






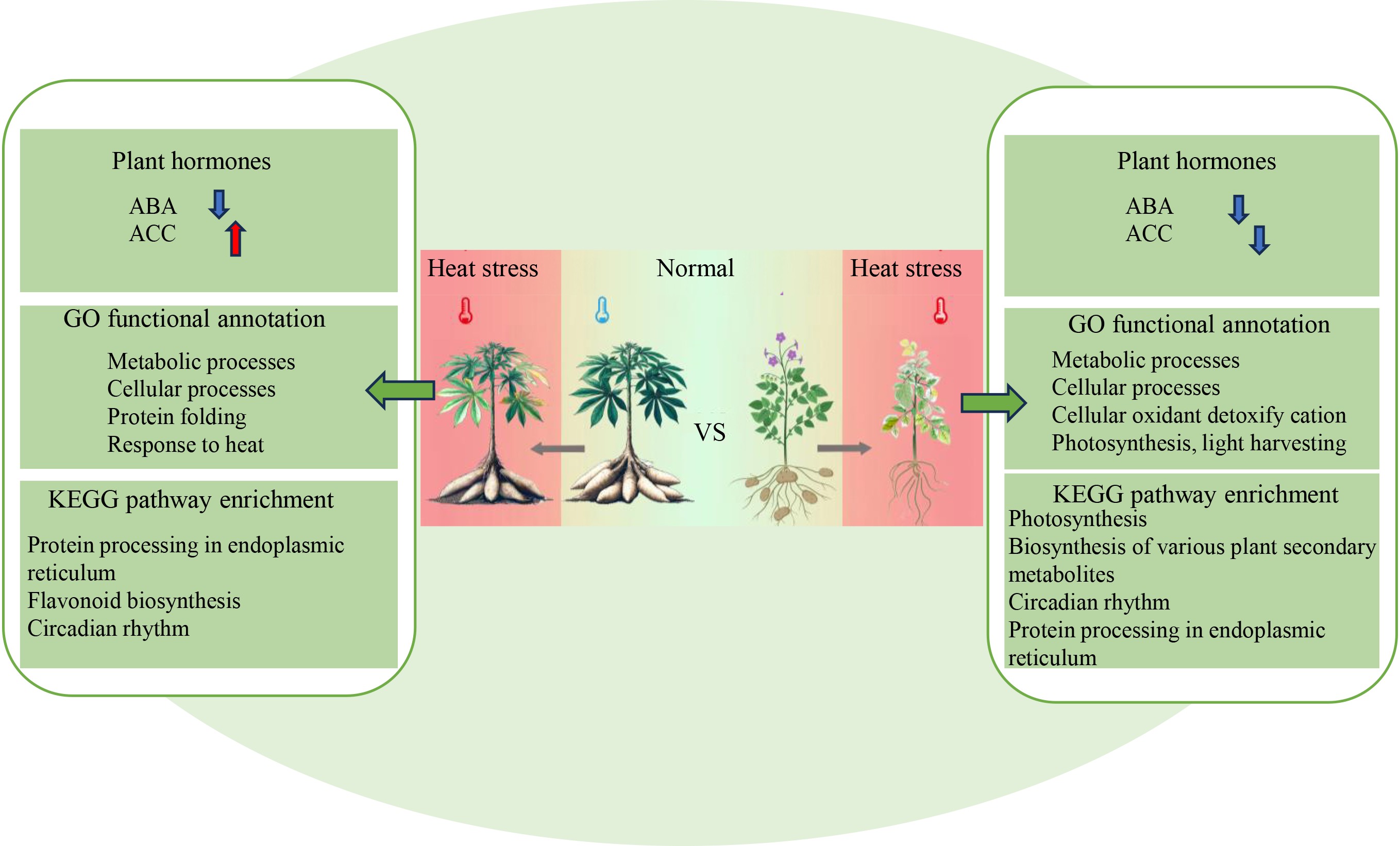



-
Global warming poses a significant threat to agricultural productivity, making the improvement of crop heat tolerance an urgent priority. Cassava (Manihot esculenta Crantz), a perennial shrub of the Euphorbiaceae family, thrives at temperatures between 27–35 °C and is considered one of the world's three major root crops alongside potato and sweet potato[1]. Due to its high starch content, cassava is often called the 'king of starch'[2]. It is mainly cultivated in hot regions such as Guangdong, Guangxi, and Hainan in China[3]. Although cassava exhibits considerable tolerance to high temperatures, the molecular basis governing its heat response remains largely unclear. Previous studies have indicated that pyruvate metabolism and phenylalanine pathways are involved in the heat response of cassava leaves and veins, respectively[4], while other studies have linked ROS regulation through PP2C1-mediated dephosphorylation of catalase and ascorbate peroxidase to heat tolerance[5]. However, detailed transcriptomic and hormonal responses of cassava to high temperature are still lacking.
Potato (Solanum tuberosum L.), in contrast, is a temperate crop with an optimal growth temperature of 18–22 °C. Temperatures above 25 °C inhibit tuberization and significantly reduce yield[6]. At 39 °C, severe growth inhibition occurs, including halted stem elongation and leaf damage[7]. Heat-tolerant potato varieties exhibit relatively stable tuberization and photosynthetic efficiency under heat stress[8].
Plant hormones, as endogenous metabolites of plants themselves, are an important signaling factor throughout the entire growth and development process of plants, playing a significant role in the growth and development cycle of plants. Research shows that when Arabidopsis faces high-temperature stress, the hypocotyl and petiole show substantial growth with a high content of IAA. This proves IAA plays a role in thermomorphogenesis when Arabidopsis is under high temperature[9]. Also, the PIF4, the TF controlling circadian rhythm, can lead to hypocotyl growth. Some researchers proved that AtPIF4 can modulate the auxin biosynthesis pathway and change the level of IAA by inducing mutants in Arabidopsis, such as yuc8, taa1, and cyp7982[10]. In some ways, giving plants extra hormones can also enhance their heat tolerance. A study shows that giving potatoes extra SA can help potatoes raise their heat tolerance[11]. Nowadays, more researchers think that ABA and SA are the signal molecules that induce heat tolerance in plants through heat acclimation[12,13].
Heat Shock Transcription Factors (HSF) plays an important role in plant growth and development, particularly under abiotic stress conditions. They help plants cope with extreme environmental stresses such as high temperature, salinity, drought, and intense light by regulating stress-responsive genes[14]. Among them, HSF1 has been identified as a key transcription factor (TF) specifically involved in the heat shock response (HSR), functioning differently from other TFs such as C-MYC[15]. In Arabidopsis, overexpression of ZmHSF1 has been shown to significantly increase the production of heat shock proteins (HSPs) compared to wild-type plants, indicating that ZmHSF1 plays a central role in mediating HSR[16]. Multiple studies have demonstrated that HSFs are essential for plant stress responses, primarily through the regulation of HSP expression under abiotic stress conditions[17]. Thus, the level and composition of HSP expression are widely used as indicators for selecting plant genotypes with high heat tolerance[18−20].
Weighted gene co-expression network analysis (WGCNA) is increasingly being used to dissect how tropical crops cope with environmental stresses, uncovering the key regulatory circuits that operate under adverse conditions. In cassava, for example, WGCNA of high-light-stressed transcriptomes has been combined with targeted studies of the NADP-ME gene family to identify candidate regulators that underpin photoprotective responses, providing prime targets for improving high-light tolerance[21]. In banana, a co-expression network constructed from cold-stressed peel samples revealed the transcription factor MabHLH355 and other hubs tightly linked to reactive oxygen species (ROS) scavenging, outlining the molecular framework that governs chilling-injury avoidance in this climacteric fruit[22]. Collectively, these studies demonstrate that WGCNA not only accelerates the discovery of stress-resilience genes and networks in tropical crops but also furnishes molecular markers and breeding targets for enhancing crop performance under climate-related stresses.
This study aimed to elucidate the differences between cassava and potato in their responses to high-temperature stress by integrating hormonal profiling and RNA-seq data. Using WGCNA, key gene modules and constructed regulatory networks highlighting critical differences between these two crops were identified. These findings provide molecular insights into why cassava is more heat-tolerant than potato and offer valuable resources for breeding heat-tolerant crop varieties.
-
The stems of cassava (SC205 and KU50) were grown in pots, under greenhouse conditions (14/10 h of light/dark, 28/25 °C day/night) until 45 d. Specifically, SC205 is a heat-tolerant local variety commonly grown in tropical regions of China, characterized by robust growth and stable yield under high-temperature conditions. KU50 is an internationally cultivated high-yield variety with moderate heat tolerance and good starch quality, widely used in comparative physiological and molecular studies. For potato, 'Qingshu No. 9' and 'FAVORITA' were used; these are representative temperate cultivars with contrasting heat sensitivities — 'Qingshu No. 9' being relatively heat-sensitive, while 'FAVORITA' exhibits better heat tolerance. The tubers of potato (Qingshu No. 9 and FAVORITA, qing9 and F) were grown in pots, under greenhouse conditions (14/10 h of light/dark, 25/23 °C day/night) until 40 d, at the Institute of Tropical Bioscience and Biotechnology (Haikou, China).
HT stress treatment of plants
-
Cassava and potato plants of the same height were selected as the control (CK) and treatment (PG). The heat treatment of cassava was in 45 °C greenhouse conditions (14/10 h of light/dark, 45 °C day/night) for 3 d. The heat treatment of potato was in 40 °C greenhouse conditions (14/10 h of light/dark, 40 °C day/night) for 3 d. During the HT stress time, the CK and the PG both had the same amount of water.
Detection and analysis of plant hormones
-
Leaves were sampled at 0, 6, 48, and 72 h after heat treatment, with two biological replicates for each time point. The samples were immediately frozen in liquid nitrogen and ground in a chilled mortar. For extraction, 100 mg of ground tissue was mixed with 1 mL of cold 50% acetonitrile (ACN) solution. The mixture was sonicated for 3 min and extracted for 30 min at 4 °C, followed by centrifugation at 12,000 rpm for 10 min at 4 °C. The supernatant was passed through an RP-SPE column, washed with 1 mL of 100% methanol and 1 mL of deionized water, and eluted with 1 mL of 30% ACN. The eluate was evaporated to dryness under nitrogen and reconstituted in 200 μL of 30% ACN for analysis.
Total RNA extraction, cDNA library construction, and high-throughput sequencing
-
Leaves were collected at 0, 2, 6, 10, 24, and 48 h after heat treatment for RNA sequencing, with three biological replicates per time point. Total RNA was extracted using a polysaccharide and polyphenol plant RNA extraction kit (Tiangen Biotech, Code No. DP441). RNA integrity and contamination were assessed using 1% agarose gel electrophoresis. mRNA was enriched using Oligo(dT) magnetic beads, and cDNA libraries were constructed with the NEBNext® Ultra™ RNA Library Prep Kit for Illumina®. Library concentrations were preliminarily measured using a Qubit 2.0 Fluorometer, and library quality was evaluated using an Agilent 2100 Bioanalyzer (Agilent Technologies, CA, USA). Qualified libraries were sequenced on an Illumina platform.
Bioinformatics analysis of Illumina data
-
The transcriptome sequencing data were filtered using the SOAPnuke software (v1.5.6)[23] developed by BGI. Reads containing adapters, reads with more than 5% unknown bases (N), and reads where more than 20% of bases had a quality score below 15 were removed. The filtered, clean reads were saved in FASTQ format. The clean reads were then aligned to the reference genomes using HISAT2 (v2.1.0)[24]. The cassava reference genome used was GCF_001659605.2_M.esculenta_v8 from the NCBI cassava database, and the potato reference genome was GCF_000226075.1_SolTub_3.0 from the NCBI potato database.
DEGs analysis
-
High-throughput sequencing was used to obtain read count data for gene expression levels across samples. Differential expression analysis was performed using DESeq2 (v1.4.5) in R (version 4.2.1), with a threshold of |log2FC| ≥ 1 and Q value ≤ 0.05 to identify differentially expressed genes (DEGs) compared to the control. The DEGs were then subjected to KEGG (
www.kegg.jp ) and GO (www.geneontology.org ) enrichment analyses to identify the main gene changes enriched under heat stress in cassava and potato.Weighted gene co-expression network analysis (WGCNA) and function annotation
-
The differentially expressed genes (DEGs) in cassava and potato were selected. Using R (version 4.2.1), a WGCNA was constructed to divide these DEGs into modules based on their expression patterns. The power parameters, minimum module size, and thresholds for merging similar modules were set according to the specific conditions. The similarities and differences in module clustering between cassava and potato DEGs were compared. Then, interaction networks were built for the differentially expressed genes in the specific modules, and finally, candidate heat-tolerant genes in cassava were identified.
Real-time quantitative PCR analysis
-
To verify the accuracy of the Illumina sequencing data, reverse transcription was performed, and the expression of selected differentially expressed genes was analyzed using a real-time quantitative PCR (RT-qPCR) kit from Accurate Biotechnology (Hunan) Co., Ltd. (Product No: AG11728) and an RT-qPCR detection kit (Product No: AG11701). The RT-qPCR was performed on a Bio-Rad CFX96 instrument. The reaction mixture consisted of 1 μL cDNA template, 0.4 μL of each primer, 10 μL PCR Mix, and 8.2 μL RNase-free water, with a total volume of 20 μL. The RT-qPCR conditions were as follows: initial denaturation at 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s, and 60 °C for 30 s. Primer information is provided in Supplementary Table S1. The cassava TUBLIN gene and the potato ACTIN gene were used as internal reference genes[25]. All RT-qPCR reactions were run with four technical replicates per sample. Statistical evaluation was performed in GraphPad Prism 8.0 (GraphPad Software, San Diego, CA, USA) using Tukey's HSD test; means bearing different letters differ significantly at p < 0.05. Illustrations were prepared with the same software.
-
To evaluate the differences in heat tolerance between cassava and potato, both tissue-cultured and potted seedlings were compared under HT stress. The results indicated that the cassava cultivar KU50 exhibited greater heat tolerance than SC205, while the potato cultivar FAVORITA (F) showed higher tolerance than Qingshu No. 9 (qing9).
Thermal imaging revealed that cassava leaves in the HT treatment group (PG) exhibited higher surface temperatures than those in the control group (CK), indicating the impact of heat stress (Fig. 1a). After 3 d of HT treatment, both tissue-cultured and potted cassava seedlings displayed visible symptoms such as yellowing leaves, whereas CK plants remained green (Fig. 1c, e).
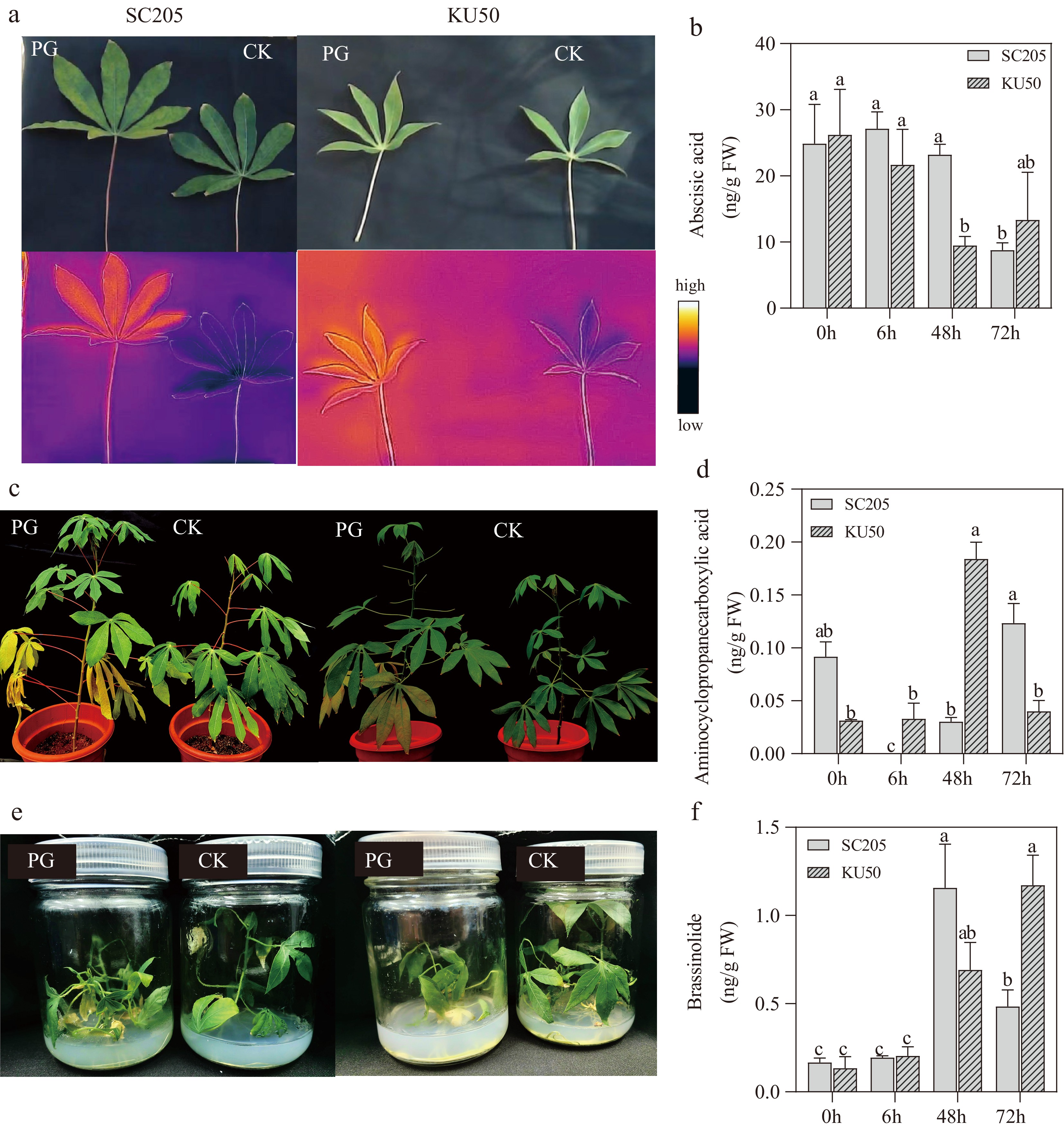
Figure 1.
Phenotypic changes in cassava under high-temperature treatment. (a) Infrared leaf temperature comparison between SC205 and KU50 under control (CK) and heat treatments (PG). (b) ABA content changes in SC205 and KU50 under high-temperature stress. (c) Phenotypic alterations of potted seedlings (control vs 72 h heat treatment) in SC205 and KU50. (d) ACC content changes in SC205 and KU50 under high-temperature stress. (e) Phenotypic alterations of tissue-cultured seedlings (control vs 72 h heat treatment) in SC205 and KU50. (f) BR content changes in SC205 and KU50 under high-temperature stress. Different lowercase letters indicate significant differences at p ≤ 0.05.
In potato, HT stress caused more severe symptoms, particularly stem wilting (Fig. 2c). Hormone profiling further revealed species-specific responses. In cassava, KU50 showed a 4.5-fold increase in ACC content, along with decreases of 63% in ABA and 36% in BR compared with CK; SC205 showed a similar pattern, with a 1.9-fold increase in ACC, and decreases of 65% in ABA and 35% in BR (Fig. 1b–f). In contrast, potato cultivars exhibited a decline in ACC and ABA under HT conditions: ACC decreased by 81% in F and 63% in qing9, while ABA decreased by 55% and 42%, respectively. Notably, BR was undetectable in all potato samples (Fig. 2b, d), suggesting a potential role of BR signaling in the greater heat resilience observed in cassava.
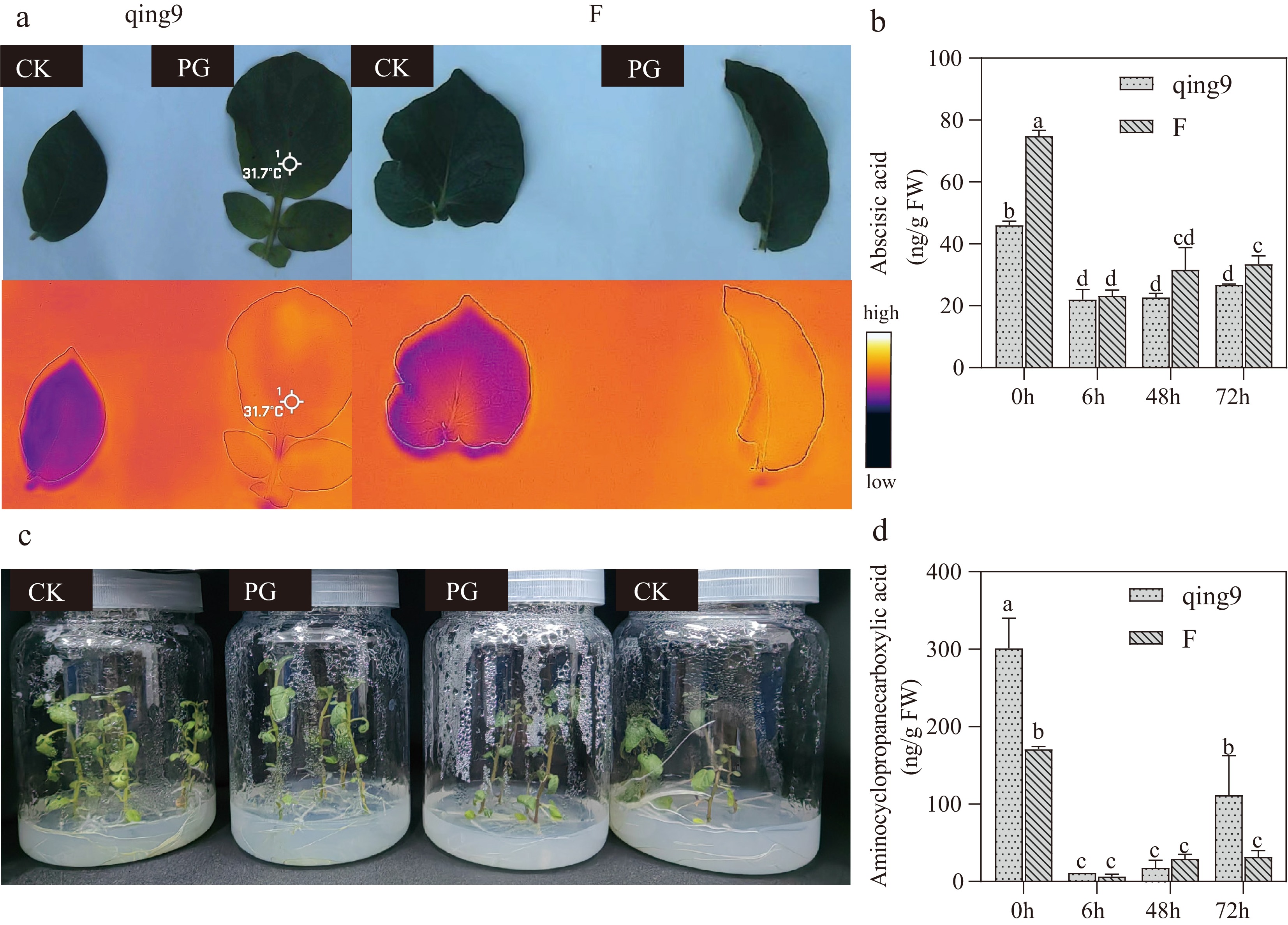
Figure 2.
Phenotypic changes in potato under high-temperature treatment. (a) Infrared leaf temperature comparison between Qingshu No. 9 and FAVORITA under control (CK) and heat treatments (PG). (b) ABA content changes in Qingshu No. 9 and FAVORITA under high-temperature stress. (c) Phenotypic alterations of tissue-cultured seedlings (control vs 72 h heat treatment) in Qingshu No. 9 and FAVORITA. (d) ACC content changes in Qingshu No. 9 and FAVORITA under high-temperature stress. Different lowercase letters indicate significant differences at p ≤ 0.05.
Phenotypic and transcriptomic responses of cassava and potato to HT stress
-
Under high-temperature (HT) stress, cassava and potato exhibited strikingly different phenotypic and molecular responses. Cassava maintained relatively stable growth with only slight leaf yellowing, whereas potato suffered severe dehydration and wilting, reflecting its lower heat tolerance. Hormonal heatmaps further indicated that cassava displayed moderate changes in hormone levels, while potato showed dramatic fluctuations (Fig. 3). To elucidate the molecular basis of these differences, RNA-seq analysis was performed on 36 cassava and 36 potato samples, each with three biological replicates. Sequencing generated high-quality data, with cassava libraries yielding an average of 6.68 Gb per sample and detecting 27,701 genes, while potato libraries produced 6.42 Gb per sample with 26,714 genes; Q20 and Q30 values exceeded 90%, confirming data reliability (Supplementary Tables S2 & S3). Differential expression analysis revealed contrasting patterns between species: in cassava, the heat-tolerant cultivar KU50 exhibited 186 continuously up-regulated genes and 426 down-regulated genes, whereas the sensitive SC205 had 285 up-regulated and 611 down-regulated genes (Fig. 4a–d). Conversely, in potato, the heat-tolerant cultivar FAVORITA (F) displayed 660 up-regulated and 1,092 down-regulated genes, while the sensitive Qingshu No. 9 (qing9) showed 357 up-regulated and 496 down-regulated genes (Fig. 4e–h). Notably, cassava thermosensitive varieties exhibited more differentially expressed genes (DEGs) than heat-tolerant varieties, whereas the opposite trend was observed in potato. Functional enrichment analysis further revealed species-specific adaptations: cassava DEGs were primarily enriched in protein folding, heat response, and endoplasmic reticulum-associated protein processing, while potato DEGs were associated with antioxidant detoxification, photosynthesis, and light-harvesting pathways (Fig. 5a, b). Key differences between the two species included enrichment of genes associated with the photosynthetic electron transport chain and phenylalanine biosynthesis, which may contribute to the enhanced heat resilience observed in cassava.
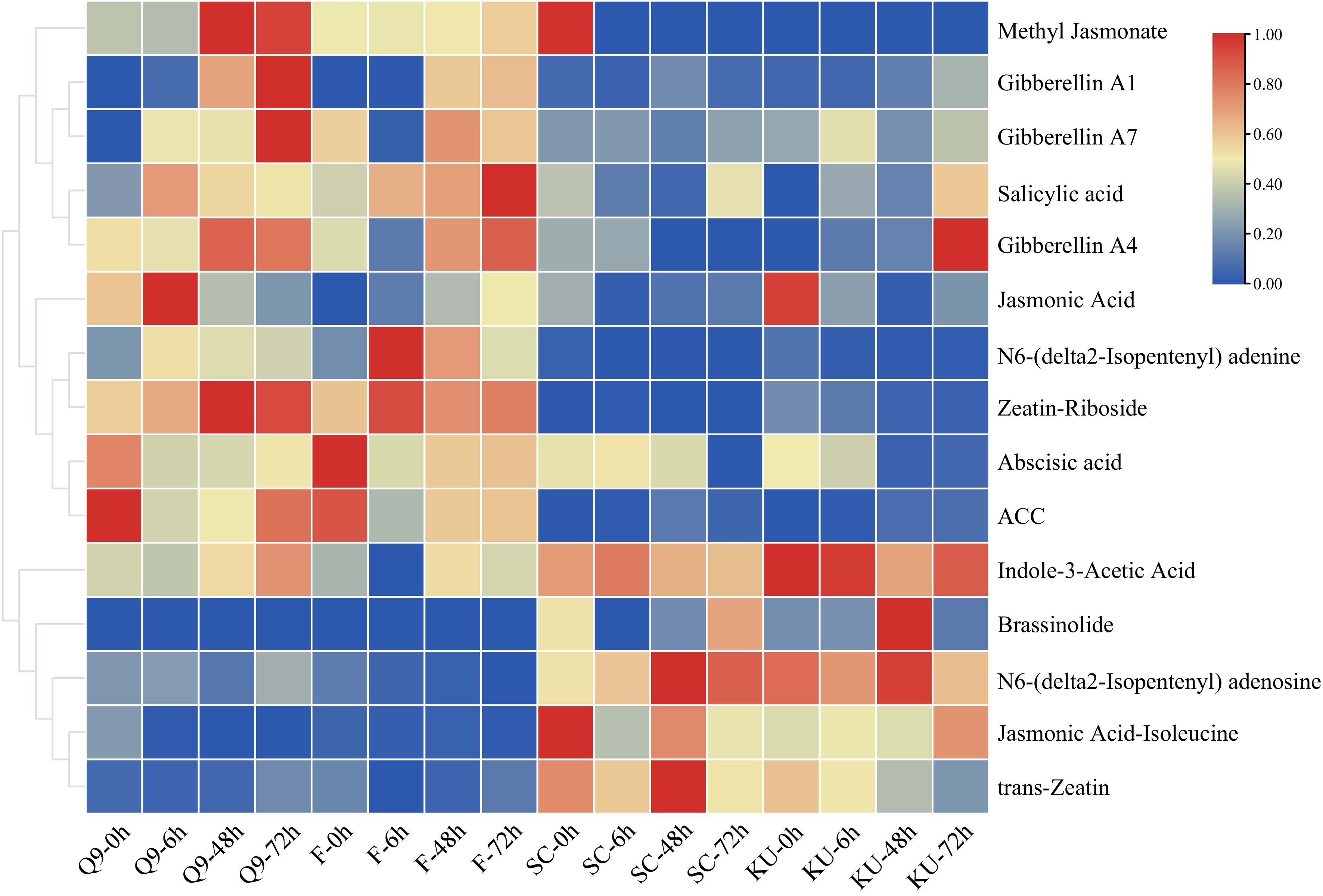
Figure 3.
Heatmap of hormone content changes in potato and cassava under high-temperature treatment. qing9 denotes Qingshu No. 9, F denotes FAVORITA, SC denotes cassava SC205, and KU denotes cassava KU50.
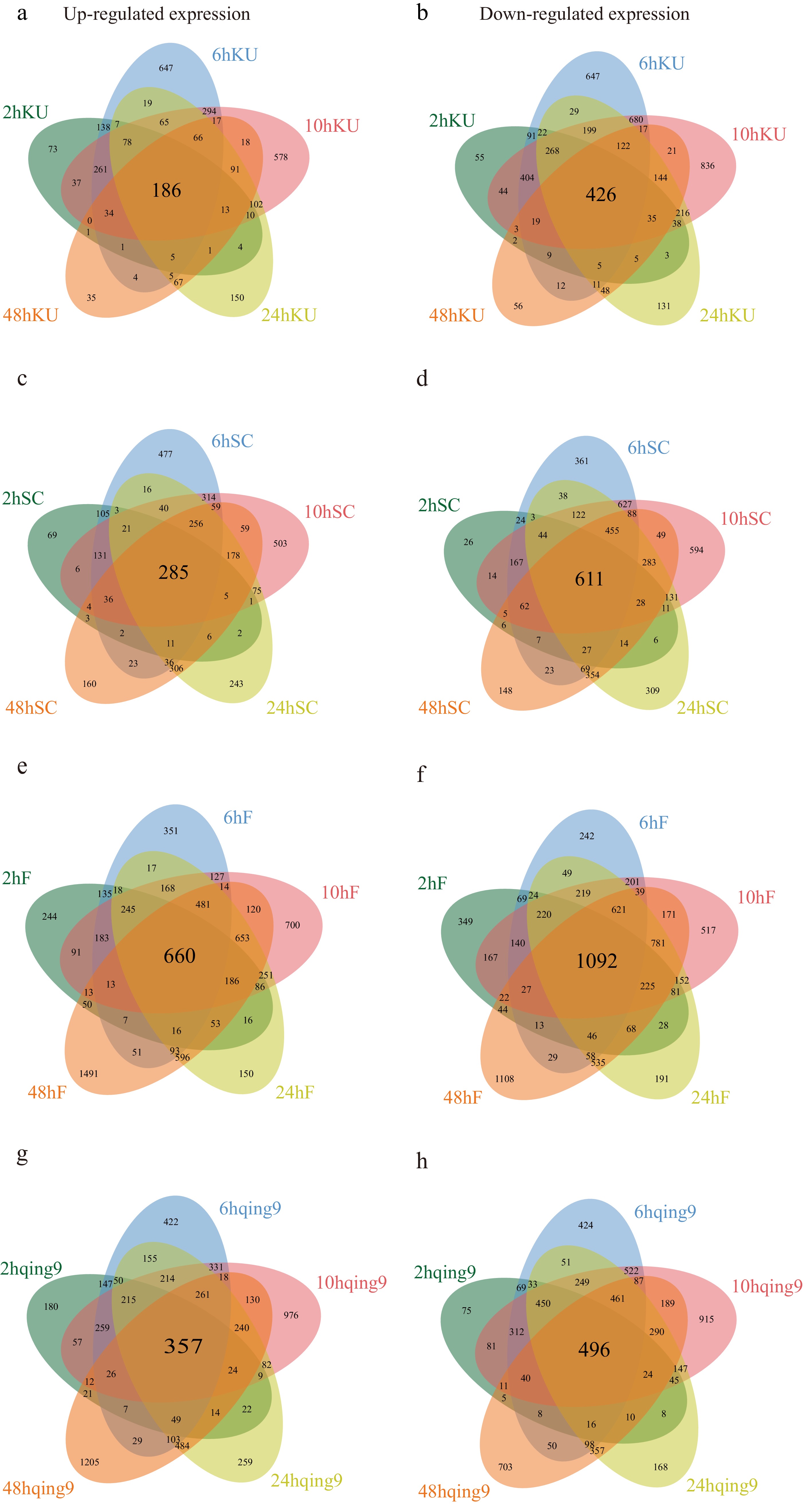
Figure 4.
Venn diagrams of differentially expressed genes (DEGs) in cassava and potato under heat treatment. (a) Venn diagram of up-regulated DEGs in KU50 (heat vs control). (b) Venn diagram of down-regulated DEGs in KU50 (heat vs control). (c) Venn diagram of up-regulated DEGs in SC205 (heat vs control). (d) Venn diagram of down-regulated DEGs in SC205 (heat vs control). (e) Venn diagram of up-regulated DEGs in FAVORITA (heat vs control). (f) Venn diagram of down-regulated DEGs in FAVORITA (heat vs control). (g) Venn diagram of up-regulated DEGs in Qingshu No. 9 (heat vs control). (h) Venn diagram of down-regulated DEGs in Qingshu No. 9 (heat vs control).
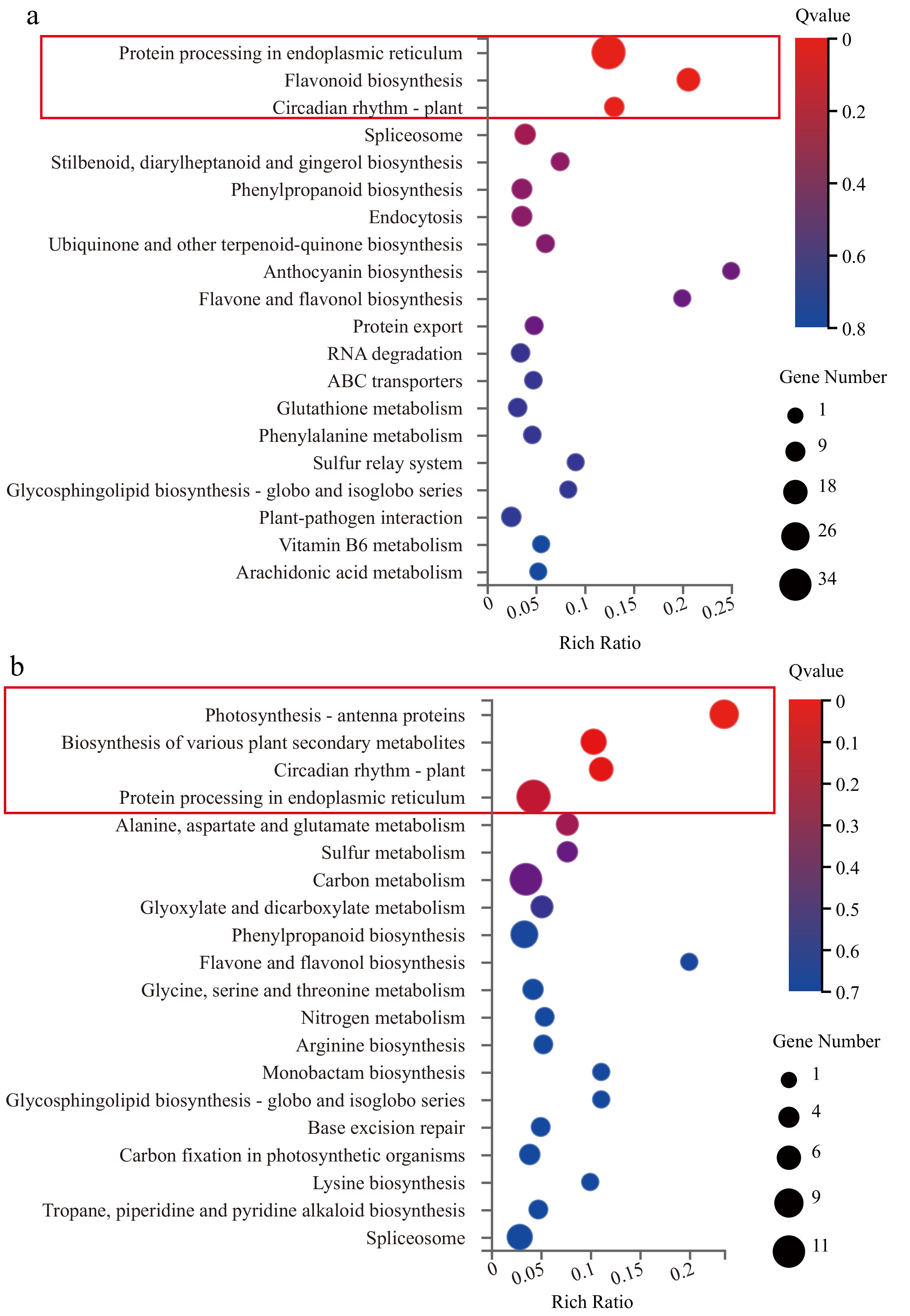
Figure 5.
Comparison of enriched KEGG metabolic pathways between cassava and potato. (a) KEGG enrichment of consistently expressed differential genes shared in cassava. (b) KEGG enrichment of consistently expressed differential genes shared in potato.
Comparative analysis of heat-responsive transcription factors in cassava and potato
-
Transcription factor (TF) prediction revealed distinct yet overlapping patterns of TF family representation in cassava and potato under high-temperature (HT) stress, highlighting their critical roles in stress signaling and transcriptional regulation. In cassava, the most abundant TF families were MYB (12%), bHLH (9%), NAC (7%), FAR1 (5%), HD-ZIP (5%), and HSF (3%) (Fig. 6a). These TF families are widely implicated in regulating plant responses to heat stress: MYB factors participate in secondary metabolism and stress signal transduction; bHLH members modulate growth and environmental adaptation; NAC TFs are known for controlling senescence and stress tolerance; FAR1 and HD-ZIP families are involved in light signaling and development; and HSFs (heat shock factors), although fewer in number, act as master regulators of heat stress by inducing heat shock protein (HSP) expression. In potato, the dominant TF families included MYB (12%), bHLH (8%), ERF (8%), C2H2 (7%), WRKY (7%), and HSF (2%) (Fig. 6b). The higher representation of ERF and WRKY transcription factor families in potato indicates a predominant activation of ethylene-mediated and defense-related signaling pathways under high-temperature stress. In contrast, the enrichment of NAC and HD-ZIP families in cassava highlights distinct regulatory mechanisms underlying its adaptive responses. Despite these differences, both species exhibited MYB and bHLH as core TF families, reinforcing their conserved roles in heat stress response. Collectively, these findings demonstrate that TF-mediated regulation in cassava is more closely associated with protein homeostasis and growth adjustment, whereas potato relies more on defense signaling and ROS detoxification pathways during heat stress.

Figure 6.
Comparison of transcription factors under high-temperature stress between cassava and potato. (a) Statistical chart of transcription factors in cassava under heat stress. (b) Statistical chart of transcription factors in potato under heat stress.
Comparative analysis of hormone biosynthesis pathways in cassava and potato under heat stress
-
To elucidate the hormonal regulation underlying differential heat stress responses, hormone content measurements were integrated with transcriptomic profiling, focusing on ethylene and abscisic acid (ABA) biosynthesis pathways. Ethylene biosynthesis in plants primarily involves three enzymatic steps: conversion of methionine to S-adenosylmethionine (SAM), catalysis by 1-aminocyclopropane-1-carboxylic acid synthase (ACS), the rate-limiting enzyme, and conversion of ACC to ethylene by ACC oxidase (ACO). In cassava, 18 differentially expressed genes (DEGs) were identified in this pathway, including six SAM-related, eight ACS-related, and four ACO-related genes (Fig. 7a). In contrast, potato exhibited 25 DEGs, comprising four SAM-related, 13 ACS-related, and eight ACO-related genes (Fig. 7b). The expression of ACS and ACO genes showed strong time-dependent changes, which aligned with variations in ACC content observed under HT stress. Similarly, in the ABA biosynthesis pathway, cassava displayed 16 DEGs, notably including nine NCED-related genes—encoding 9-cis-epoxycarotenoid dioxygenase, a key rate-limiting enzyme in ABA synthesis—along with ZEP, ABA2, and AAO3 (Fig. 7c). Potato, by comparison, had 13 DEGs in this pathway, including five NCED-related genes, and additional ZEP, NSY, ABA2, and AAO3 genes (Fig. 7d). Importantly, cassava exhibited a greater proportion of up-regulated genes in both pathways than potato, consistent with the observed hormonal profiles where ACC levels rose significantly while ABA declined under HT stress. These findings indicate that cassava and potato employ distinct hormonal regulation strategies, with cassava maintaining a stronger ethylene response and a more dynamic ABA regulatory network. Furthermore, the identification of ACS and NCED as core rate-limiting enzymes provides promising genetic targets for engineering enhanced heat tolerance in crop species.
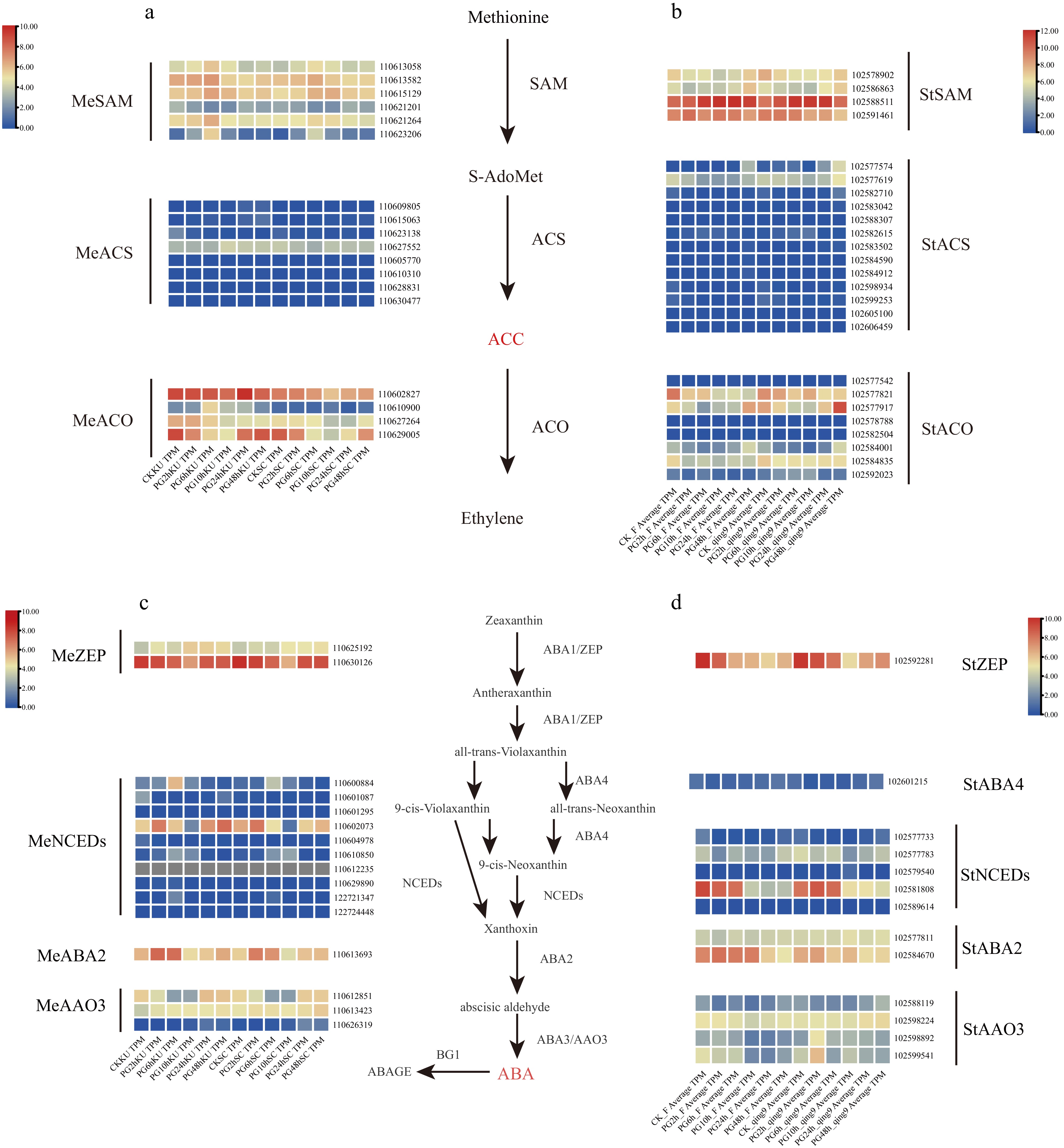
Figure 7.
Hormone biosynthesis pathway responses in cassava and potato under high-temperature stress (Heatmap: red, up-regulation; blue, down-regulation). (a) Ethylene biosynthesis pathway in cassava under heat stress. (b) Ethylene biosynthesis pathway in potato under heat stress. (c) ABA biosynthesis pathway in cassava under heat stress. (d) ABA biosynthesis pathway in potato under heat stress.
WGCNA reveals distinct heat-responsive regulatory modules in cassava and potato
-
To further dissect gene co-expression patterns associated with heat stress, WGCNA was employed on persistently differentially expressed genes in cassava and potato. In cassava, the analysis identified 13 distinct modules, among which the blue module exhibited a strong positive correlation with heat shock factors (HSFs) and heat shock proteins (HSPs). Interestingly, most genes in this module were down-regulated under high-temperature conditions, suggesting a regulatory mechanism aimed at moderating excessive protein-folding responses. Another key module in cassava, the dark purple module, was enriched in genes related to the photosynthetic electron transport chain, which showed a consistent up-regulation trend (Fig. 8a). This indicates that cassava maintains photosynthetic efficiency as part of its heat tolerance strategy.
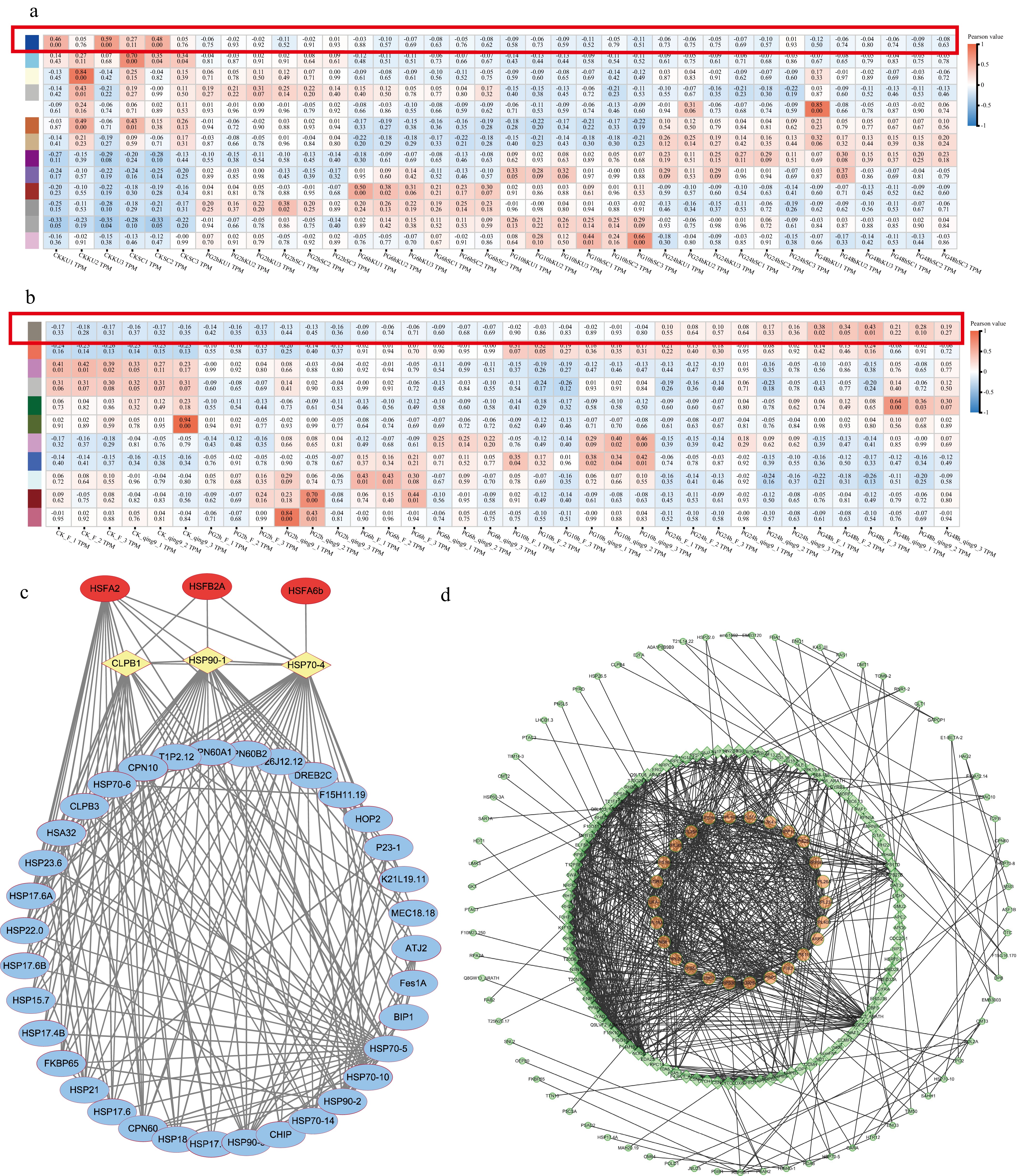
Figure 8.
Co-expression network construction of consistently expressed genes under high-temperature stress in cassava and potato. (a) Module partition of the co-expression network of consistently expressed genes in cassava under heat stress. (b) Module partition of the co-expression network of consistently expressed genes in potato under heat stress (blue, down-regulated genes; red, up-regulated genes). (c) Network construction of the cassava blue module. (d) Network construction of the potato antiquewhite4 module.
In potato, WGCNA identified 11 modules, with the antiquewhite4 module showing strong association with HSFs and HSPs, but in contrast to cassava, these genes were predominantly up-regulated under heat stress, suggesting an intense activation of the heat shock response (Fig. 8b). Additionally, the purple and grey modules in potato were enriched for photosynthesis-related genes, which displayed marked down-regulation, implying significant impairment of photosynthetic capacity under high-temperature stress.
Further network topology analysis revealed species-specific regulatory hubs. In the blue module of cassava, key regulators such as HSFA2, HSFB2A, and HSFA6b were identified, which directly regulated downstream HSP genes or exerted control through intermediates including CLPB1, HSP90-1, and HSP70-4, thereby establishing a robust yet precisely coordinated heat stress response network. Conversely, the potato antiquewhite4 module was dominated by genes involved in secondary metabolite biosynthesis and reactive oxygen species (ROS) detoxification, indicating a reliance on chemical defense and antioxidant systems rather than stable protein homeostasis. This divergence underscores fundamentally different adaptive strategies: cassava prioritizes photosynthetic stability and controlled HSP expression, while potato relies on metabolite reprogramming and stress scavenging systems to cope with heat stress (Fig. 8c & d).
Validation of cassava and potato RNA-seq data by RT-qPCR
-
To verify the reliability of the RNA-seq results, a subset of heat-responsive genes was selected for RT-qPCR validation. Expression trends between RT-qPCR and RNA-seq showed strong consistency, with correlation coefficients (R2) above 0.90 (Fig. 9). Genes such as HSP70 and HSFA2 were significantly upregulated in potato but only moderately induced in cassava, matching the transcriptome profiles. Similarly, ACS-related genes exhibited higher induction in cassava compared to potatoes, reflecting species-specific hormonal regulation patterns. These results confirm that the transcriptomic data accurately represent gene expression changes under high-temperature stress, providing a robust basis for subsequent analyses.
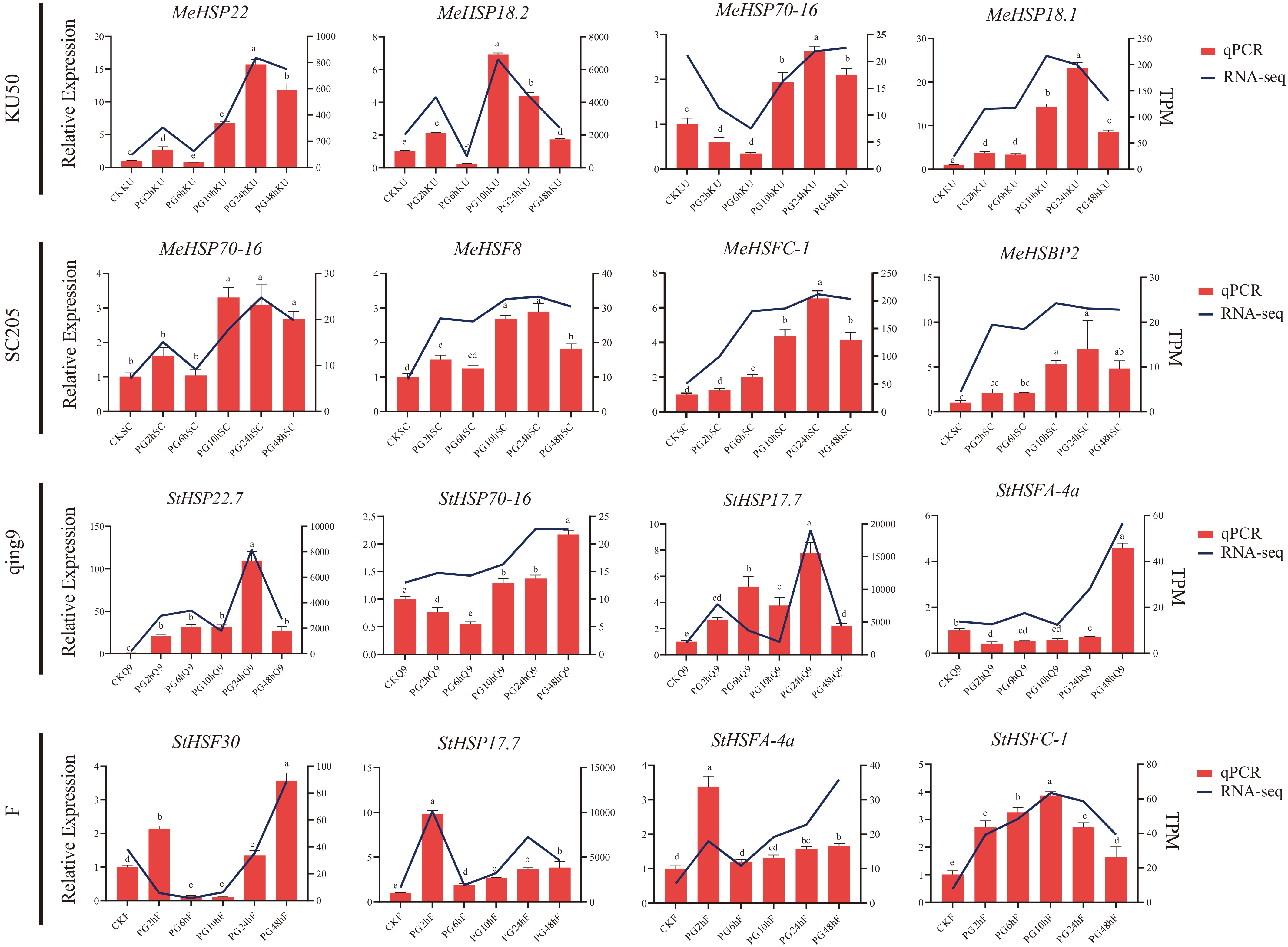
Figure 9.
RT-qPCR validation of differentially expressed genes in heat-stressed leaves of cassava and potato. Bars represent gene expression levels determined by RT-qPCR; lines indicate expression trends from RNA-seq data. Lowercase letters in the figures/tables were used to indicate the results of Tukey's HSD test; means bearing different letters differ significantly at p < 0.05.
-
The increasing occurrence of global high-temperature events poses a serious challenge to crop productivity, making the understanding of plant heat tolerance mechanisms a research priority. While most studies focus on temperate crops, the unique adaptive strategies of tropical species remain largely unexplored. Tropical crops, through long-term acclimation to high-temperature and high-light environments, have evolved specialized molecular mechanisms that minimize heat-induced damage and sustain growth. This study selected cassava and potato for comparative transcriptome analysis because they are both important members of the Euphorbiaceae and Solanaceae families, respectively, and share similar physiological processes such as starch biosynthesis and tuberous organ development, while exhibiting distinct adaptations to environmental stresses. Cassava is a typical tropical crop with strong heat and drought tolerance, whereas potato is a temperate crop that is more sensitive to heat stress. By comparing their physiological, hormonal, and transcriptomic responses under high-temperature conditions, this study aimed to identify conserved and divergent molecular mechanisms underlying heat stress adaptation in storage-root and tuber crops. This comparison provides a scientific basis for understanding the evolution of heat tolerance and offers potential genetic targets for improving stress resilience in both tropical and temperate crops.
Phenotypic adaptation strategies
-
Maintaining cell wall integrity is a well-established strategy for enhancing thermotolerance in plants such as Arabidopsis, maize, and coffee[26−28]. In the early phases of heat stress, the cell wall not only serves as a mechanical barrier but also functions as a sensor, initiating signal transduction and structural remodeling—such as increased cellulose deposition—to improve thermal resistance[29,30].
Compared to potato, cassava exhibits distinct structural features that contribute to its heat resilience. These include more compact cellular organization, thicker cell walls enriched in pectin and cellulose, and higher degrees of stem lignification and cutinization[31−34]. Such anatomical traits strengthen cell wall rigidity and reduce vulnerability to thermal damage. In this study, visual assessments under high-temperature conditions clearly showed that cassava maintained superior morphological stability relative to potato (Figs 1e & 2c). These structural advantages likely play a key role in helping cassava maintain cellular homeostasis and withstand prolonged heat stress.
Hormonal regulation under heat stress
-
Hormonal analysis revealed distinct adaptation strategies. To withstand heat stress, plants have evolved hormone signal transduction pathways that relay the heat signal and orchestrate downstream responses, enabling them to adapt to environmental pressure. Salicylic acid (SA) has been widely recognized for its role in enhancing thermotolerance by promoting heat shock protein (HSP) accumulation, reactive oxygen species (ROS) scavenging, and proline biosynthesis[35]. Abscisic acid (ABA), a central stress-response hormone, modulates downstream heat shock factors (HSFs) and sucrose metabolism, contributing to ROS regulation and reduction of heat-induced peroxides[36,37]. Cytokinin (CTK) levels often rise early in heat stress, concurrent with a decline in ABA. This dynamic shift facilitates protein targeting to chloroplasts, helping restore photosynthesis and activate root antioxidant enzymes, ultimately improving heat resilience[38−40]. In spring barley, BR-deficient or BR-insensitive mutants display lower HSP expression under heat stress, suggesting that brassinosteroids (BRs) are involved in heat signal modulation[41]. Ethylene biosynthesis is also differentially regulated under heat stress and plays a dual role in resource allocation and thermotolerance, as shown in rice[42,43].
This data shows that cassava maintains a more balanced hormonal profile under heat stress compared to potato. In cassava, SA levels remained consistently elevated, aligning with its protective role. The early rise in CTK alongside reduced ABA likely reflects a strategic redistribution of resources from storage to vegetative tissues, supporting growth and stress mitigation (Supplementary Figs S1c, i & h). Elevated ethylene may promote selective leaf senescence, reducing metabolic load (Supplementary Fig. S1g). Notably, BR responses varied by genotype, suggesting differential regulation among cassava varieties (Supplementary Fig. S1f).
In contrast, potato showed sharp declines in ABA, jasmonic acid (JA), and ethylene, reflecting a lack of active heat adaptation (Supplementary Figs S1k, n & o). Rising auxin and cytokinin levels under prolonged stress may represent a compensatory attempt to repair damaged tissues through cell division, though high concentrations of ZT and IAA have been linked to inhibited tuberization (Supplementary Figs S1i & m). GAs increased in potato, likely contributing to excessive shoot elongation and further inhibiting tuber development (Supplementary Fig. S1l).
Transcriptomic strategies for heat tolerance
-
Transcriptome profiling revealed that cassava adopts a resource-conservation strategy under heat stress, downregulating energy-intensive processes while enhancing mechanisms that sustain essential functions. Genes related to heat shock proteins (HSPs) in cassava showed little or negative induction, likely due to high basal expression, allowing rapid protein homeostasis without activating de novo synthesis (Supplementary Fig. S2a). Simultaneously, upregulation of photosynthetic electron transport genes suggests maintenance of energy production through cyclic electron flow, which mitigates heat-induced photoinhibition (Supplementary Fig. S3a). This trait also aligns with previous findings: under heat stress, enhanced expression of the cyclic electron transport chain helps crops mitigate heat-induced damage[44].
Conversely, potato exhibited a reactive stress response characterized by strong induction of HSP genes, upregulation of ER protein folding pathways, and enhanced synthesis of secondary metabolites (Fig. 5b). WGCNA confirmed that potato relies heavily on transcriptional and translational activation, DNA repair, and antioxidant detoxification modules, reflecting an energy-intensive repair strategy rather than proactive stress mitigation (Fig. 8d). This divergence implies that evolutionary adaptation to tropical environments has endowed cassava with constitutive and energy-efficient protective mechanisms, whereas potato relies predominantly on energy-intensive post-stress repair processes.
Overall, these findings demonstrate that cassava possesses inherently proactive and resource-efficient mechanisms for heat tolerance, involving structural robustness, moderate hormonal adjustments, and optimized transcriptomic regulation. In contrast, potato depends on high-energy repair processes, which may explain its greater sensitivity to heat stress. Understanding these divergent strategies provides a foundation for introducing heat tolerance traits from tropical crops into temperate species to enhance resilience under global climate change.
-
This study reveals distinct heat stress responses between cassava and potato at physiological, hormonal, and transcriptomic levels. Cassava showed stronger heat tolerance with stable growth and moderate hormone changes, while potato suffered severe damage and larger hormonal fluctuations. Transcriptomic analyses highlighted species-specific pathways: cassava focused on protein folding and ER-associated processes, whereas potato emphasized antioxidant detoxification and photosynthesis. Differential regulation of key heat-responsive transcription factors and hormone biosynthetic genes provided molecular evidence for the enhanced thermotolerance observed in cassava. Co-expression network analysis further illustrated differing regulatory modules between the two crops. Validation by RT-qPCR confirmed the reliability of the transcriptome data. These insights provide valuable targets for breeding heat-resilient crops amid global climate challenges.
Future research can further utilize multi-omics technologies (such as genomics, proteomics, and metabolomics) to comprehensively elucidate the interaction relationships and regulatory networks of heat tolerance-related genes, and clarify their specific functions and mechanisms in the response to heat stress.
This study was supported by the Hainan Provincial Natural Science Foundation (Grant No. 324MS122), the National Natural Science Foundation of China (Grant No. 32360458), and the Startup Funds for the Double First-Class Disciplines of Crop Science at Hainan University (Grant No. RZ2100003362), all of which contributed to the successful completion of this research.
-
The authors confirm contributions to the paper as follows: study conception and design: Guo X, Wang W; data collection: Luo J, Li H, Xiao J, Wang S; analysis and interpretation of results: Luo J, Wang S, Guo X, Wang W; draft manuscript preparation: Luo J. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0025-0031)
-
Received 5 August 2025; Accepted 11 October 2025; Published online 10 November 2025
-
Integrated hormone and transcriptome profiling reveals a steady-state strategy in cassava vs dramatic fluctuation in potato under heat stress.
WGCNA identified 13 cassava and 11 potato co-expression modules, uncovering opposite regulation of HSPs/TFs (down), and photosynthetic-lignin genes (up).
Validated DEGs provide a ready-to-use molecular resource for breeding heat-tolerant tropical and temperate crops.
- Supplementary Table S1 Primers used in this study.
- Supplementary Table S2 Cassava transcriptome sequencing data and alignment results.
- Supplementary Table S3 Potato transcriptome sequencing data and alignment results.
- Supplementary Fig. S1 Comparison of hormone bar charts between cassava and potato under high-temperature treatment.
- Supplementary Fig. S2 Comparative heatmap clustering of HSPs in cassava and potato under heat stress.
- Supplementary Fig. S3 Comparative heatmap clustering of photosynthetic electron transport chain components in cassava and potato under heat stress.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Luo J, Xiao J, Li H, Wang S, Wang W, et al. 2025. Comparative hormonal and transcriptomic analysis of heat stress responses in cassava and potato. Tropical Plants 4: e036 doi: 10.48130/tp-0025-0031 

Comparative hormonal and transcriptomic analysis of heat stress responses in cassava and potato
- Received: 05 August 2025
- Revised: 09 October 2025
- Accepted: 11 October 2025
- Published online: 10 November 2025
Abstract: Improving crop heat tolerance is essential under the increasingly frequent extreme temperature conditions worldwide. Cassava (Manihot esculenta Crantz) is a tropical crop with strong resistance to high temperature and high light, whereas potato (Solanum tuberosum L.) is a temperate crop highly sensitive to heat stress. To elucidate the differences in heat stress responses between cassava and potato, this study combined transcriptome sequencing with hormonal profiling. The results revealed that cassava exhibited significant changes in hormone signaling with relatively lower amplitude, whereas potato showed large fluctuations in both hormone levels and signaling pathways. Transcriptome analysis identified numerous differentially expressed genes (DEGs) in cassava cultivars KU50 and SC205 and in potato cultivars FAVORITA and Qingshu No. 9. Weighted gene co-expression network analysis (WGCNA) identified 13 regulatory modules in cassava and 11 in potato. In cassava, heat-responsive transcription factors and heat shock proteins (HSPs) were generally downregulated, while genes involved in photosynthetic electron transport and lignin biosynthesis were upregulated; the opposite trend was observed in potato. RT-qPCR validation confirmed the transcriptome data. These findings provide valuable gene resources for breeding heat-tolerant crop varieties and offer insights into the molecular mechanisms underlying heat tolerance in tropical vs temperate crops.
-
Key words:
- Heat stress /
- Potato /
- Cassava /
- Transcriptome /
- Hormonal regulation /
- Heat tolerance mechanism