








-
A major problem in global health is the lack of affordable and effective drugs available to fight many diseases[1], including antibacterial resistance (AMR), and oxidative stress–associated diseases. Despite the availability of inexpensive antibiotics, due to AMR, these antibiotics are no longer effective against serious infections[2]. Globally, it was estimated that about 1.27 million deaths in 2019 were attributed to infection with AMR bacteria[3]. In early 2024, the World Health Organization (WHO) released a list of bacterial pathogens to be prioritized in research and development, and public health management to combat AMR[4]. This includes the multi-drug resistant ESKAPE pathogens: Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Pseudomonas aeruginosa, Acinetobacter baumannii, and Enterobacter species. In addition, the bacterium Mycobacterium tuberculosis, a causative agent of tuberculosis disease, is also a significant clinical disease burden due to its ability to develop AMR. In 2022, tuberculosis caused 1.3 million deaths, making it the top infectious disease killer globally[5]. Furthermore, oxidative stress is another major problem, as it is associated with several kinds of diseases, such as reduced immune response to infections, neurodegenerative disorders[6], cardiovascular disease, chronic pulmonary disease, chronic kidney disease, and cancer[7].
Natural products have long played a critical role in addressing antibacterial and antioxidant diseases. The discovery of penicillin in 1928 exemplifies how fungi have yielded transformative medicines[8]. Over the decades, several fungal secondary metabolites or mycochemicals derived from fungi have been reported with biological activities including anticancer, antibacterial, anti-inflammatory, and antiarthritic activities[9]. Mycochemicals can include a wide range of secondary metabolites produced by fungi or mushrooms, including terpenoids, lipids, polysaccharides, and proteins[10]. Recently, there has been a growing interest in studies of fungi residing in plant tissues, known as endophytic fungi, due to their genetic diversity, which makes them a rich reservoir for the discovery of novel biologically active secondary metabolites[11].
Endophytic fungi are known to produce secondary metabolites that help their host plants defend against herbivores and parasites, stimulate nutrient metabolism, and adapt to the environment[12]. This phenomenon is thought to result from plant-fungi symbiosis, leading to mechanisms such as horizontal gene transfer, co-evolution, shared biosynthetic gene clusters, and chemical crosstalk[12,13]. Hence, in exploring sources for biologically active compounds, medicinal plants were often targeted for isolation of biologically active secondary metabolites from endophytic fungi[14].
To further expand the sourcing of novel and biologically active compounds from endophytic fungi, the OSMAC approach has been employed in several studies[15]. This technique is conceptualized and referred to as 'one strain, many compounds' (OSMAC), based on the principle that a single microbial strain has the potential to produce multiple compounds depending on specific growth conditions[15]. This approach utilizes the defense mechanisms of endophytic fungi under competitive environments. Under stress, endophytic fungi release secondary metabolites which vary according to the necessity of their survival[15,16]. Hence, modifications in the composition of the growth medium, pH levels, temperature, the inclusion of enzyme inhibitors, oxygen availability, or the type of culture vessel can result in the production of different sets of secondary metabolites[16].
Recent efforts have underscored the antibacterial potential and diversity of secondary metabolites derived from the endophytic fungi of Annonaceae species[17]. The Annonaceae, or the custard family, predominantly thrives in tropical and subtropical regions, encompasses around 135 genera and 2,500 species[18]. Their medicinal properties include antimicrobial activities such as antibacterial, antimalarial, leishmanicidal, antiviral, trypanocidal, and antiprotozoal, as well as other uses such as antitumor, anticholinesterase, antioxidant, among others[19]. These medicinal properties are attributed to phytochemicals produced by the plants, including flavonoids, alkaloids, acetogenins, and essential oil constituents[19].
The Philippines has been recognized as having a very rich diversity that hosts about 13,500 plant species, of which 1,500 have been reported to have medicinal usage[20]. Alkaloids, highly oxygenated cyclohexenoids, phenylpropanoids, and flavonoids isolated from several genera of Philippine Annonaceae, such as Uvaria[21], Goniothalamus[22], Phaeanthus[23], and Drepananthus[24] have shown potent antibacterial, antitubercular, cytotoxic, antidiabetic, and anti-obesity activities. Despite this rich biodiversity, efforts to explore the biological activity of endophytic fungi extracts remain at an early stage. While there have been reports of the biological activities of endophytic fungi derived from Annonaceae species, none have focused so far on the endophytic fungi of endemic Annonaceae. By investigating these hosts, the present study highlights an untapped reservoir of microbial diversity with potential novel biologically active compounds. Among the Annonaceae species that were selected many have been recognized for their biologically active plant extracts and traditional medicinal uses. For instance, plant extracts from Dasymaschalon clusiflorum var. megalanthum Merr. have demonstrated antimicrobial properties[25], while Platymitra arborea (Blanco) Kessler has shown anticancer potential[26]. Meanwhile, as of the current publication date, no reports exist regarding the bioactivity or traditional use of Orophea cumingiana S. Vidal and Artabotrys cagayanensis Merr.
To combat the plight against AMR of ESKAPE pathogens and oxidative stress associated diseases, this study explored the antibiotic and antioxidant potential of secondary metabolites derived from the endophytic fungi of four Philippine endemic Annonaceae plants: O. cumingiana, A. cagayanensis, P. arborea, and D. clusiflorum var. megalanthum. This study represents the first report documenting both the identity and biological activity of endophytic fungi from Philippine endemic Annonaceae, underscoring their value in natural product-based drug discovery, and their potential contribution to addressing antimicrobial resistance and oxidative stress-related diseases.
-
The reagents used were all analytical grade, including culture media, bacterial strains, and antibiotics. Potato dextrose agar (PDA), agar-agar powder, Mueller Hinton Broth (MHB), and malt extract powder were purchased from TMMedia (TMMedia, Rajasthan, India). Ampicillin, streptomycin, ciprofloxacin, amikacin, resazurin, D-mannitol, D-glucose, cottonseed flour, lactalbumin, and sodium tartrate were procured from Sigma-Aldrich (Sigma-Aldrich, Saint Louis, MO, USA). Yeast extract powder was purchased from Himedia (Himedia, Mumbai, India). Sucrose (saccharose) and potassium phosphate dibasic were obtained from Techno Pharmachem (Techno Pharmachem, Bahadurgarh, India). Glycerol, calcium carbonate, and sodium sulfate were acquired from Loba Chemie Pvt. Ltd. (Mumbai, India). Ammonium sulfate was obtained from Mallinckrodt (Mallinckrodt, Staines-upon-Thames, United Kingdom). Molasses, brown rice, and oatmeal were acquired from a local supermarket in the Philippines. The organic solvents used for extracting secondary metabolites, ethyl acetate (EtOAc), and methanol (MeOH), were also of analytical grade and procured from RCI Labscan (Bangkok, Thailand).
Isolation and fermentation of endophytic fungi
-
Orophea cumingiana S. Vidal, Artabotrys cagayanensis Merr., and Platymitra arborea (Blanco) Kessler were collected at the Northwestern University Ecotourism Park and Botanic Garden (18°9′4.28′′ N, 120°34′12.00′′ E), while Dasymaschalon clusiflorum var. megalanthum was collected from Kalbario Patapat Natural Park (18°30′19.51′′ N, 120°54′27.50′′ E) in September 2023. Voucher specimens were deposited in the Herbarium of Northwestern Luzon, Philippines (HNUL) for accession numbers. Leaves were sequentially immersed in 70% ethanol (1 min), sterile distilled water, and a final rinse of sterile distilled water. Stems underwent immersion in 70% ethanol (1 min), 3% sodium hypochlorite (NaClO) (10 min), and then 70% ethanol (30 s). Leaves were then cut into five small segments (5 mm × 5 mm), while stems were cut into four to five small portions (5 mm × 5 mm). These segments were individually transferred onto PDA plates (supplemented with 100 mg/L streptomycin sulfate and 100 mg/L ampicillin), and incubated at 25–30 °C for three to four weeks. This procedure was carried out in triplicate for each plant sample. Fungal isolates grown from the plate were purified by inoculating a small piece of fungal colony from the margin onto a new PDA culture medium[27]. The endophytic fungal isolates used in this study were deposited at the UST Collection of Microbial Strains (UST-CMS) for accession numbers.
Selected endophytic fungi were fermented in three different liquid fermentation media such as YM 6.3 (malt extract 10 g/L, D-glucose 4 g/L, yeast extract 4 g/L), ZM ½ (molasses 5 g/L, oatmeal 5 g/L, saccharose 4 g/L, mannitol 4 g/L, D-glucose 1.5 g/L, calcium carbonate 1.5 g/L, lactalbumin 0.5 g/L, ammonium sulfate 0.5 g/L)[20], Q6 ½ (D-glucose 2.5 g/L, glycerol 10 g/L, cotton seed flour 5 g/L)[21], and one solid fermentation medium such as brown rice containing BRFT (yeast extract 1 g/L, sodium tartrate 0.5 g/L, K2HPO4 0.5 g/L)[22]. For the liquid medium, 200 mL of the solution was poured into a 500-mL culture flask, while for the solid medium, 56 g of brown rice and 200 mL of BRFT solution were poured into 500-mL culture flasks. The fermenting cultures were monitored daily for growth and glucose content with the use of glucose strips (Medi-Test, Macherey-Nagel, Düren, Germany).
Extraction of secondary metabolites
-
For cultures subjected to liquid-state fermentation, extraction was performed 72 h after the culture medium tested negative for glucose, indicating the end of the fermentation[28]. The culture medium was filtered through cheesecloth to separate the mycelium from the broth. The mycelium was discarded, and the culture filtrate was transferred into a separatory funnel. Exhaustive liquid-liquid extraction was then carried out by multiple extractions with ethyl acetate (EtOAc)[29]. The obtained EtOAc extract was filtered with anhydrous sodium sulfate (Na2SO4 to ensure the complete removal of any remaining water[29,30]. The EtOAc extract was then dried under reduced pressure using a rotary evaporator. Defatting of the extracts was performed using liquid-liquid extraction between 90% aqueous methanol and n-heptane[31]. The methanol layer was collected, filtered, transferred into a vial, and dried until further use.
For brown rice cultures, the extraction process began by submerging the culture medium in EtOAc overnight, followed by filtration to separate the mycelia from the filtrate. This step was repeated three times. The filtered EtOAc extract was then dried under reduced pressure, and redissolved in 90% MeOH followed by extraction with n-heptane. The resulting aqueous methanol extract was dried using a reduce rotary evaporator[28]. The same procedure was carried out for the fat removal, filtration, and drying of the methanolic layer.
The normality of extract yield across media was assessed using the Shapiro–Wilk test. Further statistical analysis was carried out using the Kruskal–Wallis test to assess differences in secondary metabolite production across culture media, followed by a multiple comparisons test via two-stage linear step-up procedure of Benjamini, Krieger, and Yekutieli (BKY) to identify specific group differences.
Antibacterial screening against ESKAPE pathogens
-
The antibacterial activity of selected methanol crude fungal extracts against ESKAPE bacteria was determined using resazurin microtiter assay (REMA), as described by de Jesús Dzul-Beh et al.[32], with slight modifications. The antibacterial screening was performed in two stages: a primary screening to examine the antibacterial activity of the crude fungal extracts at 1,000 µg/mL, followed by the determination of the minimum inhibitory concentration (MIC) of the positive extracts using the same assay. Extracts with a yield of less than 7 mg were not considered for antibacterial investigation.
Test microorganisms such as S. aureus (USTCMS1182), P. aeruginosa (USTCMS10013), and K. pneumoniae (USTCMS1042) were acquired from UST-CMS. Other bacterial strains, including A. baumannii (ATCC 19606), antibiotic-resistant E. faecium (ATCC 27270), and E. hormaechei (ATCC 700323), were procured from Fil-Anaserve, Inc. An inoculum of the test microorganism from the glycerol stock was transferred into sterile MHB, and incubated at 37 °C. After 18−24 h, the optical density of the MHB culture was measured at 625 nm (OD625) using a UV-Vis spectrophotometer. The MHB culture was then diluted with an appropriate volume of MHB to obtain an absorbance between 0.08 and 0.1, matching the 0.5 McFarland Standard (1.5 x 108 CFU/mL).
The crude fungal extracts were dissolved in pure dimethyl sulfoxide (DMSO) and then diluted with sterile water to reach a working concentration of 6,000 µg/mL in 24% DMSO. For the positive control, 700 µg/mL of amikacin (for E. faecium), and ciprofloxacin (for other ESKAPE bacteria) were prepared in pure DMSO and then diluted with sterile water to reach a working concentration of 168 µg/mL in 24% DMSO. Meanwhile, 24% DMSO in sterile water was prepared as negative control, with a final well concentration of 4% as described in the assay method.
For the preliminary screening, 100 μL of the test culture was dispensed into separate wells of a clear 96-well microtiter plate containing 20 μL of the test samples and controls. The final well concentration of the test samples was 1,000 µg/mL in 4% DMSO. A sterility control containing 120 μL of sterile MHB was also included. All samples and controls were tested in triplicate. Afterward, the plates were sealed and incubated at 37 °C. After 18 h of incubation, 20 μL of 0.02% resazurin solution was added to each well, and incubated at 37°C for another 2−4 h until the negative control showed a color change from blue to pink. In this assay, pink indicated bacterial growth, whereas blue indicated inhibition of bacterial growth. Samples exhibiting blue to dark violet color were considered to have positive antibacterial activity, and were further assayed to determine their MIC.
For the determination of the MIC of active extracts, a two-fold serial dilution of the test samples was initially performed, resulting in a concentration range from 1,000 to 7.8125 µg/mL. Then, the same assay procedure as the preliminary screening was followed for the incubation of samples and controls with the test culture. The lowest concentration of the extract that exhibited a blue color was recorded as the MIC of the sample.
DPPH assay for radical scavenging activity
-
The active extracts from the anti-ESKAPE screening were further subjected to antioxidant evaluation. Their free-radical scavenging activity was assessed using the DPPH assay, following the method of Bungihan & Matias[33], with slight modifications. Stock solutions of the crude extracts and positive controls (BHT and ascorbic acid) were prepared in MeOH (3 mg/mL). A 0.1 mM DPPH solution was prepared and further diluted (1:10) with MeOH. In a 96-well microplate, 50 µL of each sample stock solution was mixed with 100 µL of the adjusted DPPH solution, resulting in a final concentration of 100 µg/mL per well. The plate was incubated at room temperature for 30 min, after which absorbance was measured at 490 nm using a microplate reader (Promega GloMax® Discover System). The radical scavenging activity (%RSA) was calculated using Eq. (1). Extracts with > 50% RSA at 1,000 µg/mL were further evaluated for dose–response activity using a two-fold serial dilution across eight concentrations (1,000–7.81 µg/mL) to determine IC50 values. For ascorbic acid, a 12-point two-fold serial dilution at a lower concentration range (333–0.16 µg/mL) was prepared for IC50 determination. IC50 values for both extracts and controls were calculated using GraphPad Prism 9.
$ {\text{%}}\; Inhibition=\left[\dfrac{{{{A}_{Control}}-A}_{sample}}{{A}_{Control}}\right]\times 100 $ (1) Microplate Alamar Blue assay (MABA) against Mycobacterium tuberculosis
-
The active extracts from anti-ESKAPE screening were sent to the University of Illinois Chicago (Chicago, USA) for antitubercular activity screening using MABA. Stock solutions of the extracts were prepared in DMSO (2 mg/mL), and tested at a single concentration of 64 µg/mL. Test solutions were prepared in Middlebrook 7H12 medium (7H9 broth supplemented with 0.1% w/v casitone, 5.6 µg/mL palmitic acid, 5 mg/mL bovine serum albumin, and 4 mg/mL catalase, filter-sterilized) in a final volume of 100 µL per well in black 96-well microplates. A 100 µL inoculum of M. tuberculosis H37Rv (2 × 105 CFU/mL) was then added to each well, yielding a final volume of 200 µL. Plates were incubated at 37 °C for 7 d, after which 12.5 µL of 20% Tween 80 and 20 µL of Alamar Blue (Trek Diagnostic, Westlake, OH, USA) were added. Following 16–24 h of further incubation, fluorescence was measured (excitation 530 nm, emission 590 nm). The percentage inhibition of bacterial growth was calculated relative to untreated bacteria-only controls using Eq. (2).
$ {\text{%}}\; Inhibition=\left[\dfrac{{{{F}_{Control}}-F}_{sample}}{{F}_{Control}}\right]\times 100 $ (2) Identification of the antibacterial-producing endophytic fungi
-
From the stock culture, a block of culture was transferred to the sterile PDA plates, and incubated for 7–10 d at 28 °C to observe the culture characteristics of the endophytic fungi. The culture characteristics such as color, surface structure, pattern, margin, growth, and others were observed and analyzed in the PDA plates[24]. The microscopic morphology was carried out by using the transparent adhesive tape mount or tape-lift mount techniques[25].
For the phylogenetic analysis, the endophytic fungi were sent to Macrogen Inc. (Macrogen Inc., Seoul, South Korea) for gDNA extraction, PCR amplification, and bidirectional sequencing of internal transcribed spacer (ITS), forward ITS5 (5'-GGAAGTAAAAGTCGTAACAAGG-3'), and reverse ITS4 (5'-TCCTCCGCTTATTGATATGC-3') region[34]. The chromatogram (AB1 file) of the sequences was analyzed using Molecular Evolutionary Genetics Analysis across Computing Platform (MEGA X) software. The individual forward and reverse sequences were aligned using MUSCLE algorithm, and the contigs generated from the alignment were saved as a FASTA file. For preliminary identification, the BLASTN tool (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ) was used, and sequences with ≥ 99% identity, and 99% query coverage were assigned to the genus level and was the basis for the retrieval of the reference sequencing from the National Center for Biotechnology information website (www.ncbi.nlm.nih.gov )[35]. To construct the phylogenetic tree, the contigs of each endophytic fungus and its corresponding reference sequences were then aligned using MEGA X software. To search for the best-fit model for the phylogenetic analysis, the model with the lowest Bayesian Information Criterion (BIC) was used. Maximum likelihood trees were constructed with 1,000 bootstrap replicates[26,27].Chemical profiling of the active crude fungal extracts
-
Thin-layer chromatography (TLC) was employed to describe the chemical profile of the active crude fungal extracts. The presence of alkaloids, indoles, terpenes, and sterols was investigated using spray reagents of Dragendorff, Van Urk-Salowski, and Vanillin-sulfuric acid, respectively. Lastly, Bornträger reagent was used to detect the presence of anthraquinones, anthrones, and coumarins.
-
Forty-two endophytic fungi were isolated from the leaves and stems of four Annonaceae species in this study. From this, 12 isolates presented with distinct morphology and were investigated for their antibacterial activities (Table 1).
Table 1. Endophytic fungi isolated from the leaves and stems of Philippine endemic Annonaceae species.
Plant code Host plant Plant part Total number of isolates Endophytic fungus code of selected isolates 001 O. cumingiana Leaves 7 NA-001-Lv-C NA-001-Lv-D NA-001-Lv-E NA-001-Lv-F NA-001-Lv-G Stem 7 NA-001-St-B NA-001-St-D NA-001-St-H 002 A. cagayanensis Leaves 6 NA-002-Lv-X Stem 6 NA-002-St-C 003 P. arborea Leaves 3 − Stem 4 NA-003-St-C 017 D. clusiflorum var. megalanthum Leaves 7 KA-017-Lv-A Stem 2 − Total 42 12 − indicates none. Effects of media on fungal growth and secondary metabolite production
-
The mass of secondary metabolites produced by the 12 fungal isolates varied significantly depending on the growth medium (Fig. 1a). Among the four tested media, brown rice (BR) supported the highest yields, ranging from ~40 to 700 mg. In contrast, cultures grown in YM, ZM, and Q6 media produced markedly lower amounts, generally below 50 mg. Statistical analysis confirmed highly significant differences between BR and all other media (**** p < 0.0001). Moreover, comparison among the liquid media (Fig. 1b) revealed significant differences, with Q6 supporting higher metabolite production than ZM and YM (** p < 0.01).
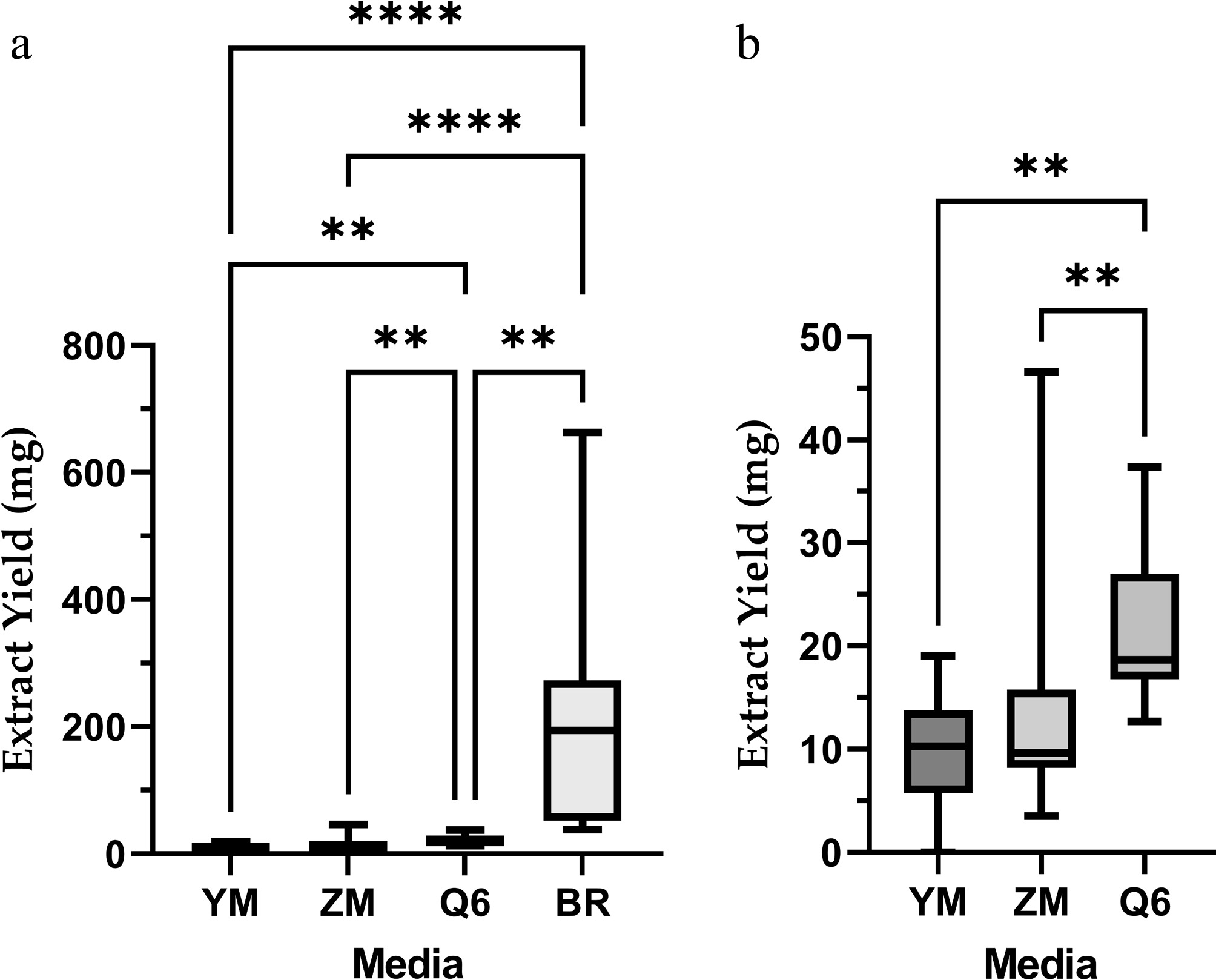
Figure 1.
(a) Mass of secondary metabolites produced by 12 fungal isolates cultivated in different media: YM = yeast malt medium; ZM = ZM ½ media; Q6 = Q6 ½ media; BR = brown rice media. (b) Enlarged view of panel (a) showing YM, ZM, and Q6. Statistical significance is denoted as follows: ** p < 0.01; **** p < 0.0001.
Antibacterial activity of fungal extracts against ESKAPE
-
Of the 48 fungal extracts, six extracts were excluded from the antibacterial screening due to their low yield (less than 7 mg), or absence of mycelium (Table 2). Results from the antibacterial screening revealed 23 extracts with antibacterial activity in at least one ESKAPE pathogen. Notably, BR-001-St-D exhibited with the most promising antibacterial activity and presents high extract yield. This extract exhibited antibacterial activity against five ESKAPE pathogens, with MIC values ranging from 7.81 to 250 µg/mL.
Table 2. Extract yield and minimum inhibitory concentrations (MIC, µg/mL) of selected endophytic fungal isolates of Annonaceae species against ESKAPE pathogens.
Isolate code Extract code Extract yield (mg) MIC (µg/mL) Ef Sa Kp Ab Pa Eh NA-001-Lv-C YM-001-Lv-C-BE 7.3 − − − − − − ZM-001-Lv-C-BE 3.5 u u u u u u Q6-001-Lv-C-BE 37.4 − − − − 1,000 62.5 BR-001-Lv-C 54.3 − 62.5 − − − − NA-001-Lv-D YM-001-Lv-D-BE 5 u u u u u u ZM-001-Lv-D-BE 9.8 − − − 1,000 500 − Q6-001-Lv-D-BE 16.7 − − − − − − BR-001-Lv-D 38.2 − − − − − − NA-001-Lv-E YM-001-Lv-E-BE No growth u u u u u u ZM-001-Lv-E-BE 46.6 − − 7.81 1,000 − − Q6-001-Lv-E-BE 16.9 − − − − − − BR-001-Lv-E 195.7 − − − − − − NA-001-Lv-F YM-001-Lv-F-BE 13.1 − − − − − 250 ZM-001-Lv-F-BE 10.3 − − − − − − Q6-001-Lv-F-BE 18.4 − − − − − − BR-001-Lv-F 193.3 − 62.5 − − − − NA-001-Lv-G YM-001-Lv-G-BE 5.2 u u u u u u ZM-001-Lv-G-BE 9.1 − − − − 250 − Q6-001-Lv-G-BE 17.1 − − − − 500 − BR-001-Lv-G 41.9 − − − − − − NA-001-St-B YM-001-St-B-BE 12.1 − − − − 500 − ZM-001-St-B-BE 9.5 − − − 500 1,000 − Q6-001-St-B-BE 14.7 − − − 1,000 125 − BR-001-St-B 296.2 − − − − 250 − NA-001-St-D YM-001-St-D-BE 14 − 1,000 − 250 1,000 − ZM-001-St-D-BE 17.2 − 1,000 − 7.81 62.5 − Q6-001-St-D-BE 18.9 − − − 500 62.5 − BR-001-St-D 202.3 − 125 15.63 250 7.81 250 NA-001-St-H YM-001-St-H-BE 10.5 − − 15.63 500 1,000 − ZM-001-St-H-BE 8.5 1,000 1,000 7.81 500 − − Q6-001-St-H-BE 23.6 − − − − − − BR-001-St-H 51.4 − − − − − − NA-002-Lv-X YM-002-Lv-X-BE 18.3 1,000 125 − − 1,000 − ZM-002-Lv-X-BE 7.9 − − − − − − Q6-002-Lv-X-BE 12.7 − − − − − − BR-002-Lv-X 122.5 − − − − − − NA-002-St-C YM-002-St-C-BE 7.3 − − − 250 − 500 ZM-002-St-C-BE 8.1 − − − − − − Q6-002-St-C-BE 28.1 − − − − − − BR-002-St-C 195.9 − − − − − − NA-003-St-C YM-003-St-C-BE 28.2 500 62.5 − 125 1,000 1,000 ZM-003-St-C-BE 13.7 − − − 500 500 − Q6-003-St-C-BE 14.5 − 62.5 − 250 1,000 − BR-003-St-C 196.4 − − − − − − KA-017-Lv-A YM-017-Lv-A-BE 19.1 − − − − − − ZM-017-Lv-A-BE 12 − − − − − − Q6-017-Lv-A-BE 21.1 − − − − − − BR-017-Lv-A 663.1 − − − − − − Culture media: YM = yeast malt medium; ZM = ZM ½ media; Q6 = Q6 ½ media; BR = brown rice media. ESKAPE pathogens: Ef = E. faecium; Sa = S. aureus; Kp = K. pneumoniae; Ab = A. baumannii; Pa = P. aeruginosa; Eh = E. hormaechei; − = no antibacterial activity; u = untested; Amikacin (28 µg/mL) was used as positive control for E. faecium and ciprofloxacin (28 µg/mL) was used as positive control for other ESKAPE pathogens. The extract with the most promising antibacterial activity is in bold. Antioxidant activity of fungal extracts
-
Extracts that showed antibacterial activity against ESKAPE were further evaluated for their antioxidant activity. From the 23 anti-ESKAPE active extracts, 18 extracts exhibited antioxidant properties as well, with radical scavenging activity ranging from 50% to 99%, as presented in Table 3. ZM-001-Lv-E was the most potent antioxidant with an IC50 of 56.03 µg/mL, and with %RSA comparable to antioxidant drugs—ascorbic acid and BHT. A dose-response curve was not performed for extracts with less than 50% RSA at 1,000 µg/mL. For extracts with undetermined IC50, a different concentration is recommended to identify their IC50 values.
Table 3. IC50 values (µg/mL), and radical scavenging activity of anti-ESKAPE active extracts (at 1,000 µg/mL), AA, and BHT in DPPH assay.
Extract code IC50 (µg/mL) %RSA ± SD Q6-001-Lv-C-BE − 13.52 ± 2.28 BR-001-Lv-C − 31.26 ± 5.99 ZM-001-Lv-D-BE 102.7 72.31± 0.94 ZM-001-Lv-E-BE 56.03 77.06 ± 0.27 YM-001-Lv-F-BE − 48.77 ± 2.23 BR-001-Lv-F − 28.36 ± 2.92 ZM-001-Lv-G-BE 65.85 68.74 ± 1,74 Q6-001-Lv-G-BE 87.47 58.67 ±2.86 YM-001-St-B-BE ND 54.34 ± 0.58 ZM-001-St-B-BE ND 56.65 ± 0.62 Q6-001-St-B-BE 200.9 58.02 ± 4.49 BR-001-St-B ND 52.04 ± 0.82 YM-001-St-D-BE ND 62 ± 1.67 ZM-001-St-D-BE 133.6 61.46 ± 0.52 Q6-001-St-D-BE 205.2 74.28 ± 1.34 BR-001-St-D 443.6 82.31 ± 1.28 YM-001-St-H-BE 435.7 88.99 ± 2.12 ZM-001-St-H-BE ND 53.16 ± 0.89 YM-002-Lv-X-BE 213.5 98.85 ± 0.23 YM-002-St-C-BE 121 59.85 ± 0.94 YM-003-St-C-BE ND 91.73 ± 1.11 ZM-003-St-C-BE − 11.33 ± 3.31 Q6-003-St-C-BE 63.29 61.32 ± 4.39 AA 1.18 76.79 ± 1.04 BHT 24.57 78.59 ± 0.82 − = no antioxidant activity; ND = not determined; BHT= Butylated hydroxytoluene; AA= Ascorbic acid The extract with the most promising antioxidant activity is in bold. Antitubercular activity of crude fungal extracts
-
In determining the antitubercular activity of the crude fungal extracts, those with antibacterial activity against ESKAPE were screened against M. tb. H37Rv. Promising extracts, which showed at least 90% of growth inhibition of the viral bacteria, were BR-001-Lv-F and YM-002-Lv-X-BE (Table 4). However, some extracts were not screened for antitubercular activity due to the limited amount of samples.
Table 4. Percent inhibition activity of anti-ESKAPE fungal extracts (64 µg/mL) against M. tb. H37Rv in MABA.
Extract code % Inhibition Q6-001-Lv-C-BE 0 BR-001-Lv-C 0 ZM-001-Lv-D-BE 16% ZM-001-Lv-E-BE 24% YM-001-Lv-F-BE 34% BR-001-Lv-F 90% ZM-001-Lv-G-BE NT Q6-001-Lv-G-BE 32% YM-001-St-B-BE 22% ZM-001-St-B-BE 17% Q6-001-St-B-BE NT BR-001-St-B 16% YM-001-St-D-BE 20% ZM-001-St-D-BE 21% Q6-001-St-D-BE 2% BR-001-St-D 0 YM-001-St-H-BE 4% ZM-001-St-H-BE NT YM-002-Lv-X-BE 96% YM-002-St-C-BE NT YM-003-St-C-BE 21% ZM-003-St-C-BE 11% Q6-003-St-C-BE 19% NT= not tested; Most promising extracts are highlighted in bold. Identification of endophytic fungi with antibiotic potential
-
The 11 active endophytic fungal isolates that produced antibacterial extracts against ESKAPE were identified using their morphology (Fig. 2), and phylogenetic tree analysis through MEGA X software. The active endophytic fungi from O. cumingiana effective against ESKAPE pathogens consist of five genera, including Diaporthe (NA-001-Lv-C, NA-001-Lv-F), Fusarium (NA-001-Lv-D, NA-001-Lv-G, NA-001-St-B), Chaetomium (NA-001-Lv-E), Chromolaenicola (NA-001-St-D), and Corynespora (NA-001-St-H). The endophytic fungal isolate NA-001-Lv-C was identified as D. tectonendophytica, showing 99% ML support in the phylogenetic tree. D. tectonendophytica is characterized by white colonies on both the front and reverse sides, with aerial hyphae, a radiate pattern, and an irregular margin. Moreover, the endophytic fungal isolate NA-001-Lv-F was identified as D. longicolla with 98% ML support based on its phylogenetic tree results. It was described as having white colonies with furrowed, zonate, irregular, spreading growth, aerial hyphae, and no reproductive cells. Endophytic fungal isolates NA-001-Lv-D and NA-001-Lv-G were identified as Fusarium species with ML support of 87% and 89%, respectively. Both Fusarium isolates were described as having white colonies on the front and reverse sides, a circular pattern, and a smooth margin. However, NA-001-Lv-D had a larger colony after 7 d, and exhibited chlamydospores and ellipsoidal conidia, while NA-001-Lv-G had a smaller colony and wedge-shaped macroconidia. Both Fusarium species had aerial mycelia. NA-001-St-B was identified as F. decemcellulare with 100% ML, and is described as having red colonies at the center, white colonies at the edges, aerial hyphae, a circular pattern, a smooth margin, and the presence of microconidia.
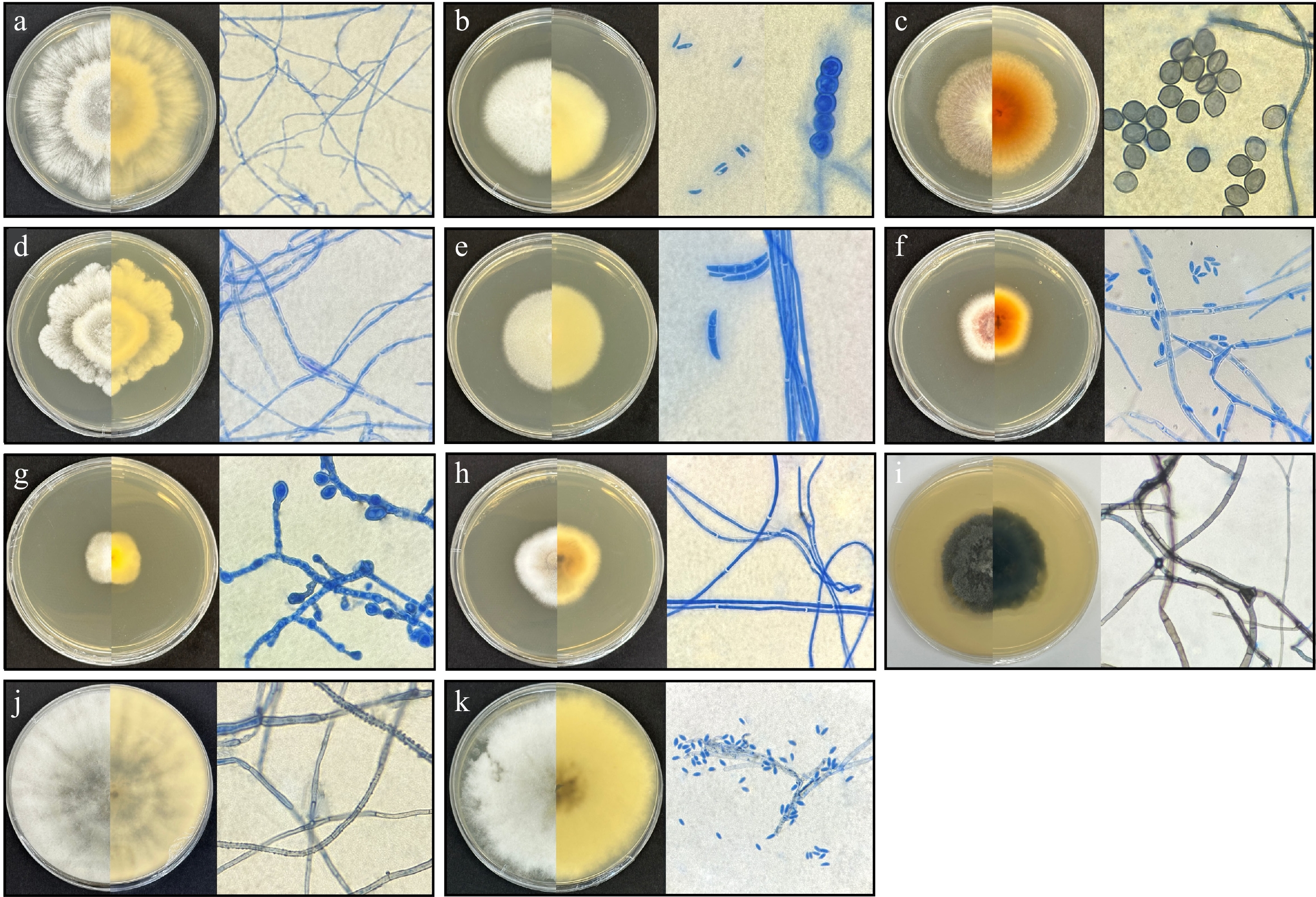
Figure 2.
Culture morphology and microscopic morphology of active endophytic fungi observed under a compound light microscope at 1,000× magnification (oil immersion objective). (a) NA-001-Lv-C. (b) NA-001-Lv-D. (c) NA-001-Lv-E. (d) NA-001-Lv-F. (e) NA-001-Lv-G. (f) NA-001-St-B. (g) NA-001-St-D. (h) NA-001-St-H. (i) NA-002-Lv-X. (j) NA-002-St-C. (k) NA-003-St-C.
Endophytic fungal isolate NA-001-Lv-E, with 66% ML, was identified as Chaetomium sp., characterized by white to brown colonies on the front and yellow to orange colonies on the reverse, aerial hyphae, and limoniform ascospores as reproductive cells. NA-001-St-D was identified as Chromolaenicola sp. with 98% ML support. It was described as having white to pale brown colonies on both the front and reverse sides, growing slowly with an irregular pattern, and containing germinated conidium from the hyphae. NA-001-St-H was identified as C. cassiicola with 98% ML support in the phylogenetic tree. It is characterized by yellowish-gray to white colonies on both the front and reverse sides, featuring a cottony texture and smooth margins, with no reproductive cells present.
The two endophytic fungal isolates from A. cagayanensis were identified as Endomelanconiopsis sp. (NA-002-Lv-X) and Lasiodiplodia sp. (NA-002-St-C). Endomelanconiopsis sp. has only 18% ML support, and is described as having white, dark green to black colonies on both the front and reverse sides, with aerial hyphae, a furrowed, zonate pattern, and an irregular margin. Lasiodiplodia sp. has 65% ML support and is described as having white to gray colonies on both the front and reverse sides, aerial hyphae, a thick cottony surface, a radiate pattern, and a smooth margin. Both endophytic fungal isolates have sterile mycelia, with no reproductive cells present. The endophytic fungal isolate from P. arborea (NA-003-St-C) was identified as D. eschscholtzii with 98% ML support and is described as having white to gray colonies, a raised surface structure, a radiate pattern, an irregular margin, spreading growth, and the presence of ellipsoidal conidia.
Chemical profile of the active extracts
-
Mycochemical screening via TLC was employed to describe the chemical profiles of the active extracts. Extracts with the same fungal isolates showed small variations with respect to the present mycoconstituents (Table 5). Results from the mycochemical analysis of active extracts indicate the presence of anthrones, coumarins, terpenes and sterols, and indoles. Notably, chemical diversity was apparent in the TLC profile of the active extracts (Fig. 3).
Table 5. Mycoconsitutents present in the active crude fungal extracts.
Extract code ALK ANQ ANT C TS IN Q6-001-Lv-C-BE − − − − − − BR-001-Lv-C − − + − ++ ++ ZM-001-Lv-D-BE − − − − − − ZM-001-Lv-E-BE − − ++ − + ++ YM-001-Lv-F-BE − − − − − + BR-001-Lv-F − − + − + + ZM-001-Lv-G-BE − − − − − − Q6-001-Lv-G-BE − − − − − − YM-001-St-B-BE − − − − + − ZM-001-St-B-BE − − − − − − Q6-001-St-B-BE − − − − − − BR-001-St-B − − − − + − YM-001-St-D-BE − − − + − − ZM-001-St-D-BE − − + − − − Q6-001-St-D-BE − − − − − − BR-001-St-D − − ++ − − − BR-001-St-E − − − − − − ZM-001-St-H-BE − − − + − − Q6-001-St-H-BE − − − − − − YM-002-Lv-X-BE − − + − − − YM-002-St-C-BE − − − + − − YM-003-St-C-BE − − + − + − ZM-003-St-C-BE − − + − + − Q6-003-St-C-BE − − + − + − ALK = alkaloids; ANQ = Anthraquinones; ANT = Anthrones; C = Coumarins; TS = Terpenes and sterols; IN = indoles; + indicates the relative abundance of certain mycoconstituent; − = absence; The most promising extract is highlighted in bold. 
Figure 3.
TLC profile of active crude fungal extracts. The TLC plate was viewed under (a) visible light, and (b) 365 nm using 7:2 EtOAc-MeOH solvent system and ethanol sulfuric acid spray reagent for derivatization. Active extracts were separated per fungal isolates by solid lines.
-
In the present study, 48 fungal extracts were isolated from four Annonaceae species. From these, 12 distinct fungal species were selected and cultivated in four different media (YM 6.3, ZM ½, Q6 ½, and BRFT), which produced 23 crude fungal extracts with antibacterial activity against ESKAPE pathogens (Table 2). The potency of the active crude extracts was classified based on their MIC values following the criteria established by de Jesús Dzul-Beh et al., wherein weak activity was defined as MIC 1,000–500 µg/mL, moderate activity as 500–250 µg/mL, and strong activity as ≤ 125 µg/mL[32]. Most of the extracts were found to be active against P. aeruginosa, A. baumanni, and S. aureus, with varying range of antibacterial strength from weak to strong. Additionally, five extracts were found active against E. hormaechei with a varying antibacterial strength (weak to strong) while three extracts were revealed to have weak antibacterial activity against E. faecium.
Because previous studies have demonstrated overlap between antioxidant and antibacterial mechanisms[36−38], the active anti-ESKAPE extracts were further evaluated for antioxidant and antitubercular properties. In the DPPH assay, most anti-ESKAPE extracts also displayed antioxidant activity. The dual activity is consistent with reports highlighting the role of reactive oxygen species (ROS) in antibacterial mechanisms, particularly in mediating bacterial cell death[36,38]. For instance, May et al. emphasized the repurposing potential of antioxidant drugs such as Auranofin and Ebselen, which inhibit thioredoxin reductase in M. tuberculosis and S. aureus[39]. Thioredoxin reductase is a critical enzyme for bacterial redox balance, as many bacteria lack glutathione synthase, and rely instead on the thioredoxin system to counteract ROS from host immunity or antibiotic stress. Similarly, Naqvi et al. showed evidence showing that natural product–derived antioxidants can act as 'natural antibiotics', with antibacterial mechanisms including impairment of membrane integrity, disruption of nucleic acid synthesis, and inhibition of bacterial respiration[38]. In fact, in the study by Bag & Chattopadhyay, synergistic interactions on antibacterial and antioxidant efficacy of essential oils were observed[40].
For targeting ESKAPE pathogens, the most promising extract was BR-001-St-D, which conferred strong to moderate antibacterial activity against five ESKAPE pathogens (MIC = 250−7.81 µg/mL). BR-001-St-D was isolated from Chromolaenicola sp. However, since the present identification was based solely on ITS sequencing, we were only able to identify this fungal isolate up to its genus level. Notably, all extracts in our studies derived from Chromolaenicola sp. demonstrated inhibitory activity against S. aureus, K. pneumoniae, A. baumannii, and P. aeruginosa. This aligns with a recent report in which three Chromolaenicola species (C. chiangraiensis MFLUCC 24-0058, C. siamensis MFLUCC 24-0057, C. thailandensis MFLUCC 24-0056) were shown to possess antibacterial and antifungal activity against Bacillus subtilis and Mucor plumbeus[41].
Of particular interest, the present extract was found to contain anthraquinones, a class of compounds well-documented for their antibacterial potential. In a study by Vidal et al., an anthraquinone compound—1,8-dihydroxyanthraquinone (danthron)—was isolated from a marine Chromolaenicola sp. (HL-114-33-R04)[42]. While danthron was initially characterized by its antioxidant and angiogenic properties[43], a study by Wei et al. has demonstrated its direct antibacterial effects[44]. Specifically, danthron was shown to exert potent activity against S. aureus through cell membrane disruption, leading to loss of bacterial viability[44].
For targeting tuberculosis, the most promising extracts were YM-002-Lv-X-BE and YM-001-Lv-F, derived from Endomelanconiopsis sp. and D. longicolla, respectively. Chemical profiling revealed the presence of terpenes and sterols, anthraquinones, and indoles in these extracts. Previous studies have established the antibacterial potential of compounds isolated from D. longicolla[45,46]. For instance, Liu et al. reported nine biologically active metabolites from this fungus, including a novel terpene, Longidacid A, which exhibited antitubercular activity by inhibiting an M. tb. protein enzyme at 50 µM[45]. Similarly, extracts of Endomelanconiopsis sp. have been documented for antibacterial activity against S. aureus and antifungal activity against Candida albicans[47], as well as antioxidant properties[48]. In a study by Pushpavathi & Krishnamurthy, dereplication of the ethyl acetate extract of Endomelanconiopsis endophytica identified several antibacterial metabolites, including Butylparaben, Sakuranetin, Tenuazonic acid, and 3,4-dimethoxycinnamic acid[48]. Notably, 3,4-dimethoxycinnamic acid, a phenylpropanoid derivative and a common precursor of coumarins, has been shown to inhibit bacterial filament formation by targeting the FtsZ protein, ultimately leading to impaired cell division and proliferation[49]. However, to date, we are the first to report antitubercular activity from this genus.
Other noteworthy antibacterial fungal isolates identified in this study include C. cassiicola, D. tectonendophytica, and F. decemcellulare. Extracts from D. tectonendophytica (MIC = 1,000–62.5 μg/mL) and F. decemcellulare (MIC = 1,000–125 μg/mL) remain largely underexplored, making this the first report to demonstrate their antibacterial potential. Previous work by Gagana et al., showed that ethyl acetate extracts of C. cassiicola were active against Salmonella typhi but inactive against K. pneumoniae, P. aeruginosa, and S. aureus[50]. In contrast, the present findings revealed that C. cassiicola grown in ZM ½ and Q6 ½ media exhibited strong activity against K. pneumoniae, as well as weak activity against E. faecium, S. aureus, A. baumannii, and P. aeruginosa. This discrepancy may stem from differences in culture media, since Gagana et al. used potato dextrose broth. Such variation in media composition likely triggered the biosynthesis of distinct secondary metabolites in the present active extracts, which would not have been produced under potato dextrose broth. Importantly, this underscores the value of the OSMAC strategy in enhancing the chemical diversity of fungal metabolites. Accordingly, chemical profiling of the active extracts (Fig. 3, Table 5) showed that even extracts derived from the same fungal isolate displayed notable differences in metabolite composition.
Focusing on the novelty of the present findings, Chromolaenicola sp., Endomelanconiopsis sp., and D. longicolla, are reported here for the first time as endophytic fungi associated with endemic Philippine Annonaceae. Chromolaenicola sp. has previously been isolated from various plant families, including Asteraceae, Bromeliaceae, Fabaceae, Ranunculaceae, and Spindaceae[41], and the marine-derived Chromolaenicola sp. HL-114-33-R04, which has been noted for its antioxidant properties[42]. However, there have been no prior reports of Chromolaenicola sp. isolated from Annonaceae, specifically from the endemic Philippine plant O. cumingiana. Similarly, this study is the first to report D. longicolla that was isolated from O. cumingiana. Although this endophytic fungus has also been reported from Cananga odorata (Annonaceae)[51], Glycine max (Fabaceae), and Pyrus sp. (Rosaceae)[52]. The endophytic fungus Endomelanconiopsis sp. has been documented in multiple plant families, including Malvaceae, Olacaceae, Ochnaceae[53], Fagaceae[54], and Lauraceae[55], but we are the first to record Endomelanconiopsis sp. isolated from A. cagayanensis.
In addition to revealing antibacterial potential, the present study highlights the importance of culture medium selection in optimizing both metabolite diversity and yield. Among the media tested, brown rice proved particularly effective in providing the highest crude extract yields (Fig. 1). Brown rice is rich in phenolics, flavonoids, γ-oryzanol, vitamins, and minerals that serve as biosynthetic precursors or inducers for fungal metabolism[56]. Under solid-state fermentation, these nutrients are more readily available, stimulating enhanced fungal growth and secondary metabolite production[57]. Moreover, an increase in production of biologically active compounds such as phenolic acids, flavonoids, and coumaric acids was observed in several fungi after fermentation with brown rice[57]. Previous studies have consistently shown that solid substrates such as rice outperform liquid broths, sometimes producing tenfold to hundredfold higher yields. For instance, VanderMolen et al. compared six liquid broths and five solid-state substrates and reported that, across three fungal strains, solid media—particularly rice—yielded significantly greater metabolite output[58].
It is noteworthy, however, that higher extract yields in brown rice did not necessarily correspond to stronger antibacterial activity. In several cases, extracts from liquid media, although produced in smaller quantities, exhibited greater potency. This suggests that liquid media may induce the biosynthesis of unique or more biologically active metabolites absent in brown rice cultures. For example, Hewage et al. demonstrated that Dothidiomycetes sp. CR17 produced azaphilone derivatives, tricyclic polyketides, and austidol in potato dextrose broth, while in Czapek malt medium, the same fungus produced four novel polyketides, 4-hydroxybenzaldehyde, and austidol[59]. Such findings emphasize that the choice of medium not only influences metabolite yield but also determines the chemical diversity and bioactivity of the compounds produced. Accordingly, further purification and profiling of the present extracts are necessary to determine the optimal culture conditions for specific biologically active metabolites.
-
This study highlights the untapped potential of extracts from endophytic fungi isolated from Philippine endemic Annonaceae species as sources of novel antibacterial and antioxidant compounds. It is the first to report the antibacterial potential of fungal endophyte extracts from O. cumingiana, A. cagayanensis, P. arborea, and D. clusiflorum var. megalanthum, which are sources of extracts with diverse bioactivities. These findings underscore the long-term value of endophytic fungi from the Annonaceae family as sources of biologically active compounds that could help combat antibacterial-resistant bacterial diseases and oxidative stress-related disorders, which are global health problems that remain unresolved due to limited resources for lead compound discovery. The identified endophytic fungi of notable importance include Chromolaenicola sp., with anti-ESKAPE activities; Chaetomium sp., with antioxidant properties; and Diaporthe longicolla and Endomelanconiopsis sp., with anti-tuberculosis activity. Furthermore, although brown rice as a medium could result in a high yield of extract, it does not guarantee bioactivity, as certain media may only produce compounds with bioactivity. Further purification of the active extracts or dereplication is recommended to determine the identity of biologically active compounds and their mechanism of action. Moreover, species-level identification of the fungi is recommended to determine which endophytic fungal species hold the greatest potential for the treatment of various diseases.
-
The authors confirm their contributions to the paper as follows: conceptualization, supervision, project administration and writing—review and editing: Macabeo AP, Bungihan ME; methodology and validation: Macabeo AP, Bungihan ME, Fajardo MT, Mendoza G, Arturo HC, Suyo LR; formal analysis and investigation: Fajardo MT, Mendoza G, Arturo HC, Suyo LR; data curation: Mendoza G, Arturo HC; writing—original draft preparation: Fajardo MT, Mendoza G; visualization: Bungihan ME. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and are available from the corresponding author on reasonable request.
-
This study was funded by the Department of Science and Technology–Grants-in-Aid (DOST-GIA) through the National Research Council of the Philippines (NRCP). We acknowledge Michael A Calaramo and the Northwestern University Ecotourism Park and Botanic Gardens (NUEBG) for their contribution to the collection and identification of Philippine endemic Annonaceae plants used in this study. We also extend our gratitude to the University of Santo Tomas Collection of Microbial Strains (UST-CMS) for providing the bacterial cultures.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Fajardo MT, Mendoza G, Arturo HC, Suyo LR, Macabeo AP, et al. 2026. Endophytic fungi associated with Philippine endemic Annonaceae species yield extracts with anti-ESKAPE, antitubercular, and antioxidant activities. Studies in Fungi 11: e007 doi: 10.48130/sif-0026-0005 

Endophytic fungi associated with Philippine endemic Annonaceae species yield extracts with anti-ESKAPE, antitubercular, and antioxidant activities
- Received: 15 April 2025
- Revised: 02 January 2026
- Accepted: 19 January 2026
- Published online: 05 March 2026
Abstract: Rising cases of antibacterial resistance and oxidative stress-related disorders prompted the search for new compounds. In this study, endophytic fungi isolated from Philippine endemic Annonaceae plants were investigated as potential sources of antibiotic and antioxidant mycochemicals. Twelve isolates were cultured in four media (YM 6.3, ZM ½, Q6 ½, BRFT), extracted with ethyl acetate and methanol, yielding 48 crude extracts. Antibacterial activity was assessed using REMA for ESKAPE pathogens, and MABA MIC90 for Mycobacterium tuberculosis H37Rv, while antioxidant potential was determined via the DPPH assay. Twenty-three extracts were found to be inhibitory against ESKAPE pathogens, two against M. tuberculosis H37Rv, and 18 demonstrated free-radical scavenging properties. Morphological characterization and ITS-based phylogenetic analysis with MEGA X identified 11 distinct endophytic fungi as the sources of these bioactive extracts. Notably, the BRFT extract from Chromolaenicola sp. exhibited strong anti-ESKAPE activity (MIC = 250–7.81 µg/mL), while BRFT extracts from Diaporthe longicolla and Endomelanconiopsis sp. inhibited M. tuberculosis H37Rv by 90% and 96% at 64 µg/mL, respectively. In addition, the ZM extract from Chaetomium sp. (IC50 = 56.03 µg/mL) exhibited radical scavenging activity comparable to BHT (IC50 = 24.57 µg/mL), and ascorbic acid (IC50 = 1.18 µg/mL). Among the tested media, BRFT supported the highest metabolite production. Mycochemical screening revealed anthrones, terpenes, sterols, and indoles as major constituents. This study provides the first documentation of Chromolaenicola sp. and D. longicolla from O. cumingiana and Endomelanconiopsis sp. from A. cagayanensis, representing novel records of these genera in Annonaceae, and as promising sources of novel antibiotics and antioxidants.
-
Key words:
- Antibiotic /
- Antioxidant /
- ESKAPE /
- Mycobacterium tuberculosis H37Rv /
- Fungal secondary metabolites /
- Natural product