









-
Plants are constantly challenged by various biotic and abiotic stresses, resulting in yield loss of crops[1]. Numerous investigations have revealed that plants possess the capacity to establish adaptive defense mechanisms in response to diverse biotic and abiotic stressors. The primary cellular reaction is the immediate synthesis of reactive oxygen species (ROS), commonly referred to as oxidative burst[2].There are proteins in plants that control the burst of ROS and protect plants from harm[3,4]. Respiratory burst oxidase homologs (Rbohs), which operate as nicotinamide adenine dinucleotide phosphate (NADPH) oxidases, have been extensively investigated for their role in the enzymatic generation of reactive oxygen species[5]. Rboh proteins are absent in prokaryotes, but found in a wide variety of eukaryotes[6]. Mounting evidence indicates that plant Rboh proteins play crucial roles through regulating ROS production in plant ontogeny and its associated mechanisms for coping with biotic and abiotic challenges[7−9]. ROS scavenging enzymes such as superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), and ascorbate peroxidase (APX) play an essential role in regulating ROS levels and oxidative damage[10].
A wide range of plant species have been found to possess Rboh genes, such as Arabidopsis, tomato, tobacco, potato, maize, watermelon, barley, alfalfa, grape, and cotton.[11,12]. The Arabidopsis Rboh family has 10 members, AtRbohA-J, which were best characterized and demonstrated to be involved in a variety of physiologic processes[13,14]. Among them, AtRbohD exhibited constitutive expression across all tissues and demonstrated pronounced responsiveness to various stress conditions[15−17]. In the atrbohD mutant with AtRbohD gene deletion, ROS production failed to be induced by inoculation with the fungus Alternaria brassicicola, indicating that the ROS induced by pathogen infection was AtRbohD-dependent[18]. Currently, AtRbohD-dependent ROS production was considered to be a ubiquitous mechanism in response to pathogen attack as a key component of plant innate immunity[19]. Based on plant innate immunity theory, the detection of pathogen-associated molecular patterns (PAMPs) by cell surface-localized pattern recognition receptors (PRRs) initiates PAMP-triggered immunity (PTI). Conversely, the recognition of microbial effectors introduced into plant cells by resistance (R) proteins activates effector-triggered immunity (ETI), typically associated with a hypersensitive response (HR)[20]. During both PTI and ETI, ROS production was predominantly dependent on RbohD[21]. In PTI, fungal chitin and bacterial flagellin, the typic PAMPs from fungal and bacterial pathogens, respectively, could trigger AtRbohD-dependent ROS production[22−25], which might regulate lignin biosynthesis, and callose deposition[5,26,27]. In ETI, AtRbohD was associated with HR in response to Pst.DC3000 expressing effector factor avrRpm1, showing reduced cell death in atrbohD mutants after inoculation with Pst.DC3000 (avrRpm1)[14]. Moreover, atrbohD mutants showed different responses in different plant-pathogen systems[28]. For instance, atrbohD mutants showed increased resistance to biotrophic oomycete Peronospora parasitica Emco5 and necrotrophic fungus Alternaria brassicicola[14,18], compared to the wild type, but were more susceptible compared to wild-type to the biotrophic fungus Golovinomyces cichoracearum, and necrotrophic bacterium Dickeya dadantii[17,29,30]. These findings underscore the complex regulatory role of AtRbohD-mediated ROS generation in modulating plant-pathogen interactions.
Emerging evidence has demonstrated the involvement of RbohD in plant responses to diverse abiotic stresses. In Arabidopsis, the ERF74-RbohD-ROS signaling cascade has been identified as a key mediator of osmotic and hypoxic stress responses[31]. Salinity stress upregulates RbohD expression through ABI4, a central component of abscisic acid signaling, leading to ROS overaccumulation that ultimately causes membrane damage and reduced seed vigor[32]. RbohD collaborates with the pattern recognition receptor FLS2 to coordinate drought and salt stress adaptation through modulating the expression of phytochrome-interacting factor 4 (PIF4), and fine-tuning the accumulation of stress-related metabolites and associated biosynthetic gene expression[33,34]. However, another study revealed that salt stress-induced ROS production depends on AtECA4 and clathrin-mediated trafficking of plasma membrane-localized RbohD, a process distinct from flg22-triggered immune responses[3]. These findings collectively demonstrate that RbohD orchestrates distinct signaling pathways in response to different abiotic stresses, highlighting its multifaceted roles in stress adaptation.
Several studies have demonstrated that RbohD-dependent ROS production is involved in the signal pathways mediated by different phytohormones under different stresses. In Chinese white pear, it was reported that the transcription of PbRbohD was upregulated by SA, ABA, and MeJA[35]. In cell death induced by the potato Ny-1 gene against potato virus Y, the expression of RbohD occurs specifically on the lesion border, and SA was required for the responses in both the cell death zone and the surrounding tissue, suggesting that RbohD was required for the spatial accumulation of SA and conversely regulated by SA transcriptionally[36]. The flg22-triggered oxidative burst at an early stage of the plant immune response was dependent on RbohD and regulated by ET signaling[37]. Ethylene response factor 74 (ERF74) could directly bind to the promoter of RbohD and activate its expression under different abiotic stresses[31]. Notably, the coordinated action of AtRbohD and AtRbohF contributes significantly to stomatal defense mechanisms by modulating stomatal closure via ABA signaling cascades[37−39]. Recently, it was also found that RbohD modulated ROS metabolism during seed germination under salinity stress along with Abscisic acid 4 (ABI4), a key component in ABA signaling, and Vitamin C Defective 2 (VTC2), another component of ROS production and clearance[32].
As the exclusive source of natural rubber for commercial purposes, Hevea brasiliensis holds significant economic value across tropical nations. In NR production, rubber trees often suffer from various biotic and abiotic stresses, which results in NR yield loss every year[40,41]. Consequently, it is of great significance to investigate the molecular basis of rubber tree stress tolerance for developing novel resistance strategies. It was reported that ROS signaling was involved in innate immunity in rubber tree[42], and 10 genes encoding Rboh proteins were identified in the rubber tree genome[43]. However, the functions of these Rbohs in disease resistance remain elusive. In this study, a homolog of RbohD in rubber tree (HbRbohD) was identified, and its expression profiles, subcellular localization, and ROS-induced ability investigated. Furthermore, the functions of HbRbohD in disease resistance and salinity tolerance were determined in HbRbohD transgenic plants. The results provide valuable information for future research on biotic and abiotic responses mediated by HbRbohD-dependent ROS production in rubber trees.
-
Hevea brasiliensis (Reyan 7-33-97) seedlings were cultivated in potting soil at 28 °C. The leaves of rubber trees, during the light green period, were used in this study. Arabidopsis thaliana Columbia ecotype, and Nicotiana benthamiana grew in a growth chamber set at 22 °C with 60% relative humidity, under a 12 h light/24 h cycle provided by fluorescent lamps (200 μE/m2/s).
Isolation and sequence analysis of the HbRbohD gene
-
Total RNA of rubber tree leaves was extracted using an RNA Extraction Kit (Tiangen DP441) . The first-strand cDNA was obtained by using Revert Aid First Strand cDNA Synthesis Kit (Thermo Fisher K1621). To identify potential homologs of AtRbohD (AT5G47910) in rubber tree, a BlastX analysis was conducted against the transcriptome database using its nucleotide sequence as the query. Among the candidate sequences, Refseq XM_021809230.1 exhibited significant homology and was subsequently designated as HbRbohD. The full-length CDS sequence of HbRbohD was amplified using the first-strand cDNA as a template and verified by sequencing. The amino acid sequences of HbRbohD were deduced by DNAMAN software. The conserved domains were predicted using the SMART website (
http://smart.embl-heidelberg.de/ ). The multiple alignments of amino acid sequences were performed using ESPript 3.0 (http://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi ), and GeneDoc 2.7.0. The neighbor-joining phylogenetic tree with bootstrap values was generated through Clustal X 2.0 and MEGA 7.0.Pathogen inoculation
-
For the expression patterns assay of HbRbohD, Colletotrichum gloeosporioides (anthracnose pathogen), and Oidium heveae Steinm (powdery mildew pathogen) were used to infect the leaves of the rubber tree during the light green period. The grafted seedlings of rubber tree clone Reyan7-33-97 were inoculated by foliar spraying of C. gloeosporioides spore suspension (5 × 105 spores/mL) and O. heveae Steinmann spore suspension (4 × 105 spores/mL) respectively, and the seedlings without inoculation were used as control. The control and inoculated seedlings were cultivated under greenhouse conditions at 25 °C with 90% relative humidity. Leaf samples were collected at 0, 1, 2, 3, and 5 d post-inoculation (dpi), immediately flash-frozen in liquid nitrogen, and preserved at −80 °C for subsequent RNA isolation. Each biological replicate consisted of two leaves pooled from three individual seedlings, with no repeated sampling from the same plant.
Botrytis cinerea was used for the disease assay of HbRbohD overexpression lines. B. cinerea conidia were collected and suspended with suspension (1% Sabouraud Maltose Broth + 0.05% Tween 20) at 2.5 × 105 conidia/mL 5 μL droplets of conidia suspension were inoculated on 4-week-old leaves of A. thaliana and preserved with plastic wrap for 3 or 5 d. The size of the spots was quantified through diameter measurements and subsequent analysis.
Chitin and Flg22 treatment
-
According to the protocol previously reported by Zhang et al., mesophyll protoplasts were prepared from rubber tree leaves[44−48]. 10−5 μg/mL of flg22 and 200 μg/mL of chitin were used to treat the protoplast cells, respectively. Mesophyll protoplasts treated with flg22 and chitin for 0, 5, 10, and 15 min were collected for RNA extraction.
Phytohormone treatment
-
For phytohormone application, Reyan7-33-97 seedlings were foliar-treated with aqueous solutions containing 5 mM salicylic acid (SA), 1 mM methyl jasmonate (MeJA), 0.5 mM ethephon (ET), 50 μM abscisic acid (ABA), or 200 μM gibberellic acid (GA), following the methodology described by Yang et al.[49]. All solutions were prepared using distilled water supplemented with 0.1% (v/v) ethanol as a solvent. Control plants received 0.1% (v/v) ethanol aqueous solution only. The treated seedlings were maintained under greenhouse conditions at 25 °C. Leaf samples were collected at 0, 12, 24, and 48 h post-treatment, immediately flash-frozen in liquid nitrogen, and stored at −80 °C for subsequent RNA isolation. Each biological replicate consisted of two leaves pooled from three individual seedlings, with no repeated sampling from the same plant.
Salt treatment
-
The suspension callus of the Brazilian Reyan 7-33-97 variety was produced according to the previously published study by Luan et al.[50]. The suspension callus cultured on the solid medium for 7 d was transferred to a liquid medium containing 150 mmol/L NaCl and subjected to shaking culture. The suspended callus in the medium was collected at 0, 10, 30, and 60 min, respectively. The suspended callus growing in salt-free culture medium was used as a control.
RNA isolation, cDNA synthesis, and qRT-PCR
-
A commercial kit designed for polysaccharide- and polyphenol-rich tissues (Tiangen, DP441) was used for total RNA isolation using First-strand cDNA was synthesized according to the manufacturer's protocol of a reverse transcription system (Thermo Fisher K1691). Real-time quantitative PCR was conducted with a SYBR Green-based detection system (Vazyme Biotech, Q711-02) on a LightCycler 96 platform (Roche). Gene expression quantification was performed by normalizing to either baseline levels (0 d) or untreated controls. The transcript abundance of HbRbohD was determined through comparative Cᴛ (2−ΔΔCᴛ) analysis, with HbActin serving as the endogenous control for data normalization.
Subcellular localization analysis
-
The coding sequence of the HbRbohD gene was amplified and inserted into the transient expression vector pEGAD-GFP for subcellular localization analysis. The recombinant plasmid pEGAD-HbRbohD-GFP was verified by sequencing. Genetic transformation was performed via Agrobacterium tumefaciens-mediated infiltration, with red fluorescent MIEL-RFP[51], and green fluorescent pEGAD-GFP empty vector serving as dual-spectral reference markers. The distribution of fluorescence in leaves of Nicotiana benthamiana was observed by laser confocal microscopy (Leica TCS SP8).
Transient expression and ROS measurement in mesophyll protoplasts of rubber tree
-
To investigate the function of HbRbohD on ROS production and accumulation, the coding region of HbRbohD was ligated into pUC19-35S-Flag vector and transiently expressed in rubber tree mesophyll protoplasts. The isolation and transformation of rubber tree mesophyll protoplasts refers to the previous method by Zhang et al.[44]. Following a 2-h transformation period, intracellular ROS levels were quantified at 3-min intervals over a 15-min duration using the Reactive Oxygen Species Assay Kit (Beyotime Institute of Biotechnology, Haimen, China) in accordance with the manufacturer's protocol. Fluorescence measurements were obtained at an excitation wavelength of 485 nm, and an emission wavelength of 530 nm using a microplate fluorescence reader.
Construction of HbRbohD overexpressing against Col-0 background
-
The binary recombinant vector pCAMBIA1300-HbRbohD-HA was constructed and transformed into Agrobacterium tumefaciens GV3101. The A. tumefaciens GV3101 harboring the HbRbohD gene was transformed into Col-0 using the A. tumefaciens-mediated flower dip method. Positive T1-generation transgenic lines were screened with 30 mg/L hygromycin. Genomic DNA from positive transgenic lines was extracted and PCR was performed by using an amplification primer pair for the HbRbohD gene. The positive transgenic seedlings detected by PCR were verified by semi-quantitative RT-PCR with HbActins as the reference gene.
Determination of ROS antioxidant enzyme activity, and malondialdehyde (MDA) concentration
-
The fresh Arabidopsis leaf inoculated with B. cinerea for 2 d (1 g) was homogenized with 10 ml of 25 mM potassium phosphate buffer (pH 7.4). The homogenate was centrifuged at 10,000 g for 25 min at 4 °C, and the supernatant was used as an enzyme extract to determine the activity of ROS scavenging enzymes. The activity of SOD, POD, and CAT was measured according to the manufacturer's instructions of the Total Superoxide Dismutase (T-SOD) assay kit (A001-1-2, Nanjing Jiancheng, China), Peroxidase assay kit (A084-3-1 Nanjing Jiancheng, China), and Catalase (CAT) assay kit (A007-1-1, Nanjing Jiancheng, China) on a Multiskan Spectrum (Infinite M200 PRO, Tecan, Austria). Malondialdehyde content was quantified according to the manufacturer's instructions of the Malondialdehyde (MDA) assay kit (A003-1-2, Nanjing Jiancheng, China).
Seed germination analysis
-
The seeds of Arabidopsis were subjected to surface sterilization by immersion in 70% ethanol for 1 min, followed by treatment with a 1% sodium hypochlorite solution for 10 min. Subsequently, the seeds were thoroughly rinsed with autoclaved distilled water a minimum of five times. The sterilized seeds were then transferred onto ½ MS medium supplemented with 200 mM mannitol and 150 mM sodium chloride. The plates were stratified in darkness at 4 °C for a period of 48 h. Then, the seeds were transferred to a growth chamber at 22 °C under 24 h light conditions. Seed germination was determined by radicle emergence. For each germination test, more than 50 seeds per genotype were used, and three experimental replications were performed.
Statistical analysis
-
Statistical analysis was performed with SPSS version 10.0 (SPSS Inc., Chicago, USA). The data for quantitative RT-PCR, disease test, ROS production and accumulation, seed germination, cotyledon greening, defense enzyme activity, and MDA content were analyzed based on Student's t-test or one-way ANOVA with Duncan's multiple range tests. Different letters labeled on the bar graphs in the figure indicate significant differences (p < 0.05).
-
The nucleotide sequence of Arabidopsis respiratory burst oxidase homolog D (AtRbohD) (AT5G47910) was used as a query to perform a BlastX search against the NCBI database to obtain the homolog of RbohD in rubber tree. Refseq No. XP_021679868.1 was selected as a candidate and named as HbRbohD. The full length CDS of HbRbohD was amplified by RT-PCR, and the sequence was verified by sequencing. The full-length CDS of HbRbohD was 2,733 bp, encoding a 910 amino acid polypeptide with a 102.5 kDa molecular weight. The amino acid alignment showed that HbRbohD, as well as the known RbohD from the other plants, contained NADPH_Ox, EF-hand, and NAD-binding conserved domains and had the highest homology with RbohD from Arabidopsis (Fig. 1a). All ten RBOH proteins from A. thaliana and HbRbohD were used to generate a Neighbour-joining tree (Fig. 1b). Phylogenetic tree analysis revealed that HbRbohD was clustered in the same branch as AtRbohD, suggesting that HbRbohD was the homolog of AtRbohD in rubber tree.
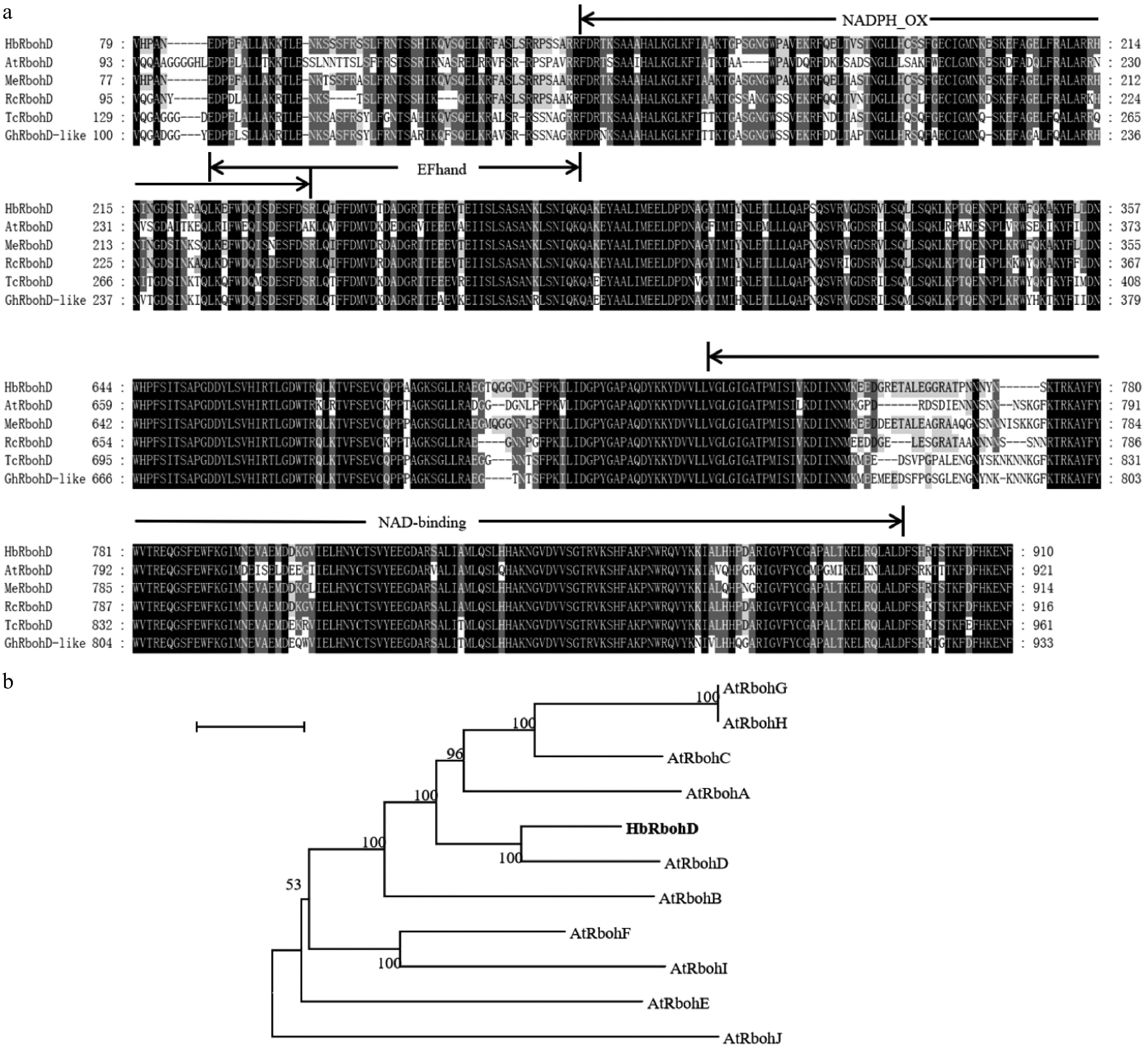
Figure 1.
Multiple sequence alignment and phylogenetic analysis of HbRbohD. (a) Comparative analysis of RbohD homologs across diverse plant species. Conserved domains are highlighted, with regions identical in all examined sequences shown in black, while residues matching HbRbohD are denoted in gray. AtRbohD was from Arabidopsis thaliana (NP_199602), MeRbohD was from (XP_021624267), RcRbohD was from Ricinus communis (XP_015570593), TcRbohD was from (EOY14749), GhRbohD-like was from (XP_040943195). (b) Phylogenetic tree of HbRbohD with Rboh proteins from A. thaliana, including AtRbohA (NP_196356), AtRbohB (NP_172383), AtRbohC (NP_199919), AtRbohD (NP_199602), AtRbohE (NP_173357), AtRbohF (NP_564821), AtRbohG (NP_194239), AtRbohH (NP_200809), AtRbohI (NP_192862), AtRbohJ (NP_190167).
Cis-elements assay
-
To examine the potential role of HbRbohD in stress responses and its association with phytohormone-mediated signaling cascades, Plantcare online tools were used to analyze the cis-elements of the promoter region located 3,000 bp upstream of the HbRbohD coding region. The results showed that the cis-elements include MYB and MYC elements, CAAT-box, box-4, and ARE elements. In addition, the promoter of HbRbohD contained different stress response elements and phytohormone response elements, such as the GA-responsive element (GARE), ABA-responsive element (ABRE), SA-responsive element (SARE), defense and stress responsive element (TC-rich repeats), and low-temperature response element (LTR) (Supplementary Table S1). These results indicate that HbRbohD might be involved in growth development, biotic and abiotic stress responses in rubber tree.
HbRbohD responded to stresses and phytohormone treatments
-
To investigate the potential roles of HbRbohD in the responses to biotic and abiotic stresses, the expression patterns of HbRbohD were analyzed under conditions of phytopathogen infection, salt and drought treatments. The expression levels of HbRbohD were monitored at various time points post-inoculation with C. gloeosporioides and O. heveae Steinm, which cause anthracnose and powdery mildew, respectively. The results showed a significant up-regulation of HbRbohD at 72 h post-inoculation (hpi) with both C. gloeosporioides and O. heveae Steinm (Fig. 2a & b). Treatment with typical PAMPs chitin and flg22 triggered the upregulation of HbRbohD gene expression in mesophyll protoplasts of rubber trees (Fig. 2c & d). In addition, HbRbohD expression was significantly up-regulated in suspension callus culture at 30 min post-treatment with salt (Fig. 2e). These findings suggest that HbRbohD is involved in immune response and salt tolerance of rubber trees.
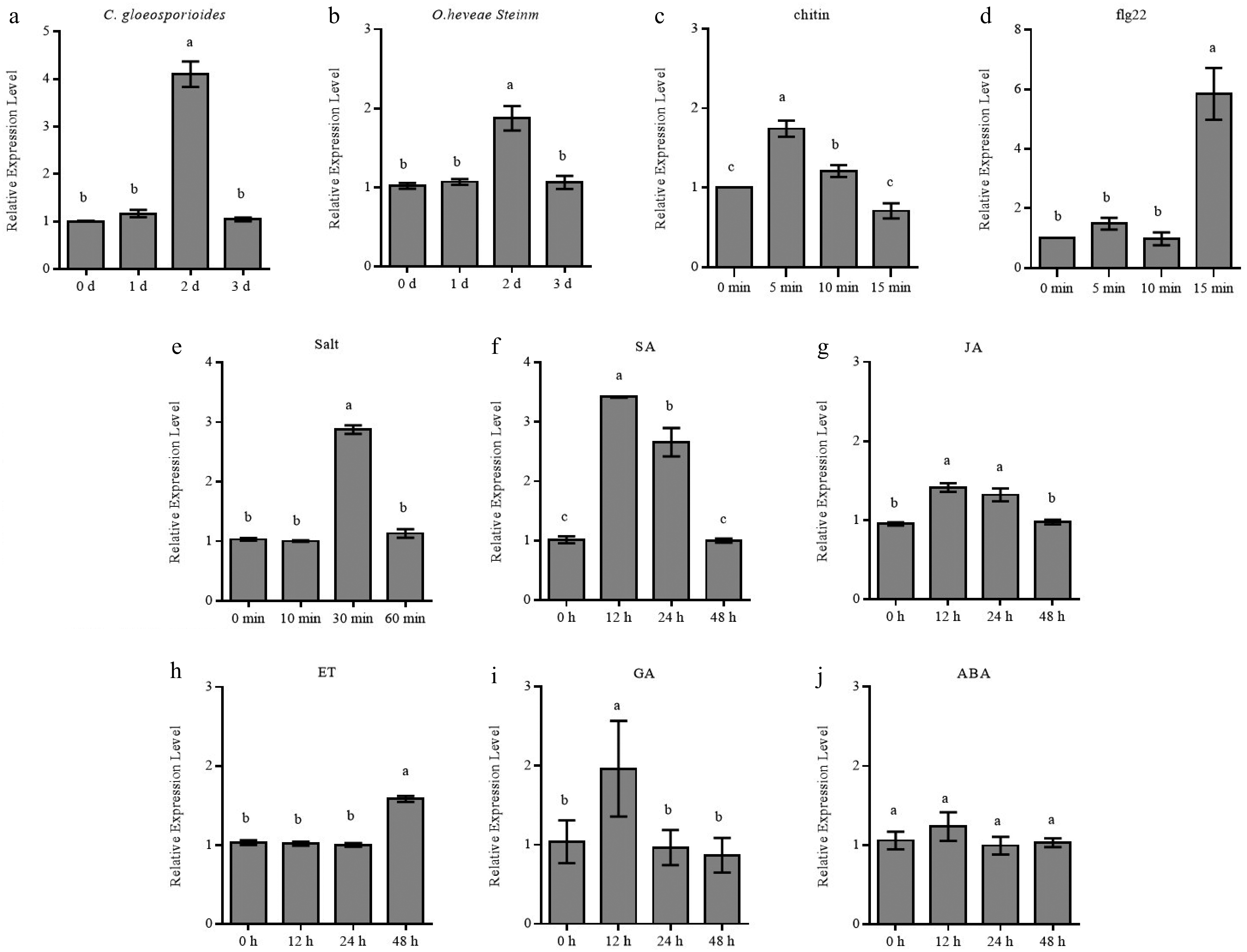
Figure 2.
Expression patterns of HbRbohD in response to stresses and phytohormone treatments. The expressions of HbRbohD in rubber tree leaves inoculation with (a) C. gloeosporioides, and (b) O. heveae Steinm, in rubber tree mesophyll protoplasm treated with (c) chitin, and (d) flg22, in rubber tree suspension callus cultures treated with (e) salt, and treatment with various phytohormone, such as (f) SA, (g) JA, (h) ET, (i) GA, and (j) ABA were quantitatively assessed through qRT-PCR analysis, with transcript abundance being normalized against baseline measurements obtained at day 0 (control). The expression level was normalized using actin as housekeeping gene. Data are shown as the means ± SD from three independent experiments and the mean values assigned dissimilar superscript letters differ significantly (p < 0.05).
To investigate the role of HbRbohD in various phytohormone-mediated signaling pathways, its expression changes in rubber seedlings treated with different phytohormones were examined. The results revealed that expression of HbRbohD was rapidly and significantly up-regulated by more than three times after 12 h treatment with SA (Fig. 2f), and increased by about 1–2 times after treatments with JA, ET, and GA (Fig. 2g–i), but there were no significant changes after treatment with ABA (Fig. 2j). These results suggest that HbRbohD plays a broad role in multiple phytohormone signaling pathways in rubber trees, with a particularly strong involvement in SA-mediated signaling.
HbRbohD is located on the plasma membrane, and induces ROS accumulation
-
To determine the subcellular localization of HbRbohD, HbRbohD was fused with GFP and transiently expressed in N. benthamiana leaves via Agrobacterium-mediated infiltration. MIEL-RFP was the marker of plasma membrane and nucleus[51]. In tobacco leaves expressing GFP and MIEL-RFP, fluorescence of GFP was uniformly distributed throughout the entire cells and MIEL-RFP was mainly located on plasma membrane and nucleus. When HbRbohD-GFP fusion and nucleocytoplasmicmarker (MIEL1-RFP) were transiently co-expressed in N. benthamiana leaves, the HbRbohD-GFP overlaps with the MIEL1-RFP on the plasma membrane but not on the nucleus (Fig. 3a). This results indicate that HbRbohD is localized to the plasma membrane.
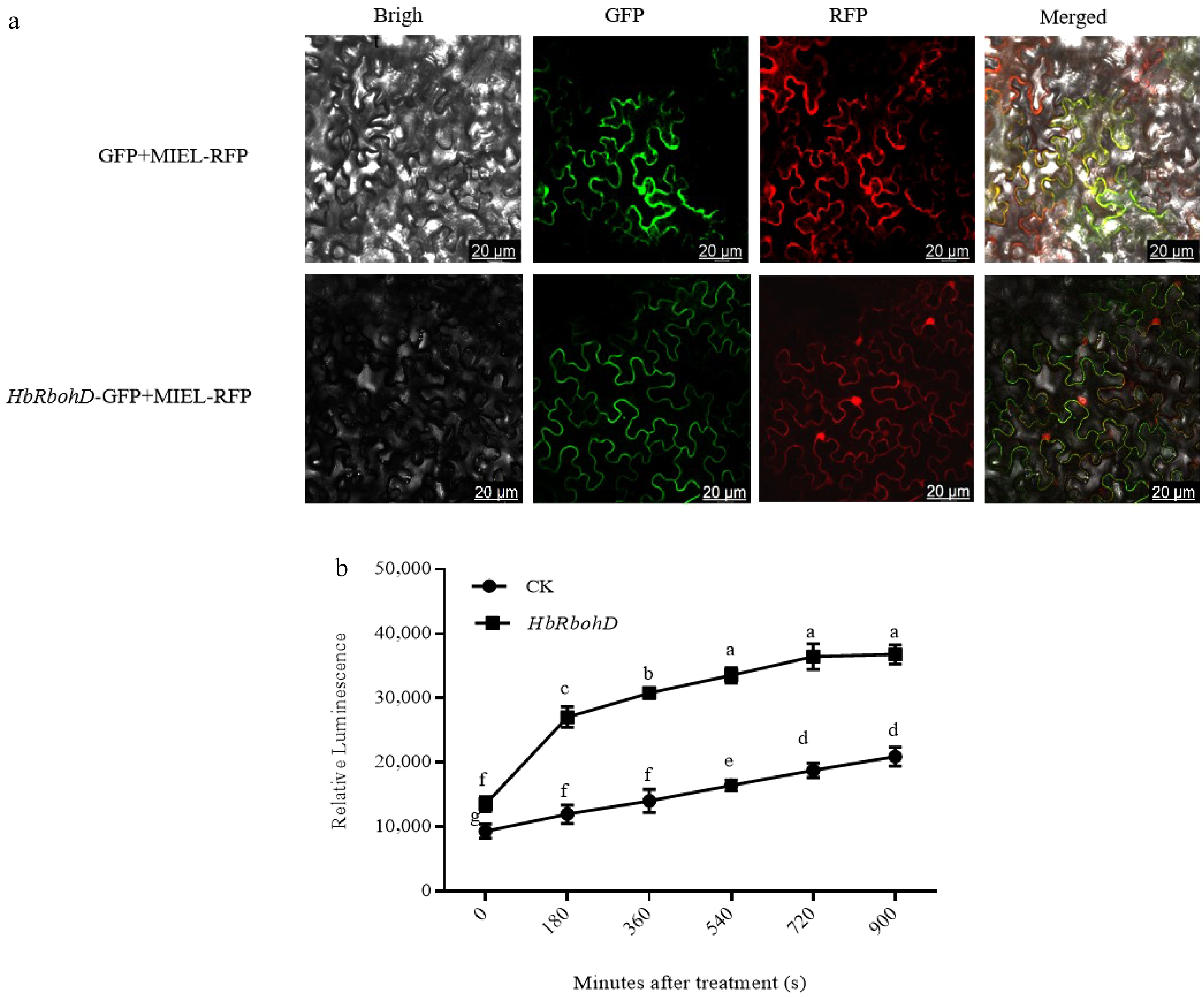
Figure 3.
Subcellular localization of HbRbohD and the function of HbRbohD on ROS production. (a) Subcellular localization of HbRbohD. Tobacco leaves co-expressing GFP/MIEL-RFP and HbRbohD-GFP/MIEL-RFP were viewed with fluorescent microscope 24 h after infiltration (bar, 20 μm). (b) The function of HbRbohD on ROS production. The oxidative degree in protoplasts was detected by dichlorofluorescin diacetate (DCFH-DA) and the fluorescence intensity was measured every 3 min for 15 min. Data presented are the mean ± SD from three independent experiments and the mean values assigned dissimilar superscript letters differ significantly (p < 0.05).
RBOHs have been identified as the predominant catalytic agents generating reactive oxygen species in plants[9]. To explore the role of HbRbohD in ROS generation, ROS levels were measured using dichlorofluorescein diacetate (DCFH-DA) in rubber tree mesophyll protoplasts overexpressing HbRbohD (Fig. 3b). Data indicated a marked rise in ROS accumulation in protoplasts overexpressing HbRbohD compared to the control group, highlighting the pivotal role of HbRbohD in driving ROS production.
Overexpression of HbRbohD enhanced plant resistance to fungal pathogens, and seed tolerance to osmotic stresses
-
To investigate the role of HbRbohD in plant disease resistance, HbRbohD transgenic Arabidopsis plants were generated under the control of the 35S promoter in the Col-0 background, designated as 35S::HbRbohD. The transgenic lines were confirmed through genomic PCR and RT-PCR analyses (Supplementary Fig. S1). Subsequently, the phenotypic response of 35S::HbRbohD plants to the necrotrophic fungus Botrytis cinerea and A. brassicicola were assessed. Upon inoculation with B. cinerea and A. brassicicola, necrotic lesions were observed on both Col-0 and 35S::HbRbohD seedlings at 3 d post-inoculation (Fig. 4a). The necrotic lesion size on Col-0 plants was significantly smaller compared to that on 35S::HbRbohD plants (Fig. 4b). These results suggested that HbRbohD enhances Arabidopsis resistance to B. cinerea and A. brassicicola, highlighting its positive role in plant disease resistance.
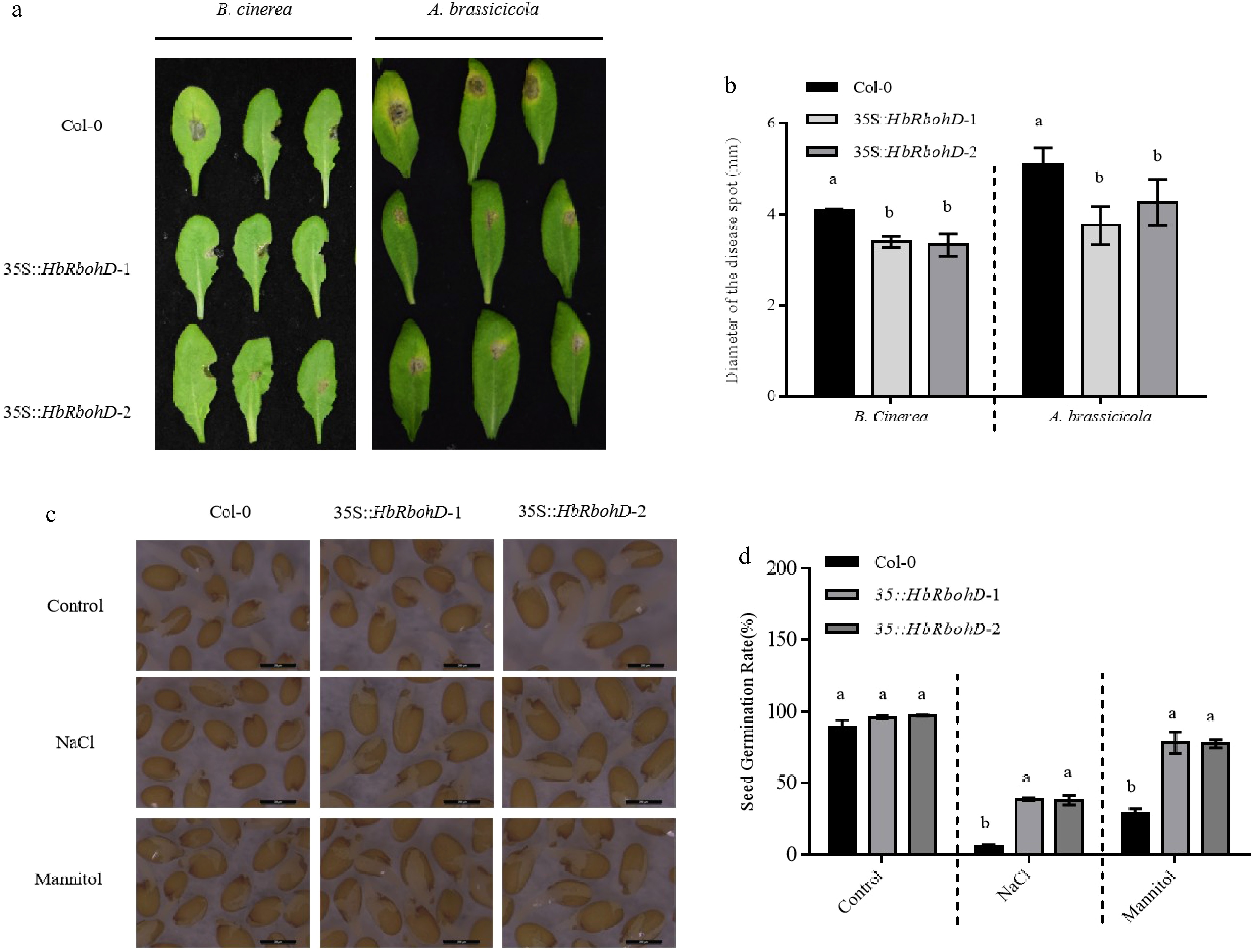
Figure 4.
Disease and seed osmotic tolerance assay of Col-0 and 35S::HbRbohD. (a) Disease symptoms of A. thaliana Col-0 and 35S::HbRbohD to B. cinerea and A. brassicicola. (b) Statistical analysis of necrotic lesion in WT and 35S::HbRbohD. The symptoms observation and the lesion measure were performed at 3 d post inoculation with B. cinerea and A. brassicicola, respectively. (c) Seed germination of Col-0 and 35S::HbRbohD on ½ MS medium with exogenous NaCl treatment (150 mM) and mannitol treatment (200 mM). (d) The quantitative analysis of germination rates of Col-0 and 35S::HbRbohD on ½ MS medium with exogenous NaCl treatment (150 mM), and mannitol treatment (200 mM). Data presented was the mean ± SD from three independent experiments and the mean values assigned dissimilar superscript letters differ significantly (p < 0.05).
Previous studies have demonstrated that AtRbohD plays a crucial role in plant resistance to salt stress[34]. To investigate the function of HbRbohD under osmotic stress conditions, the seed germination of both wild-type (WT) and 35S::HbRbohD transgenic lines under NaCl and mannitol treatments were examined (Fig. 4c). Comparative analysis revealed that the seed germination rates of 35S::HbRbohD transgenic plants were significantly higher than those of WT plants (Fig. 4d). These findings suggest that HbRbohD positively regulates osmotic stress tolerance, particularly in response to salinity.
Overexpression of HbRbohD changed the transcripts of defense-related genes
-
Given that the PTI inducers (Chitin/flg22) and salicylic acid (SA) treatments could significantly induce the expression of HbRbohD (Fig. 3), the expression profiles of defense-related genes, including PTI marker genes such as AtFRK1 and AtCYP71, and key components of the SA signaling pathway such as AtPAL, AtSID2, AtPR1, and AtNPR1, were analyzed by qRT-PCR in Col-0 and 35S::HbRbohD inoculated with B. cinerea. As shown in Fig. 5, without inoculation with B. cinerae (0 d post inoculation), the expression levels of AtFRK1, AtCYP71, AtPAL, AtSID2, AtPR1, and AtNPR1 in 35S::HbRbohD lines were increased significantly compared to that in Col-0. At 1 and 2 d post inoculation with B. cinerae, although there were differences in the expression levels of these genes, their expression trends in the Col-0 and 35S::HbRbohD lines were similar, that is, the expression level in 35S::HbRbohD was significantly higher than that in Col-0 (Fig. 5). These results demonstrated that overexpression of HbRbohD enhanced the PTI and SA signaling pathway in plants. Taking into account the results shown in Fig. 4a, it suggested that HbRobhD enhanced plant resistance to fungal pathogens through SA-mediated defense response.
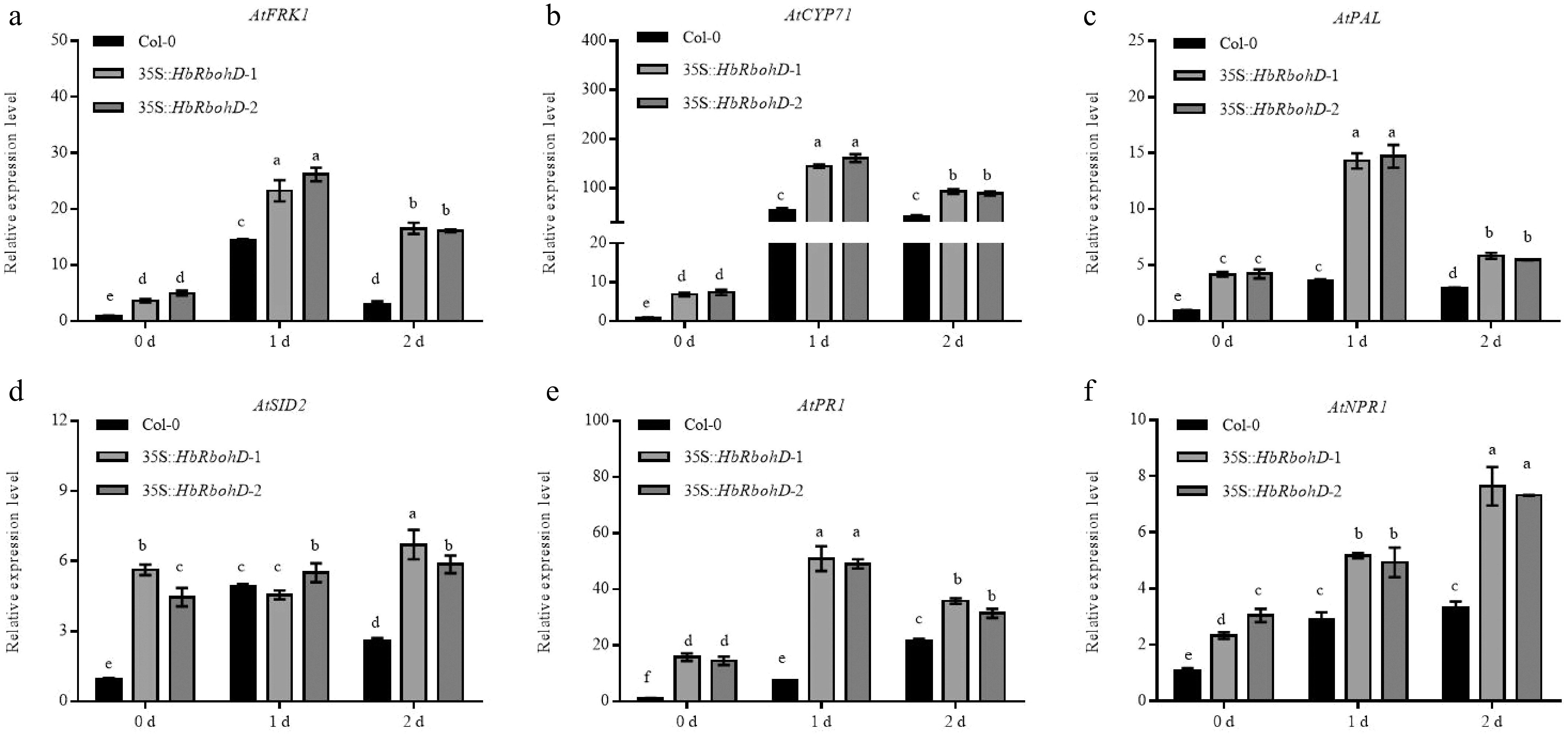
Figure 5.
Relative expression assay of defense related genes in Col-0 and 35S::HbRbohD inoculated with B. cinerea. The expression of defense related genes, such as (a) AtFRK1, (b) AtCYP71, (c) AtPAL, (d) AtSID2, (e) AtPR1, and (f) AtNPR1, were analyzed by qRT-PCR in Col-0 and 35S::HbRbohD inoculated with B. cinerea. Data are shown as the means ± SD from three independent experiments. Different letters above columns indicate a significant difference (p < 0.05).
HbRbohD enhanced plant antioxidant enzyme activities
-
To elucidate the potential mechanism by which HbRbohD confers plant resistance, the activities of key antioxidant enzymes (SOD, POD, CAT), and malondialdehyde (MDA) content in both Col-0 and 35S::HbRbohD transgenic lines were measured. The results showed that 35S::HbRbohD exhibited significantly enhanced activities of POD and CAT, but not SOD, compared to those of Col-0 control plants (Fig. 6a–c). Conversely, MDA content in 35S::HbRbohD plants was markedly lower than that in Col-0 plants (Fig. 6d). These findings indicated that HbRbohD contributes to plant resistance by regulating antioxidant enzyme activities and reducing oxidative damage.
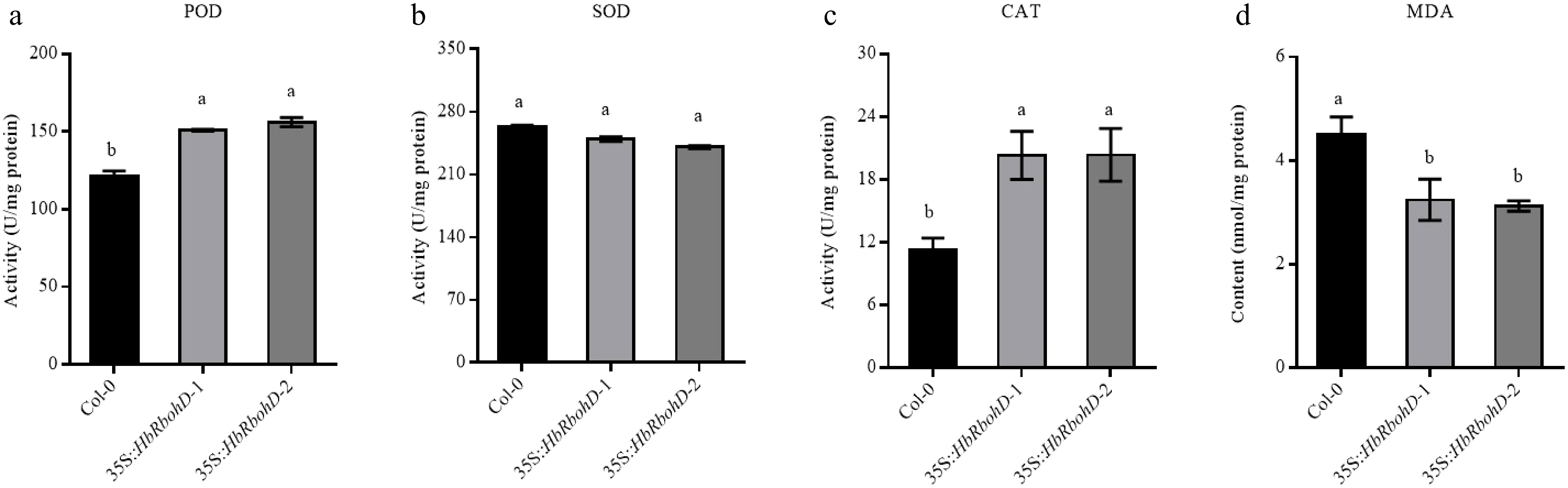
Figure 6.
Activity of antioxidant enzymes and MDA content in Col-0 and 35S::HbRbohD. The key antioxidant enzyme activities of (a) POD, (b) SOD, (c) CAT, and the content of (d) MDA were analyzed in Col-0 and 35S::HbRbohD, respectively. Data presented was the mean ± SD from three independent experiments and the mean values assigned dissimilar superscript letters differ significantly (p < 0.05).
-
The Rboh protein family in plants functions as a central regulator of reactive oxygen species biosynthesis and participates in modulating plant morphogenesis, and development, and responds to abiotic/biotic stresses[15]. Structural analyses have revealed that all identified plant Rboh proteins share conserved domains, including six transmembrane helices, a cytosolic C-terminal region containing NADPH- and FAD-binding sites, and an N-terminal region featuring phosphorylation sites and EF-hand motifs for calcium binding[12]. The present sequence analysis of HbRbohD from rubber tree demonstrated similar structural features, including six transmembrane helices, NADPH_Ox and NAD-binding domains, a FAD-binding domain, and three EF-hand motifs (Supplementary Fig. S1). Notably, a difference in EF-hand motif number was observed—three in AtRbohD vs two in HbRbohD. Since EF-hand motifs are critical for calcium binding and directly regulate RbohD activity, this structural variation suggests that although HbRbohD likely functions similarly to AtRbohD, their calcium responsiveness may differ significantly.
ROS serve as crucial signaling molecules in plant responses to diverse abiotic and biotic stresses, and Rboh proteins directly catalyze ROS production in plants[16,28]. The present study demonstrates that HbRbohD expression in rubber tree mesophyll protoplasts significantly induced ROS generation (Fig. 3b), confirming the function of HbRbohD in ROS production. Additionally, ROS signaling exhibits remarkable spatiotemporal specificity, achieved through precise regulation of ROS production and scavenging across various organelles and subcellular compartments, including chloroplasts, mitochondria, peroxisomes, and apoplast[52]. The subcellular localization analysis revealed that HbRbohD is predominantly localized to the plasma membrane (Fig. 4), suggesting its involvement in apoplastic ROS production. This finding is particularly significant as an apoplastic ROS burst represents one of the earliest detectable events in plant stress responses, typically triggered by extracellular stimuli[53]. In Arabidopsis, the AtRbohD-mediated reactive oxygen species burst represents a conserved stress response mechanism that is activated by diverse biotic and abiotic challenges, particularly pathogen infection and salt stress[17]. The present results reveal that HbRbohD expression was significantly upregulated in rubber trees following inoculation with fungal pathogens (C. gloeosporioides and O. heveae Steinm) and exposure to abiotic stresses (salt and drought treatments) (Fig. 2). These findings strongly indicate that HbRbohD-mediated ROS production plays a pivotal role in rubber trees' defense responses against pathogen attack, salinity, and drought conditions.
ROS signaling is connected with the response to phytohormones, including salicylic acid (SA), jasmonic acid (JA), ethylene (ET), abscisic acid (ABA), gibberillin (GA), and auxin[54−56]. Phytohormones have also been proven to be extensively involved in plant biotic and abiotic stresses[47−51]. The analysis of cis-elements revealed the presence of various stress and phytohormone response elements in the promoter regions of HbRbohD gene, such as MYB and MYC which were reported to participate in drought, low temperature, and salt stress responses[57], low temperature response element (LTR), drought response element (MBS), defense and stress responsiveness (TC-rich repeats), gibberellin response element (GARE), abscisic acid response element (ABRE), salicylic acid response element (SARE), and jasmonic acid response element (JARE) (Supplementary Table S1). The expression patterns of HbRbohD under pathogen inoculation and phytohormone treatment by qRT-PCR showed that exogenous SA, JA, ET, and GA could significantly promote the transcriptional level of HbRbohD, but not ABA (Fig. 2f–j), which basically supports the results of cis-element analysis. Although no ethylene response element was detected in the promoter of the HbRbohD gene, HbRbohD still responded to ET, probably because HbRbohD responded to ET indirectly rather than directly.
Extensive research has revealed the critical role of AtRbohD in plant immune responses as well as the tolerance to salinity and drought[17]. In the present study, the expression of HbRbohD is significantly up-regulated by chitin and flg22 (Fig. 2c, d). Overexpression of HbRbohD induces ROS accumulation in rubber tree mesophyll protoplasts (Fig. 3b), and increases the resistance of Arabidopsis to the pathogens (Fig. 4a). On the other hand, the germination rates of AtRbohD-overexpression seeds are significantly lower than those of the wild type. However, the present results showed that the germination rates of 35S::HbRbohD seeds are significantly higher than that of wild type (Fig. 4b), suggesting the distinct regulation pathway of salt tolerance between AtRbohD and HbRbohD. A few other studies have shown that ABA and GA signaling pathways salinity are involved in salinity tolerance during seed germination[32,58,59]. The present study exhibits the expression of HbRbohD significantly up-regulated quickly at 12 h after GA treatment, but not ABA (Fig. 2i & j). Thus, HbRbohD might be involved in responses to plant salt stress through the GA signaling pathway.
In plants, ROS scavenging enzymes, such as superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), and ascorbate peroxidase (APX), play crucial roles in maintaining ROS homeostasis and mitigating oxidative damage[13]. The present study revealed that 35S::HbRbohD transgenic lines exhibited enhanced activities of POD, and CAT, but not SOD (Fig. 6a–c), along with reduced malondialdehyde (MDA) content (Fig. 6d). These results suggest that the improved resistance of 35S::HbRbohD to biotic and abiotic stresses may be attributed to the elevated activity of ROS-scavenging enzymes, which effectively maintain cellular redox balance in plants.
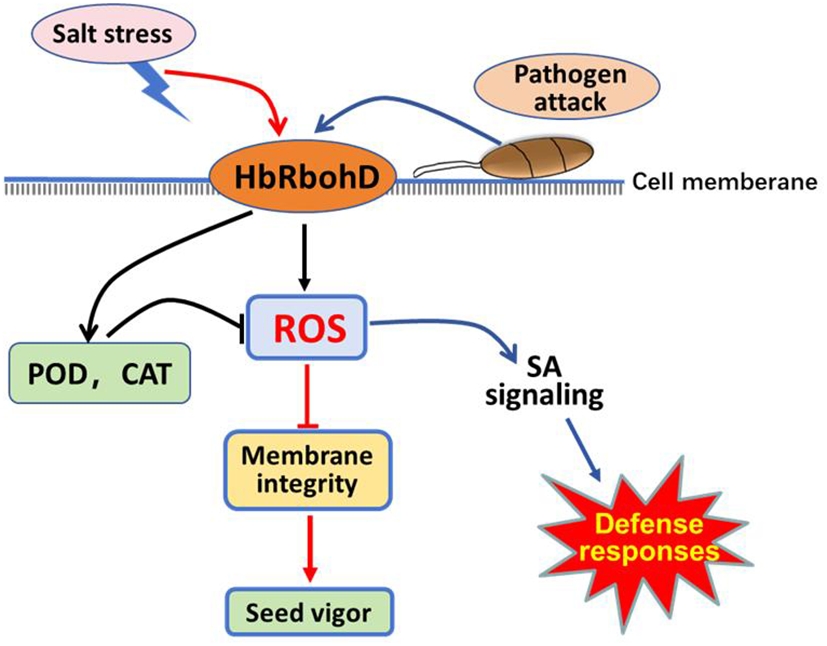
Taken together, this study identified a homolog of the respiratory burst oxidases homolog D gene of rubber tree HbRbohD, and performed analysis of its promotor and expression profiles under biotic and abiotic stress conditions. Further analysis demonstrated that HbRbohD was located on the cell membrane and was involved in the production of ROS. Overexpression of HbRbohD in Arabidopsis plants enhanced resistance to pathogenic fungi such as B. cinerea and A. brassicicola, and promoted tolerance to salt and mannitol with increased expression of multiple defense response-related genes and activities of antioxidant enzymes such as SOD and CAT. These findings suggest that salt stress and pathogen attack promoted HbRbohD expression and antioxidant enzyme activities to maintain ROS homeostasis in plants, which regulate cell membrane integrity, seed germination ability, and the expression of defense-related genes, especially SA signaling genes, enabling plants to respond to biotic and abiotic stresses (Fig. 7). This study offered new insights into the contribution of HbRbohD to plant biotic and abiotic resistance through modulating ROS homeostasis.
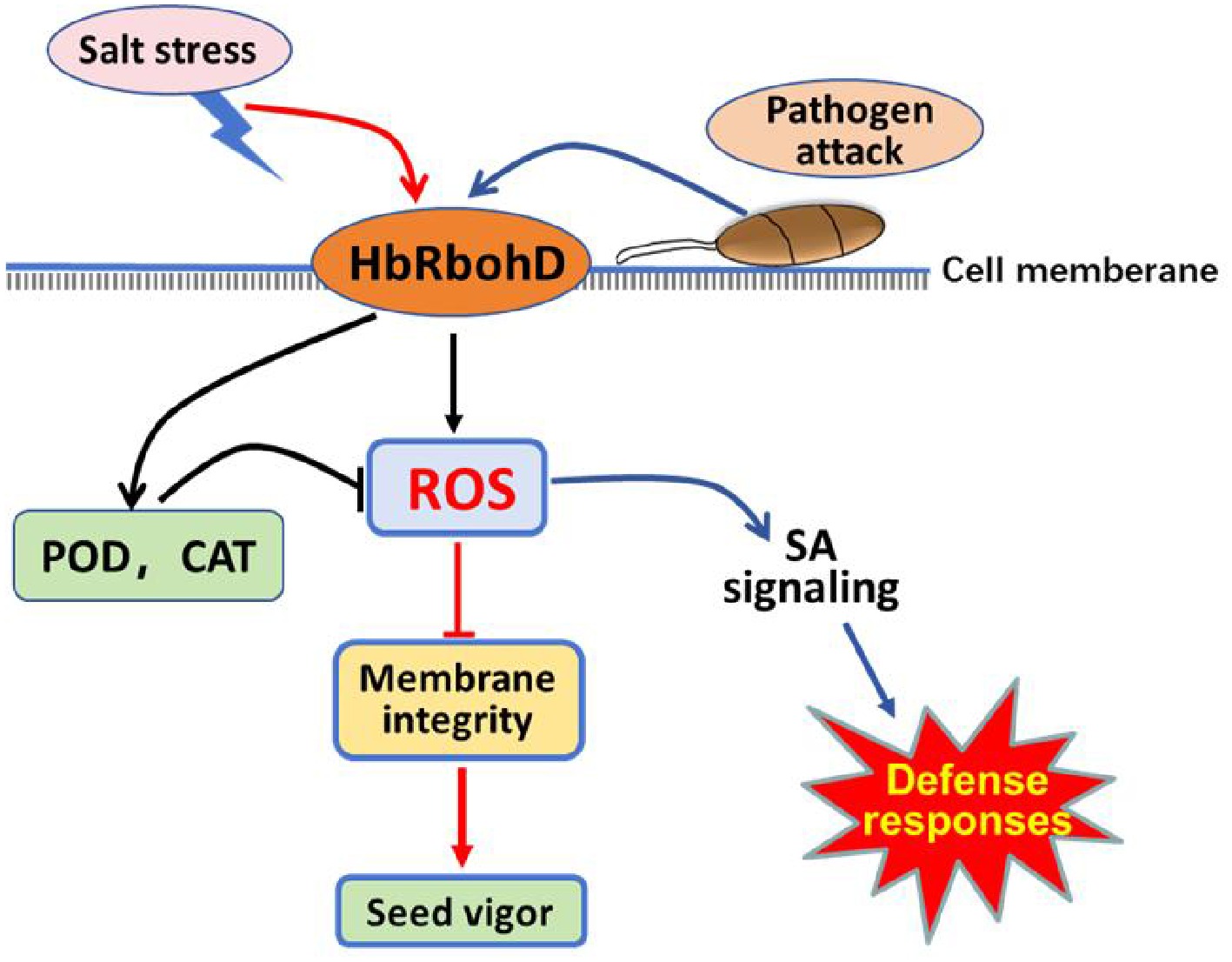
Figure 7.
Proposed model of respiratory burst oxidase homolog D of rubber tree (HbRbohD) signaling in response to biotic and abiotic stressed. HbRbohD regulates ROS homeostasis through antioxidant enzymes to regulate cell membrane integrity and seed vigor under salinity stress conditions (red arrows). HbRbohD also regulates the expression of defense-related genes,especial, enabling plants to respond to pathogen attack (blue arrows). The black arrows represent the common pathway shared by salt stress and pathogen attack.
This work was supported by the Hainan Provincial Natural Science Foundation of China (352RC649), and the National Natural Science Foundation of China (32260716).
-
The authors confirm contributions to the paper as follows: study conception and design: Luo H, An B; data collection: Wang M, Song Y; analysis and interpretation of results: Wang M, Song Y; draft manuscript preparation: Wang Q, Chen J, He C. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0025-0029)
-
Received 23 April 2025; Accepted 28 August 2025; Published online 13 November 2025
-
A respiratory burst oxidases homolog D (RbohD) gene in rubber tree (Hevea brasiliensis), which responds to phytohormone treatments and biotic/abiotic stresses, was identified.
HbRbohD induced ROS accumulation in rubber tree mesophyll protoplasts.
HbRbohD overexpression in Arabidopsis exhibited increased antioxidant enzyme activity, enhanced plant resistance to Botrytis cinerea and Alternaria brassicicola, increased the expression level of defense genes, and promoted seed tolerance to salt and mannitol.
- Supplementary Table S1 Analysis of cis-acting elements in the 3,000 bp upstream fragment of the HbRbohD.
- Supplementary Fig. S1 Molecular characterization of HbRbohD-Overexpressing Arabidopsis lines.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang M, Song Y, An B, Wang Q, Chen J, et al. 2025. Rubber tree respiratory burst oxidases homolog D contributed to plant biotic and abiotic resistance. Tropical Plants 4: e037 doi: 10.48130/tp-0025-0029 

Rubber tree respiratory burst oxidases homolog D contributed to plant biotic and abiotic resistance
- Received: 23 April 2025
- Revised: 06 August 2025
- Accepted: 28 August 2025
- Published online: 13 November 2025
Abstract: Reactive oxygen species (ROS) serve as signaling molecules in the responses to biotic and abiotic stresses in plants. The respiratory burst oxidase homologue (Rboh) proteins play a crucial role in ROS production. Among them, RbohD-dependent ROS production showed a high degree of biotic and abiotic stress responsiveness. In this study, the homologous gene of rubber tree (Hevea brasiliensis) respiratory burst oxidase homolog D was amplified, and named as HbRbohD. The full-length of the HbRbohD gene contained 2,730 bp, encoding a 910 amino acid polypeptide with conserved NADPH_Ox, EF-hand, and a NAD-binding domain. Putative cis-elements assay showed that the promoter of HbRbohD contained SARE, JARE, GARE, ABRE, TC-rich repeats, LTR, and MBS that could be triggered by diverse biotic and abiotic stimuli. RT-qPCR data revealed that the expression of HbRbohD was induced by multiple phytohormones and pathogenic fungi such as Oidium heveae Steinm and Colletotrichum gloeosporioides. HbRbohD was predominantly localized in the plasma membrane and exhibited significant modulation of reactive oxygen species (ROS) in plant cells. Arabidopsis plants overexpressing HbRbohD had enhanced resistance to Botrytis cinerea and Alternaria brassicicola, and promoted tolerance to salt and mannitol with increased activities of dismutase (SOD), catalase (CAT), and decreased MDH content. The present results suggest that HbRbohD contributes to plant biotic and abiotic stresses by modulating the defense response, and the activity of antioxidant enzymes. This study provides new insights into the response of rubber trees to biotic and abiotic stresses through modulating HbRbohD-mediated ROS homeostasis.
-
Key words:
- Hevea brasiliensis /
- HbRbohD /
- Disease resistance /
- Defense enzymes