








-
Lecanora is the second richest and cosmopolitan genus of lichenized fungi that is classified in the family Lecanoraceae (Lecanorales, Lecanoromycetes)[1]. The genus is characterized by a crustose thallus, apothecial ascomata with generally reddish-brown discs, the presence of calcium oxalate crystals in the amphithecium, atranorin in the grayish white to gray thallus, Lecanora-type asci that are commonly eight-spored but rarely multispored, and the production ofcolorless, almost ellipsoid to broadly ellipsoid ascospores[2−4]. Vegetative reproduction is also commonly found with the presence of soredia containing atranorin, oxalate crystals in the amphithecium, and filiform conidia[3,4]. The genus represents one of the largest and morphologically most diverse groups of lichen-forming fungi, with approximately 1,000 described species globally[5] and ~250 accepted species in Hyde et al.[1], widely distributed from the Arctic and Antarctic to subtropical and tropical regions[4,6−11]. Members of the genus are primarily crustose lichens, commonly found colonizing bark, wood, rocks, and soil in all ecosystems[4,7,9,12−15]. In addition, many species produce a wide range of secondary metabolites such as atranorin, usnic acid, and various depsides and depsidones, which are important in chemotaxonomic studies and are utilized for their antioxidant, antiviral, antimicrobial, and cytotoxic properties[16−19].
Lecanora was established by Erik Acharius in Luyken[20] and subsequently typified by L. subfusca. Lecanora species are traditionally recognized by their lecanorine apothecia, which are characterized by a thalline margin that includes algal cells and asci containing hyaline, nonseptate ascospores[2,21]. Traditionally, the core genus Lecanora accommodated a large majority of species in the family and thus became a taxonomic repository for crustose lichens with lecanorine apothecia. Over the last two centuries, the genus was defined in a broad sense on the basis of a few stable morphological characteristics, particularly the presence of a thalline apothecial margin and simple ascospores. This traditional circumscription led to Lecanora becoming a "catch-all" genus for numerous unrelated species that superficially resembled each other, contributing to taxonomic inflation and artificial groupings[21]. However, Eigler[22] demonstrated the heterogeneity of the genus and segregated the taxon into several groups according to the microscopic characteristics of the ascomata. In subsequent studies based on ascus types, many Lecanora taxa were reclassified in different genera, families, and subclasses[2,23−27].
Recent advances in molecular phylogenetics have confirmed that Lecanora in its traditional sense is polyphyletic[2,9]. Multilocus analyses using the nuclear ribosomal internal transcribed spacer (ITS), mitochondrial small subunit (mtSSU), and protein-coding genes such as rpb1 and rpb2 have revealed that many species previously placed in Lecanora are more closely related to other genera. This has led to the segregation of several lineages into distinct genera, including Lecanoropsis, Myriolecis, Palicella, Protoparmeliopsis, Pulvinora, and Vainionora[2,26,27−31]. Despite these advances, taxonomic delimitation within Lecanora remains problematic in certain species complexes, where morphological convergence and cryptic speciation are commonly found. Ongoing integrative taxonomic approaches combining morphology, chemistry, ecology, and multilocus phylogeny are essential for resolving these issues and achieving a stable classification[32,33].
In this study, we introduce a new taxon, Lecanora rhododendri, collected from Yunnan, China, based on its morphology and molecular analyses coupled with preliminary chemical test (5% KOH and Lugol's reagent). The topology based on a combined ITS and mtSSU dataset revealed that Lecanora rhododendri sp. nov. formed a separate branch and clustered with L. flavoviridis, L. darlingiae, L. alboflavida, and L. orientoafricana within Lecanora s. str.
-
The sample was collected from dead and dying branches of Rhododendron in May 2019 in Yunnan, China. The sample was observed under a Motic SMZ-171 stereo microscope (Motic Xiamen, China) and a compound microscope (Nikon ECLIPSE 80i, Japan). Measurements of its morphological features (viz. the apothecia, amphithecia, amphithecial cortex, hypothecium, parathecium, hymenium, epihymenium, paraphyses, asci, and ascospores) were made by using the Tarosoft® Image FrameWork program. Figures were processed with Adobe Photoshop CS6 Extended v.10.0 software (Adobe Systems, USA). The herbarium material was deposited in the Herbarium of Cryptogams, Kunming Institute of Botany Academia Sinica (KUN-HKAS), Kunming, China, and duplicated in the Herbarium of Mae Fah Luang University (Herb. MFLU). Faces of Fungi[34] and Index Fungorum numbers[35] were obtained for the novel taxon.
DNA extraction, polymerase chain reaction amplification, and sequencing
-
The DNA was extracted from fruiting bodies using the E.Z.N.A. Forensic DNA Kit (OMEGA® biotek). Polymerase chain reaction (PCR) was used to amplify specific gene regions: ITS was amplified using the primers ITS5 and ITS4[36], and mtSSU was amplified using the primers mtSSU1 and mtSSU3R[37]. The amplification reactions were performed in 25-μL final volumes containing 8.5 μL of sterilized ddH2O, 12.5 μL of Easy Taq PCR Super Mix, 1 μL of each forward and reverse primer, and 2 μL of the DNA template. The PCR thermal cycle program for ITS and mtSSU gene amplification was 94 °C for 3 min initially, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 50 s, elongation at 72 °C for 1.30 min, and a final extension at 72 °C for 10 min. The PCR products were purified and sequenced with the same primers at Sangon Biotech, Shanghai, China. The chromatograms of the sequences were checked to ensure the quality of the sequences using BioEdit v.5[38]. DNA Star v. 5.1 was used to make consensus sequences.
Phylogenetic analyses
-
The newly generated ITS and mtSSU sequences were subjected to a nucleotide BLAST search (
https://blast.ncbi.nlm.nih.gov/Blast.cgi , accessed on 10 August 2025) to identify closely related taxa of the new collection. The sequence data of Lecanora species and closely related taxa in the family Lecanoraceae were downloaded from NCBI's GenBank (Table 1) following recent publications[3,4,9,11]. Downloaded sequences were aligned with the sequences obtained in the current study using MAFFT v. 7.511 (http://mafft.cbrc.jp/alignment/server )[39]. If necessary, single gene alignments were manually improved with BioEdit v.5[38], and the combined ITS and mtSSU sequence dataset was analyzed using the maximum likelihood (ML) and maximum parsimony (MP) criteria.Table 1. Taxa used in the phylogenetic analyses and their GenBank accession numbers.
Taxa Voucher details Country GenBank accession no. ITS mtSSU Glaucomaria carpinea Herbarium Malíček 14828 Czech Republic OQ717895 − Glaucomaria carpinea Kondratyuk S. 21337 (KW-L) Ukraine MK672827 MK693683 Glaucomaria rupicola KRAM: Flakus 29527 Bolivia OL604012 OL604094 Glaucomaria rupicola Leavitt 18-460c BRY-C USA MZ243629 − Japewia aliphatica J. Maliček 9764 Czech Republic MN547352 MN547343 Japewia tornoensis Printzen s. n. Canada HQ650656 HQ660559 Lecanora albella J. Maliček 7336 Czech Republic KY548048 KY502423 Lecanora alboflavida 0 Coppins s. n. United Kingdom KY548045 KY502428 Lecanora alboflavida 1 KRAM: Flakus 29124 United Kingdom OL604044 OL604124 Lecanora alboflavida KRAM: Flakus 29143 Bolivia OL604042 OL604122 Lecanora alboflavida KRAM: Rodriguez-Flakus 3997 Bolivia OL604034 OL604116 Lecanora allophana 0 J. Malíček 9491 (hb. JM) Finland KY548051 KY502416 Lecanora allophana 1 J. Malíček 9626 (hb. JM) Russia KY548050 KY502421 Lecanora argentata J. Malíček 1963 Czech Republic KT630245 KT630264 Lecanora barkmaniana Maliček 10602 Russia MK778605 MK778530 Lecanora caesiorubella F: Lumbsch 19094a USA − JQ782666 Lecanora campestris Arup U225 Sweden AF159930 − Lecanora cenisia J. Maliček 5869 Austria KY548047 KY502425 Lecanora crystalliniformis Wang et al. 19-63013 China ON807164 ON807171 Lecanora crystalliniformis Wang et al. 17-56082 China − ON807172 Lecanora crystalliniformis Wang et al. 17-56085 China ON807163 ON807173 Lecanora crystalliniformis Wang et al. 17-56554 China ON807162 − Lecanora crystalliniformis Tang & Lui L09 China − ON807174 Lecanora darlingiae Lendemer 46644 USA − MH481360 Lecanora expersa J. Maliček 9624 Russia KY548053 KY502420 Lecanora flavidomarginata Flakus 29951 Bolivia OL604056 OL604135 Lecanora flavidomarginata Flakus 28943 Bolivia OL603996 OL604077 Lecanora flavoviridis Papong 6539 Thailand JQ782711 JQ782675 Lecanora fulvastra Flakus 26717 Bolivia − OL604089 Lecanora gangaleoides HTL19923a USA MG554660 JQ782676 Lecanora glabrata Lubeck & Kukwa 17811 Poland MN387101 − Lecanora helva Papong 6444 Thailand JQ782716 JQ782679 Lecanora horiza n/a Spain KT453772 KT453821 Lecanora hybocarpa H.T. Lumbsch s. n. Spain EF105412 EF105417 Lecanora intricata KRAM: Flakus 29565b Bolivia OL604030 OL604112 Lecanora intumescens J. Malíček 8480 (hb. JM) Czech Republic KY548040 KY502441 Lecanora intumescens TRH L-19015 Norway OQ945715 − Lecanora masana JL46185 USA MK573928 − Lecanora orientoafricana P. Kirika 2205 Kenya JQ900619 JQ900617 Lecanora phaeocardia Papong 3492 (F) Thailand JQ782724 JQ782687 Lecanora pulicaris 0 J. Malíček 10262 (hb. Malíček) Russia MK778611 MK778539 Lecanora pulicaris 1 J. Malíček 10263 (hb. Malíček Russia MK778612 MK778540 Lecanora rhododendria MFLU 20-0045a Chinaa PV241604a PV470111a Lecanora rugosella Arup L97561 Sweden AY398712 − Lecanora subimmersa HTL19103b Australia JQ782733 JQ782697 Lecanora subrugosa Arup L98188 Austria AY398711 − Lecanora substerilis Maliček 202 Slovakia KT630243 KT630254 Lecanora tropica F: Papong 6440 Thailand JN943720 JQ782699 Lecanora vainioi Papong 7120 Thailand JN943716 JQ782702 Lecanora variolascens Maliček 8422 Austria KY548038 KY502445 Lecanora zeorina Wang et al. 19-63051 China ON807167 ON807168 Lecanora zeorina Wang et al. 19-63070 China ON807166 ON807169 Lecanora zeorina Wang et al. 19-62893 China ON807165 ON807170 Lecanoropsis saligna 491_DE Germany MT938982 MT939209 Lecanoropsis subintricata 604_JP Japan MT939010 MT939239 Lecidella carpathica ZX 20140367-2 China KT453741 KT453831 Lecidella meiococca Ekman 3101 (BG) Sweden AF517929 AY300893 Palicella glaucopa 0 P. Rodriguez Flakus 2539 (FR) Argentina KJ152486 KJ152471 Palicella glaucopa 1 P. Rodriguez Flakus 2504b (FR) Argentina KJ152485 KJ152472 Parmotrema reticulatum K.H. Moon 14834 Japan KP942516 KP942523 Parmotrema tinctorum TNS-L-Y.O. 5947 Japan KF129464 KF129521 Polyozosia altunica Xahidin 20071910 China MH698406 MH698407 Polyozosia contractula Brodo 31501 (DUKE) USA HQ650604 DQ986898 Polyozosia perpruinosa Arup U176 Sweden AF070025 − Protoparmeliopsis achariana Arup U155 Sweden AF070019 DQ787342 'Protoparmeliopsis crystalliniformis' BDNA-L-0000298 South Korea MW832796 MW832802 'Protoparmeliopsis crystalliniformis' BDNA-L-0000349 South Korea MW832797 MW832803 Protoparmeliopsis garovaglii Leavitt 089 (BRY-C) USA KT453728 KT453818 Protoparmeliopsis garovaglii M. Haji Moniri (KW-L) Iran MK672841 MK693689 Protoparmeliopsis garovaglii 19-63070 China ON807161 ON807175 Protoparmeliopsis garovaglii 19-63467 China ON807160 ON807176 Protoparmeliopsis kopachevskae KoLRI 040276 South Korea MK672847 − Protoparmeliopsis muralis SK 765 Romania KP059048 KP059054 Protoparmeliopsis muralis Schmull s. n. Germany HQ650653 HQ660556 Protoparmeliopsis nashii Knudsen 19030 n/a ON447553 ON367825 Protoparmeliopsis peltata n/a USA KT453722 KT453860 Protoparmeliopsis peltata 14619 Iran ON807159 ON807177 Protoparmeliopsis zareii SK 480 Iran KP059049 KP059055 Pulvinora stereothallina E. A. Davydov no. 14817 & L. S. Yakovchenko (LE-L15134) Russia MW257118 MW257159 Pulvinora stereothallina E. A. Davydov no. 14820 (ALTB) Russia MW257112 MW257152 a The new taxon is indicated in bold. The ex-type strains are indicated by black bold and the unavailable sequence is indicated by "–". The phylogenetic analyses were performed by ML using the RAxML-HPC2 package on XSEDE (8.2.8)[40,41] via the CIPRES Science Gateway platform[42], with the GTR+I+G model of nucleotide substitution, which was selected independently for each locus using MrModeltest 2.3[43]. A rapid bootstrap was set up at 1,000 replicates. The MP analysis was executed with the heuristic search option in PAUP (Phylogenetic Analysis Using Parsimony) v.4.0b10 with the following parameter settings: Characters unordered with equal weight, random taxon addition, branch swapping with the tree bisection–reconnection (TBR) algorithm, and branches collapsing if the maximum length was zero. Alignment gaps were treated as missing characters in the analysis of the combined dataset, where they occurred in relatively conserved regions. Trees were inferred using the heuristic search option with 1,000 random sequence additions, with maxtrees set at 1,000. Descriptive tree statistics were used for parsimony; tree length (TL), the consistency index (CI), the retention index (RI), the relative consistency index (RC), and the homoplasy index (HI) were calculated for the generated trees. Kishino–Hasegawa tests[44] were performed in order to determine whether the trees were significantly different. The ML and MP bootstrap values equal or greater than 60% are given above each node. The phylogenetic trees were visualied in FigTree v.1.4[45] and edited using Microsoft Office PowerPoint 2010 and Adobe Illustrator CS3 (Adobe Systems Inc., USA). The sequences generated in this study have been deposited in NCBI's GenBank (Table 1).
-
The result of the nucleotide BLAST search showed that the new collection is closely related to Lecanora inversa (voucher KRAM_L_74751) with 90.55% similarity (identities: 498/550 bp with three gaps) in the ITS, and is similar to L. alboflavida (vouchers KRAM : Flakus 29143, KRAM : Flakus 28262, KRAM : Rodriguez-Flakus 3997, KRAM : Flakus 29196, and KRAM : Flakus 29124) and L. inversa (voucher Flavonora inversa KRAM L 74752) with 98.14% similarity (identities: 580/591 bp with one gap) in the mtSSU. Therefore, the phylogenetic tree of 56 representative species (including the new collection) in Lecanoraceae was constructed. The combined sequence alignments comprised 80 taxa, with Japewia aliphatica (voucher J. Maliček 9764) and J. tornoensis (voucher Printzen s. n.) as the outgroup taxa. The dataset comprised 1,697 characters, including gaps (1–753 bp for the ITS, and 754–1,697 bp for the mtSSU). The RAxML analysis yielded the best scoring tree (Fig. 1) with a final ML optimization likelihood value of –18,787.258374. The sequence alignment had 1,055 distinct alignment patterns, with 34.39% undetermined characters or gaps. The estimated base frequencies were A = 0.279342, C = 0.212476, G = 0.231333, T = 0.276849, with the substitution rates AC = 1.226615, AG = 3.167475, AT = 1.707134, CG = 1.553882, CT = 5.510086, and GT = 1.000000. The gamma distribution shape parameter alpha was 0.294643, and the tree length was 5.258968. The MP dataset consisted of 885 constant characters, 157 variable parsimony-uninformative characters, and 655 parsimony-informative characters. Eighteen equally parsimonious trees were generated with a tree length of 3,628 steps (CI = 0.402, RI = 0.675, RC = 0.271, HI = 0.598), of which the first tree was shown to be the most parsimonious tree according to the Kishino–Hasegawa test.
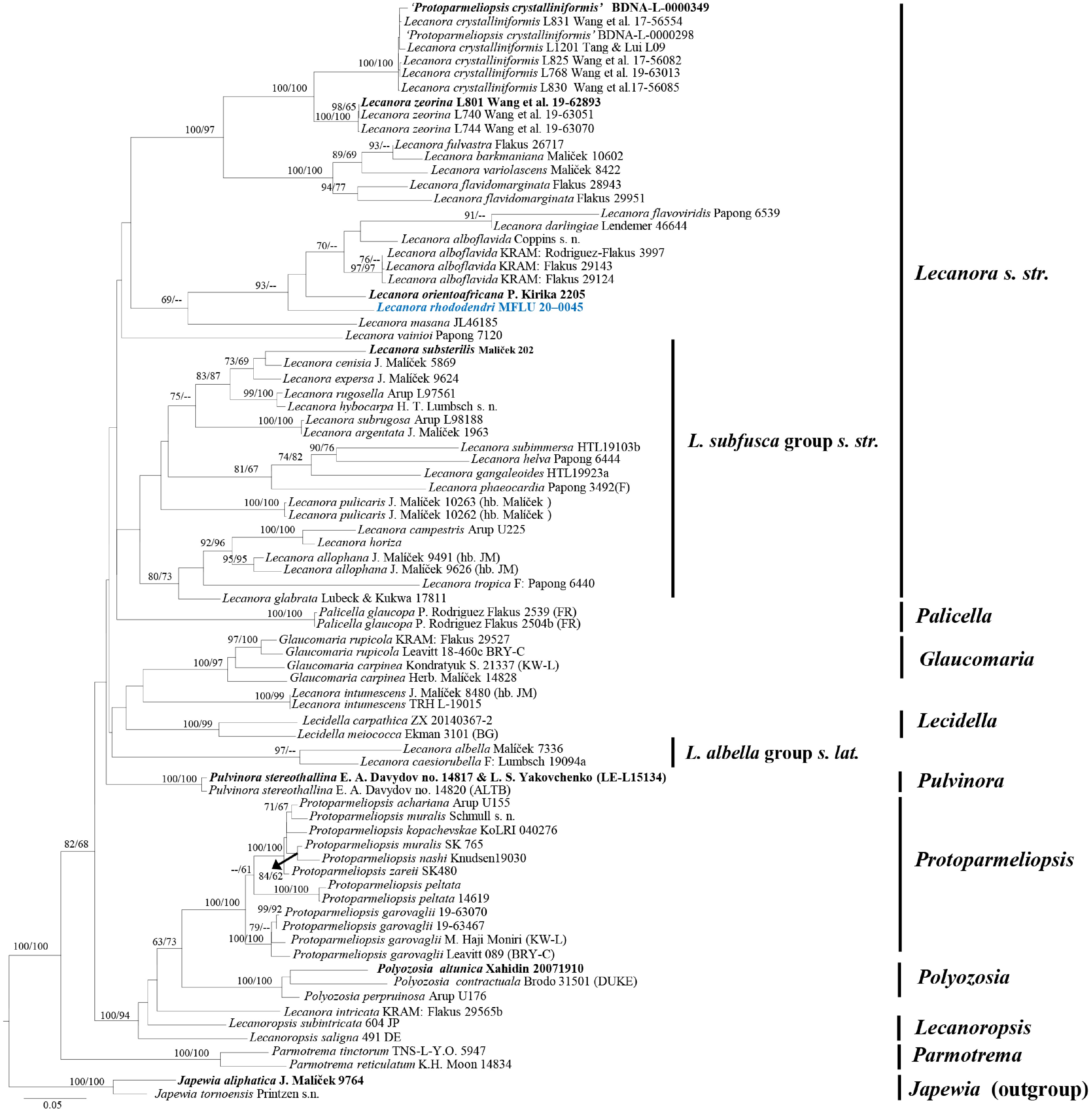
Figure 1.
Phylogram generated from RAxML based on combined ITS and mtSSU sequence data. Bootstrap support values for ML and MP equal to or greater than 60% are indicated as ML/MP at the nodes. The newly generated species in this study is indicated in blue bold text and the ex-type strains are indicated by black bold text. The scale bar represents the expected number of nucleotide substitutions per site.
Phylogenetic trees inferred from the ML and MP analyses showed a similar topology and concurred with previous studies[9]. Most described Lecanora species (such as Lecanora s. str. in Li et al.[9]) clustered within the Lecanora s. str. clade in this study, comprising L. alboflavida (vouchers: Coppins s. n., KRAM: Rodriguez-Flakus 3997, KRAM: Flakus 29143, and KRAM: Flakus 29124), L. barkmaniana (voucher: Maliček 10602), L. crystalliniformis (vouchers: Wang et al. 17-56554, Tang & Lui L09, Wang et al. 17-56082, Wang et al. 19-63013, and Wang et al. 17-56085), L. crystalliniformis (as Protoparmeliopsis crystalliniformis' vouchers: Lee & Kim 2020-000149 and Lee 2020-000098), L. darlingiae (voucher: Lendemer 46644), L. flavidomarginata (vouchers: Flakus 28943 and Flakus 29951), L. flavoviridis (voucher: Papong 6539), L. fulvastra (voucher: Flakus 26717), L. masana (voucher: JL46185), L. orientoafricana (voucher: Kirika 2205 [F]), L. vainioi (voucher: Papong 7120), L. variolascens (voucher: Maliček 8422), L. zeorina (vouchers: Wang et al. 19-62893, Wang et al. 19-63051, and Wang et al. 19-63070). The new species, L. rhododendri (MFLU 20-0045), was also included in this clade and is basal to L. orientoafricana, L. alboflavida, L. darlingiae, and L. flavoviridis.
All species of Lecanora subfusca group s.str. in Li et al.[9] also formed a distinct subclade and was basal of Lecanora s. str. in the present study, comprising L. allophana (vouchers: J. Malíček 9491 [hb. JM] and J. Malíček 9626 [hb. JM]), L. argentata (voucher: J. Malíček 1963), L. campestris (voucher: Arup U225), L. cenisia (voucher: J. Malíček 5869), L. expersa (voucher: J. Malíček 9624), L. gangaleoides (voucher: HTL19923a), L. glabrata (voucher: Lubeck & Kukwa 17811), L. helva (voucher: Papong 6444), L. horiza, L. hybocarpa (voucher: H. T. Lumbsch s. n.), L. phaeocardia (voucher: Papong 3492), L. pulicaris (vouchers: J. Malíček 10263 [hb. Malíček] and J. Malíček 10262 [hb. Malíček]), L. rugosella (voucher: Arup L97561), L. subimmersa (voucher: HTL19103b), L. subrugosa (voucher: Arup L98188), L. substerilis (voucher: Maliček 202), and L. tropica (voucher: F: Papong 6440). However, Lecanora albella group s. lat. viz. L. albella (voucher: Malíček 7336) and L. caesiorubella (voucher: F: Lumbsch 19094a) formed a separate lineage, distant from the Lecanora s. str. clade and clustered with Lecidella, Lecanora intumescens (vouchers: J. Malíček 8480 [hb. JM] and TRH L-19015) and Glaucomaria.
The genus Palicella, namely P. glaucopa (vouchers: P. Rodriguez Flakus 2539 (FR) and P. Rodriguez Flakus 2504b (FR)), constituted an independent lineage and was closely related to Lecanora subfusca group s.str. Other representative genera, namely Glaucomaria, Lecidella, Parmotrema, Polyozosia, Protoparmeliopsis, and Pulvinora, formed well-resolved clades basal to Lecanora and Palicella, except for Lecanoropsis in the present study. Two Lecanoropsis species, namely L. subintricata (voucher: 604 JP) and L. saligna (voucher: 491 DE), formed separate branches, were basal to Lecanora intricata (voucher: KRAM: Flakus 29565b), and clustered with the genera Protoparmeliopsis and Polyozosia with strong support values (100% ML and 94% MP; Fig. 1).
Our new species, Lecanora rhododendri (MFLU 20-0045), formed an independent branch basal to L. orientoafricana, L. alboflavida, L. darlingiae, and L. flavoviridis with significant support in the ML analysis (93% ML) but low support in the MP analysis (32% MP; Fig. 1). These species also clustered with L. masana with 69% support in the ML analysis.
Lecanora rhododendri Chaiwan, sp. nov. (Fig. 2)
Index Fungorum number: IF901032; Faces of Fungi number: FoF17546
Etymology: The epithet "rhododendri" refers to the host genus, Rhododendron, from which the fungus was collected.
Holotype: MFLU 20-0045

Figure 2.
Lecanora rhododendri (MFLU 20-0045, holotype) (a), (b) Appearance of lecanorine apothecia on the host's surface. (c), (d) Section of the apothecia in water. (e) Apothecium stained with 5% KOH. (f) Paraphyses. (g) Close-up of the asci in an apothecium. (h), (i) Asci embedded in the paraphyses. (j) Asci and paraphyses stained with Lugol's reagent. (k), (l) Ascospores. Scale bars: (a) = 1 mm, (b)–(e) = 500 μm, (f)–(j) = 20 μm, (k), (l) = 10 μm.
Associated with dead and dried branches of Rhododendron sp. Thallus crustose, areolate to squamulose, verrucose, thin to thick, whitish; margin distinct; roundish to concave, greenish gray to brownish black. Photobiont trebouxioid. Apothecia 0.6–2.0 mm in diameter, lecanorine, numerous, rounded, dispersed or aggregated, sessile, strictly constricted at the base, rising singly on the areoles; disc black-brown, velvety, in one plane, pruinose; margin inconspicuously merged with the thallus, prominent, thick, smooth, entire to flexuose. Amphithecial cortex uniform, gelatinous, inspersed with crystals, hyaline, 45–63 μm thick, with the hyphae growing out basally. Amphithecium with small to large crystals (= melacarpella-type). Hypothecium hyaline, 25–30 μm high. Parathecium hyaline, with grayish crystals, 5–7 μm thick. Hymenium hyaline, 55–70 μm high, clear, yellowish in KOH. Epihymenium brownish, 10–12 μm thick. Paraphyses septate, sparingly branched, apically slightly swollen at tips, hyaline. Asci 63–85 × 10–14 μm, eight-spored, unitunicate, clavate, Lecanora-type, apically round, with I+ in Lugol's iodine. Ascospores 20–30 × 5–10 μm, hyaline, simple, subglobose to obovoid, aseptate. Asexual morph: Undetermined.
Chemistry: Thallus and apothecial margin K+ yellow in 5% KOH and I+ blue in Lugol's reagent.
Material examined: China, Yunnan, Zhaotong City, 24˚21894' N, 100˚29927' E, 2465 m above sea level on the bark of dead and dried branches of Rhododendron sp., 6 May 2019, H.L. Li & O. Janson, RD-023 (MFLU 20-0045, holotype); ibid., KUN-HKAS 150900 (isotype).
Notes: In the present study, Lecanora rhododendri sp. nov., has a close phylogenetic relationship with L. orientoafricana (voucher P. Kirika 2205) and also clustered with L. alboflavida (vouchers KRAM: Rodriguez-Flakus 3997, KRAM: Flakus 29143, and KRAM: Flakus 29124) and L. masana (voucher JL46185). Morphologically, L. rhododendri is typical of Lecanora in having a crustose, areolate, whitish thallus with an indeterminate, smooth to irregularly wrinkled margin; sessile, lecanorine apothecia with black-brown, velvety, planar, pruinose discs, with small to large crystals of amphithecium; eight-spored Lecanora-type asci; and hyaline, subglobose to obovoid, aseptate ascospores[2,12]. Lecanora orientoafricana differs from the new species in having smaller apothecia (0.4–1.4 mm in diameter vs. 0.6–2.0 mm), with light red-brown to brown, matt, planar or concave, sparsely grayish pruinose discs, smaller asci (50–60 × 10–12 vs. 63–85 × 10–14 μm), and smaller ascospores (12.5–15.5 × 6.0–8.5 µm vs. 20–30 × 5–10)[46]. Lecanora alboflavida can be distinguished from the new species in having a thick, smooth to more often postulate, continuous, minutely granular-verrucose or minutely areolate, sorediate, greenish-gray to gray thallus, and rarely producing apothecia, asci and ascospores; if present, immature apothecia are scattered, planar on the surface of the thallus, circular or distorted, with a thick margin, with planar, subpruinose, light brown discs[3,47]. Unfortunately, there are only a few reports of L. alboflavida. A comprehensive morphological study was carried out by Laundon[47], who synonymized the species under the genus Ochrolechia. Lecanora masana has a grayish-white, epruinose thallus without sorediate or isidia; lecanorine, sessile apothecia with yellow to yellow-green discs; an amphithecium without crystals; eigh-spored, clavate, Lecanora-type asci; and broadly ellipsoid ascospores[48].
Lecanora orientoafricana was reported growing on bark in a degraded montane forest dominated by Podocarpus falcatus, Rapanea melanophloes, and Faurea saligna at an altitude of 3,240 m in Kenya[46]. Lecanora alboflavida is rarely known, with very rare oceanic species, mostly occurring on the acidic bark of old trees from the British Isles, Norway, France, and Macaronesia[3,49]; however, the type specimen (Taylor's saxicolous specimen from 1836) was described on transition rocks[3,47]. Lecanora masana is endemic to high elevations in the southern Appalachian Mountains and is narrowly distributed in the southeastern United States[50]. The species was reported in different habitats, including spruce–fir forests, northern hardwood forests, and shrub balds, growing on the bark of Abies fraseri, Betula spp., Picea rubens, and ericaceous shrubs[50]. However, our new species occurred on the bark of dead and dried branches of Rhododendron sp. at high altitudes (2,465 m above sea level) in Yunnan, China, where the forests are dominated by Rhododendron species.
The ITS nucleotide pairwise comparison revealed that Lecanora rhododendri (MFLU 20-0045) differs from L. orientoafricana (voucher P. Kirika 2205, ex-type) in 57/550 bp with three gaps (10.36%), L. alboflavida (voucher KRAM : Flakus 29124) in 55/515 bp with three gaps (10.68%), L. alboflavida (voucher Coppins s. n.) in 65/553 bp with four gaps (11.75%), and L. masana (voucher JL46185) in 100/550 bp with seven gaps (18.18%). The mtSSU nucleotide pairwise comparison revealed that the new species differs from L. orientoafricana (voucher P. Kirika 2205, ex-type) in 22/606 bp with 6 gaps, L. alboflavida (voucher KRAM : Flakus 29124) in 91/866 bp with 49 gaps (10.51%), and L. alboflavida (voucher Coppins s. n.) in 76/828 bp with 49 gaps (9.19%). On the basis of its morphological distinctiveness, phylogenetic analyses, and the nucleotide-based pairwise comparison, L. rhododendri is introduced here as a new species.
-
The discovery of Lecanora rhododendri sp. nov. enhances the recognition of cryptic and previously overlooked species diversity within this complex genus. As highlighted in recent studies, traditional morphological classifications have often underestimated the number of distinct lineages because of convergent features such as thalline apothecial margins and simple ascospores[2,32]. The combination of detailed morphological assessment and molecular phylogenetic analysis is therefore essential to accurately delimit species boundaries in Lecanora[33,51]. Using the phylogenetic results coupled with the morphological characteristics, the chemical study of KOH and Lugol's reagent test, and DNA polymorphism, we propose a new species in this study, which improves our current understanding of Lecanora fungi. The novelty of L. rhododendri could be justified on the basis of a combination of diagnostic morphological features, including the thallus, ascospore dimensions, and pigmentation of the apothecial disc. These distinctions are consistent across multiple collections and supported by clear phylogenetic separation from the related species L. alboflavida, L. masana, and L. orientoafricana[2,28,46].
Lecanora rhododendri sp. nov. is characterized by sessile, lecanorine apothecia with planar, pruinose discs; small to large crystals of the amphithecium; hyaline hypothecia lacking chlorinated xanthones; eight-spored, Lecanora-type asci; and hyaline, aseptate ascospores. Unfortunately, mature ascospores of L. rhododendri are rarely found; only a few immature ascospores could be observed in the present study. Most species within the genus Lecanora exhibit an amyloid reaction at the ascal apex, characterized by a distinct blue coloration in Lugol's iodine (I+), particularly in species possessing the Lecanora-type ascus. This positive reaction serves as a key diagnostic feature and is valuable for confirming generic or species-level placement. The presence or absence of this reaction can also aid in distinguishing Lecanora from morphologically similar taxa that lack an amyloid response. The ascal apex of L. rhododendri exhibits an amyloid reaction (I+), showing a positive response when tested with Lugol's iodine. Additionally, the thallus of Lecanora species exhibits a KOH reaction, showing a positive response when gelatinous distinctly delimited from the medulla and contains only crystals (granules) which are soluble in KOH but remain gelatinous[52,53]. A preliminary chemical study showed that the thallus and apothecial margin of L. rhododendri turned to K+ yellow in 5% KOH and I+ blue in Lugol's reagent. These diagnostic features are in agreement with typical members of Lecanora s. str. However, additional morphological and chemical studies are necessary to fully understand the circumscription of species in this group.
The monophyly of Lecanora rhododendri placed the species in Lecanora s. str. Lecanora rhododendri formed a stable lineage as a sister to L. orientoafricana (voucher P. Kirika 2205) and clustered with L. alboflavida (vouchers Coppins s. n., KRAM: Rodriguez-Flakus 3997, KRAM: Flakus 29143, and KRAM: Flakus 29124) and L. masana (voucher JL46185), as well as L. darlingiae (voucher Lendemer 46644) and L. flavoviridis (voucher Papong 6539). This phylogenetic result concurred with teh finding of Li et al.[9,54] that these sorediate species (L. alboflavida, L. darlingiae, and L. orientoafricana) and a species with a dark hypothecium (L. flavoviridis) always clustered into a monophyletic clade. Notably, L. alboflavida (vouchers Coppins s. n.) formed a distinct branch with other collections of L. alboflavida and clustered with L. darlingiae (voucher Lendemer 46644) and L. flavoviridis (voucher Papong 6539) in the present study; the intraspecific variation of L. alboflavida (vouchers Coppins s. n.) needs to be clarified, pending further study. Unfortunately, the phylogenetic position of many Lecanora species and their allied genera in Lecanoraceae could not be resolved in the present study, concuring with other previous studies that Lecanora is highly polyphyletic and its phylogeny at higher taxonomic levels remained unresolved[2,3,55,56]. This unstable phylogenetic position might have occurred because of the phylogenetic markers (ITS and mtSSU) and the numbers of available taxa on the tree. Members of the Lecanora group also strongly demand future taxonomic changes based on multilocus genetic data (e.g., ITS, mtSSU, rpb1, and rpb2).
Lecanora species can thrive in extreme conditions, such as higher altitudes, dry areas, and low-nutrient environments. Their ability to survive in these areas allows them to occupy ecological niches that would otherwise be unavailable to other organisms[28,52,53]. In terms of its ecology and distribution, the new species, L. rhododendri, appears to have a restricted habitat preference, being confined to a high elevation (2,465 m above sea level) on corticolous Rhododendron sp. in Yunnan, China. This niche specificity may contribute to its previous misidentification or exclusion from broader surveys, particularly those focused on more common lowland taxa. The biogeographical pattern observed here reflects similar trends reported for narrowly endemic lichens in mountainous or isolated environments[57,58]. Approximately, about 119 Lecanora species have been reported from China thus far[9,54,59−70], distributed across low to high elevation gradients (altitudes of 500–4,500 m). Among them, 25 species were classified within Lecanora s. str. However, these species lack the molecular data required for a comprehensive systematic revision to clarify their placement in Lecanora[9].
In this study, Lecanora rhododendri is introduced. It was found at an altitude of 2,465 m in forests dominated by Rhododendron sp. in Yunnan, China. We propose the recognition of this taxon as a new species within Lecanora, based on phylogenetic evidence, its morphological characteristics, and DNA polymorphism. Its discovery underscores the importance of continued taxonomic exploration in understudied habitats and highlights the role of integrative approaches in revealing hidden fungal diversity.
-
The authors confirm their contributions to the paper as follows: conceptualization, data curation, and formal analysis: Chaiwan N, Phookamsak R, Jiang H, Thiyagaraja V; funding acquisition: Promputtha I, Phookamsak R; investigation, methodology, and writing − original draft: Chaiwan N, Phookamsak R, Jiang H, Thiyagaraja V; project administration: Chaiwan N, Phookamsak R; supervision: Promputtha I, Phookamsak R; writing − review and editing: Phookamsak R, Jiang H, Promputtha I, Bhat J, Dawoud TM. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article. Sequences generated in this study were submitted to GenBank.
-
Napalai Chaiwan would like to thank the Chiang Mai University (CMU) Proactive Researcher grant of Chiang Mai University (Grant No. EX010059) and Chiang Mai University. The authors thank Li Huili and Owen Janson for their help with collecting the specimens. We also thank Shaun Pennycook from Manaaki Whenua - Landcare Research for his assistance in naming the new species. Prof. David L Hawksworth from Plant and Fungal Comparative Biology, Royal Botanic Gardens, Kew is thanked for his assistance in finding literature in Lecanora masana; Hongbo Jiang appreciates the Postdoctoral Directional Training Foundation of Yunnan Province (Grant No. E33O38E261) under Yunnan Provincial Department of Human Resources and Social Security, Yunnan, China, and the 1st batch of the national (Chinese) postdoctoral overseas attraction program in 2023 from Ministry of Education of China (MOE). Rungtiwa Phookamsak sincerely acknowledges the Talent Introduction Plan of Kunming Institute of Botany, Chinese Academy of Sciences, Yunnan Revitalization Talent Support Program 'Young Talent' Project (Grant No. YNWR-QNBJ-2020-120), Yunnan Revitalization Talent Support Program: High-end Foreign Expert Project (Grant No. XDYC-GDWZ-2024-0016), Independent Research of Department of Economic Plants and Biotechnology, Yunnan Key Laboratory for Wild Plant Resources, Kunming Institute of Botany, Chinese Academy of Sciences (Grant No. Y537731261), Yunnan Provincial Department of Human Resources and Social Security, Yunnan Province Foreign Expert Project (Grant No. 202505AO120002) and Yunnan Intelligence Union Program for Young Scientists, Yunnan, China (Grant No. 202503AM140005). Jayarama D. Bhat and Turki M. Dawoud gratefully acknowledge the financial support under the Distinguished Scientist Fellowship Programme (DSFP), King Saud University, Riyadh, Saudi Arabia.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chaiwan N, Phookamsak R, Jiang H, Thiyagaraja V, Bhat J, et al. 2026. Lecanora rhododendri sp. nov. (Lecanoraceae, Lecanorales) associated with Rhododendron sp. in Yunnan, China. Studies in Fungi 11: e008 doi: 10.48130/sif-0026-0006 

Lecanora rhododendri sp. nov. (Lecanoraceae, Lecanorales) associated with Rhododendron sp. in Yunnan, China
- Received: 08 September 2025
- Revised: 08 January 2026
- Accepted: 19 January 2026
- Published online: 13 March 2026
Abstract: During a survey of fungi on Rhododendron in China, Lecanora rhododendri sp. nov., was identified. In this paper, it is introduced with a detailed morphological description and illustrations. The species' novelty is justified on the basis of its distinct morphological characteristics and phylogenetic analyses of a concatenated internal transcribed spacer (ITS) and mitochondrial small subunit (mtSSU) sequence data using maximum likelihood and maximum parsimony criteria, as well as ITS and mtSSU nucleotide base comparisons with related taxa. A preliminary chemical test with 5% KOH and Lugol's reagent on the thallus, apothecial margin, and asci was also observed. The fungus was collected from dead branches of Rhododendron sp. in Yunnan, China. In the phylogenetic analyses of a combined ITS and mtSSU sequence dataset, the species formed a separate branch and has a close relationship with Lecanora orientoafricana in the genus Lecanora (Lecanoraceae, Lecanorales). Morphologically, the new isolate is typical of Lecanora in having a crustose thallus; apothecial, sessile ascomata with black-brown discs; Lecanora-type clavate asci; and hyaline, subglobose to obovoid aseptate ascospores.
-
Key words:
- Lecanoromycetes /
- Lichenized fungi /
- New species /
- Rhododendron inhabiting fungi /
- Taxonomy