epigenetic status of OsFIE1, including both DNA methylation and H3K9me2 levels, is sensitive to temperature. Heat stress reduces the methylation of OsFIE1 and increases its transcriptional activity, which negatively correlates with seed size and results in smaller seeds[60]. Collectively, these studies highlight the pivotal roles of histone methylation in integrating environmental cues to modulate flowering time and seed development, underscoring its importance in maintaining reproductive success under stress conditions.
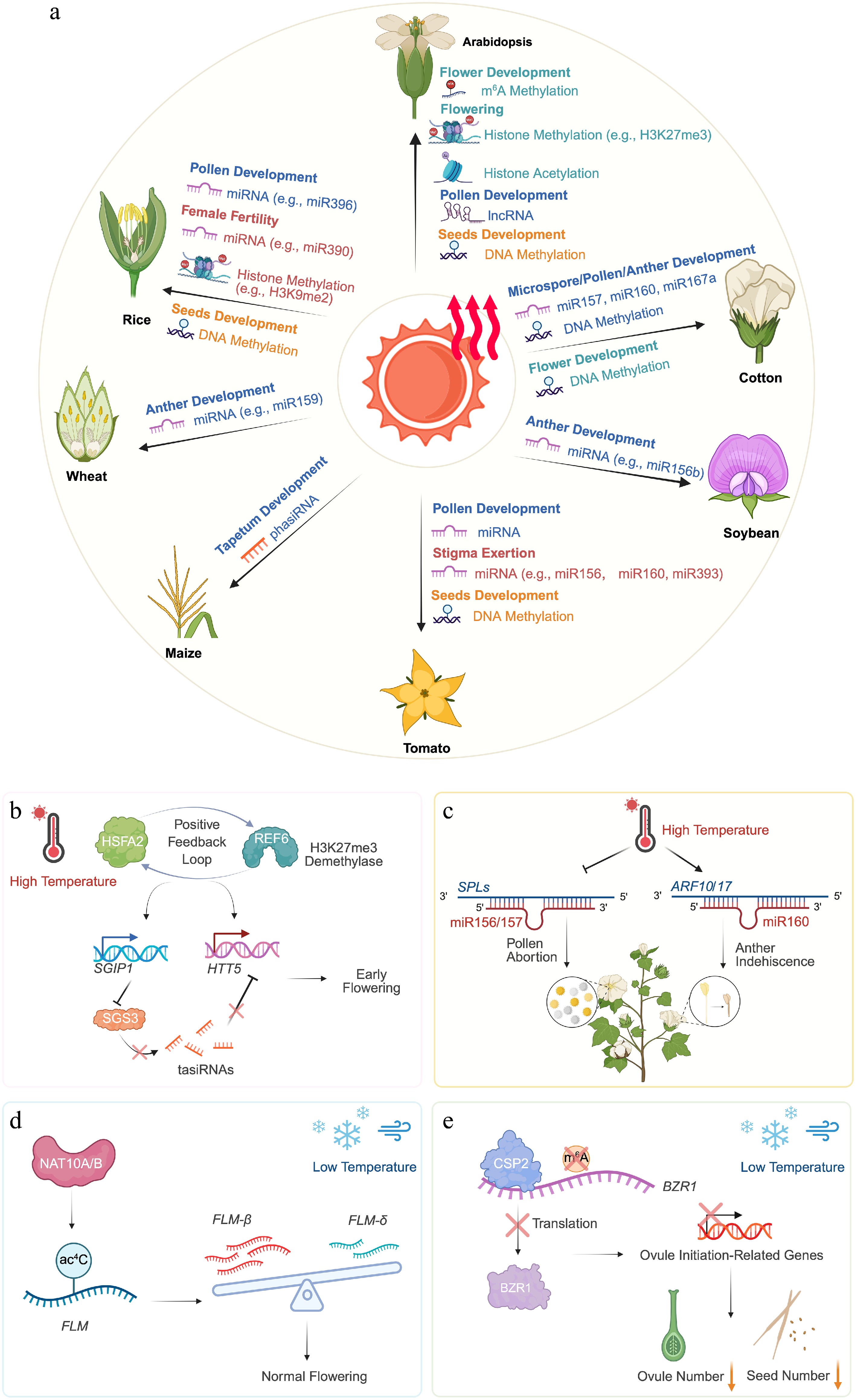
Figure 2.
Epigenetic mechanisms safeguarding plant reproduction under abiotic stress. (a) Epigenetic regulation plays a pivotal role in maintaining the proper progression of sexual reproduction in plants under heat stress. Multiple epigenetic mechanisms are involved in modulating heat stress responses, including DNA methylation, histone methylation, and histone acetylation, as well as noncoding RNAs such as lncRNAs, miRNAs, and siRNAs. Together, these regulatory mechanisms influence flowering time and the development of floral organs and seeds. (b) The H3K27me3 demethylase REF6 establishes a positive regulatory feedback loop with the heat-responsive transcription factor HSFA2. This loop induces the expression of SGIP1, promotes degradation of SGS3, and consequently derepresses the tasiRNAs target HTT5, thereby contributing to heat-induced flowering. (c) In cotton, overexpression of miR157 or miR160 compromises auxin signaling, thereby enhancing the sensitivity of male fertility to heat stress and leading to microspore abortion or anther indehiscence. (d) Under low ambient temperatures, Arabidopsis NAT10A/B promote temperature-responsive ac4C deposition on FLM transcripts, thereby modulating alternative splicing of FLM and preventing extremely late flowering. (e) In Arabidopsis, brief exposure to chilling induces the RNA-binding protein CSP2, which binds to BZR1 mRNA and modulates its m6A modification, thereby reducing the mRNA translation efficiency of BZR1, inhibiting ovule initiation, and decreasing seed number. Created in BioRender.
https://BioRender.com/l6d6j9a .Dynamics of histone acetylation in plant reproduction under environmental stresses
-
Histone acetylation is a major form of histone-related epigenetic modification and plays an important role in helping plants adapt to environmental stresses, particularly during the reproductive stages. In Arabidopsis thaliana, HISTONE DEACETYLASE 6 (HDA6) enhances thermotolerance by modulating histone acetylation levels. Loss of HDA6 disrupts the expression of genes associated with reproductive development under heat stress, thereby impairing sexual reproduction[61]. Similarly, HISTONE DEACETYLASE 9 (HDA9) regulates seeds' responses to drought and salt stresses. In this context, hda9-1 and hda9-2 mutants exhibit increased stress sensitivity and accelerated seed germination, likely caused by elevated histone acetylation at the promoters of stress-responsive genes[62]. In addition, POWERDRESS (PWR) interacts with HDA9 to cooperatively mediate histone deacetylation, which is important for floral transition under heat stress[63,64]. In the pwr mutant, transcriptional levels of heat-responsive factors such as PHYTOCHROME INTERACTING FACTOR 4 (PIF4) are reduced, resulting in decreased thermosensitivity during floral transition, indicating that PWR-mediated deacetylation is essential for thermomorphogenesis[65]. Moreover, histone acetylation regulates seed germination under stress. SILENT INFORMATION REGULATOR2 HOMOLOG (SRT2), a histone deacetylase, represses the expression of VESICLE-ASSOCIATED MEMBRANE PROTEIN 714 (VAMP714) to modulate seed germination under salt stress[66].
Histone modifications in plants' stress memory
-
Plants can establish short-term stress memories, and the formation and maintenance of such memories are closely associated with histone methylation. The H3K4 methyltransferases SET DOMAIN GROUP 25 (SDG25) and ARABIDOPSIS TRITHORAX 1 (ATX1) deposit H3K4me3 marks at heat stress-responsive loci, preventing the establishment of DNA methylation at these regions and sustaining the proper expression of heat-inducible genes during and after heat exposure[67]. To initiate flowering, plants require prolonged cold exposure (vernalization) and maintain this cold memory throughout their growth. During prolonged cold, CASEIN KINASE II (CK2), induced by low temperature, phosphorylates the H3K27 methyltransferase, promoting genome-wide accumulation of H3K27me3 in Arabidopsis. This cold–CK2–PRC2 signaling pathway facilitates PRC2 enrichment at the FLC locus[68], increasing H3K27me3 levels and switching transcription from activation to repression. After vernalization, FLC remains transcriptionally silenced throughout the subsequent growth stages, ensuring normal flowering. This cold-induced epigenetic memory is eventually erased in the next generation[69−71].
-
In plants, noncoding RNAs are broadly classified into small noncoding RNAs (small ncRNAs) and long noncoding RNAs (lncRNAs) according to their length[72,73]. Small noncoding RNAs include microRNAs (miRNAs) and siRNAs, which are processed by DCL proteins and function in association with AGO family proteins to mediate post-transcriptional or transcriptional gene silencing[72,74]. Most lncRNAs are primarily transcribed by RNA polymerase II and function through modulating transcriptional and epigenetic processes[75,76]. Under environmental stress conditions, noncoding RNAs play pivotal roles in safeguarding the normal progression of sexual reproduction in plants.
Roles of small RNAs in regulating reproduction in plants under environmental stresses
-
miRNAs and siRNAs are important post-transcriptional regulators that modulate plants' reproductive development, ensuring proper anther and ovule formation and contributing to fertility under both normal and stress conditions (Fig. 2a). miRNAs play crucial roles in regulating anther development in plants. For example, miR319 promotes secondary cell wall thickening in the anther endothecium of Arabidopsis by repressing TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR 24 (TCP24)[77]. Similarly, AUXIN RESPONSE FACTOR 17 (ARF17), which is expressed in the microspores and tetrads and is required for tetrad wall synthesis, is directly targeted by miR160, thereby critically regulating the development of pollen[78]. miRNAs also participate in heat stress responses, enhancing reproductive thermotolerance in plants. In Arabidopsis, miR156 regulates normal growth under heat stress by repressing the expression of SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) transcription factors. Given that SPL proteins also play critical roles in flowering time and the development of gametophytes (e.g., in pollen wall formation), this suggests that this epigenetic regulatory module may be essential for ensuring proper sexual reproduction under stress conditions[79−81]. In rice, the transcription factor GROWTH-REGULATING FACTOR 4 (OsGRF4) is encoded by a semi-dominant quantitative trait locus (QTL) for grain size and weight (GS2)[82,83]. OsGRF4 is post-transcriptionally regulated by miR396. In GS2, OsGRF4 mRNA escapes miR396-mediated degradation, leading to accumulation of OsGRF4, which promotes nitrogen storage and metabolism, and consequently increases grain size and yield[84]. However, under high-temperature conditions, OsGRF4 in GS2 modulates the transcription and splicing of reproduction-related genes in the anthers, thereby reducing thermotolerance, pollen viability, and seed set[85]. Thus, the miR396-mediated post-transcriptional regulation of OsGRF4 plays a pivotal role in the development of rice under both nitrogen deficiency and heat stress[84,85]. Most studies on the effects of stress on sexual reproduction in rice have focused on male fertility, whereas female fertility has received comparatively less attention. Emerging evidence indicates that female fertility is also regulated by miRNAs. miR390 cooperates with AGO7 to form the RNA-induced silencing complex (RISC), which triggers siRNA biogenesis and suppresses the expression of auxin response factors[86−89]. The thermo-sensitive female sterility 1 (tfs1) mutant, which carries a point mutation in AGO7, disrupts RISCs' formation and reduces the accumulation of siRNAs, ultimately compromising female fertility under high-temperature conditions[89]. High night temperatures also impair reproductive development and grain-filling in rice, and recent studies have revealed regulatory roles for miRNAs in this response. Several high night temperature-responsive miRNAs that differ between heat-tolerant and heat-sensitive rice lines, target mRNAs involved in heat-related biological processes[90]. In wheat (Triticum aestivum), GAMYB, a target of miR159, participates in flower development and gibberellin (GA) signaling in aleurone cells[91,92]. Notably, the expression of TamiR159 is downregulated after 2 h of heat treatment, suggesting a role in thermotolerance[93]. Moreover, the overexpression of TamiR159 in rice causes delayed heading and male sterility, indirectly supporting its importance in maintaining normal anther development in wheat under heat stress[94]. miRNAs also regulate male thermotolerance in cotton, exhibiting stage-specific expression patterns during anther development[95]. In cotton, overexpression of miR157 or miR160 disrupts auxin signaling, rendering male fertility more sensitive to heat stress and leading to phenotypes such as microspore sterility or anther indehiscence (Fig. 2c)[96]. Proper miRNA regulation is therefore critical for the application of the three-line hybrid system in cotton, particularly under heat stress. For instance, the CMS-D2 restorer line based on cotton cytoplasm is heat-sensitive, severely restricting the utility of the three-line system. Differential expression of heat-responsive miRNAs has been observed among cotton restorer lines carrying distinct cytoplasms under high-temperature stress[97]. In heat-sensitive restorer lines, downregulation of mtr-miR167a_R+1 leads to increased expression of its target AUXIN RESPONSE FACTOR 8 (ARF8), disrupting auxin signaling and ultimately reducing pollen viability[97]. In soybean (Glycine max), downregulation of specific miRNAs contributes to protecting male fertility from heat stress. For example, the overexpression of gma-miR156b in Arabidopsis leads to heat-sensitive anther fertility, highlighting its conserved regulatory role[98]. In tomato (Solanum lycopersicum), heat-responsive miRNAs have also been detected at the post-meiotic stage and in mature pollen[99]. Additionally, several miRNAs are downregulated in the pistils under heat stress, contributing to the regulation of stigma exsertion[100]. In maize (Zea mays), phased siRNAs (phasiRNAs) play critical roles in anther development under high-temperature conditions. DCL5, a monocot-specific protein, regulates the biogenesis of 24-nt phasiRNAs, which facilitate heat stress responses and maintain male fertility[101,102]. Mutations in DCL5 abolish the production of 24-nt phasiRNAs, making the dcl5 mutant temperature-sensitive during meiosis and leading to abnormal development in the anthers[102]. Collectively, these studies highlight the central roles of miRNAs and siRNAs in safeguarding plants' reproductive development under environmental stress conditions.
lncRNA-dependent control of fertility in response to temperature and photoperiod
-
lncRNAs provide an additional epigenetic layer influencing reproductive development under environmental stresses. Upon heat stress, numerous pollen-specific lncRNAs are induced in Arabidopsis thaliana, suggesting that lncRNAs contribute to the reproductive response to thermal stress[103]. Beyond temperature, lncRNAs also play critical roles in regulating fertility in response to varying photoperiods. Photoperiod-sensitive male sterility (PSMS) in rice is characterized by male sterility under long-day conditions and fertility under short-day conditions. Two loci, PHOTOPERIOD-SENSITIVE GENIC MALE STERILITY 1 (PMS1) and PMS3, were identified in the PSMS line Nongken 58S[104]. PMS1 encodes a lncRNA, PMS1T, which is preferentially expressed in young panicles. Under long-day conditions, PMS1T is targeted by miR2118, leading to the production and accumulation of 21-nt phasiRNAs and ultimately causing male sterility[105,106]. PMS3 encodes another lncRNA, the LONG-DAY SPECIFIC MALE-FERTILITY-ASSOCIATED RNA (LDMAR)[107,108]. Proper accumulation of LDMAR is critical for maintaining male fertility under long-day conditions. Compared with wild-type Nongken 58, the Nongken 58S line carries a single-nucleotide polymorphism (SNP) within LDMAR that alters its secondary structure and increases DNA methylation at its promoter region. This epigenetic modification suppresses the transcription of LDMAR, triggers premature programmed cell death (PCD) in developing anthers, and consequently leads to long-day-dependent male sterility[107]. Moreover, the restoration of fertility following a shift to short-day conditions is delayed in Nongken 58S[108]. Together, these findings highlight lncRNAs as key epigenetic regulators that fine-tune male reproductive development and fertility in response to environmental and photoperiodic cues.
-
RNA modifications constitute an important post-transcriptional regulatory layer that enables plants to fine-tune their gene expression in response to environmental stresses (Fig. 2a). These modifications dynamically influence RNA metabolism, including stability, splicing, and translation, thereby shaping stress-responsive gene expression programs[109,110].
m6A RNA modificationin regulating reproductive development in plants under temperature stress
-
Among the diverse RNA modifications identified in plants, N6-methyladenosine (m6A) is the most abundant internal modification of mRNA and is deposited, removed, and interpreted by conserved writer, eraser, and reader proteins[111]. Increasing evidence indicates that m6A plays a central role in coordinating plants' stress responses in a context- and tissue-dependent manner. In Arabidopsis thaliana, m6A dynamically modulates the variability of gene expression in floral tissues under heat stress, thereby affecting reproductive thermotolerance and fertility outcomes. For example, the disruption of m6A demethylation mediated by the RNA demethylase ALKBH10 impairs the heat-induced activation of numerous genes, ultimately leading to shortened siliques and reduced seed fertility[112]. In addition, m6A modification also participates in the regulation of sexual reproduction in plants in response to cold stress. Under low-temperature conditions, the Arabidopsis RNA-binding protein COLD SHOCK PROTEIN 2 (CSP2) modulates the m6A methylation of BRASSINAZOLE RESISTANT 1 (BZR1) mRNA, thereby affecting BZR1's translation efficiency (Fig. 2e)[113]. This negative regulatory mechanism disrupts normal sexual reproductive development, leading to a significant reduction in ovule number and ultimately resulting in shortened siliques and decreased seed set[113].
mRNA acetylation as a post-transcriptional regulator of flowering under cold stress
-
In addition to histone acetylation, recent studies have revealed that mRNA acetylation also plays an important role in plants' responses to environmental cues. In Arabidopsis, the mRNA N4-acetylcytidine (ac4C) writers N-ACETYLTRANSFERASE 10A/B (NAT10A/B) are rapidly induced under low-temperature conditions, leading to ac4C modification of FLOWERING LOCUS M (FLM) transcripts (Fig. 2d)[114]. This modification facilitates the recruitment of SPLICING FACTOR 1 (SF1) to FLM, thereby modulating its alternative splicing. Through this mechanism, Arabidopsis fine-tunes flowering time under cold conditions and prevents excessive flowering delay[114].
-
Chromatin-level epigenetic modifications mediate plants' reproductive responses to abiotic stress. It balances genomic stability, gene regulation, and physiological adaptations during critical developmental stages such as gametogenesis and fruit development.
Maintaining the chromatin structure during meiosis is foundational for fertility, but heat can disturb this process. For instance, heat stress impairs the centromeres' structure and segregation of meiotic chromosomes in Arabidopsis, leading to defects and reduced fertility[115]. Heat stress also likely influences chromatin's conformation in pollen through critical chromatin modifications[116]. Stress conditions often relax epigenetic silencing, potentially allowing TEs to mobilize. A transposon surveillance system operating at the chromatin level is crucial to maintain male fertility during stress and prevent the activation of deleterious TEs in the germline[117]. A single-cell transcriptome atlas and chromatin accessibility survey of cotton anthers revealed that tapetal cells are particularly sensitive to heat. Heat reduces the chromatin accessibility of specific genes, such as QUARTET 3 (QRT3) and CYTOCHROME P450 703A2 (CYP703A2), silencing them and causing defects in pollen wall formation and male sterility[118]. These results further illustrate that chromatin-level stress responses are deeply integrated with reproductive development.
Beyond structural integrity, chromatin modifications also regulate key stress-response genes. Overexpression of the chromatin remodeling ATPase genes AtCHR12 or AtCHR23 reduced seed germination in Arabidopsis, especially under salt or heat stress. Epigenetic repression of multifunctional stress regulators enables plants to balance stress tolerance with reproductive development by preventing harmful overactivation of defense pathways[119]. A critical question regarding these chromatin modifications is their heritability. In Arabidopsis, stress-triggered loss of gene silencing can be transmitted to the next generation. However, the male germline possesses an active reprogramming system that selectively resets these marks. This ensures that only a subset of stress-induced epialleles is heritable, preventing long-term destabilization of the germline's epigenome[116].
The upstream signals driving these chromatin-level changes are linked to the cell's physiological state. Redox status may act as a metabolic sensor that connects abiotic stress to chromatin remodeling[120]. The connection between cellular physiology and genetic regulation is further supported by multi-omics analyses. In tomato fruit, transcriptome analyses suggest that epigenetic mechanisms mediate heat-induced transcriptional changes. Genes involved in chromatin remodeling and histone methylation are enriched among those upregulated in response to heat[121].
-
In this review, we have emphasized the central role of epigenetic mechanisms, including DNA methylation, histone modifications, noncoding RNAs, and chromatin remodeling, in coordinating reproductive development in plants under environmental stresses. These multilayered regulatory networks enable developmental plasticity, mediate stress memory, and, in some cases, facilitate transgenerational adaptation. We have also discussed how emerging high-resolution technologies, from single-cell and spatial multi-omics to long-read epigenomics and chromatin conformation profiling, are transforming our understanding of stress-responsive epigenetic regulation and paving the way for strategies to enhance reproductive resilience and crops' performance in challenging environments.
High-resolution epigenomic dissection of reproductive stress responses
-
Plants' reproductive organs are embedded within complex tissues, making it difficult to resolve cell-type-specific epigenetic regulation using bulk approaches. Recent advances in single-cell and spatial multi-omics have begun to overcome these limitations. Single-cell RNA sequencing (scRNA-seq) has revealed pronounced transcriptional heterogeneity and cell type-specific sensitivity to heat stress during anther and stamen development in both Arabidopsis and crops[118,122], whereas fixation-based based scRNA-seq methods have extended the applicability of this technology to recalcitrant and field-grown samples[123]. However, understanding stress adaptation requires us to move beyond the transcriptome to directly profile epigenetic states. Techniques such as the single-cell assay for transposase-accessible chromatin using sequencing (scATAC-seq) enable the profiling of chromatin's accessibility, allowing the identification of cell-type-specific regulatory elements and stress-induced changes in chromatin's openness. In maize, scATAC-seq of developing anthers under heat stress pinpointed cis-regulatory elements whose accessibility is altered in heat-sensitive cell types, such as the tapetum or microspores[124]. Similarly, single-cell chromatin immunoprecipitation sequencing (scChIP-seq) methods, though technically challenging, are emerging to profile histone modifications (e.g., H3K27me3, H3K4me3) at a cellular resolution, potentially revealing how stress reshapes the epigenome in specific lineages leading to gametes[125]. In parallel, long-read-based approaches such as Fiber-seq and advances in capturing chromatin conformation have uncovered fine-scale cis-epigenetic features and dynamic three-dimensional genome organization in plants' reproductive cells[126,127]. Such innovations are redefining the study of reproductive stress, moving from descriptive, tissue-level analyses to integrated, cell-resolved insights into the epigenetic mechanisms.
Transgenerational epigenetic regulation under stress
-
A defining feature of epigenetic regulation is its potential persistence through mitosis and, in some cases, across generations. Although stress-induced epigenetic memory has been observed in plants, the principles determining which marks are transmitted through the germline and which are reset remain poorly understood. In Arabidopsis, many stress-induced DNA methylation changes arise at epigenetically unstable loci and are actively erased by DNA demethylases such as DME and ROS1, suggesting a selective filtering mechanism that limits the inheritance of deleterious epialleles[54]. Future studies need to systematically define which epigenetic features, including specific DNA methylation contexts, histone modifications, or small RNA pathways, are more likely to escape germline reprogramming. A central unresolved question is how stress-induced epigenetic states are selectively inherited across generations. It remains unclear whether specific reader proteins protect certain epigenetic marks during gametogenesis, or whether epigenetic reprogramming differs between male and female germlines. Evidence from rice indicates that multigenerational drought or heat stress can induce heritable DNA hypomethylation at specific loci, contributing to adaptive phenotypic variation[53,128]. Using isogenic lines, isolated gametes or embryos, and high-resolution technologies like Fiber-seq will be key to decoding epigenetic inheritance, potentially allowing the deliberate stabilization of beneficial stress memories to enhance a crop's resilience without changing its DNA.
Epigenetic engineering and crop improvement
-
The growing understanding of epigenetic regulation in plants' stress responses offers new avenues for crop improvement. Beyond traditional genetic engineering, "epigenetic engineering" aims to stably modulate gene expression to enhance complex traits such as stress resilience. Targeting key regulators, such as histone demethylases, deacetylases, or chromatin remodelers, can reprogram entire gene networks, providing broader tolerance than single-gene edits. More precise approaches utilize epigenome editing, such as CRISPR-dCas9 fused to effector domains to modify histone marks (e.g., for H3K27me3) or DNA methylation at specific loci, enabling the targeted activation or repression of stress-responsive genes[129,130]. Ensuring stability, specificity, and heritability remains a key challenge. Complementarily, harnessing natural epigenetic variation through epiQTL mapping or epigenome-wide association studies allows the identification of beneficial epialleles for breeding. Integrating high-throughput epigenotyping with genomic selection could accelerate the development of "epi-bred" crops with enhanced resilience.
In conclusion, integrating single-cell and single-molecule epigenomics, deciphering transgenerational epigenetic inheritance, and leveraging advanced epigenetic engineering will be key to developing climate-resilient crops and safeguarding global food security.
-
The authors confirm their contributions to the paper as follows: study design and writing the manuscript: Zhu R, Qi L, Qian W; figure preparation: Zhu R, Qi L. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article, as no datasets were generated or analyzed during the current study.
-
This work was supported by the China Postdoctoral Science Foundation (2024T170028 and 2023M740096 to Lijuan Qi), and the National Natural Science Foundation of China (32270288 to Weiqiang Qian).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Ruixian Zhu, Lijuan Qi
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhu R, Qi L, Qian W. 2026. Epigenetic networks in sexual reproduction of plants under environmental stresses. Seed Biology 5: e011 doi: 10.48130/seedbio-0026-0006 

Epigenetic networks in sexual reproduction of plants under environmental stresses
- Received: 21 December 2025
- Revised: 23 January 2026
- Accepted: 27 January 2026
- Published online: 08 April 2026
Abstract: Sexual reproduction in plants is precisely regulated by intricate epigenetic networks under environmental stress, which are crucial for reproductive success and crop yield. This review systematically summarizes recent advances in epigenetic mechanisms underlying sexual reproduction in plants in response to stresses such as heat, cold, drought, and salinity. We focus on the dynamic roles of DNA methylation, histone modifications, noncoding RNAs, and chromatin remodeling during reproductive development. Stress induces methylation reprogramming at specific genomic loci and influences transposon activity via the RNA-directed DNA methylation pathway. Histone modifications, including H3K27me3, H3K4me3, and acetylation, integrate environmental cues to regulate flowering time, endosperm development, and gametophyte formation. Noncoding RNAs, such as microRNAs, phased small interfering RNAs, and long noncoding RNAs, participate in stress responses through post-transcriptional regulation to maintain normal pollen and ovule development. Furthermore, changes in chromatin's accessibility and structure coordinate stress memory and transgenerational adaptation. These epigenetic layers interact to form a multidimensional regulatory network, enabling developmental plasticity under stress conditions. This review provides an integrated perspective on the epigenetic regulation of plant reproduction under environmental challenges and highlights potential epigenetic strategies for improving crops in terms of stress resistance.
-
Key words:
- Epigenetics /
- Sexual reproduction /
- Environmental stresses