








-
Anthurium andraeanum, a tropical perennial species in the family Araceae, is highly valued in the global floriculture industry for its vivid spathes and unique floral morphology[1,2]. As a leading cut flower species, it is widely cultivated for ornamental purposes and floral design[3−6]. However, during postharvest handling and vase life, cut A. andraeanum is prone to senescence, characterized by peduncle base browning, spadix necrosis, and spathe bluing or loss of glossiness[7], which markedly reduces its ornamental value and commercial quality. These symptoms are primarily associated with water imbalance, impaired nutrient metabolism, and oxidative damage.
During the vase period, continuous transpiration can lead to a decline in fresh weight and tissue water loss if water uptake is inadequate[8]. Concomitantly, excessive reactive oxygen species (ROS) promote membrane lipid peroxidation, leading to disruption of membrane structure and cellular integrity[9]. Malondialdehyde (MDA), the final product of lipid peroxidation, serves as a key indicator of oxidative stress and cell senescence[10]. Antioxidant enzymes such as superoxide dismutase (SOD) and peroxidase (POD) play critical roles in scavenging ROS and maintaining cellular homeostasis[11]. Moreover, chlorophyll degradation is widely regarded as a physiological marker of plant senescence and directly affects photosynthetic performance and flower coloration. In A. andraeanum, spathe discoloration and quality decline are closely associated with the breakdown of chlorophyll rather than anthocyanin loss, as the red coloration of the spathe is largely determined by cell structure and vacuolar pH. Recent work has further demonstrated that chlorophyll degradation strongly correlates with spathe yellowing and the loss of ornamental value during vase life[6], making it a reliable indicator for evaluating postharvest senescence[5, 12].
In recent years, plant growth regulators (PGRs) have been widely used in postharvest preservative solutions to extend vase life in ornamental species, with diverse responses across plant types[13]. Among them, Gibberellic acid (GA3) has been shown to delay senescence and enhance membrane stability, significantly prolonging the vase life of cut Dianthus caryophyllus, Narcissus pseudonarcissus, and Rosa chinensis[14−16]. 6-Benzylaminopurine (6-BA) can suppress ethylene biosynthesis and slow the degradation of chlorophyll and proteins, demonstrating effective preservation results in Petunia × atkinsiana, Dahlia pinnata, and Rosa chinensis[17−20]. For cut A. andraeanum, prior research indicates that GA3 and 6-BA can alleviate senescence symptoms and improve vase longevity and visual quality[21−23]. Notably, 6-BA application has been shown to markedly improve postharvest performance, although the response depends on cultivar and concentration. In 14 of 15 tested cultivars, 6-BA treatment increased vase longevity by up to 35 d, with the optimal concentration typically between 100 and 200 mg·L−1 [24]. Concentrations ranging from 37.5 to 300 mg·L−1 have also been reported to enhance freshness and maintain flower quality, particularly below 150 mg·L−1. Likewise, GA3 applied alone or in combination with spermine can substantially extend vase life by enhancing phenolic metabolism and antioxidant enzyme activity[21]. However, most existing research has focused on external factors such as season, cultivar, and placement conditions, while investigations on hormone treatments are relatively limited. In particular, studies combining hormone treatments with general preservative solutions are scarce. Moreover, most work has examined a single hormone, predominantly 6-BA, with relatively fewer reports on GA3. This study employed a sucrose + 8-hydroxyquinoline (8-HQ)-based preservative solution with gradient concentrations of GA3 and 6-BA to assess their effects on vase life, fresh weight change, water balance, antioxidant enzyme activities, MDA levels, and chlorophyll degradation in cut A. andraeanum. The objective was to identify optimal hormone application rates and explore their regulatory mechanisms. The findings aim to support the formulation of effective preservative solutions and advance the application of plant hormones in postharvest cut flower management, contributing to the development of green, efficient, and residue-free preservation technologies.
-
Cut flowers of A. andraeanum 'Michigan', a widely cultivated cultivar in the commercial market, were used in this study. Samples were collected from Greenhouse No. 6 of the Fujian Forestry Science and Technology Experimental Center, located in Nanjing County, Zhangzhou City, Fujian Province, China. Flowers were selected at the stage of full spathe expansion, with uniform size, straight and thick stems, vibrant color, healthy foliage, and free from pests and diseases.
Treatment
-
The basal ends of cut A. andraeanum 'Michigan' stems were trimmed at a 45° angle by 1.5 cm under distilled water, and all stems were standardized to a final length of 18 cm. Each stem was inserted into a preservation bottle filled to a depth of approximately 8 cm. The bottle mouths were sealed with Parafilm® to reduce evaporation. Treated flowers were maintained in a controlled environment chamber with day conditions (7:00 am to 6:00 pm) set at 25 °C and 10,000 lx light intensity, and night conditions (6:00 pm to 7:00 am) set at 18 °C and 0 lx. Each treatment included three replicates, with six cut flowers per replicate (Table 1). Relevant physiological parameters were measured every 5 d. The preservative solution was refreshed every 10 d. Each treatment consisted of 18 flower stems, arranged into three biological replicates, with six stems per replicate. To analyze physiological and biochemical responses, three petal samples were collected daily from each treatment (n = 3, as biological replicates) and assessed for SOD and POD activities, MDA content, and chlorophyll concentration. For analysis, petals from three flowers per treatment were pooled, ground in liquid nitrogen, and stored at −80 °C until further use.
Table 1. Experimental treatment formulations.
Treaments Sucrose (g·L−1) 8-HQ (mg·L−1) 6-BA (mg·L−1) GA3 (mg·L−1) CK1 20 200 0 0 CK2 Distilled water T1 20 200 10 0 T2 20 200 20 0 T3 20 200 30 0 T4 20 200 0 50 T5 20 200 0 100 T6 20 200 0 150 Vase life assessment
-
The vase quality of cut A. andraeanum flowers was observed at 5-day intervals throughout the experimental period, beginning on Day 0, the day of stem insertion. Vase life was assessed as the duration (in days) from the onset of vase placement until the occurrence of peduncle base browning, spadix necrosis, and spathe bluing or loss of glossiness, which indicated a decline in ornamental quality[21,22].
Fresh weight and water balance
-
The fresh weight and water balance of cut A. andraeanum petals were determined using a gravimetric method from day 0 of vase insertion. Fresh weight was measured every 5 d using a precision electronic balance. This method provides a quantitative assessment of water absorption dynamics and transpirational loss in cut flowers, contributing to the evaluation of postharvest hydration efficiency and overall water status.
The rate of fresh weight change was calculated using the following formula:
$ \text{Fresh weight change rate}\; ({\text{%}})=\dfrac{FW_t-FW_0}{FW_0}\times100 $ (1) where, FWt is the fresh weight at day t, and FW0 is the initial fresh weight.
The calculation of petal water balance was performed according to a documented procedure[25].
Involving differences in weight due to water uptake and loss:
$ \text{Water uptake }(W_a)=W_1-W_0 $ (2) where, W1 denotes the measured weight of the vase solution together with the container on the specified day, and W0 refers to the initial weight recorded at the start of the experiment.
$ \text{Water loss }(W_1)=T_1-T_0 $ (3) where, T1 indicates the combined weight of the vase, container, and flower on the measurement day, and T0 refers to the initial total weight at the beginning of the assessment.
$ \text{Water balance }(W_b)=W_a-W_1 $ (4) SOD and POD activities
-
Superoxide dismutase (SOD, E.C. 1.15.1.1) activity was assessed via the NBT photochemical reduction method, with slight modifications in buffer composition and reagent concentrations[26]. Petal samples weighing 0.5 g each (n = 3 per treatment per day, representing three biological replicates) were homogenized in 5 mL of ice-cold 0.05 M phosphate-buffered saline (PBS, pH 7.8) using a chilled mortar and pestle until a uniform extract was achieved. The control tube and blank tube contained buffer instead of the enzyme solution. All reaction tubes were gently shaken. One tube was kept in the dark as the control, while the remaining tubes were illuminated under a daylight lamp (approx. 4,000 lx, 24 °C) for 20 min to initiate the photochemical reaction. Following incubation, absorbance at 560 nm was recorded using a spectrophotometer, with the dark-incubated control serving as the baseline reference.
SOD activity, normalized to fresh weight (Ug·g−1 FW), was calculated using Eq. (5) provided below:
$ \begin{matrix}\mathrm{SOD}\; \mathrm{activity}\; \left(\mathrm{Ug}\cdot\mathrm{g}^{-1}\; \mathit{\mathrm{\mathit{FW}}}\right)=\dfrac{\text{(}\mathit{\text{A}_{\text{CK}}-\text{A}_{\text{E}}})\times\mathit{\text{V}_{\text{T}}}}{0.5\times\mathit{\text{A}_{\text{CK}}\times\text{V}_{\text{S}}\times\text{W}}}\end{matrix} $ (5) where, ACK is the absorbance of the control tube, AE is the absorbance of the sample tube, VT is the total volume of enzyme extract (mL), Vs is the volume of enzyme used in the reaction (mL), W is the fresh weight of the sample (g).
Peroxidase (POD, E.C. 1.11.1.7) activity was determined using the guaiacol colorimetric method, with modifications to buffer composition and reagent concentrations[27]. Three biological replicates (n = 3) were prepared for each treatment and sampling day, with 0.5 g of petal tissue used per replicate. A blank control with buffer replacing enzyme extract was included for each set. All measurements were performed in triplicate. POD activity was defined as the enzyme quantity required to increase the absorbance at 470 nm by 0.01 per minute under the specified assay conditions, and expressed as one unit (U). POD activity was quantified using Eq. (6) outlined below:
$ \rm{P}OD\; activity\; (U\cdot min^{-1}\cdot g^{-1}\; FW)=\dfrac{A_{470}\times V_T}{0.01\times W\times V_S\times t} $ (6) where, A470 represents the total absorbance change at 470 nm throughout the reaction, Vt denotes the overall volume of the crude enzyme extract (mL), Vs corresponds to the aliquot volume of enzyme extract employed in the assay (mL), W indicates the fresh weight of the tissue sample (g), and t refers to the duration of the reaction (min).
MDA content
-
Malondialdehyde (MDA) content was determined using the thiobarbituric acid (TBA) method, with slight modifications to the original protocol[28]. For the extraction of MDA, 0.5 g of petal tissue was collected per sample (n = 3 biological replicates per treatment per day) and homogenized in 5 mL of 10% TCA on ice. A blank control (distilled water instead of extract) was included, and each sample was measured in triplicate.
The concentration of MDA was determined according to Eqs (7) and (8) provided below:
$\rm C\;({\text µ}mol\cdot L^{-1})= 6.45 \times (A_{532}-A_{600})-0.56 \times A_{450} $ (7) $ \rm MAD\;content\;({\text µ}mol\cdot L^{-1})=\dfrac{C\times V_T}{V_S\times W}$ (8) where, A532, A600, and A450 represent the absorbance values at 532, 600, and 450 nm, respectively, Vt denotes the total volume of the reaction mixture (mL), Vs corresponds to the volume of extract used in the assay (mL), and W refers to the fresh mass of the sample tissue (g).
Chlorophyll content
-
Chlorophyll content was determined using an acetone–ethanol (1:1, v/v) extraction method. Petal tissues (n = 3 per treatment·day-1) were weighed, cut into fine strips, and immersed in 20 mL of extraction solvent in sealed test tubes. Samples were incubated at room temperature in the dark until fully decolorized. A parallel extraction was performed using 95% ethanol following the same procedure. After extraction, the solvent volume was adjusted to 20 mL to account for evaporation loss. The absorbance of the supernatant was measured at 663 and 645 nm using a UV-Vis spectrophotometer. Distilled water served as the blank control, and all measurements were conducted in triplicate.
Chlorophyll content was calculated using the following Eqs (9)–(12):
$ \begin{matrix}{\text{Chl}}_{\text{a}}=\mathrm{12.7}\cdot {\text{OD}}_{\text{663}}-\mathrm{2.69}\cdot {\text{OD}}_{\text{645}}\end{matrix} $ (9) $ \begin{matrix}{\text{Chl}}_{\text{b}}=\mathrm{22.9}\cdot {\text{OD}}_{\text{645}}-\mathrm{4.86}\cdot {\text{OD}}_{\text{663}} \end{matrix} $ (10) $ \begin{matrix}{\text{C}}_{\text{v}}={\text{Chl}}_{\text{a}}+{\text{Chl}}_{\text{b}} \end{matrix} $ (11) $ \begin{matrix}{\text{C}}_{\text{A}}=\dfrac{{\text{C}}_{\text{V}}\times {5 \times 100}}{{1000 \times {\rm S}}}=\dfrac{{0.5 \cdot {\rm{Cv}}}}{\text{W}} \end{matrix} $ (12) where, CV is the chlorophyll concentration (μg·mL−1), CA represents the chlorophyll content per unit leaf area and per unit volume, respectively, S denotes the leaf area used for pigment extraction (cm2), and W represents the fresh mass of the analyzed sample (g).
Correlation analysis
-
Pearson's correlation coefficients were calculated between vase life and the physiological parameters SOD activity, POD activity, MDA content, and chlorophyll content using SPSS version 27.0. The correlation matrices were visualized as heatmaps with color gradients generated through the Chiplot online platform at
www.chiplot.online .Statistical analysis
-
Data analysis was conducted using SPSS version 27.0 and Excel 2021 to calculate means and standard deviations (SD). Each treatment for physiological and biochemical assays included three biological replicates. Prior to one-way analysis of variance (ANOVA), Levene's test was used to confirm homogeneity of variances across groups (p > 0.05 indicating equal variances). Significant differences between treatment groups were determined by one-way ANOVA followed by Tukey's multiple range test at p < 0.05. Figures illustrating these differences (a, b, c, etc.) were created using OriginPro 2021.
-
Different concentrations of 6-BA and GA3 significantly affected the duration of vase life and the maintenance of ornamental traits of A.andraeanum cut flowers (p < 0.05) (Table 2). The T3 was the most effective, extending vase life to 25.33 d, which was 14.3% (3.16 d) and 9.3% (2.16 d) longer than the CK1 and CK2 control groups, respectively. Similarly, T6 resulted in a vase life of 24.83 d, representing increases of 12.0% (2.66 d) and 7.2% (1.66 d) over CK1 and CK2. In contrast, low concentrations of GA3 (50–100 mg·L−1) significantly reduced vase life to 18.33–18.50 d, indicating a concentration-dependent effect of GA3 on senescence. Morphological observations revealed that even on Day 25, cut flowers in T3 and T6 retained acceptable ornamental quality and were markedly superior to the control groups, which showed clear signs of senescence (Fig. 1). The treated flowers exhibited relative maintenance of spathe size and color, and spadix integrity compared to the severely degraded controls. Collectively, the results indicate that 6-BA at 30 mg·L−1 effectively delays postharvest senescence and prolongs the period of acceptable visual quality, while GA3 exerts a positive effect only at higher concentrations (150 mg·L−1).
Table 2. Effects of plant growth regulators on the vase life of A. andraeanum cut flowers.
Treaments Vase life (d) CK1 22.17 ± 0.48c CK2 23.17 ± 0.60abc T1 23.83 ± 0.60abc T2 22.67 ± 0.49bc T3 25.33 ± 0.42a T4 18.50 ± 0.43d T5 18.33 ± 0.67d T6 24.83 ± 0.48ab Data are expressed as mean ± standard deviation. Different lowercase letters indicate significant differences at p < 0.05. 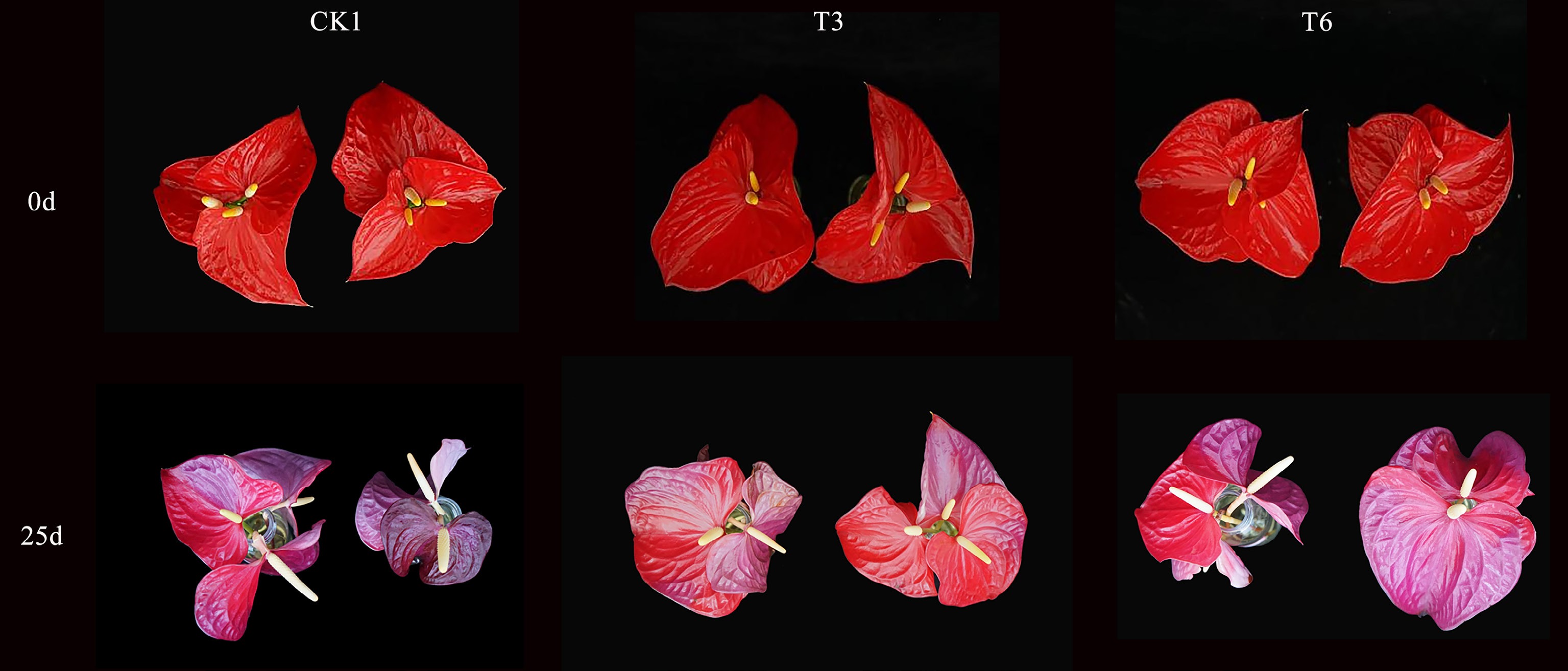
Figure 1.
Comparison of A. andraeanum cut flowers morphology on Days 0 and 25.
Postharvest rate of change of flower fresh weight and water balance of A. andraeanum cut flowers
-
Fresh weight change is a significant determinant of postharvest senescence in A. andraeanum cut flowers exhibited a marked decline in fresh weight due to water loss immediately after harvest. As shown in Fig. 2a, the fresh weight continuously declined during the vase period for all treatments, but with significant differences in the rate of decline. The control groups presented a striking contrast: CK1 suffered rapid and severe weight loss, whereas CK2 remained the most stable. Of the exogenous hormone treatments, T3 was the most outstanding, mirroring the stable pattern of CK2 and significantly alleviating the fresh weight loss. In contrast, T4 and T5 exhibited a precipitous drop in fresh weight, which led to the termination of their vase life before Day 20 (hence no subsequent data), underscoring their poor preservation effect. While T6 performed better than CK1, it was less effective than T3.
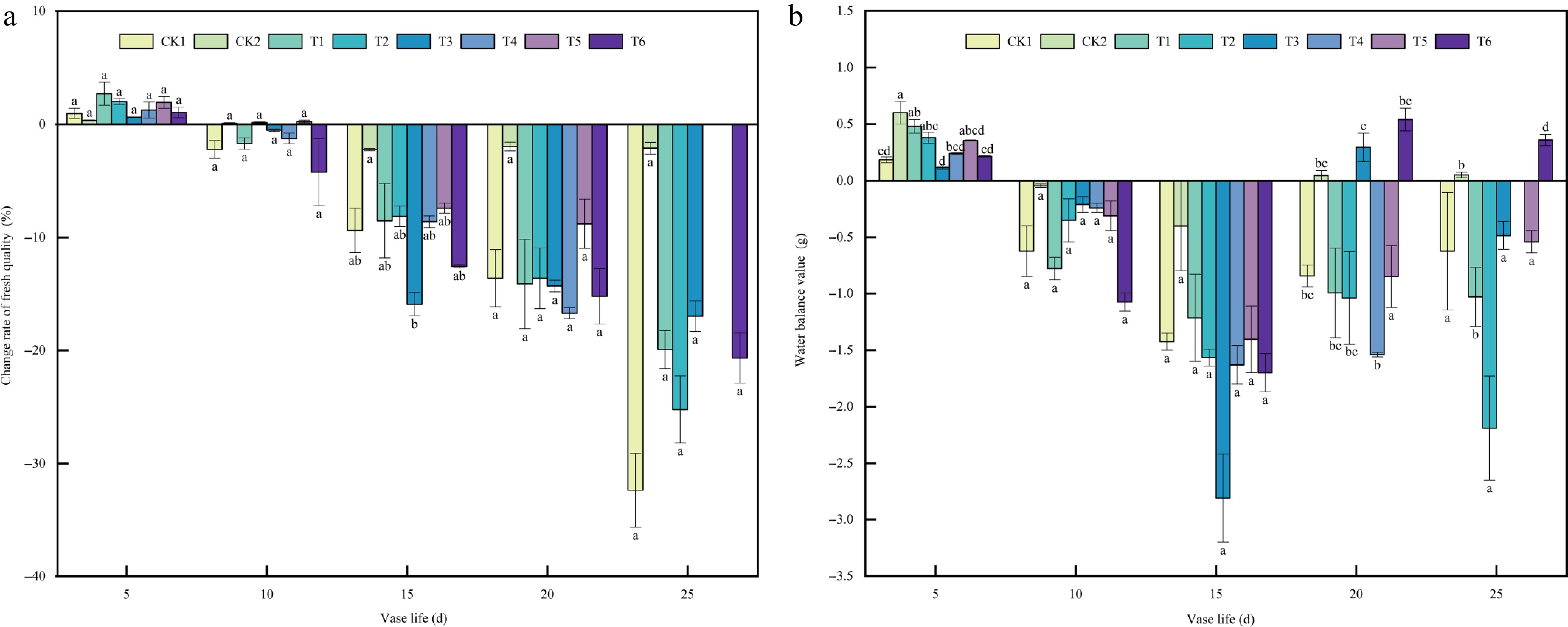
Figure 2.
Effects of plant growth regulators on (a) fresh weight variation, and (b) water balance in A. andraeanum cut flowers. Values are expressed as the mean ± SD (n = 3 biological replicates). Different lowercase letters indicate significant differences according to Tukey's multiple range test (p < 0.05). Treatments: CK1, base solution (20g·L−1 Sucrose + 200 mg·L-18-HQ); CK2, distilled water; T1, base solution + 10 mg·L−1 6-BA; T2, base solution + 20 mg·L−1 6-BA; T3, 30 mg·L−1 6-BA; T4, base solution + 50 mg·L−1 GA3; T5, base solution + 100 mg·L−1 GA3; T6, 150 mg·L−1 GA3.
Water balance reflects the dynamic relationship between water uptake and transpiration, serving as an essential parameter for evaluating water status in cut flowers. A positive water balance indicates that absorption exceeds transpiration, while a negative value reflects the opposite. The trends in water balance values closely mirrored those of fresh weight changes. Although all treatments initially exhibited a shift to negative water balance values, their recovery trends showed clear divergence in the later stages. Both T3 and T6 demonstrated superior recovery capacity, returning to positive water balance values by Day 20 after the mid-vase period decline. Furthermore, T6 exhibited more sustained improvement, maintaining the highest value until the end of the observation period. This performance was comparable to the stable control group CK2 and significantly better than treatments like T4 and T5, which showed irreversible deterioration. The ability of T3 and T6 to recover and maintain positive water balance conclusively demonstrates their significant efficacy in alleviating postharvest water stress in cut flowers (Fig. 2b).
Antioxidant enzyme activity in cut A. andraeanum flowers
-
As pivotal antioxidant enzymes in plants, SOD and POD play essential roles in maintaining redox homeostasis and delaying senescence. SOD functions as the first line of defense against oxidative stress by scavenging superoxide radicals (O2−), thereby mitigating ROS-induced cellular damage and contributing to stress tolerance and longevity regulation. POD, another key component of the antioxidant enzyme system, catalyzes the breakdown of hydrogen peroxide and participates in ROS scavenging and cell wall lignification, jointly enhancing the plant's ability to resist oxidative stress and postpone aging.
Variation trends of SOD and POD Activities are shown in Fig. 3a, b. During the early stage (Day 5), SOD and POD activities remained low across all treatments, with CK2 exhibiting significantly lower SOD activity, while POD activity showed no significant difference among treatments. By Day 10, both enzymes exhibited slight increases. SOD activity increased modestly in most hormone-treated groups, whereas POD activity rose more noticeably, with T1 reaching the highest level. At day 15, SOD activity peaked in T5 and T6, both being significantly higher than in the controls and other treatments, whereas POD activity declined overall without significant variation among treatments. By Day 20, both enzyme activities decreased, with SOD having the largest reduction in T3, while CK2 and T4 maintained relatively higher levels; POD activity also declined slightly, but T2 and T3 retained comparatively higher values. Toward the end of the vase period (Day 25), partial recovery of SOD activity was observed in several treatments, while POD activity remained low across groups. Overall, SOD activity displayed a rise-and-fall pattern with a distinct mid-term peak in T5 and T6, whereas POD activity had an early transient increase followed by a gradual decline, with CK2 peaking earlier and T3 maintaining moderate activity during the mid stage.
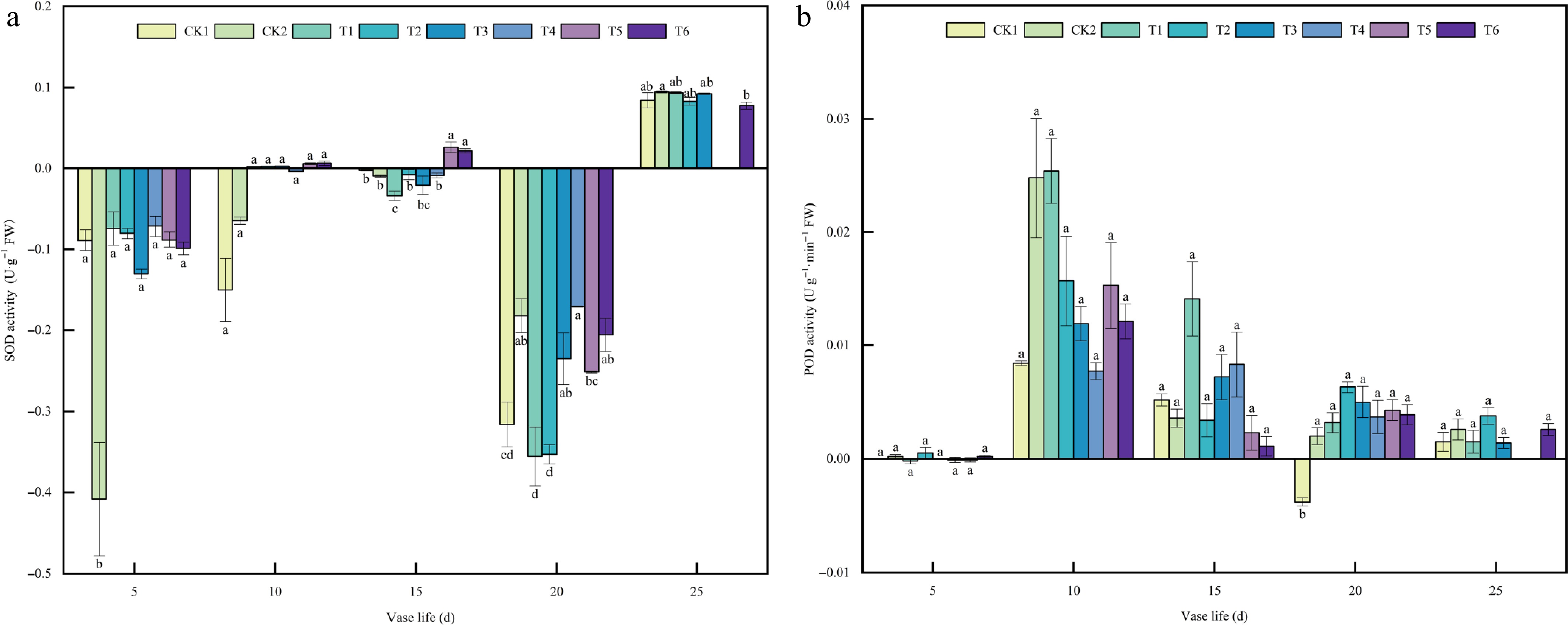
Figure 3.
Effects of plant growth regulators on (a) SOD, and (b) POD activity in A. andraeanum cut flowers. Values are expressed as the mean ± SD (n = 3 biological replicates). Different lowercase letters indicate significant differences according to Tukey's multiple range test (p < 0.05). Treatments: CK1, base solution (20g·L−1 Sucrose + 200 mg·L−18-HQ); CK2, distilled water; T1, base solution + 10 mg·L−1 6-BA; T2, base solution + 20 mg·L−1 6-BA; T3, 30 mg·L−1 6-BA; T4, base solution + 50 mg·L−1 GA3; T5, base solution + 100 mg·L−1 GA3; T6, 150 mg·L−1 GA3.
MDA content of A. andraeanum cut flowers
-
MDA, a primary byproduct resulting from lipid peroxidation within cellular membranes, serves as a key physiological indicator reflecting the extent of membrane oxidative damage and the progression of senescence in cut flowers. Generally, increased MDA levels indicate more severe membrane injury, which may accelerate the senescence or death of floral tissues. MDA content varied among treatments over the vase period (Fig. 4). At Day 5, T3 exhibited the highest MDA level, while T2 was the lowest, with the remaining treatments at intermediate levels. By day 10, MDA decreased in all treatments, with T6 maintaining a level similar to day 5. At Day 15, MDA increased in CK1, CK2, T1, T2, and T4, whereas it decreased in T3, T5, and T6; CK1 showed the largest increase, and T6 the smallest decrease. From Day 20 to 25, MDA increased in most treatments by Day 20 and then decreased by Day 25, with T3 and T6 showing substantial reductions and stabilizing at relatively low levels. Overall, MDA exhibited an early peak, mid-term fluctuations, and late-stage stabilization, with T3 and T6 maintaining relatively lower levels in the late stage.
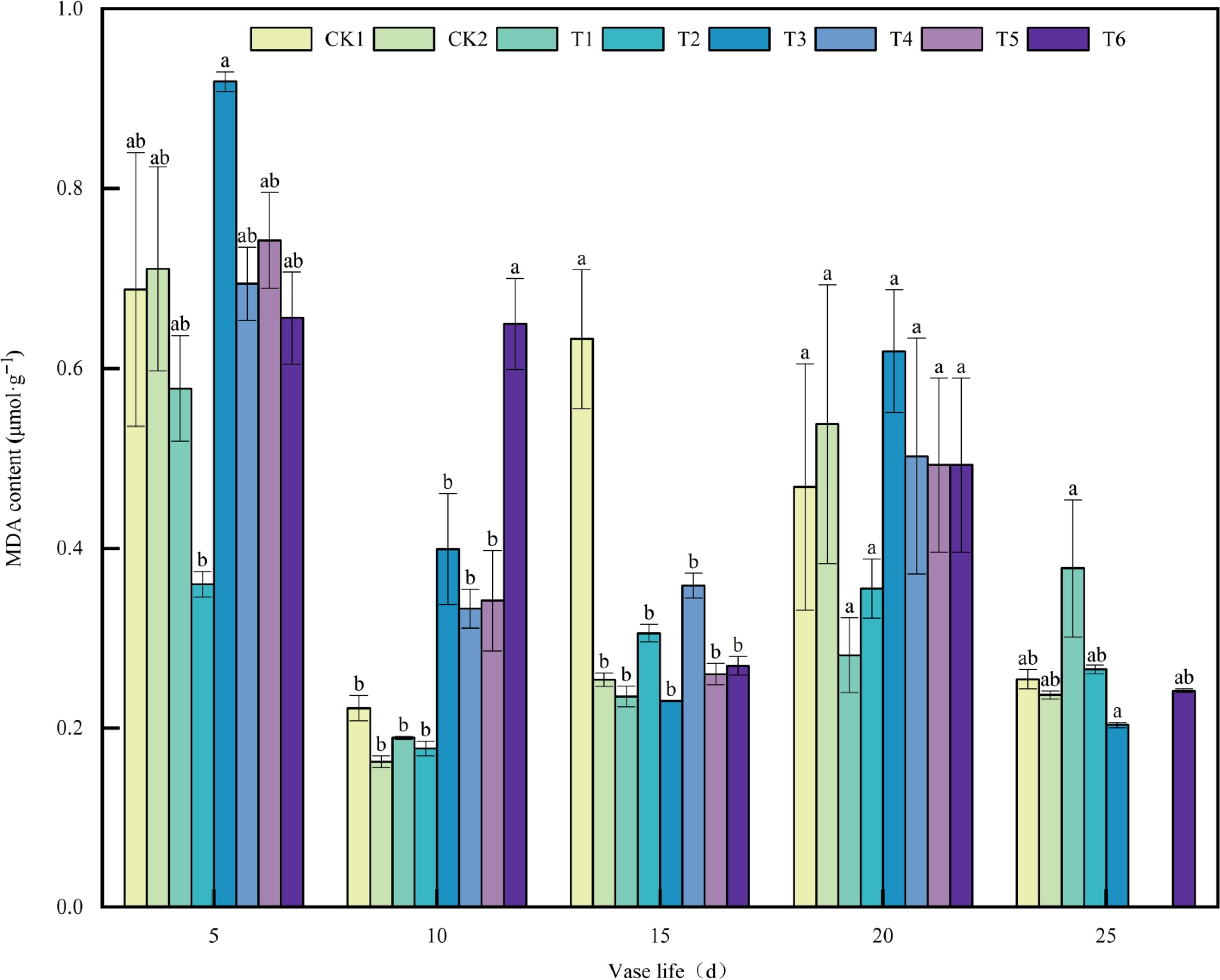
Figure 4.
Effects of plant growth regulators on MDA content in A. andraeanum cut flowers. Values are expressed as the mean ± SD (n = 3 biological replicates). Different lowercase letters indicate significant differences according to Tukey's multiple range test (p < 0.05). Treatments: CK1, base solution (20g·L−1 Sucrose + 200 mg·L−18-HQ); CK2, distilled water; T1, base solution + 10 mg·L−1 6-BA; T2, base solution + 20 mg·L−1 6-BA; T3, 30 mg·L−1 6-BA; T4, base solution + 50 mg·L−1 GA3; T5, base solution + 100 mg·L−1 GA3; T6, 150 mg·L−1 GA3.
Chlorophyll content of A. andraeanum cut flowers
-
Chlorophyll is an essential pigment for maintaining photosynthesis and the green coloration of floral organs in plants. Its content plays a pivotal role in determining the ornamental value and vase life of cut Anthurium flowers. As chlorophyll degradation is closely associated with the senescence process, changes in its content serve as an important physiological indicator of cut flower aging. Chlorophyll content showed dynamic changes among treatments throughout the vase period (Fig. 5). On Day 5, CK1 and T6 displayed the highest chlorophyll levels, whereas T1 and T3 were the lowest. By Day 10, most treatments experienced a decline, with T6 and T1 retaining relatively higher values. At day 15, T3 reached the peak chlorophyll content, followed by T6, while T5 showed the lowest level. On Day 20, chlorophyll slightly decreased across treatments, ranging from CK1 to T3. By Day 25, T3 maintained the highest content, with other treatments between 6.44 and 7.37 mg·g−1. Overall, chlorophyll content dropped sharply during the early stage, fluctuated moderately during the mid-stage, and stabilized in the late stage, with T3 and T6 retaining comparatively higher levels.
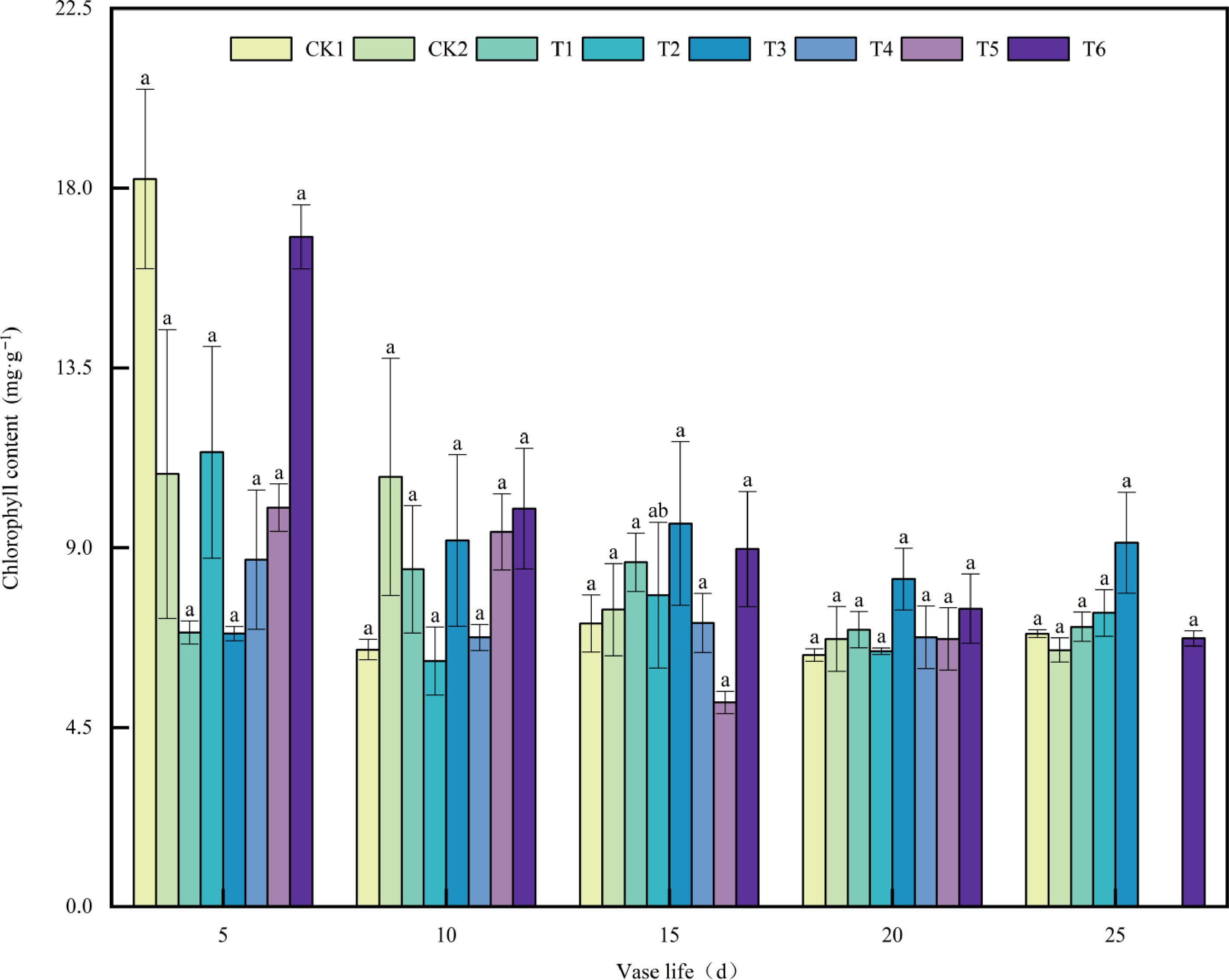
Figure 5.
Effects of plant growth regulators on chlorophyll content in A. andraeanum Cut Flowers. Values are expressed as the mean ± SD (n = 3 biological replicates). Different lowercase letters indicate significant differences according to Tukey's multiple range test (p < 0.05). Treatments: CK1, base solution (20g·L−1 Sucrose + 200 mg·L−18-HQ); CK2, distilled water; T1, base solution + 10 mg·L−1 6-BA; T2, base solution + 20 mg·L−1 6-BA; T3, 30 mg·L−1 6-BA; T4, base solution + 50 mg·L−1 GA3; T5, base solution + 100 mg·L−1 GA3; T6, 150 mg·L−1 GA3.
Correlation between vase life and physiological parameters across postharvest stages
-
To further investigate the relationship between vase life and physiological changes, Pearson correlation analyses were conducted between vase life and key parameters (SOD, POD, MDA, and chlorophyll) across different stages. As shown in Fig.6, in the first stage, no strong correlations were detected between vase life and any physiological parameter (all |r| < 0.4). In the second stage, vase life showed weak positive correlations with chlorophyll content (r = 0.318) and POD activity (r = 0.282). In the third stage, vase life was strongly and significantly correlated with chlorophyll content (r = 0.884, p < 0.05), whereas its correlations with other parameters remained weak. In the fourth stage, vase life maintained a moderate positive correlation with chlorophyll content (r = 0.563) but showed negative correlations with POD activity (r = –0.295) and MDA content (r = –0.221). These results suggest that chlorophyll content is most consistently associated with vase life, especially in the mid-to-late stages of postharvest.
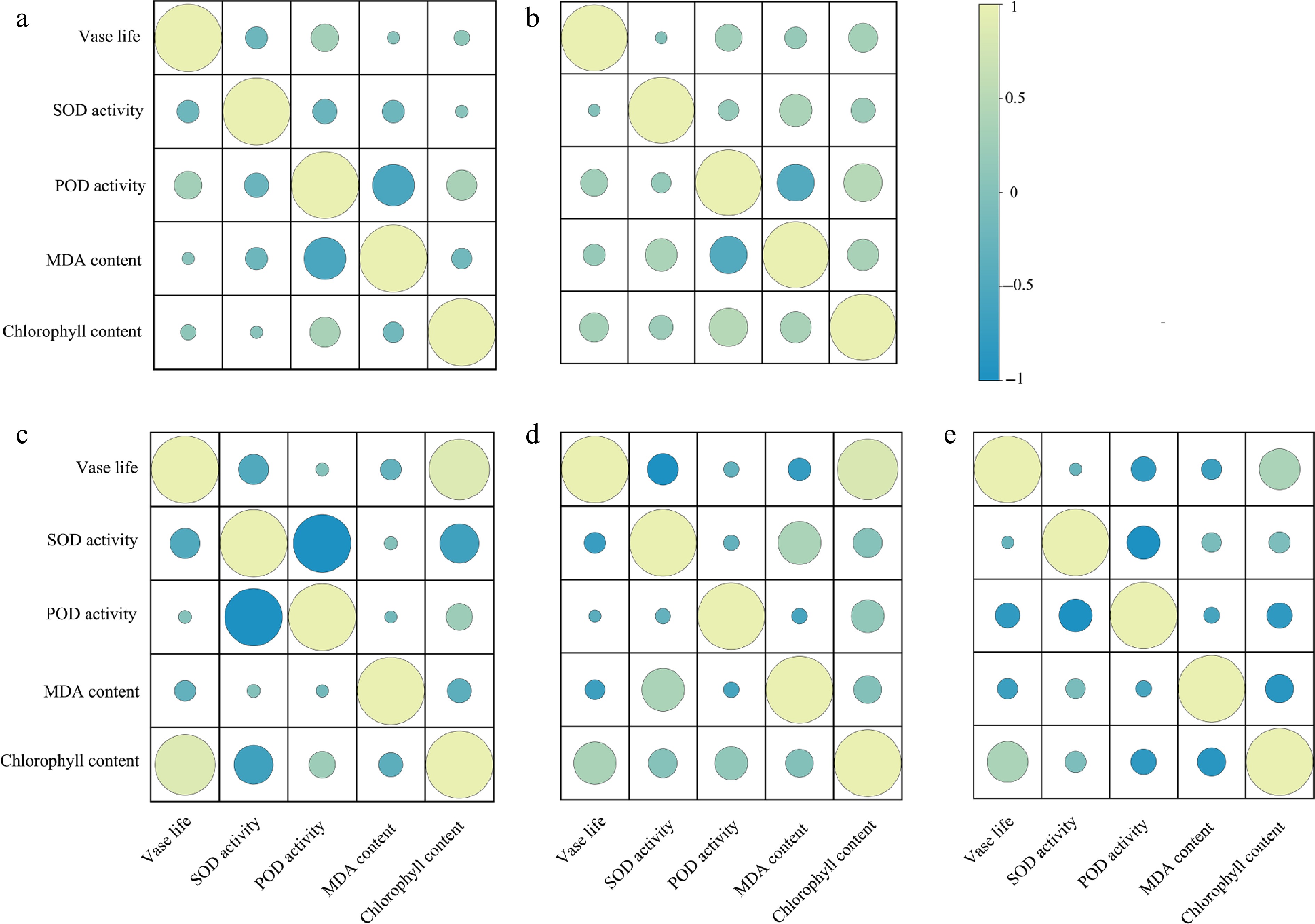
Figure 6.
Morphological changes of cut Anthurium flowers during vase life. (a) Day 5, (b) Day 10, (c) Day 15, (d) Day 20, and (e) Day 25 after vase placement.
-
Vase life is a crucial parameter for evaluating the effectiveness of cut flower preservative solutions. However, for long-lasting cut flowers such as A. andraeanum, where senescence progresses slowly, vase life alone may not comprehensively reflect their postharvest performance. Therefore, it is essential to consider physiological and biochemical changes during vase life, such as antioxidant enzyme activities, membrane lipid peroxidation, and chlorophyll content, to more accurately assess the effects of various treatments. During postharvest storage and vase display, Anthurium cut flowers often suffer from microbial proliferation and xylem blockage, which can accelerate senescence symptoms, including petal yellowing and spathe wilting[2]. Thus, the rational formulation of preservative solutions plays a critical role in prolonging vase life and enhancing ornamental quality.
Preservative solutions typically consist of water, sugars, antimicrobial agents, organic acids, and plant growth regulators. Among these, sugars provide respiratory substrates and help maintain osmotic balance, which are essential for delaying senescence[29]. Sucrose is the most commonly used carbon source in such solutions[30,31]. In addition, 8-HQ is widely used as an antimicrobial agent due to its ability to suppress bacterial growth in vase solutions. In this study, a base solution of sucrose and 8-HQ was supplemented with various concentrations of GA3 and 6-BA. Consistent with previous reports[32,33] GA3contributed to membrane stability, and 6-BA delayed organ senescence by regulating ROS metabolism. Antioxidant enzymes SOD and POD initially increased and then declined over vase life. Among all treatments, T3 (30 mg·L−1 6-BA) maintained significantly higher enzyme activities at all stages compared to the control, with a slower decline, indicating enhanced antioxidant defense. MDA, a widely recognized marker for lipid peroxidation, reflects the extent of membrane damage and is closely associated with the progression of floral senescence[34−36]. MDA content patterns in this study confirmed these trends: T3 (20 mg·L−1 6-BA) and T6 (150 mg·L−1 GA3) treatments resulted in significantly lower MDA accumulation during the late vase period compared with controls, suggesting a reduction in oxidative membrane damage. Chlorophyll content is another key physiological indicator of plant aging[37,38]. In the present study, chlorophyll levels in the spathes of the T3 group remained consistently higher than those in other treatments throughout the vase life, with an evident accumulation trend. This suggests that exogenous application of 6-BA effectively suppressed chlorophyll degradation, delayed yellowing, and maintained tissue greenness, thus enhancing ornamental quality. These findings are consistent with previous reports showing that chlorophyll breakdown is closely associated with spathe yellowing and the decline of ornamental quality in A. andraeanum, supporting the reliability of chlorophyll as an indicator for evaluating postharvest senescence in this species. Similarly, other studies have reported that hormone treatments also inhibited chlorophyll degradation in Lilium cut flowers[39], supporting the present results.
Overall, both GA3 and 6-BA were effective in improving vase life and ornamental performance in Anthurium cut flowers, with T3 showing the most consistent benefits. However, it is worth noting that stem yellowing was observed in some hormone-treated groups, and the underlying mechanism remains unclear, warranting further investigation. This phenomenon could be related to high hormone concentrations, altering vascular tissue activity, or disrupting the balance of growth regulators. Future studies could investigate the underlying physiological and molecular mechanisms and explore whether combining GA3 and 6-BA at optimized concentrations could mitigate such adverse effects while maintaining or enhancing vase life. In addition, the regulatory mechanisms of GA3 and 6-BA on ethylene biosynthesis and signaling are not fully elucidated, and future research at the molecular level is needed to better understand their modes of action.
-
Exogenous application of plant growth regulators significantly contributed to prolonging vase life, enhancing ornamental quality, and maintaining physiological and metabolic stability in A. andraeanum cut flowers. Both GA3 and 6-BA treatments were effective in promoting water balance, delaying yellowing and fresh weight loss, and enhancing the activities of antioxidant enzymes such as SOD and POD. These effects improved the scavenging of ROS, inhibited the accumulation of MDA, and consequently reduced lipid peroxidation and oxidative damage to cell membranes. Among all treatments, 30 mg·L−1 6-BA showed the most pronounced preservative effect, characterized by stable fresh weight changes, prolonged maintenance of antioxidant enzyme activity, the slowest rate of chlorophyll degradation, and a vase life extended by 14.26% compared to the control. Correlation analysis further indicated that chlorophyll content was closely associated with vase life, highlighting its potential as a key physiological indicator for evaluating postharvest quality. These findings provide both theoretical support and technical guidance for optimizing preservative formulations and promoting the commercial application of Anthurium cut flower preservation technology, with promising practical implications.
We would like to thank the editor and reviewers for their helpful comments on the manuscript. The Project of National Key R & D Program (Grant No. 2023YFD1600504), Fujian Provincial Natural Science Foundation of China (Grant Nos 2023J01283, 2022J01639), the National Natural Science Foundation of China (Grant Nos 32101583, 31901353), the Innovation and Application Engineering Technology Research Center of Ornamental Plant Germplasm Resources in Fujian Province (Grant No. 115-PTJH16005), the key research and development program of the Ningxia Hui Autonomous Region in China (Grant No. 2022BBF02041).
-
The authors confirm contribution to the paper as follows: study conception and design, methodology, supervision: Zhao K, Zhou Y; data curation, draft manuscript preparation: Tong E, Zhan S; resources: Zhu W; writing − review and editing: Huang R, Zhao Y, Wen Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and also available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Ruiliu Huang, Yuqing Zhao
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Huang R, Zhao Y, Wen Y, Tong E, Zhan S, et al. 2026. Effects of exogenous 6-BA and GA3 on vase life and postharvest physiology of Anthurium andraeanum cut flowers. Technology in Horticulture 6: e003 doi: 10.48130/tihort-0025-0038 

Effects of exogenous 6-BA and GA3 on vase life and postharvest physiology of Anthurium andraeanum cut flowers
- Received: 29 June 2025
- Revised: 29 October 2025
- Accepted: 06 November 2025
- Published online: 28 January 2026
Abstract: Anthurium andraeanum is a prominent cut flower species widely cultivated for ornamental purposes. To investigate the effects of exogenously applied plant hormones on postharvest longevity and senescence in cut flowers, a preservative solution containing sucrose and 8-hydroxyquinoline (8-HQ) was supplemented with varying concentrations of 6-benzylaminopurine (6-BA) and gibberellic acid (GA3). Changes in fresh weight, antioxidant enzyme activities, malondialdehyde (MDA) content, and chlorophyll levels were systematically monitored. Both 6-BA and GA3 treatments significantly extended vase life, enhanced antioxidant capacity, reduced MDA accumulation, and delayed chlorophyll degradation. The most pronounced effect was observed with 30 mg·L−1 6-BA, which extended vase life to 25.33 d—a 14.26% increase compared to the control—while maintaining higher peroxidase (POD) activity and increasing chlorophyll content. These results provide a theoretical foundation for improving the postharvest quality and storage of A. andraeanum cut flowers.
-
Key words:
- Anthurium andraeanum /
- Cut flowers /
- 6-benzyladenine (6-BA) /
- Gibberellic Acid (GA3) /
- Vase life