









-
The first member of the Tubulin Polymerization Promoting Protein (TPPP) family was identified from bovine brain and named p25[1]. It received its current name when its tubulin-polymerizing and microtubule-stabilizing effects were established[2,3] (for a recent review of TPPP see Diaz et al.[4]). The domain characteristic of TPPP proteins, which covers most of their sequence, is named after the original protein as 'p25alpha' (Pfam05517 or IPR008907). It was also shown that there are two main types of the p25alpha domain, and thus TPPP, a long and a short one[5]. In addition to the different lengths, the two types are characterized by distinctive sequential similarities and differences. The most typical is that the C-terminus of the long TPPP contains a highly conserved region of about 30–32 amino acids, which is absent in the short type[3]. Part of this conserved region is the GxGxGxxGR motif[3]. Long TPPP occurs in practically all animals; the only exception is the placozoan Trichoplax adhaerens[5]. It is present in some phyla of flagellated (!) fungi[6−8], and in other early branching Opisthokonta[9]. There is a very strong correlation between the incidence of TPPP proteins/p25alpha domains and the existence of eukaryotic flagellum/cilium, a microtubule-based organelle[7,10−12]. A different type of TPPP also occurs in fungi, the so-called fungal type TPPP, in which the characteristic C-terminal region is present twice[6] (Fig. 1). Both types are present simultaneously in several fungal species. Very recently, a TPPP that contains not only the C-terminus but the entire p25alpha domain twice has also been identified in a few Opisthokonta species, including a fungal one ('double long TPPP')[9]. The present article systematically examines and reviews the occurrence of different types of TPPPs in the phyla of fungi, discussing the phylogenetic relationships between them.
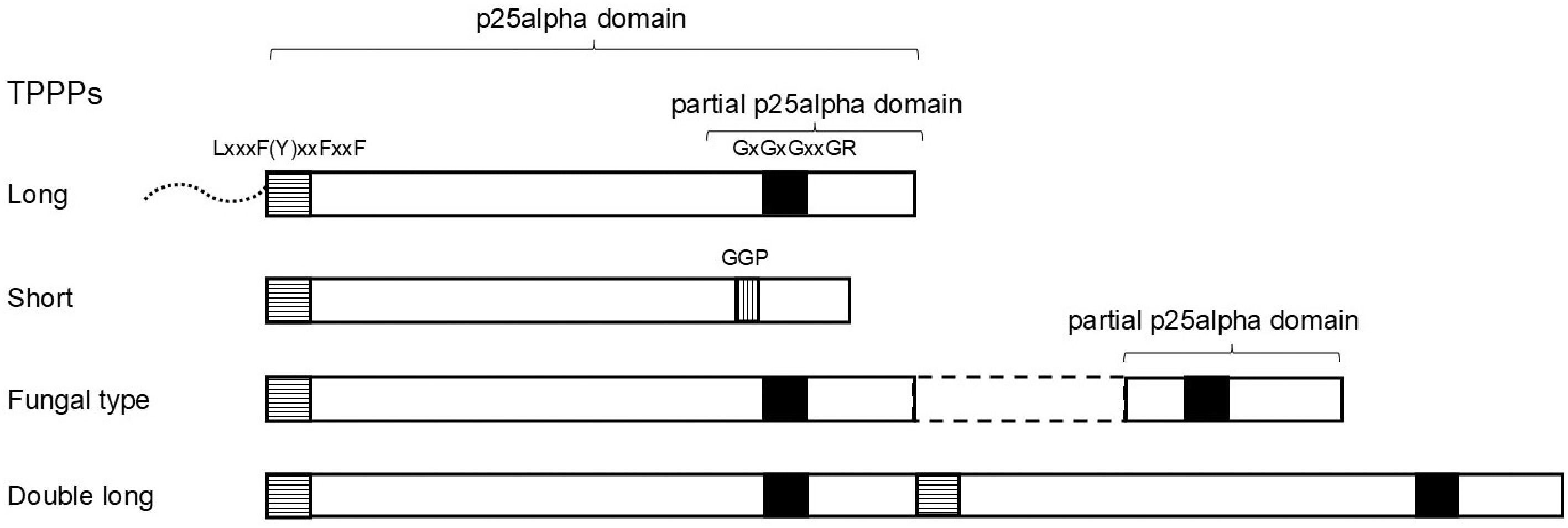
Figure 1.
Schematic structures of various TPPPs. Squares indicate conservative sequences. Dotted line indicates an unstructured N-terminus present in some long TPPPs. Dashed lines indicate a sequence of fungal type TPPP that does not belong to the p25 alpha domains.
-
Accession numbers of protein and nucleotide sequences refer to the National Center for Biotechnology Information (NCBI) GenBank database unless otherwise stated. NCBI Blast was used to search databases (
https://www.ncbi.nlm.nih.gov/BLAST/ )[13]. Whole sequences of various TPPPs were used as queries against databases to find similar sequences in fungi using the BLASTP program. The queries were Chytriomyces hyalinus KAJ3249223, Batrachochytrium dendrobatidis XP_006680205, and Spizellomyces punctatus XP_016604112. If a TPPP was found in a species of a given phylogenetic branch, then its sequence was used as a query within the whole branch. The hits were considered as TPPPs when the BLAST E-value was lower than 10−5, and the query and hit were reciprocal best hits[14,15]. If the hits occurring in a species were at least 95% identical, only one was included in the phylogenetic analysis.Some TPPP sequences of Aphelidiomycota and Sanchytriomycota were retrieved from previous studies[16−18].
Phylogenetic analysis
-
Multiple alignments of sequences (Supplementary Fig. S1) were conducted using the Clustal Omega program[19]. The N-terminal amino acids (fewer than ten) before the first p25alpha domain were trimmed after alignment. Phylogenetic trees were constructed based on the maximum-likelihood (ML) method. The MEGA11 program[20] was used for ML analysis. Bootstrap values were calculated by bootstrap analyses of 500 replicates with the WAG model[21].
-
The BLAST search resulted in numerous TPPPs in all phyla of flagellated fungi, except Neocallimastigomycota, but none were found in later-branching terrestrial phyla. It should be noted that Aphelidiomycota is not included in the fungi in the NCBI databases but is included in fungi according to the latest classification[22,23], so the search was extended to this phylum. Most of the hits were fungal type and long TPPPs. The double long TPPP occurs only in one Chytridiales species (Chytridiales sp. JEL 0842, KAJ3407993). In some Rozellomycota species, there were hits in which proteins contained a large part of the p25alpha domain (approximately the first two-thirds), which accounts for approximately half of their sequences. It is questionable whether these can be considered as TPPPs.
TPPPs of Chytridiales
-
Most of the hits occurred in the Chytridiomycota phylum. It was noticed that species of the Chytridiales, an order of the Chytridiomycota phylum, often have three TPPPs, one long TPPP and two fungal type ones. Therefore, we performed a phylogenetic analysis of the TPPPs belonging to this order. If the proteins occurring in a species were at least 95% identical, only one was included in the analysis. In addition to the genera classified in the order Chytridiales by the NCBI database, we also included those genera whose species belong to this order according to recent studies[24]; as well as those species of the genera (Blyttiomyces, Cladochytrium, Entophlyctis) that clearly belong to another order.
Figure 2 shows the ML phylogenetic tree of TPPPs of the Chytridiales. Species known not to belong to the order Chytridiales are in separate positions: Blyttiomyces helicus, Cladochytrium replicatum, and Entophlyctis helicus. Phlyctochytrium arcticum joins them as a fourth. Within Chytridiales, three clades (two of them with maximal support) are distinguished. Two clades contain fungal type TPPPs; within both, species belonging to the families Chytriomycetaceae and Chytridiaceae are separated. The third clade contains long TPPPs and, in a weakly supported sister position, TPPPs of uncertain classification from Chytridiales.
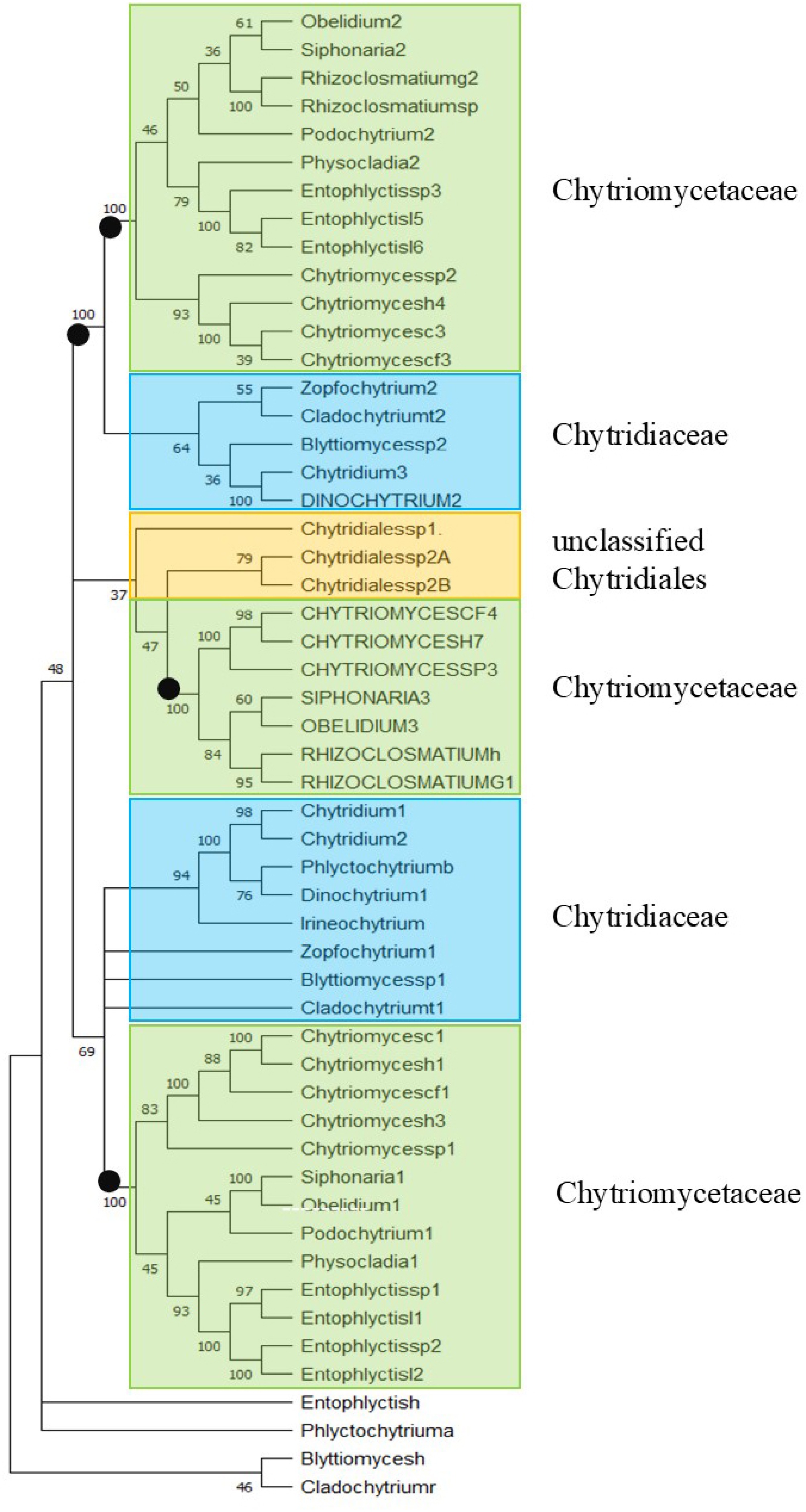
Figure 2.
Phylogenetic tree of TPPPs of Chytridiales by ML analysis. The accession numbers of proteins are listed in Supplementary Table S1. Capital letters indicate long TPPPs. Bootstrap values (500 replicates) above 33% are shown at branch nodes. Branches corresponding to partitions reproduced in less than 33% bootstrap replicates are collapsed. Black circles indicate maximal (100%) bootstrap values at main branches. Boxes represent families. Color code: Chytriomycetaceae (green), Chytridiaceae (blue), unclassified Chytridiales (orange).
Most species belonging to the families Chytriomycetaceae and Chytridiaceae, Chytriomyces, Entophlyctis, Obelidium, Physocladia, Podochytrium, Siphonaria, and Blyttiomyces, Chytridium, Cladochytrium, Dinochytrium, Zopfochytrium, respectively, contain two kinds of fungal-type TPPP. The tree provides information for some genera whose species affiliation was previously disputed (see the Discussion section).
Dinochytrium kinnereticum also belongs to the Chytridiaceae; both TPPPs in it have the corresponding position. Both fungal-type TPPP clades contain a D. kinnereticum protein; however, one of them does not meet the criteria for fungal-type TPPPs, as it lacks the second, partial p25alpha domain. However, the position shown in the tree is indeed correct! D. kinnereticum TPPP (KAJ3212964) shows very high homology to the Chytridium lagenaria KAI8846486 protein, which is in the sister position to it. Comparing the sequences of the two proteins and their corresponding whole genome shotgun (WGS) nucleotide sequences, we see that they show a high degree of identity/similarity throughout, and the second, partial p25alpha domain is also found in the WGS sequence of D. kinnereticum. Thus, either the corresponding hypothetical protein was annotated incorrectly, or a shorter protein was indeed transcribed by alternative splicing (Cf. Supplementary Fig. S2).
TPPPs of Chytridiomycota
-
Figure 3 shows the ML phylogenetic tree of TPPPs of the Chytridiomycota phylum. Clades follow the individual orders: Caulochytridales, Chytridiales, Cladochytriales, Lobulomycetales, Polychytriales, Rhizophlyctidales, Rhizophydiales, Spizellomycetales, Synchytriales. It can also be seen that long and fungal type TPPPs belonging to the same order are in the same clade, although they are distinct within the clade. Chytridiales are found in two different clades. Both clades contain the Chytriomycetaceae and Chytridiaceae families. The Lobulomycetales also form two separate clades. B. helicus + Rhizophlyctiales + Spizellomycetales form a well-supported (bootstrap value 90%) clade within which families are distinct.
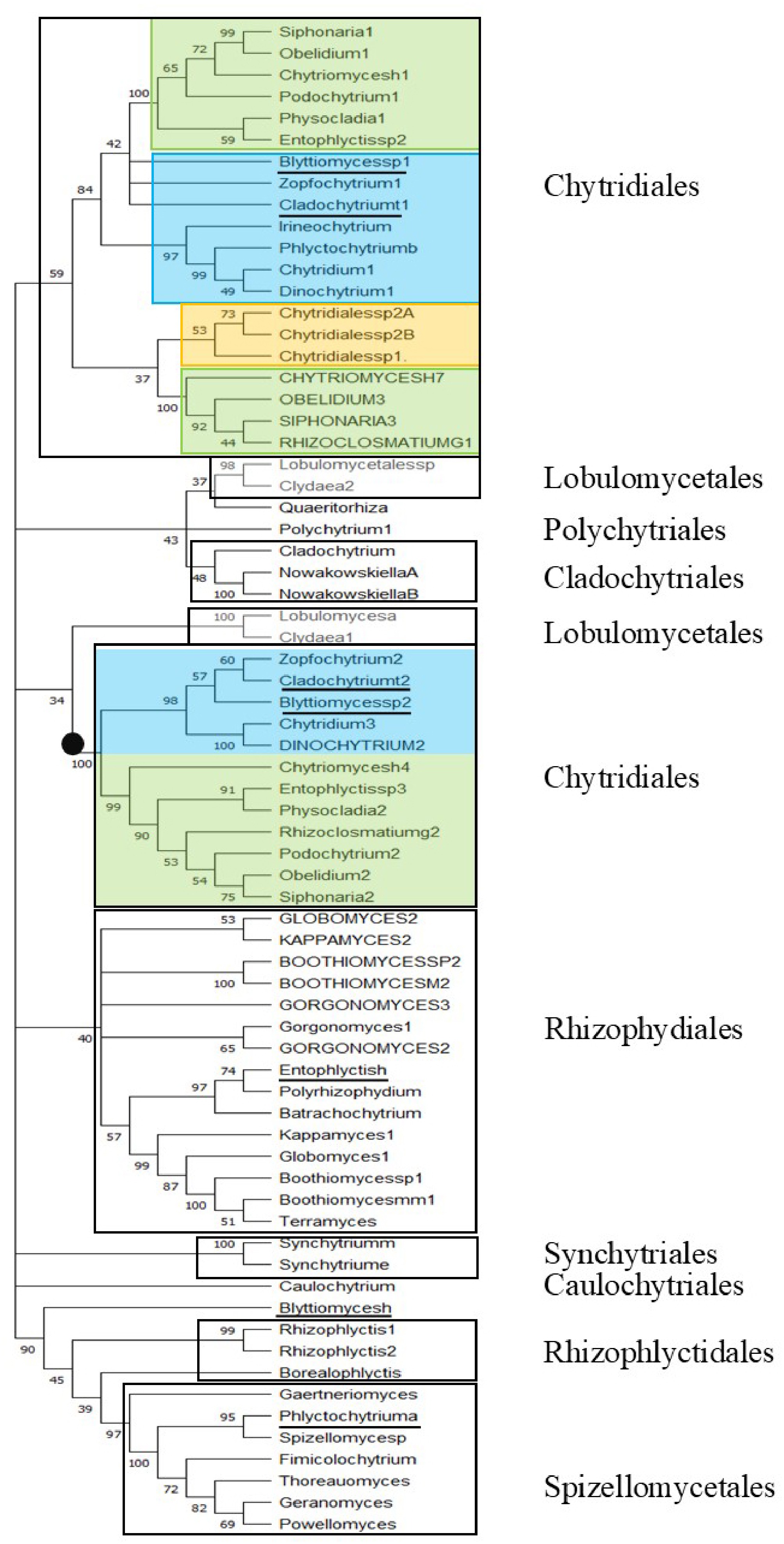
Figure 3.
Phylogenetic tree of TPPPs of Chytridiomycota by ML analysis. The Accession Numbers of proteins are listed in Supplementary Table S1. Capital letters indicate long TPPPs. Underlining indicates species whose taxonomic classification have been disputed. Bootstrap values (500 replicates) above 33% are shown at branch nodes. Branches corresponding to partitions reproduced in less than 33% bootstrap replicates are collapsed. The black circle indicates the maximum (100%) bootstrap value of a main branch. Boxes represent orders. Color code as in Fig. 2.
TPPPs of fungi
-
Figure 4 shows the ML phylogenetic tree of TPPPs of fungi. It includes not only TPPPs found by BLAST search, but also some whose sequences have been published in recent years but are not yet listed in the NCBI database. These are TPPPs of Aphelidiomycota and Sanchytriomycota; the latter one has recently been defined as a new phylum[25]. The Aphelidiomycota species are Paraphelidium tribonematis[26,27], Aphelidium tribonematis, and Aphelidium insulamus[28]; the Sanchytriomycota species are Amoeboradix gromovi and Sanchytrium tribonematis[25,29]. P25alpha-containing proteins of Rozellomycota were not included in the analysis because, on the one hand, they do not contain the entire domain, and on the other hand, the domain only makes up half or even less of their sequence.
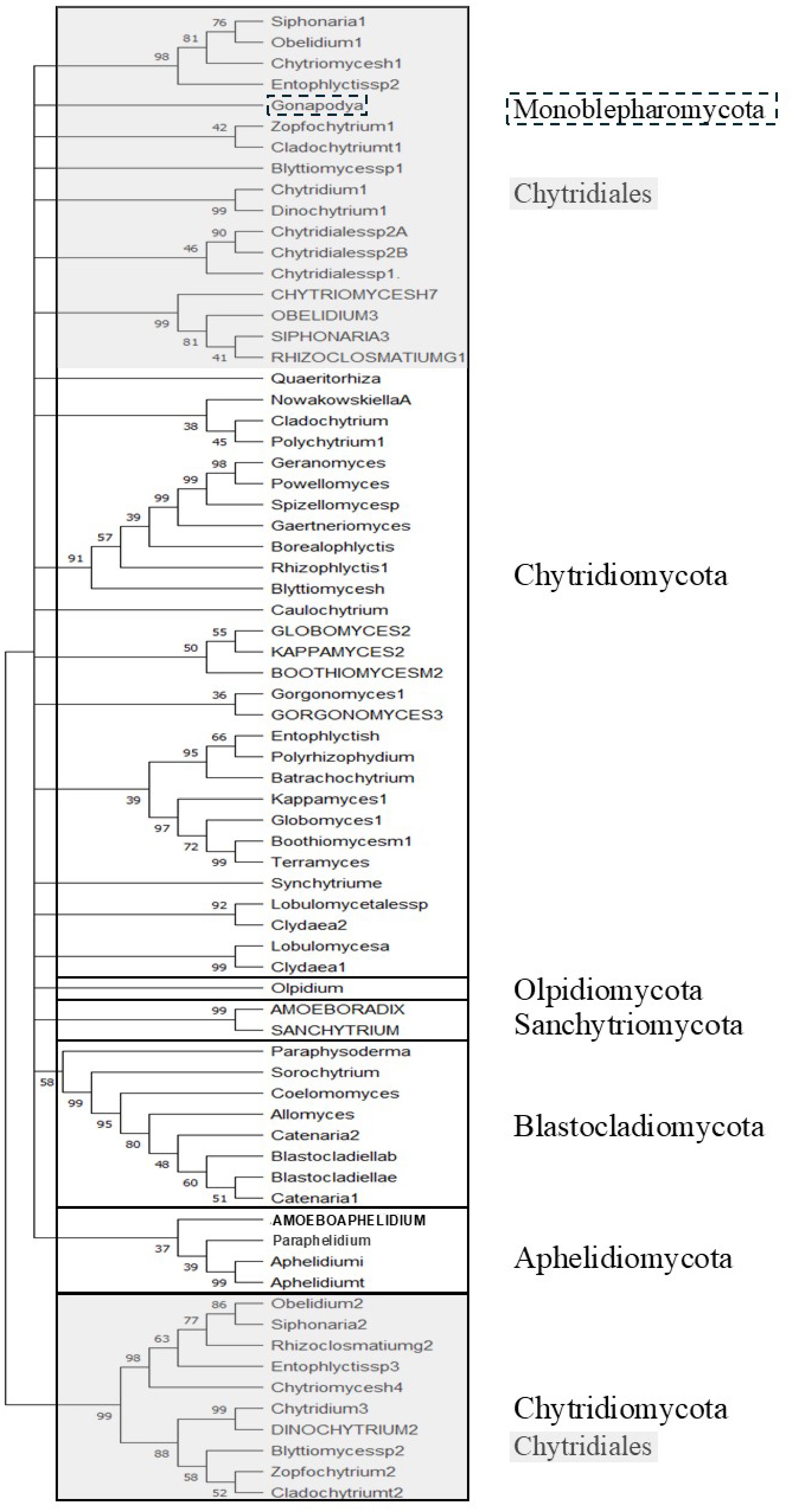
Figure 4.
Phylogenetic tree of TPPPs of fungi by ML analysis. The Accession Numbers of proteins are listed in Supplementary Table S1. Capital letters indicate long TPPPs. Bootstrap values (500 replicates) above 33% are shown at branch nodes. Branches corresponding to partitions reproduced in less than 33% bootstrap replicates are collapsed. Boxes represent phyla. Grey background indicates the Chytridiales order.
The tree corresponds to species phylogeny; the TPPPs belonging to each phylum form well-separated clades: Aphelidiomycota, Chytridiomycota, Sanchytriomycota, Blastocladiomycota, and Olpidiomycota. The exception is the single Monoblepharomycota (Gonapodya), which is wedged among the Chytridiomycota. Here too, we have the second Chytridiomycota clade (with very high bootstrap value), which contains TPPPs belonging to the order Chytridiales that have their 'pair' in the first Chytridiomycota clade. The second Chytridiomycota clade is sister to all the others. Since the choice of root was arbitrary, the tree can also be represented with the Aphelidiomycota at this position. Then the second clade of Chytridiomycota is sister to TPPPs of all the other 'traditional' fungi.
-
Phylogenetic analysis has confirmed that in some cases where the position of certain species was questionable or controversial, the more recently accepted positions are the correct ones, as the positions of the TPPPs also correspond to these. According to Fig. 2, Entophlyctis species belong to the Chytridiales; however, E. helicus does not, as has been shown previously[30,31]. The phylogenetic position of B. helicus is still uncertain; however, Blyttiomyces sp. JEL0837 was placed with Chytridiaceae, both here and in the study by Amses et al.[24]. The same can be said about Cladochytrium tenue, while C. replicatum belongs to Cladochytriales[24,32]. C. tenue is a sister to Zopfochytrium polystomum, which was previously named Cladochytrium polystomum but was renamed by Powell and co-workers after they showed that it belongs to the Chytridiaceae[32].
In Fig. 3, B. helicus + Rhizophlyctiales + Spizellomycetales form a well-supported clade within which families are distinct. This is consistent with the most recent phylogeny of species[24]. The positions of Blyttiomyces sp. JEL0837, B. helicus, C. tenue, and E. helicus TPPPs differ from the positions of other species of the given genera but are consistent with the most recent species phylogeny[24]. Moreover, P. arcticum TPPP is not among the Chytridiales, as would be consistent with the accepted phylogenomics and as the Phlyctochytrium bullatum TPPP is located, but in the Spizellomycetales clade, in a sister position with S. punctatus (bootstrap value 95%). However, recent studies[24,33] place P. arcticum exactly in this position, which means that the discrepancy applies not only to the given protein but to the species. In accordance with this, the species has been classified as Triparticalcar arcticum[24,33], as in the original article by Barr, who first defined the order Spizellomycetales[34] (Triparticalcar is a genus of Spizellomycetales; its other member is T. equi).
TPPPs and flagellum in fungi
-
It is known that the presence of TPPP is strongly associated with that of the flagellum[10], an organelle that extends from the plasma membrane and is built on a complex microtubule structure called the axoneme. The axoneme has a characteristic structure ('9+2'), with nine symmetric doublet microtubules surrounding two singlet microtubules in the center[35]. Each cilium has a centriole at its base, named also as a basal body, composed of microtubules arranged in nine triplets. The relationship between TPPP and flagellum is primarily based on bioinformatics[10,36], but there are also experimental data. FAP265 protein, the TPPP paralog of the biflagellated green alga, Chlamydomonas reinhardtii. was shown to be localized in the basal bodies and in the flagella, and its necessity in the formation of flagella was proven by using null mutants of FAP265[37]. In Plasmodium genus (P. falciparum, P. yoelli), TPPP orthologs (PFL1770c, Py05543) are necessary for male gametocyte exflagellation[38,39].
The above relationship also applies to fungi[6]; however, no experimental data are available. Species of the early branching flagellated phyla of fungi, with one exception (Neocallimastigomycota), all contain TPPP, while the later branching non-flagellated fungi do not (Fig. 5, Table 1). However, in Neocallimastigomycota there is a protein (apicortin) that has a partial p25alpha domain, precisely, the most conserved C-terminal part[6]. This protein also occurs in some other fungal phyla (Monoblepharomycota, Chytridiomycota, Rozellomycota)[6]. It is interesting that most of the zoospores of fungi have a single posterior flagellum, but some Neocallimastigomycota species can be polyflagellated[41]. More importantly (and curiously), they lack the centriole, the base of the flagellum[41]. One can speculate whether there is a cause-and-effect relationship between the lack of centriole and that of TPPP.
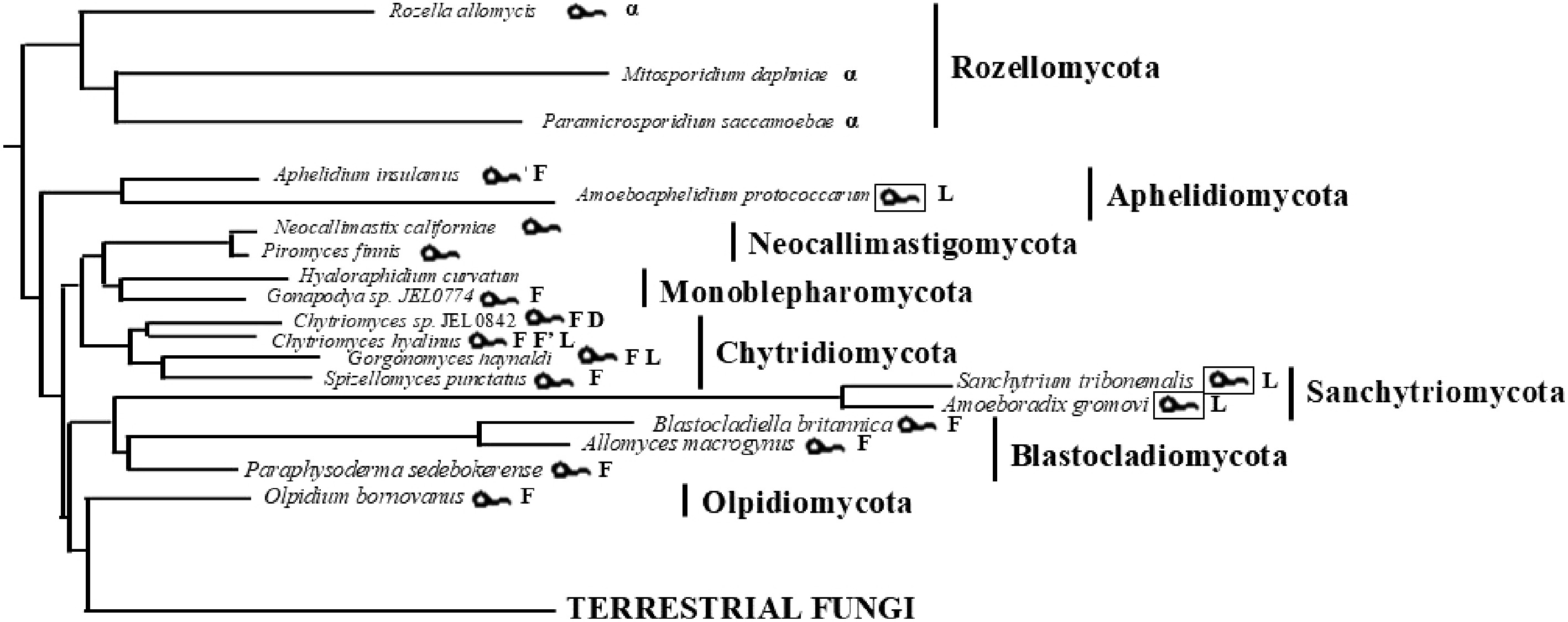
Figure 5.
Occurrence of flagellum and TPPP-like proteins in the Kingdom of fungi. The tree is based on the species phylogeny published in previous studies[24,40]. The presence of flagellum, fungal-type TPPPs, long, TPPP, double-long and TPPP-like p25alpha-containing domain proteins is labeled by the following symbols:

Table 1. Occurrence of TPPPs in various fungi. How many species are there in the given phylogenetic unit in which it occurs?
Phylogenetic
unitTPPP Long Fungal Fungal2 Double-long Blastocladiomycota Phylum − 10 − − Chytridiomycota Phylum 14 53 17 1 Chytridiomycetes Class 14 53 17 1 Caulochytridales Order − 1 − − Chytridiales Order 7 19 17 1 Chytriomycetaceae Family 7 11 12 − Chytridiaceae Family − 7 5 − Cladochytriales Order − 3 − − Lobulomycetales Order − 3 − − Polychytriales Order − 1 − − Rhizophlyctidales Order − 2 − − Rhizophydiales Order 7 11 − − Synchytriales Order − 2 − − Spizellomycetales Order − 10 − − Monoblepharidomycota Phylum − 1 − − Olpidiomycota Phylum − 1 − − Sanchytriomycota Phylum 2 − − − Aphelidiomycota Phylum 1 3 − − Kingdom of fungi 17 68 17 1 Among early-branching fungi, there are also some that have lost their flagellum. These include the Microsporidians of the Rozellomycota phylum[42], the Hyaloraphydium curvatum of the Monoblepharomycota phylum[43], and some other cases, in the phyla Sanchytriomycota[25], and Aphelidiomycota (genus Amoeboaphelidium)[27]. These Sanchytriomycota and Aphelidiomycota species possess a highly reduced, non-motile posterior flagellum, named pseudocilium. They are at an intermediate position between flagellated and non-flagellated fungi. Many, but not all, of the flagellar proteins were lost in these species[25]. However, the Sanchytriomycota A. gromovi and S. tribonematis, and the Aphelidiomycota Amoeboaphelidium protococcorum do have a TPPP, a long one, while Amoeboaphelidium occidentale does not (Fig. 5). A. tribonematis and P. tribonematis, which belong also to the phylum Aphelidiomycota and did not lose their flagellum, both possess a fungal-type TPPP. It seems that the occurrence of a pseudocilium is connected to the presence of the long, but not the fungal type TPPP. One could only speculate whether this is by chance or for a reason.
Although Microsporidians (class Microsporidea, phylum Rozellomycota) lost their flagellum, some of their species (Mitosporidium daphniae and Paramicrosporidium saccamoebae) have TPPP-like proteins (Fig. 5) in which proteins contain a large part of the p25alpha domain (approximately the first two-thirds), which accounts for approximately half of their sequences. It can be hypothesized that the absence of the last part of the p25alpha domain, which is responsible for microtubule binding, is not independent from the loss of flagellum since there is no need for the stabilization of the axonemal microtubules of the flagellum. The situation is almost the same in the flagellated Rozellomycota Rosella allomycis, in which the protein contains a somewhat larger part of the domain beside additional long sequences not characteristic of TPPPs.
TPPP paralogs in fungi
-
Figure 5 and Table 1 summarize the occurrence of TPPPs in early branching flagellated fungi. The most common TPPP is the fungal type one, which occurs in the majority of the phyla: Aphelidiomycota, Monoblepharomycota, Chytridiomycota, Blastocladiomycota, and Olpidiomycota. Long TPPP can be found in Chytridiomycota, Aphelidiomycota, and Sanchytriomycota.
A unique TPPP (double long) is found in a Chytridiales species, in which the entire p25alpha domain is present twice (Chytridiales sp. JEL 0842, KAJ3407993). This is the only case among fungi; only a few other examples are known in other Opisthokonta[9]. In the phylogenetic analysis, the two domains were considered as 'apparent long TPPPs' (Chytridialessp2A, Chytridialessp2B) (Figs 2–4). In each case, they form a highly supported clade, sister to each other. This indicates that the double-length protein is the result of an intraspecific duplication.
One of the orders of Chytridiomycota, Chytridiales, is unique in that it has two kinds of fungal-type TPPPs. One of them always forms a distinct clade on the trees, outside Chytridiales or Chytridiomycota (Figs 3, 4). It is puzzling that it only occurs in the order Chytridiales, and yet on the phylogenetic trees it is found outside Chytridiales and Chytridiomycota, respectively, and in fact is in a sister position to all fungal TPPPs, or at least to TPPPs of all 'traditional' fungi, without Aphelidiomycota. This actually suggests that we are dealing with a distinct TPPP. If this paralog were the result of a gene duplication within Chytridiales/Chytridiomycota, then it should be within Chytridiales/Chytridiomycota on the phylogenetic tree. However, if it arose earlier, why is its presence limited to only one order? It suggests a selective loss in the other fungal lineages.
The effect of long TPPPs on the dynamics of the tubulin/microtubule system has been experimentally demonstrated in animals[2,3,44]. The prominent role of the conservative C-terminus has also been demonstrated in this process[45,46]. In the case of fungi, there are no experimental data available, but it is reasonable to assume that long TPPPs have the same effect here as well. In fungal TPPPs, the C-terminus, which plays a prominent role in binding to tubulin/microtubules, is present twice. This may mean stronger binding or allow the binding of more tubulin/microtubules at the same time, promoting microtubule bundling. However, this still needs to be experimentally demonstrated. Thus, it is not known whether there is any other function that explains the presence of two or even three types of TPPP in fungi.
As mentioned, in Microsporidians that have lost their flagellum, although there is no TPPP in the traditional sense, there are proteins that contain the first two-thirds of the p25alpha domain, lacking the C-terminus that is crucial for microtubule binding. The loss of the flagellum in these species may have been accompanied by degeneration of the TPPP-like protein, with the loss of its C-terminus. In some fungal species that have only a rudimentary pseudocilium instead of a true flagellum, there is no fungal type TPPP, but there is a long one; however, in flagellated species of the same phylum (Aphelidiomycota), there is fungal type TPPP. This is consistent with the view that attributes special importance to the simultaneous presence of the two p25alpha domains (full and C-terminal) in microtubule binding and flagellum-connected functions.
In those fungal phyla that contain flagellated species that do not have real TPPP (Neocallimastigomycota, Rozellomycota), there is another protein, apicortin, that contains the C-terminal p25alpha part, together with another microtubule-binding domain (DCX), so it is conceivable that in these species apicortin may fulfill a similar role as TPPP elsewhere[47].
Finally, there is the question of what the need is for the two (three) types of TPPP in fungi? First, it is important to highlight that while previously only fungi were known to have fungal-type TPPP, very recently, it was shown that they are present sporadically in some early-branching animal phyla (Porifera, Cnidaria)[9]. The situation is exactly the opposite for animals and for fungi. The former contain mainly long (or animal-type) TPPP, and only occasionally the fungal type one[9]; the latter possess usually fungal type, and sometimes long TPPP. Nevertheless, these facts suggest that both types were present in the common ancestor of fungi and animals, and the shift in the ratio of TPPP paralogs occurred later. In animals, flagellum and cilium of various types and functions occur[48−51], while in fungi their presence may serve a single role, which is associated with the flagellum of zoospores[25,30,52]. This role may prefer the use of fungal-type TPPP for some reason.
Finally, it can be strengthened that TPPPs are present in the early diverging lineages of fungi, mostly connected with the presence of flagellum but they are absent in terrestrial (non-flagellated) fungi (Fig. 5), in accordance with the former suggestion[6,10] that each species possessing flagellum/cilium contains TPPPs or TPPP-like proteins.
Not applicable.
-
The author confirms sole responsibility for the following: study conception and design, data collection, analysis and interpretation of results, manuscript preparation, and approved the final version of the manuscript.
-
The data that support the findings of this study were derived from the following resources available in the public domain: National Library of Medicine (www.ncbi.nlm.nih.gov/protein/; www.ncbi.nlm.nih.gov/nuccore/), and in previous studies [16-18].
-
The author declares no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/sif-0026-0013.
- Supplementary Fig. S1 Multiple sequence alignments for figures 2, 3 and 4.
- Supplementary Fig. S2 Alignment of Chytridium lagenaria and Dinochytrium kinnereticum sequences.
- Supplementary Table S1 Accession numbers of TPPPs shown in figures 2−4.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Orosz F. 2026. Tubulin Polymerization Promoting Protein (TPPP) paralogs of flagellated fungi. Studies in Fungi 11: e015 doi: 10.48130/sif-0026-0013 

Tubulin Polymerization Promoting Protein (TPPP) paralogs of flagellated fungi
- Received: 27 December 2025
- Revised: 25 February 2026
- Accepted: 04 April 2026
- Published online: 25 May 2026
Abstract: Tubulin Polymerization Promoting Proteins (TPPPs) are a family of proteins whose occurrence is closely correlated with the presence of eukaryotic flagellum/cilium. This relationship is also observed in fungi, where these proteins are only found in species of early-branching, non-terrestrial phyla that usually reproduce by flagellated zoospores. The defining structural unit of these proteins is the p25alpha domain. TPPP paralogs differ in the length of this domain and the number of times it occurs in the protein. The paper systematically examines the occurrence of different types of TPPPs in the phyla of fungi, discussing the phylogenetic relationships between them. The types found in fungi are long TPPP, which has one full-length p25alpha domain; double long TPPP, which has two copies of the domain; and fungal type, in which the second p25alpha domain has only the C-terminal part. Of the latter, two different paralogs exist; one is found in several fungal phyla, while the other is found only in the order Chytridiales (phylum Chytridiomycota). Phylogenetic analysis has shown that although the second fungal type paralog occurs only in this single order, it is not the result of a duplication within the order. Furthermore, the phylogenetic trees of TPPPs generally agree with the species phylogeny. In several cases where the classification of individual species has been disputed, the phylogenetic tree of TPPPs confirms recent phylogenomic results (e.g., in Blyttiomyces, Entophlyctis, Cladochytrium, Phlyctochytrium genera). Finally, it can be noted that TPPPs are present in the early diverging lineages of fungi, mostly associated with the presence of a flagellum, but they are absent in terrestrial (non-flagellated) fungi.
-
Key words:
- Flagellum /
- p25alpha domain /
- Flagellated fungi /
- Phylogenetics /
- Chytridiales