









-
Oxalis vulcanicola 'Sunset Velvet' is a perennial dicotyledonous herb belonging to the Oxalidaceae family and the Oxalis genus. In recent years, it has been introduced as a new ornamental plant with excellent foliage and flowers[1]. This cultivar is widely appreciated for its unique heart-shaped leaves and the seasonal variation in leaf color. During the spring and autumn seasons, its leaves display a vibrant red color reminiscent of maple leaves, making it a popular choice among consumers and a new favorite in home gardening, earning the affectionate nickname 'Little Red Maple'. In addition, O. vulcanicola 'Sunset Velvet' has other varieties, such as 'Little Black Maple' with black-colored leaves and 'Elegant' with black-powdered foliage[2]. Another notable feature of this plant is its diurnal leaf movement, known as 'Nyctinasty'[3]. The leaves exhibit a fascinating opening and closing behavior in response to light intensity: they fully open in high-light environments and close in low-light environments. This characteristic makes it an excellent subject for studying plant light responses[4]. Furthermore, unlike other Oxalis species, O. vulcanicola 'Sunset Velvet' lacks underground bulbs and has white fibrous roots. These unique characteristics make O. vulcanicola 'Sunset Velvet' an ideal material for studying leaf color variation, nyctinasty, and bulb development. However, there are still significant challenges in gene function validation and genetic transformation due to the lack of a well-established regeneration system.
The regeneration systems for Oxalis species typically include three main pathways: somatic embryogenesis, indirect organogenesis, and direct organogenesis[5]. Somatic embryogenesis refers to the process in which cells dedifferentiate and form embryo-like structures under in vitro conditions. This pathway requires stringent environmental conditions, and the induction of somatic embryos often results in a high incidence of malformed embryos, making it less commonly used in Oxalis tissue culture. For example, Li & Chen[6] induced somatic embryos in O. triangularis ASt. Hil. through this method. In contrast, indirect organogenesis is the most commonly used tissue culture method for Oxalis species. This method involves the formation of callus from explants, followed by differentiation into shoots and roots, eventually resulting in the formation of a complete plant. For instance, Qiu et al.[7] used stem tips and leaves from three cultivars of O. obtuse to induce callus and obtain regenerated plants. Chen et al.[8] successfully induced sterile plantlets from leaf petioles of O. triangularis 'Purpurea'. However, the process of callus formation is complex and unpredictable, influenced by various internal and external factors such as plant hormone concentrations, medium composition, and plant genetic background[9]. This uncertainty leads to low efficiency in organogenesis, and the differentiation of callus often involves genomic instability and mutation risks, which can affect the genetic stability of regenerated plants[10]. Such instability can have negative effects on agricultural and horticultural applications. Moreover, the callus culture process is cumbersome and time-consuming, requiring multiple rounds of subculture, which further reduces regeneration efficiency[11].
In contrast, direct organogenesis bypasses the formation of callus and directly induces the formation of roots, shoots, or leaves from plant tissues[12]. This pathway has significant advantages, as it avoids the uncertainties and potential genomic instability associated with callus induction[13]. Studies have shown that direct organogenesis can significantly improve regeneration efficiency, and the organs produced are more similar to their natural counterparts, enhancing the physiological health and production adaptability of regenerated plants. For example, Chen et al.[14] established a rapid propagation system for O. vulcanicola 'Sunset Velvet' using stem segments with buds as explants. However, their study lacked statistical rigor, as it was based on a single experimental repetition, and did not optimize culture conditions for improved propagation efficiency. In this study, we incorporated three independent biological replicates to enhance data reliability and systematically evaluated the effects of different plant growth regulator (PGR) concentrations on cluster bud proliferation and browning. These improvements provide a more robust and efficient propagation strategy for large-scale production. Wang et al.[15] used bulbs from O. grandis as explants and induced shoot proliferation to obtain regenerated plants. These benefits also have been demonstrated across various plant species. For example, an optimized micropropagation protocol for Eucommia ulmoides facilitated direct shoot organogenesis from nodal segments with axillary buds, significantly enhancing propagation efficiency by reducing the cultivation period to only four weeks for bud elongation and proliferation[9]. In Populus deltoides, direct shoot organogenesis from leaf explants achieved high regeneration efficiency and ensured genetic stability of regenerated plants[10]. Similarly, an efficient protocol for Curculigo latifolia using a liquid culture system resulted in high genetic similarity (89%) between regenerated and mother plants, highlighting the method's ability to minimize somaclonal variation[16]. Additionally, direct shoot organogenesis in Lycium chinense was confirmed as an efficient regeneration system with genetic stability verified using ISSR markers, making it suitable for genetic transformation studies[17]. In Chrysanthemum indicum, direct shoot organogenesis from leaf and petiole explants provided a reliable system for rapid clonal propagation and genetic transformation[18].
In summary, this study uses stem segments from O. vulcanicola 'Sunset Velvet' as explants and employs direct organogenesis to induce shoot proliferation, optimize hormone formulations, and optimize rooting conditions. The goal is to establish an efficient clonal propagation system. This research will not only provide technical support for the large-scale production of O. vulcanicola 'Sunset Velvet' but also lay the foundation for gene function validation and genetic transformation experiments, further advancing the potential of this cultivar in horticultural applications and gene function studies.
-
The O. vulcanicola 'Sunset Velvet' seedlings were purchased from the Taobao store (
https://tb.alicdn.com/snapshot/index.html ) 'Xiao Wang's Four Seasons Plant Garden' on October 14, 2023. We invited Qiaoxian Xu, a teacher from the Institute of Corn and Characteristic Dry Grains of Zhejiang Academy of Agricultural Sciences, to identify the purchased plant materials. The plants were grown in 12 cm × 12 cm (diameter × height) plastic flowerpots with a substrate composed of peat soil : perlite (3:1, v:v). The plants were cultivated in a climate box at the Physiology Laboratory (Room 504), Jingyi Building, Fujian Agriculture and Forestry University, Fuzhou, Fujian Province, China (Latitude: 26°03'23" N, Longitude: 119°10'4" E). The conditions for incubation were: light intensity: 2,000 lx, provided by fluorescent lamp, with 12 h of light/12 h of dark light period. Temperature: 25 °C during the day, 22 °C at night. Humidity: Relative humidity is maintained at 60%−70%. The plants were subjected to standard care after planting.The culture medium was prepared with 4.74 g·L−1 MS (Murashige and Skoog) basal medium + 30 g·L−1 sucrose + 5 g·L−1 agar, with a pH of 5.8−6.0. The medium was autoclaved at 121 °C for 20 min. All inoculation tools, including blades, handles, tweezers of varying lengths, inoculation plates, and filter paper, were sterilized by autoclaving before use. The tissue culture chamber was maintained with light intensity at 2,000 lx, a 12-h light/dark cycle, a temperature of 25 ± 2 °C, and relative humidity of 50%−70%.
The disinfectants used were 75% alcohol and 5% sodium hypochlorite, purchased from Fuzhou Melissa Biotechnology Co., Ltd. MS medium (without agar and sucrose) was purchased from Beijing Solebao Technology Co., Ltd., (Beijing, China) while agar was purchased from Xilong Science Co., Ltd, and sucrose was sourced from Hangzhou Mobarium Biotechnology Co., Ltd.
Effect of disinfection time on sterile system establishment
-
In this experiment, healthy, pest-free stem segments from the O. vulcanicola 'Sunset Velvet' plant were selected as explants. The explants were first brushed clean of surface dirt with a soft brush, then cut into pieces of about 7 cm. The segments were soaked in a small amount of detergent (Eagle brand phosphorus-free laundry detergent) for 1 h, followed by covering the bottle openings with autoclaved gauze. They were then rinsed under running water for 3 h, followed by a 2-min rinse with sterile water. A sterile scalpel was used to remove petioles and leaves, which were sterilized with 75% ethanol for 1, 2, and 3 min, and then treated with 3% sodium hypochlorite for 10, 12, and 15 min. After that, the explants were rinsed with sterile water for 2 min and the stem segments were cut into small pieces of about 0.5 cm. These were inoculated onto sterile medium (MS + 0.50 mg·L−1 NAA + 1.00 mg·L−1 6-BA + 30 g·L−1 sucrose + 5 g·L−1 agar) for further analysis of the optimal disinfection procedure for O. vulcanicola 'Sunset Velvet' stem segments. Nine treatments (75% ethanol: 1, 2, and 3 min, and 3% sodium hypochlorite: 10, 12, and 15 min) were performed, with each treatment consisting of 15 culture bottles. Two explants were inoculated per culture bottle, totaling 30 explants per treatment, and repeated three times. After 30 d, the contamination rate and survival rate were recorded.
Adventitious shoot induction
-
Sterile stem segments with leaf petioles were used as explants for adventitious shoot induction. An orthogonal experimental design was employed to optimize the regeneration conditions. The hormone concentrations were selected based on the experimental protocol of Jiang et al.[19], which established an effective range for adventitious shoot induction in a similar plant system. These concentrations have been widely used in related studies to optimize regeneration efficiency. The MS medium was supplemented with varying concentrations of (0.10, 0.50, 1.00 mg·L−1) NAA, and (1.00, 2.00, 3.00 mg·L−1) 6-BA, along with 30 g·L−1 sucrose and 5 g·L−1 agar, with the pH adjusted to 5.8–6.0. Each culture dish contained 30 explants, and each treatment was replicated three times. After 30 d of culture, the induction rate of adventitious shoots was recorded and analyzed to determine the optimal hormonal combination.
Culture medium for healthy seedling growth
-
The 1 cm tall clump buds that have been reproduced twice in the optimal induction medium were cut into small pieces of about 0.30 g. These were inoculated onto the MS + 0.50 mg·L−1 NAA + 1.00 mg·L−1 6-BA + 30 g·L−1 sucrose + 5 g·L−1 agar + (0.00, 0.10, 0.20, 0.30, 0.40, 0.50 mg·L−1) IBA, pH 5.8–6.0. Each treatment had five culture bottles, with six explants per bottle and three replicates. After 30 d, the growth of healthy seedlings was analyzed, focusing on the multiplication rate and browning rate.
Rooting medium
-
The sterile seedlings of proliferation culture were transferred to the optimal induction medium (MS + 0.50 mg·L−1 NAA + 1.00 mg·L−1 6-BA + 30 g·L−1 sucrose + 5 g·L−1 agar), and the clumping buds obtained by two generations of culture were selected and cut off from the stem tips of about 3 cm of clumping buds with consistent growth for rooting culture, were transferred to rooting medium consisting of 1/2 MS + (0.00, 0.05, 0.10, 0.20 mg·L−1) NAA + 30 g·L−1 sucrose + 5 g·L−1 agar, pH 5.8–6.0. Each treatment included five culture bottles, with six explants per bottle and three replicates. After 30 d, root formation was assessed by measuring the rooting rate, average root number, and average root length.
Hardening and transplantation
-
The small plantlets with roots of about 3 cm long after rooting treatment were transplanted by refining seedlings. The tissue culture flask was moved outdoors, the cap was opened, the seedlings were refined under natural scattered light conditions for 3 d, the plants were removed by tweezers, the medium on the roots was washed with tap water and transplanted into a substrate of peat : perlite = 7:3 (v/v), and three plants were transplanted in each pot. After planting, watering thoroughly, and spraying 800 times liquid carbendazim (Sichuan Guoguang brand carbendazim 50%, wettable powder), the samples were covered with film to moisturize, shaded in a cool place, with a temperature kept at about 25 °C, the relative humidity is about 80%, sprayed with water 2–3 times a day, properly ventilated after 3 d, and reduced the number of water sprays after 7 d.
Data collection and statistical analysis
-
Data analysis was performed using SPSS 25.0.0.2 and Excel 2023. One-way ANOVA was used for multiple comparisons with SPSS 25.0.0.2, followed by Tukey's HSD post-hoc test to identify differences between groups. All data were expressed as mean ± standard deviation and statistical significance was considered at p < 0.05.
The following formulas were used for the statistical analysis:
(1) Contamination Rate = (Number of contaminated explants/Total number of inoculated explants) × 100%
(2) Adventitious Shoot Induction Rate = (Number of explants with adventitious shoots/Total number of inoculated explants) × 100%
(3) Seedling Multiplication Rate = (Weight of explants after 30 d/Weight of explants at inoculation) × 100%
(4) Browning Rate = (Total mass of browning explants after 30 d/Total mass of explants after 30 d) × 100%
(5) Rooting Rate = (Number of rooted plants/Total number of explants per bottle) × 100%
(6) Average Root Number = (Total number of roots per bottle/Total number of explants per bottle) × 100%
(7) Average Root Length = (Total root length per bottle/Total number of explants per bottle) × 100%
Each treatment had five culture bottles, with six explants per bottle and three replicates.
-
For the sterilization of stem segments as explants, leaf development began at the petiole region on day 3, and a few explants began to form adventitious roots at the wound sites by day 7. On day 10, the two ends of the stem segments began to expand, and by day 12, some segments induced callus formation at both ends (Supplementary Fig. S1). The experimental results indicated that as the sterilization time with 75% ethanol and 3% sodium hypochlorite increased, the contamination rate gradually decreased (Table 1). The highest contamination rate was 92.22% when the explants were treated with 75% ethanol for 1 min and 3% sodium hypochlorite for 10 min. The lowest contamination rate was 10.00% when treated with 75% ethanol for 3 min, and 3% sodium hypochlorite for 15 min. Excessively prolonged sterilization times caused damage to the explants, leading to browning and even death during subsequent culturing. The sterilization time with 75% ethanol was gradually increased, while the time for sterilization with 3% sodium hypochlorite remained fixed. As the sterilization time with ethanol increased, the germination rate initially increased and then decreased. The highest germination rate (50.00%) was observed when the explants were treated with 75% ethanol for 2 min and 3% sodium hypochlorite for 15 min, with a contamination rate of 14.44%. However, in the T9 treatment, which had the lowest contamination rate of 10.00%, the germination rate was only 16.67%. Therefore, the optimal sterilization method was 75% ethanol for 2 min and 3% sodium hypochlorite for 15 min.
Table 1. Statistics of disinfection contamination rate of 75% alcohol and 3% sodium hypochlorite.
Treatment 75% alcohol (min) 3% sodium hypochlorite
(min)Contamination rate (%) Germination rate (%) T1 1 10 92.22 ± 5.09a 0.00 ± 0.00e T2 1 12 55.56 ± 5.09b 4.44 ± 3.85e T3 1 15 33.33 ± 3.33e 24.44 ± 3.85bc T4 2 10 50.00 ± 5.77b 5.56 ± 1.92e T5 2 12 24.44 ± 3.85cd 31.11 ± 5.09b T6 2 15 14.44 ± 1.92ef 50.00 ± 3.33a T7 3 10 46.67 ± 6.67b 24.44 ± 5.09bc T8 3 12 23.33 ± 3.33de 18.89 ± 1.92cd T9 3 15 10.00 ± 3.33f 16.67 ± 3.33d The data are mean ± standard deviation, and different letters after the same column of numbers indicate significant differences between different treatments (p < 0.05). Each treatment contains 30 explants, and each treatment was repeated three times. Effect of 6-BA and NAA on adventitious shoot induction
-
For the induction of adventitious shoots using stem segments with petioles as explants, leaf development began at the petiole region on day 3, and callus formation was observed at both ends of some explants by day 12. The results showed that when the NAA concentration was 0.10 mg·L−1, the adventitious shoot induction rate increased and then decreased with increasing concentrations of 6-BA (Table 2). When the NAA concentration was 0.50 or 1.00 mg·L−1, the adventitious shoot induction rate decreased as the concentration of 6-BA increased. Adventitious shoots grew robustly in the low-concentration NAA and 6-BA medium, with lush plant growth, while in high-concentration NAA and 6-BA media, the growth was poor, and the plants were weak. The highest shoot induction rates of 74.45% were observed in groups Y4 and Y7, while the longest bud lengths were found in groups Y1 and Y4, measuring 0.96 cm and 0.99 cm, respectively. However, when the concentration of NAA was 0.50 mg·L−1 and 6-BA concentration was 1.00 mg·L−1, both the induction rate and bud length reached their maximum values. The induction rate was 74.45%, and the longest bud length was 0.99 cm. Additionally, the induced shoots were robust and displayed vibrant coloration (Fig. 1). Therefore, the optimal medium for adventitious shoot induction was MS + 0.50 mg·L−1 NAA + 1.00 mg·L−1 6-BA + 30 g·L−1 sucrose + 5 g·L−1 agar.
Table 2. Effect of different hormone combinations on induction and multiplication of O. vulcanicola 'Sunset Velvet'.
Treatment NAA
concentrations
(mg·L−1)6-BA
concentrations
(mg·L−1)Induction
rate (%)Bud length
(cm)Y1 0.1 1 38.89 ± 1.92d 0.96 ± 0.31a Y2 0.1 2 70.00 ± 3.33ab 0.69 ± 0.29b Y3 0.1 3 50.00 ± 3.33c 0.60 ± 0.18bc Y4 0.5 1 74.45 ± 6.94a 0.99 ± 0.34a Y5 0.5 2 71.11 ± 1.92ab 0.38 ± 0.10cd Y6 0.5 3 44.44 ± 5.09cd 0.28 ± 0.10d Y7 1 1 74.44 ± 5.09a 0.34 ± 0.08d Y8 1 2 71.11 ± 5.09ab 0.25 ± 0.11d Y9 1 3 60.00 ± 3.33b 0.31 ± 0.07d The data are mean ± standard deviation, and different letters after the same column of numbers indicate significant differences between different treatments (p < 0.05). Each treatment contains 30 explants, and each treatment was repeated three times. 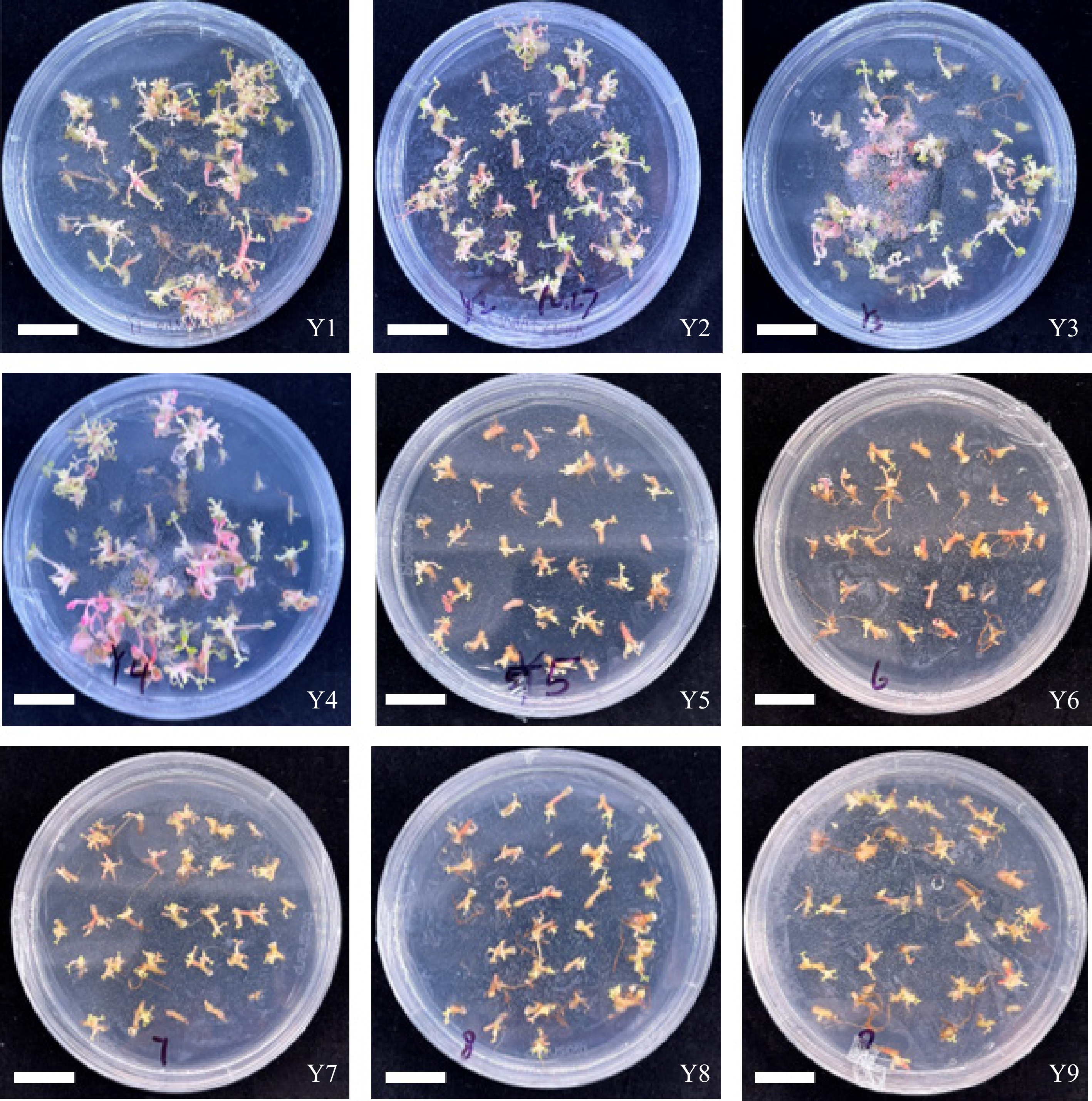
Figure 1.
Effects of 6-BA and NAA on clumping bud induction after, Y1: 0.1 mg·L−1 NAA + 1 mg·L−1 6-BA, Y2: 0.1 mg·L−1 NAA + 2 mg·L−1 6-BA, Y3: 0.1 mg·L−1 NAA + 3 mg·L−1 6-BA, Y4: 0.5 mg·L−1 NAA + 1 mg·L−1 6-BA,Y5: 0.5 mg·L−1 NAA + 2 mg·L−1 6-BA, Y6: 0.5 mg·L−1 NAA + 3 mg·L−1 6-BA,Y7: 1 mg·L−1 NAA + 1 mg·L−1 6-BA, Y8: 1 mg·L−1 NAA + 2 mg·L−1 6-BA, Y9: 1 mg·L−1 NAA + 3 mg·L−1 6-BA, the culture conditions are: light intensity at 2,000 lx, a 12-h light/dark cycle, a temperature of 25 ± 2 °C, and relative humidity of 50%−70%. Scale bars = 1 cm.
Effect of IBA on O. vulcanicola 'Sunset Velvet' plantlet growth
-
Approximately 1 cm tall O. vulcanicola 'Sunset Velvet' clumping buds that have undergone two generations in the optimal induction medium were transferred to the plantlet culture medium, new shoots began to grow from the axillary buds of the plants on day 3, and roots started to form at the base of some plants by day 7. On day 10, plants cultured in MS medium without IBA or with high concentrations of IBA began to show browning. The biomass of adventitious shoots decreased as the IBA concentration increased (Table 3). Browning rate decreased initially and then increased (Table 3, Fig. 2). In the MS medium without hormones or with 0.1 mg·L−1 IBA, the plants were relatively weak, with compact, dense shoots that were light green, and the biomass increased significantly. In the MS medium with 0.2 mg·L−1, 0.3 mg·L−1, or 0.4 mg·L−1 IBA, the plants were relatively robust, with bushy shoots that were dark green, though the biomass was lower, and the browning and mortality rates were higher. Plants cultured in MS medium with 0.5 mg·L−1 IBA were weak and showed high browning rates. The highest induction rate 20.32-fold was observed with CK, when 0.01 mg·L−1 IBA was added to the medium, the multiplication rate decreased slightly to 17.39-fold with the browning rate reduced to 10.21%.
Table 3. Effect of different IBA concentrations on the proliferation of clustered shoots.
Treatment IBA concentrations (mg·L−1) Biomass multiplication (folds) Browning rate (%) CK 0.00 20.32 ± 0.54a 18.65 ± 0.32d Z1 0.10 17.39 ± 0.70b 10.21 ± 0.33e Z2 0.20 14.99 ± 0.77c 17.98 ± 0.34d Z3 0.30 13.09 ± 0.38d 22.30 ± 0.46c Z4 0.40 11.78 ± 0.49d 26.88 ± 0.35b Z5 0.50 9.71 ± 0.59e 31.63 ± 0.19a The data are mean ± standard deviation, and different letters after the same column of numbers indicate significant differences between different treatments (p < 0.05). Each treatment contains 30 explants, and each treatment is repeated three times. 
Figure 2.
Proliferation of clustered shoots (scale bars = 1 cm), CK: 0.00 mg·L−1 IBA, Z1: 0.10 mg·L−1 IBA, Z2: 0.20 mg·L−1 IBA, Z3: 0.30 mg·L−1 IBA, Z4: 0.40 mg·L−1 IBA, Z5: 0.50 mg·L−1 IBA, the culture conditions are: light intensity at 2,000 lx, a 12-h light/dark cycle, a temperature of 25 ± 2 °C, and relative humidity of 50%−70%.
Effect of NAA on rooting in O. vulcanicola 'Sunset Velvet'
-
Without NAA, the number of roots was lower, the average root length was shorter, and the plants were smaller compared to those treated with NAA. As the NAA concentration increased, the average number of roots, average root length, and average plant height initially increased and then decreased (Table 4). In media without NAA or with 0.05 mg·L−1 NAA, a few lateral roots grew from the main root, and the root system was thinner, with some roots being gray-black and others white. The plants were pink in color with light green leaves. In media with 0.05 mg·L−1 NAA, the stems were red, and the leaves were dark green. When the NAA concentration was 0.05 mg·L−1 or 0.10 mg·L−1, rooting was 100%, but at 0.10 mg·L−1 NAA, the plants had the highest number of roots, the longest roots, and the tallest plant height, and the plants exhibited many white root hairs growing at the upper part of the medium, with gray-black roots and lateral roots forming at the base of the main root. The plant stems were red, and the leaves were green. When the NAA concentration was increased to 0.20 mg·L−1, the rooting rate decreased, and the roots turned black, with no lateral roots growing on the main root (Fig. 3).
Table 4. Effects of different concentrations of NAA on plant growth and rooting.
Treatment NAA concentrations (mg·L−1) Rooting rate (%) Number of roots Average root length (cm) Plant height (cm) G1 0.00 93.33 ± 3.33b 7.05 ± 0.10d 2.10 ± 0.49d 3.51 ± 0.06c G2 0.05 100.00 ± 0.00a 10.10 ± 0.44c 2.54 ± 0.04b 3.62 ± 0.08c G3 0.10 100.00 ± 0.00a 14.10 ± 0.10a 2.73 ± 0.15a 4.26 ± 0.05a G4 0.20 87.77 ± 1.93c 12.83 ± 0.25b 2.40 ± 0.10c 3.97 ± 0.11b The data are mean ± standard deviation, and different letters after the same column of numbers indicate significant differences between different treatments (p < 0.05). Each treatment contains 30 explants, and each treatment is repeated three times. 
Figure 3.
Thirty days of plant growth and rooting , the starting material was to cut about 3 cm from the tip of the shoot of the clumping buds cultured on the optimal proliferation medium for two generations to be used as an explant insertion into the rooting medium, G1: 0.00 mg·L−1 NAA, G2: 0.05 mg·L−1 NAA, G3: 0.10 mg·L−1 NAA, G4: 0.20 mg·L−1 NAA, the culture conditions are: light intensity at 2,000 lx, a 12-h light/dark cycle, a temperature of 25 ± 2 °C, and relative humidity of 50%−70%, scale bars = 1 cm.
Transplanting of sterile plantlets
-
As shown in Fig. 4a, the plant underwent a hardening process. This is achieved by gradually adapting the seedlings to the external environment. The hardening process involves moving the tissue culture flasks outdoors to open and cap the bottle and refining the seedlings under naturally scattered light conditions for 3 d to ensure that the seedlings are fully prepared for transplantation. After transplanting three seedlings per pot, water the plants thoroughly, then spray them 2−3 times approximately every 3 d, and reduce the frequency of watering after 7 d. A total of 60 seedlings were transplanted, of which 57 survived after 2 weeks under optimal growth conditions (cool and ventilated place, temperature 25 °C, relative humidity 80%) (Fig. 4b).

Figure 4.
Tissue culture seedlings refining situation (scale bars = 1 cm). (a) Tissue culture seedlings. (b) The growth of seedlings after refining.
-
This study established an efficient and complete regeneration system for O. vulcanicola 'Sunset Velvet' using stem segments with axillary buds as explants. Effective sterilization of explants is a prerequisite for establishing a regeneration system. By combining different sterilizing agents with varying sterilization times, the contamination rate of the explants can be reduced, and the survival rate of the explants can be improved. Common sterilizing agents used in the establishment of aseptic systems for Oxalis species include 0.1% mercuric chloride[20,21], 75% ethanol[22], and 3% sodium hypochlorite[14,15]. Ethanol treatment is usually limited to 1 min, while 3% sodium hypochlorite and 0.1% mercuric chloride are typically applied for 3 to 8 min. However, due to the toxic nature of mercuric chloride and difficulties in waste disposal, it is no longer the preferred sterilizing agent in laboratories. Therefore, this experiment used 75% ethanol and 3% sodium hypochlorite as the sterilizing agents to establish the aseptic system.
A moderate duration of ethanol exposure effectively eliminates surface contaminants, such as fungi or bacteria, while maintaining the integrity of the explants. However, prolonged exposure to ethanol may damage the cell membranes and interfere with the physiological processes required for successful germination. Previous studies have shown that short-term ethanol treatment can enhance the germination rate by preventing microbial contamination, while excessive exposure can lead to toxicity and reduced plant viability[23]. Thus, the optimal sterilization time needs to balance between sufficient disinfection and minimal damage to the explants. In this experiment, the contamination rate of O. vulcanicola 'Sunset Velvet' stem explants was lowest, and the germination rate was highest when treated with 75% ethanol for 2 min and 3% sodium hypochlorite for 15 min. Compared to sterilization times for the petioles or stem tips of other Oxalis species, such as Oxalis[24], O. triangularis 'Purpurea'[21], and O. corniculata[7], the sterilization time for O. vulcanicola 'Sunset Velvet' was relatively longer. This is likely due to the presence of trichomes on the surface of the stem segments, which make sterilization more challenging, and bacteria in the gaps between the petiole and stem are harder to eliminate. Therefore, extended sterilization time was necessary to ensure thorough disinfection. According to the results of the preliminary experiments, it is recommended to be in subsequent experiments, stem segments with buds that sprouted in the same year (from March to May) should be used, as newly formed stems are cleaner and more easily sterilized. Additionally, spring temperatures are cooler, which limits bacterial growth, making it a more suitable time for establishing aseptic systems.
Effects of NAA and 6-BA on shoot induction in O. vulcanicola 'Sunset Velvet'
-
The majority of plant tissue culture propagation techniques rely on axillary bud regeneration, which allows the regeneration plants to retain the desirable traits of the parent plant[25]. This method is the most reliable for maintaining genetic stability during in vitro propagation[26,27]. Axillary buds in stem segments originate from the axillary meristem, and young shoots typically grow more vigorously. Since the stem segments store nutrients that support the subsequent growth of axillary buds, the growth rate of the axillary buds is fast[28].
Hormones are essential for organ differentiation during tissue culture, and plant growth regulators such as auxins and cytokinins are commonly selected based on the needs of the explants at different stages of culture[29]. Direct organogenesis, such as the induction of multiple shoots, is a rapid way to obtain tissue-cultured plants. In Oxalis species, the hormones 6-BA and NAA are commonly used for shoot induction, with the typical concentration ranges being 0.50–2.00 mg·L−1 for 6-BA and 0.20–0.50 mg·L−1 for NAA[19]. In this experiment, 6-BA and NAA were used as plant growth regulators during the induction phase. NAA is a stable plant hormone that is not influenced by the plant's internal metabolism. It can enter the cells of plant tissues and maintain a high concentration to induce cell division. Therefore, high concentrations of NAA are beneficial for shoot initiation, but excessively high concentrations can cause the growth of weak multiple shoots, leading to browning and death[14].
In this experiment, the optimal concentration of NAA for inducing multiple shoots was 0.50 mg·L−1. This result is consistent with the findings of Wang et al.[15], who determined that 0.50 mg·L−1 NAA was the best concentration for inducing multiple shoots in Oxalis bowiei. When the NAA concentration was 0.50 mg·L−1 or higher, an increase in the concentration of 6-BA led to a decrease in the shoot induction rate. Therefore, the optimal medium for multiple shoot induction was MS + 0.50 mg·L−1 NAA + 1.00 mg·L−1 6-BA + 30 g·L−1 sucrose + 5 g·L−1 agar, which resulted in the highest shoot induction rate of 74.45%, consistent with the findings of Xiao & Cao[30] in tissue culture of O. triangularis 'Purpurea'.
Effects of hormones on proliferation in O. vulcanicola 'Sunset Velvet'
-
Proliferation is a key step in tissue culture, as the proliferation coefficient reflects the speed and efficiency of plant propagation. It is an important indicator for evaluating the total production of seedlings in factory-based nursery systems[31]. 6-BA is known to promote cell division and differentiation, making it widely used in plant tissue culture. When used in combination with IBA, 6-BA enhances cell division. The appropriate ratio of 6-BA to IBA not only promotes the proliferation of adventitious buds but also supports robust plant growth. Zhang et al.[32] found that, under constant 6-BA concentrations, the proliferation coefficient increased and then decreased with rising IBA concentrations, suggesting that high concentrations of IBA inhibit the proliferation of adventitious buds. IBA effectively reduces browning in plant tissue culture by regulating cell division, hormone balance, and antioxidant responses. It promotes cell division and expansion, thereby mitigating cell death and oxidative reactions that lead to browning. Studies have shown that IBA modulates plant hormone balance, particularly enhancing cytokinin synthesis, which helps suppress browning[33]. Additionally, IBA may reduce browning by enhancing antioxidant enzyme activity and decreasing reactive oxygen species (ROS) accumulation[34]. The effectiveness of IBA varies across different plant species and culture conditions. For instance, in the tissue culture of Oryza sativa and Solanum lycopersicum, IBA effectively inhibits browning and promotes callus formation[35]. However, in fruit trees such as Malus domestica and Vitis vinifera, the effect of IBA is weaker, possibly due to lower cell division response and specific endogenous hormone levels[36]. Therefore, further research into the differential responses of various plants to IBA will help optimize its application conditions. In this experiment, MS + 0.50 mg·L−1 NAA + 1.00 mg·L−1 6-BA + 30 g·L−1 sucrose + 5 g·L−1 agar reduced the browning rate of clumping buds, and when the concentration of IBA was higher than 0.1 mg·L−1, not only the proliferation coefficient decreased, but also the browning rate increased.
Effects of hormones on rooting in O. vulcanicola 'Sunset Velvet'
-
The growth and root development of regenerated plants are crucial for improving transplant survival rates. Therefore, rooting is a key step in the regeneration process. During the rooting phase, 1/2 MS medium supplemented with different concentrations of NAA is commonly used. NAA promotes cell division and expansion and induces adventitious root formation. In this experiment, O. vulcanicola 'Sunset Velvet' exhibited 100% rooting when cultured in 1/2 MS medium supplemented with 0.05 and 0.10 mg·L−1 NAA, which is consistent with the results of Chen et al.[14]. In rooting media containing only NAA, higher concentrations led to thicker roots, but plant survival rates were lower. In a study of rooting in O. corniculata, Qiu et al.[7] found that 0.1 mg·L−1 NAA resulted in the highest rooting rate, with the plants having dense and thick roots, when the NAA concentration exceeded 0.1 mg·L−1, the number of roots decreased, and plant growth was impaired, consistent with the findings of this study.
-
This study successfully established an efficient tissue culture system for Oxalis vulcanicola 'Sunset Velvet', optimizing key parameters for the initiation, shoot proliferation, and rooting of plantlets. The optimized disinfection protocol and hormone combinations were crucial in achieving high rates of shoot induction, proliferation, and rooting. Specifically, a medium containing 0.50 mg·L−1 NAA + 1.00 mg·L−1 6-BA + 30 g·L−1 sucrose + 5 g·L−1 agar was found to be highly effective for shoot induction and proliferation, with notable improvements in browning rates when IBA was included in the proliferation medium. Additionally, the rooting medium of 1/2 MS + 0.10 mg·L−1 NAA + 30 g·L−1 sucrose + 5 g·L−1 agar successfully supported the formation of healthy roots. These findings provide a robust foundation for the large-scale propagation of O. vulcanicola 'Sunset Velvet', as well as for future genetic transformation efforts, which could facilitate further genetic improvement and commercial utilization of this ornamental cultivar.
This work was supported by a grant from the National Natural Science Foundation of China (Grant No. 32471958) and the 2021 Fujian Agriculture and Forestry University 'Rural Revitalization Service Team' Support Program Project (Grant No. 11899170151).
-
The authors confirm contribution to the paper as follows: study conception and design: Wu S; carrying out the experiment, data analysis, and writing the manuscript: Wang X, Tuo W; data analysis: Zhang S, Wu T, Li J; manuscript revision: Zhai J, Wu S. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during or analyzed during the current study are available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xuexuan Wang, Wanli Tuo
- Supplementary Fig. S1 Clumping bud induction process.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang X, Tuo W, Zhang S, Wu T, Li J, et al. 2025. Efficient plant regeneration through direct shoot organogenesis of Oxalis vulcanicola 'Sunset Velvet'. Technology in Horticulture 5: e020 doi: 10.48130/tihort-0025-0017 

Efficient plant regeneration through direct shoot organogenesis of Oxalis vulcanicola 'Sunset Velvet'
- Received: 07 January 2025
- Revised: 08 March 2025
- Accepted: 14 April 2025
- Published online: 23 May 2025
Abstract: Oxalis vulcanicola 'Sunset Velvet', a cultivar of Oxalidaceae, is a perennial dicot known for its compact growth, vibrant foliage, and attractive flowers, making it a popular choice for ornamental potted plants. Despite its ornamental value and physiological significance, significant challenges remain in gene function validation and genetic transformation due to the lack of a well-established regeneration system. This study aimed to establish an effective tissue culture system for O. vulcanicola 'Sunset Velvet' to facilitate genetic transformation and large-scale propagation. Sterile stem segments with buds were used as explants, and a direct shoot organogenesis pathway was employed to initiate the culture system. Optimization of hormone combinations led to the selection of suitable induction, proliferation, and rooting media. The most effective disinfection protocol was a 2-min treatment with 75% ethanol, followed by a 15-min treatment with 3% sodium hypochlorite. The optimal medium for inducing multiple shoot formation consisted of MS + 0.50 mg·L−1 NAA + 1.00 mg·L−1 6-BA + 30 g·L−1 sucrose + 5 g·L−1 agar, achieving an induction rate of 74.45 ± 6.94% and the longest shoot length of 0.99 ± 0.34 cm after 30 d of culture. For shoot proliferation, MS + 0.50 mg·L−1 NAA + 1.00 mg·L−1 6-BA + 30 g·L−1 sucrose + 5 g·L−1 agar achieved a multiplication rate of 20.32 ± 0.54-fold, with a browning rate of 18.65 ± 0.32%. When 0.01 mg·L−1 IBA was added to the medium, the multiplication rate decreased slightly to 17.39 ± 0.70-fold with the browning rate reduced to 10.21 ± 0.33%. For rooting, the best medium was 1/2 MS + 0.10 mg·L−1 NAA + 30 g·L−1 sucrose + 5 g·L−1 agar, resulting in an average of 14.10 roots per plant, with an average root length of 2.73 ± 0.15 cm and plant height of 4.26 ± 0.05 cm, starting from stem segments with an initial height of 3 cm. These findings provide important technical support for the large-scale propagation and genetic transformation of O. vulcanicola 'Sunset Velvet'.