









-
Metallic cadmium (Cd) is a by-product of lead, copper, and zinc refining plants. It is recorded that about 20 million hectares of land are polluted by cadmium and other heavy metals in China[1]. ABC-binding cassette (ABC) transporter proteins exist in all organisms, and their family members are mainly divided into three categories: Full molecular transporters, half-molecular transporters, and soluble transporters[2,3]. They release energy by binding and hydrolyzing adenosine triphosphate (ATP) and transport various substrates across cell membranes. The substrates which they transport mainly include metal ions, polypeptides, small proteins, inorganic molecules, alkaloids, and lipids[4,5]. The full molecular transporter consists of two transmembrane domains (TMDs) and two nucleotide-binding domains (NBDs). However, the half-molecular transporter has merely one TMD and one NBD[2,6,7].
The ABC proteins of eukaryotes can be grouped into eight primary subfamilies (A_H)[6]. Both Oryza sativa and Arabidopsis thaliana have more than 120 ABC transporter genes within their genomes[8]. In Arabidopsis, 21 whole-molecule ABCB subfamily proteins were identified and grouped into three clusters[9]. The primary function of AtABCB21 is to regulate auxin distribution and serve as a supplement to the main auxin transporters ABCB1 and 19 in the ABCB subfamily[10]. The loss-of-function mutants of ABCB15, 16, 17, 18, and 22 (ABCB15–22), which are novel auxin-transporting ABCBs, showed strongly reduced the lateral root density[11]. In plants, distribution of auxin is done by auxin transporters, which include PIN, PILS, Aux1/LAX, and some other ABCB members[12].
Furthermore, under heavy metal pollution, plants regulate their growth by altering the resource allocation to various organs to better adapt to adverse conditions and survive[3]. The tap-type transporter AtABCB27 (ALS1/TAP2) is localized to the vacuolar membranes and is crucial for root development in environments with severe aluminum pollution[13]. Microscopic analysis with a Cd-sensitive probe revealed that AtABCC1 and AtABCC2 could improve plants' resistance to Cd by transferring Cd from the cytoplasm to the vacuole[14]. In rice, the TAP transporter OsABCB27 (OsALS1), which is localized in the vacuolar membrane, is involved in the aluminum stress response[15]. OsABCG36, a Cd efflux transporter, is considered to have functionally redundancy with OsABCG44[16]. In rice, the expression of OsABCG53/OsPDR20 is necessary for decreasing the accumulation of cadmium[17]. However, the expression of tomato ABCB transporter genes in response to Cd stress remains unclear.
In this study, we provide comprehensive information on the B subfamily of ABC proteins in tomato under cadmium stress. Analyses of the exon–intron organization, chromosomal distribution, motif analysis, comprehensive phylogenetics, and distribution of cis-regulatory elements have been attempted. This study may provide a foundation for further functional analysis of the ABCB transporter's role in resistance to cadmium in S. lycopersicum.
-
The characteristic domains of the ABC transporter family were obtained from the Pfam database (
http://pfam.xfam.org ). Biological sequence analysis with the HMMER software was carried out to construct the hidden Markov model (HMM) and acquire an amino acid database for ABC transporters[18]. The NCBI website and SMART software were used to identify the candidate proteins for AAA domains and ABC2 transmembrane domains and to eliminate the sequences for the missing domains[19].Phylogenetic analysis
-
A multiple-step strategy was utilized to construct the phylogenetic tree. The analysis of large protein families is particularly challenging, since they may include several domains and repetitive sequences.
Amino acid and nucleotide sequences of the tomato ABC family were compared with ClustalW. Through the neighbor-joining (NJ) tree methods and maximum likelihood, the rootless phylogenetic tree was built using MEGA 6.0 software, and each had 1,000 bootstrap replicates. The differences between the nucleotides of the SlABCB genes were computed with MEGA 6.0[20,21]. The evolutionary tree was further developed via the Evolview website (
www.evolgenius.info/evolview ). The genome annotation files of Arabidopsis (TAIR10.1) and tomato (SL 4.0) were obtained from NCBI. MCScanX was used to investigate the whole-genome collinearity of tomato and Arabidopsis[22], and TBtools was utilized to map the whole-genome collinearity.Identification of conserved motifs and gene structure
-
With the MEME (Multiple EM for Motif Elicitation) web server, 10 motifs of the SlABCB protein sequences were identified. MEGA X was used to construct their evolutionary relationships[23]. The evolutionary relationships, domains, and motifs of the tomato ABCB gene family were analyzed and integrated using TBtools[24].
Analysis of chromosomal localization and promoter elements
-
According to the tomato genome annotation files (GFF4.0, general feature format), versions ITAG4.0 and SL3.0 of the genome sequence were downloaded from the Sol Genomics Network (SGN,
https://solgenomics.net ). The location of the ABCB family members on the tomato chromosome was identified using TBtools software[24]. The ABCB family's physicochemical properties in tomato were predicted with the ExPASy-ProtParam tool.The promoter sequences of all ABCB genes located in the 2,000 bp sequence of the 5'-upstream region were downloaded from the SGN website. The PlantCARE database was used to predict the promoter elements. TBtools was subsequently used to construct heatmaps and distribution maps[25].
Plant material and cadmium treatments
-
The seeds surface of tomato (A57 variety) were disinfected for 30 s with 70% ethanol and then with 15% NaClO for 15 min. Before sowing on solid half-strength Murashige and Skoog media, the treated seeds were thoroughly rinsed with deionized water, and vernalizated for 2–4 days at 4 °C. Plants were grown in a greenhouse at a constant temperature (25 + 2 °C) and photoperiod (16/8 h light/dark cycle) for 7 days, and then transferred to half-strength modified Hoagland nutrient solutions[26] supplemented with CdCl2 to a final cadmium concentration of 10 μM. The leaves and roots of plants treated or not treated with cadmium were harvested after two hours of treatment[27]. Fresh tissues were quickly frozen with liquid nitrogen, and stored at −80 °C for RNA extraction.
Total RNA extraction and RNA sequencing
-
TRIzol RNA extraction reagent (Invitrogen, Carlsbad, CA, USA) was used to extract the total RNA from tomato plants grown in vitro, which was analyzed using RNA sequencing (RNA-Seq) and quantitative real-time polymerase chain reaction (qRT-PCR)[28]. RNase-free agarose gel electrophoresis was used to verify the RNA's integrity. RNA concentration was determined with an Agilent 2100 Bioanalyzer (Santa Clara, CA). For this, 2 μg of high-quality total RNA with a RIN ≥ 8 for each genotype and treatment were sent to BGI Genomics Co. Ltd. company (Shenzhen) to proceed with the RNA sequencing analysis.
RNA-Seq was carried out on an Illumina HiSeq 2500 platform. The raw reads obtained were filtered, and the remaining reads were mapped to the tomato genome SL3.0 (ITAG4.0). The expression level of each mRNA transcript was calculated as fragments per kilobase of transcript per million mapped reads (FPKM). According to the instructions of manufacturer, the first-strand cDNAs were synthesized by reverse transcription from RNAs treated with DNase I (Fermentas, Canada) with M-MuLV reverse transcriptase (Takara, Dalian, China). qRT-PCR was carried out with SYBR premix ExTaq (Takara) on an ABI7500 system.
qRT-PCR analysis
-
To validate the RNA-Seq results and identify the key candidates likely to be involved in cadmium tolerance, six upregulated SlABCB genes (SlABCB4, SlABCB8, SlABCB11, SlABCB15, SlABCB22, and SlABCB26) were chosen for qRT-PCR analysis. These particular genes were selected because of their strong induction under cadmium stress, their close phylogenetic relationship to functionally characterized ABCB transporters known to mediate heavy metal or auxin transport in other plants, and their promoter regions enriched with stress and hormone-related cis-elements, which imply their potential role in broader regulatory networks. Fluorescence quantitative RT-PCR was performed by adopting the 2−ΔΔCᴛ method, and the results are expressed as the relative expression levels of genes[29]. Table 1 lists the qRT-PCR primers. Heat maps were drawn using TBtools software, with different colors representing the expression levels relative to the average expression level. The original data were normalized by calculating the log2 value of the ratio of the measured expression level to the average expression level. Three biological replicates were used to determine the expression profiles, and the data are expressed as the average value and the standard error of the mean. At a significance threshold of p < 0.05, Duncan's multiple range tests were used to determine the differences between the means.
Table 1. Primers used in RT-qPCR analysis
Genes Locus Forward primers (5'→3') Reverse primers (5'→3') ABCB4 Solyc02g087870.2.1 CTACTATACAAATCCCGCAA TGCCAACATCATCCTCCGAA ABCB8 Solyc03g114950.2.1 AACCGTTCTTGTCATAGC GTCATCATGTGTGCCACT ABCB11 Solyc04g010310.2.1 CGTTCAATCTGCAAATCGGAA AAACTCACTGTTAGGACGACT ABCB15 Solyc06g009290.2.1 GGCTTTTGATGGGGGCATTG GATTTTCGCGTAGAGGGCCT ABCB18 Solyc06g009290.2.1 AACAAGCCTGAAATACCTG GAAGGCAATAAAAGTTACA ABCB20 Solyc09g008240.3.1 TCTTCATACGGTCGATCACC CGGCCAAACGAACTAGCTTG ABCB22 Solyc09g055350.2.1 GCCAGAATTATAAGCACTCT CATTCACAACCTTTGCACCA ABCB26 Solyc12g098840.3.1 GCATCTTTACTAGCGACACC TCTCCAATCATACCTCGGAC Ubiquitin Solyc10g005560.2.1 CACCAAGCCAAAGAAGATCA TCAGCATTAGGGCACTCCTT Yeast experiments
-
The primers used for amplifying the coding DNA sequences (CDSs) of the tomato ABCB candidate genes are listed in Table 1. The PCR-amplified fragments and the yeast expression vector pYES2 were restricted with of BamH1 and EcoR1. After the double enzyme digestion products were recovered with a gel extraction kit (Tiangen, Beijing, China), they were then subjected to connection via the T4 RNA ligase and then transformed into the corresponding yeast strain, as described by Elble[30]. The positive monoclonal yeast cells in fresh synthetic dextrose (SD) liquid medium were placed and cultured until they reached the logarithmic growth phase, then the yeast cells were collectedby centrifugation. The collected yeast particles were suspended in sterile water and then diluted to an optical density at 600 nm (OD600) of 1, 0.1, 0.01, or 0.001. Subsequently, equal volumes (15 μL) of the suspension were dropped onto medium plates containing different concentrations of heavy metals as indicated, and were incubated at 30 °C for approximately 7 d, with photos taken.
-
In order to clarify the tomato ABC protein families, a BLAST search of SGN genome database was carried out. We searched all the tomato ABC proteins with version ITAG4.0 of the annotation and version SL3.0 of the tomato genome. Ultimately, 154 members putatively encoding ABC gene transporter families were identified (Supplementary Fig. S1), and phylogenetic trees and functional predictions were subsequently generated. The tomato ABC family was grouped into eight subfamilies (A–I), including 9 ABCAs, 29 ABCBs, 26 ABCCs, 2 ABCDs, 2 ABCEs, 6 ABCFs, 70 ABCGs, and 10 ABCIs. Here, we focused on the ABCB subfamily because of its potential role in the Cd stress response.
With an average of 1,018 amino acids, the predicted proteins containing the ABCB gene ranged in quantity from 108 (SLABCB28) to 1,401 (SLABCB9). The molecular weights ranged from 11.8 to 142.7 kD, with an average of 110.28 kD. An essential physiological indicator of proteins is the isoelectric point (pI), which is based primarily on the percentage of acidic to basic amino acids. ABCBs had isoelectric points ranging from 5.84 to 9.45 (Table 2). The isoelectric points of most of the ABCB proteins (89.66%) were greater than 7, indicating that ABCB proteins might be alkaline proteins.
Table 2. Characterization of SlABCBs protein in tomato
Gene Name Gene ID AA MW (kD) pI Instability index Aliphatic index Structural model SlABCB1 Solyc02g071340.1 1,264 137.7 5.84 35.80 103.71 Whole-molecular transporter SlABCB2 Solyc02g071350.2 1,264 137.1 8.09 35.41 103.32 Whole-molecular transporter SlABCB3 Solyc02g087410.2 1,263 137.5 7.97 40.60 99.70 Whole-molecular transporter SlABCB4 Solyc02g087870.2 1,250 136.1 8.08 36.86 99.42 Whole-molecular transporter SlABCB5 Solyc03g005860.2 1,260 136.7 7.20 36.03 105.92 Whole-molecular transporter SlABCB6 Solyc03g093650.2 1,228 134.3 8.35 33.30 102.94 Semi-molecular transporter SLABCB7 Solyc04g010310.2 1,286 142.3 6.33 46.36 101.18 Whole-molecular transporter SLABCB8 Solyc06g009280.1 1,290 139.3 7.80 36.82 101.57 Whole-molecular transporter SLABCB9 Solyc06g009290.2 1,401 138.6 8.18 35.65 100.28 Semi-molecular transporter SLABCB10 Solyc06g072960.1 1,029 113.1 7.01 38.56 104.26 Semi-molecular transporter SLABCB11 Solyc07g018130.1 1,276 140.2 8.73 36.13 101.71 Semi-molecular transporter SLABCB12 Solyc07g064120.1 1,260 138.0 8.88 37.83 93.87 Semi-molecular transporter SLABCB13 Solyc08g076720.2 1,258 138.2 8.75 35.64 103.51 Semi-molecular transporter SLABCB14 Solyc09g008240.2 1,315 140.0 8.63 36.18 95.56 Whole-molecular transporter SLABCB15 Solyc11g067310.1 1,290 141.4 7.05 35.48 103.07 Whole-molecular transporter SLABCB16 Solyc12g098840.1 1,281 138.6 6.38 34.90 101.88 Semi-molecular transporter SLABCB17 Solyc12g098870.1 1,313 142.72 6.76 37.95 100.58 Whole-molecular transporter SLABCB18 Solyc11g067300.1 1,261 138.47 7.50 32.90 102.02 Whole-molecular transporter SLABCB19 Solyc05g013890.1 955 105.16 9.10 43.42 96.12 Whole-molecular transporter SLABCB20 Solyc03g026310.2 664 67.51 8.94 41.69 102.73 Whole-molecular transporter SLABCB21 Solyc03g114950.2 639 68.34 8.45 36.24 104.42 Semi-molecular transporter SLABCB22 Solyc03g122050.1 673 74.69 7.34 34.75 80.97 Semi-molecular transporter SLABCB23 Solyc03g122070.1 667 73.02 9.45 38.67 101.94 Whole-molecular transporter SLABCB24 Solyc09g009910.2 640 71.08 9.14 34.76 113.40 Whole-molecular transporter SLABCB25 Solyc09g055350.2 726 80.37 9.40 28.52 98.52 Semi-molecular transporter SLABCB26 Solyc00g304030.1 1,081 111.53 9.24 44.24 87.73 Whole-molecular transporter SLABCB27 Solyc12g049120.1 349 38.62 9.03 41.41 103.68 Quarter-molecular transporter SLABCB28 Solyc12g049130.1 108 11.8 8.79 35.36 101.12 Quarter-molecular transporter SLABCB29 Solyc12g070280.1 232 25.82 8.63 29.5 113.55 Quarter-molecular transporter Analysis of the number of exons, conserved motifs, and chromosomal locations
-
The conserved domains of the ABCB subfamily proteins in tomato were analyzed with TBtools software. The fifth motif was present in almost all ABCB subfamily proteins except the five genes SlABCB24, SlABCB25, SlABCB27, SlABCB28, and SlABCB29, suggesting that Motif 5 may be reasonably well conserved (Fig. 1). Each subfamily member showed some common and some unique motifs. In the cluster of SlABCB3 and SlABCB28, SlABCB28 had only one motif, namely Motif 4. As shown in Fig. 1 and Table 3, 10 distinct motifs were identified, and a schematic overview of the identified motifs is provided. Interestingly, a similar motif composition is shared by the same subgroups, indicating functional similarities among members of the same subgroups (e.g., SlABCB4 and SlABCB13, SlABCB15 and SlABCB18, etc.).

Figure 1.
Phylogenetic tree of ABCB family genes and distribution of conserved protein motifs.
Table 3. The sequences of conserved protein motifs in the ABCB gene family of the tomato genome.
Motif number Motif sequence Motif 1 AAKLANAHNFISGLPQGYETQVGERGVQLSGGQKQRIAIARAILKNPKILLLDEATSALDAESERIVQEALDR Motif 2 EIELKBVYFSYPSRPDVQILNGFSLKIPSGKTVALVGGSGSGKSTVISLJERFYDPQSGZVLJDGVBJKELQLKW Motif 3 MVGERGTQLSGGQKQRIAIARAILKBPRILLLDEATSALDAESERIVQEALDRIMINRTTVIVAH Motif 4 TVALVGESGSGKSTVISLJZRFYDPDSGEIYJDGIDIRKFNLKWLRQQMGLVSQEPVLFNDTIRENIAYGKEGAAEEE Motif 5 AGEKLIRRIRSMMFEKVLRMEIGWFDEEENSSGAJGARLSTDAATVRSLVGDRLALJVQBLATAIVALVIAFIASWRLALVMJAVQPL Motif 6 MVNRTTVVVAHRLSTIKGADVIAVIKBGVIVEKGTHDTLLN Motif 7 JRQQIGLVSQEPVLFATTIKENIAYGKEDATEEEIKEAIKLANAAKFIDKLPZGLDT Motif 8 KVSLKFVYLAIGVGVASFLZVACWTVTGERQASRIRCLYLKSVLRQDIGFFDTZTNT Motif 9 GMTLDVIKGBIEFKHVSFAYPTRPDVQILKDLSLTIPAGK Motif 10 AYEEASQIANEAVGNIRTVASFSAEEKVLELYK Gene Structure Display Server (GSDS) analysis of the structure of tomato ABCB family members helped to clarify the evolutionary relationships among the proteins. Genes within the same group usually have a similar structure, for example, SlABCB1 and SlABCB2, SlABCB16, and SlABCB17. All ABCB family members possessed 1 to 20 exons. SlABCB28 had only 1 exon, whereas SlABCB20, SlABCB21, and SlABCB25 had 15, 16, and 20 exons, respectively, which implied significant diversity in their exon–intron structure (Supplementary Fig. S2). This variation in SlABCBs was primarily caused by differences in exon–intron length, which is one of the predominant factors affecting gene size. Overall, the conserved motif compositions and similar gene structures of the ABCB members in the same group, together with the results of the phylogenetic analysis, strongly support the reliability of the group classifications.
The chromosomal distribution of the identified ABCB family members revealed that they were distributed throughout 10 chromosomes, with a maximum of six ABCB family members on chromosome 3. There were six members on chromosome 3, and five ABCB genes were located on chromosome 12. In addition, only one ABCB gene member were detected on chromosomes 1, 4, 5, and 8 (Supplementary Fig. S3). The distribution pattern of the ABCB subfamily on individual chromosomes also indicated certain physical regions with a relatively higher accumulation of multiple ABC gene clusters, such as chromosome 3, 11, and 12 at the lower end of the arms. Meanwhile, some ABCB gene clusters were present in the upper chromosome part, such as chromosome 6.
Promoter element analysis
-
Cis-acting elements within the promoter region act as transcription factors' binding targets, which govern gene expression in a tissue-specific, developmental stage-dependent, or stress-responsive manner. To systematically explore the regulatory-level functional diversity among the ABCB gene family members, 21 cis-elements were identified when the ABCB gene promoter sequences were submitted to the Plant CARE database (Fig. 2). Salicylic acid, abscisic acid (ABA), gibberellin, zein, methyl jasmonate (MeJA), and auxin were the main plant hormone-responsive elements in the upstream promoter region.

Figure 2.
Location of cis-elements in the ABCB genes' promoter sequences.
Anaerobic-, defense stress- and low temperature-induced response elements were among the stress-related cis-acting elements evaluated. The greatest proportion of cis-related elements related to plant development and growth among all the tomato ABCB family members were light-responsive elements, indicating that light responsive elements might play a significant role in photomorphogenesis.
Gene replication and collinearity analysis of the SlABCB genes
-
Investigating gene replication activities in the tomato genome allowed the investigators to better understand the amplification and evolutionary mechanism of the SlABCB gene family. TBtools software was used to investigate the collinearity of the tomato genome. The results revealed that the tomato ABCB family included a duplicated gene pair (SlABCB1 and SlABCB5) (Fig. 3). This suggests that there may be functional similarities between SlABCB1 and SlABCB5.
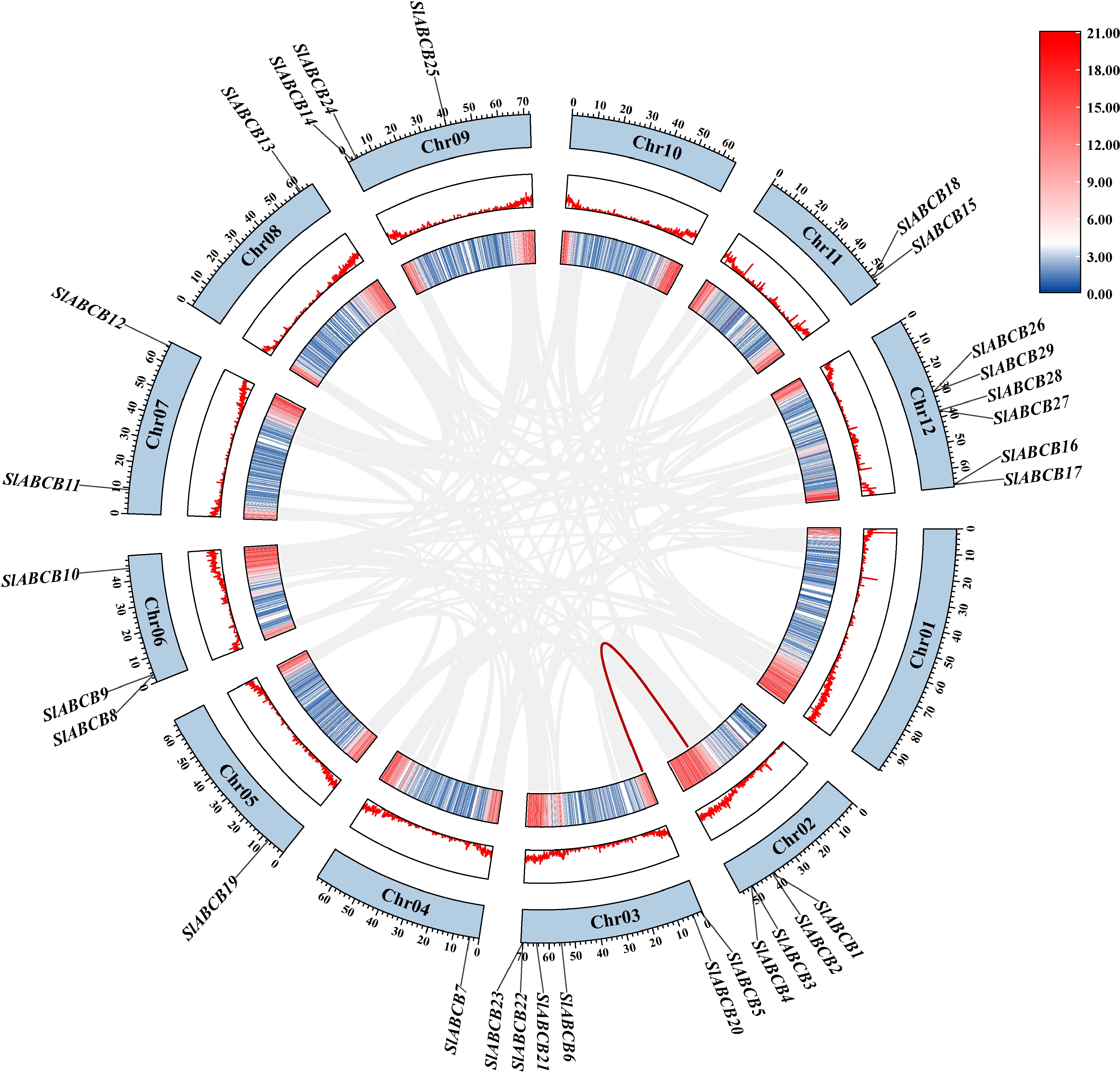
Figure 3.
Collinearity analysis of ABCB genes in tomato species from chromosome 1 to 12.
Figure 4 displays the collinear relationships of the duplicated gene pairs in the ABCB gene families in the genomes of tomato (S. lycopersicum), rice (Oryza sativa), eggplant (Solanum melongena), and Arabidopsis thaliana. Tomato and eggplant (from dicotyledonous Solanaceae) had the most collinear ABCB gene pairs, followed by tomato and Arabidopsis (from dicotyledonous plants), whereas tomato and rice (from monocotyledonous plants) had the fewest collinear ABCB gene pairs, in accordance with plant evolution laws. Ten pairs of ABCB genes were identified between tomato and Arabidopsis thaliana (Fig. 4a). SlABCB13, SlABCB19, SlABCB21, and SlABCB24 were the only pairs of the ABCB gene subfamily found in rice and tomatoes (Fig. 4b). Moreover, 16 pairs of ABCB genes were found to exist between tomato and eggplant (Fig. 4c). These revealed that ABCB genes between tomato and eggplant have relatively higher homology and highly conserved sequences.
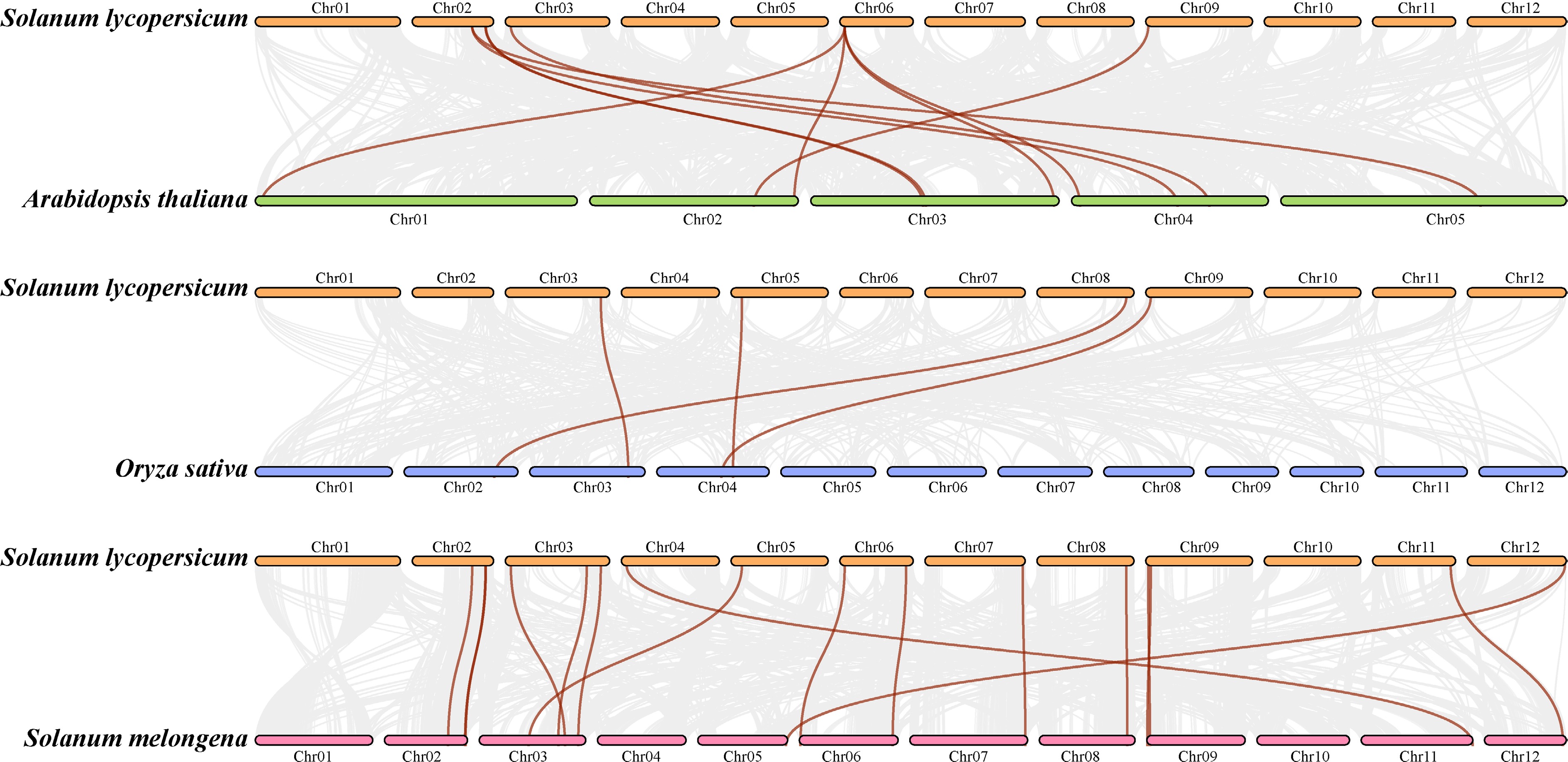
Figure 4.
Collinear correlation of the ABCB gene family between Solanum lycopersicum L. and (a) Arabidopsis thaliana, (b) Oryza sativa, and (c) Solanum melongena.
Phylogenetic relationships of the tomato ABCB gene family
-
A phylogenetic tree was constructed using the Arabidopsis ABCB family members to predict the functions of the 29 SlABCB genes in tomato and establish their homologies (Fig. 5). Tomato ABCB family members can be grouped into seven subfamilies (1–7), which include five, seven, nine, one, one, four, and two ABCB gene family members, respectively. There were two groups with the fewest members, Clust4 and Clust5, each including only one protein from the tomato ABCB family (SlABCB22 and SlABCB23). These results suggest that SlABCB22 and SlABCB23 may have specific functions in tomatoes (Fig. 5). SlABCB22 and SlABCB23 form their own unique clade each, which hints that they may have evolved separate, specialized jobs compared with the other family members. This idea is supported by our functional data for SlABCB22, which was strongly induced by cadmium and proved to be very effective at helping yeast survive cadmium stress. It might be a specialized transporter for this metal. The role of SlABCB23 is less clear, as it did not respond strongly to cadmium. Its unique evolutionary branch could mean it transports something else entirely, perhaps a specific compound that is not related to metal stress.
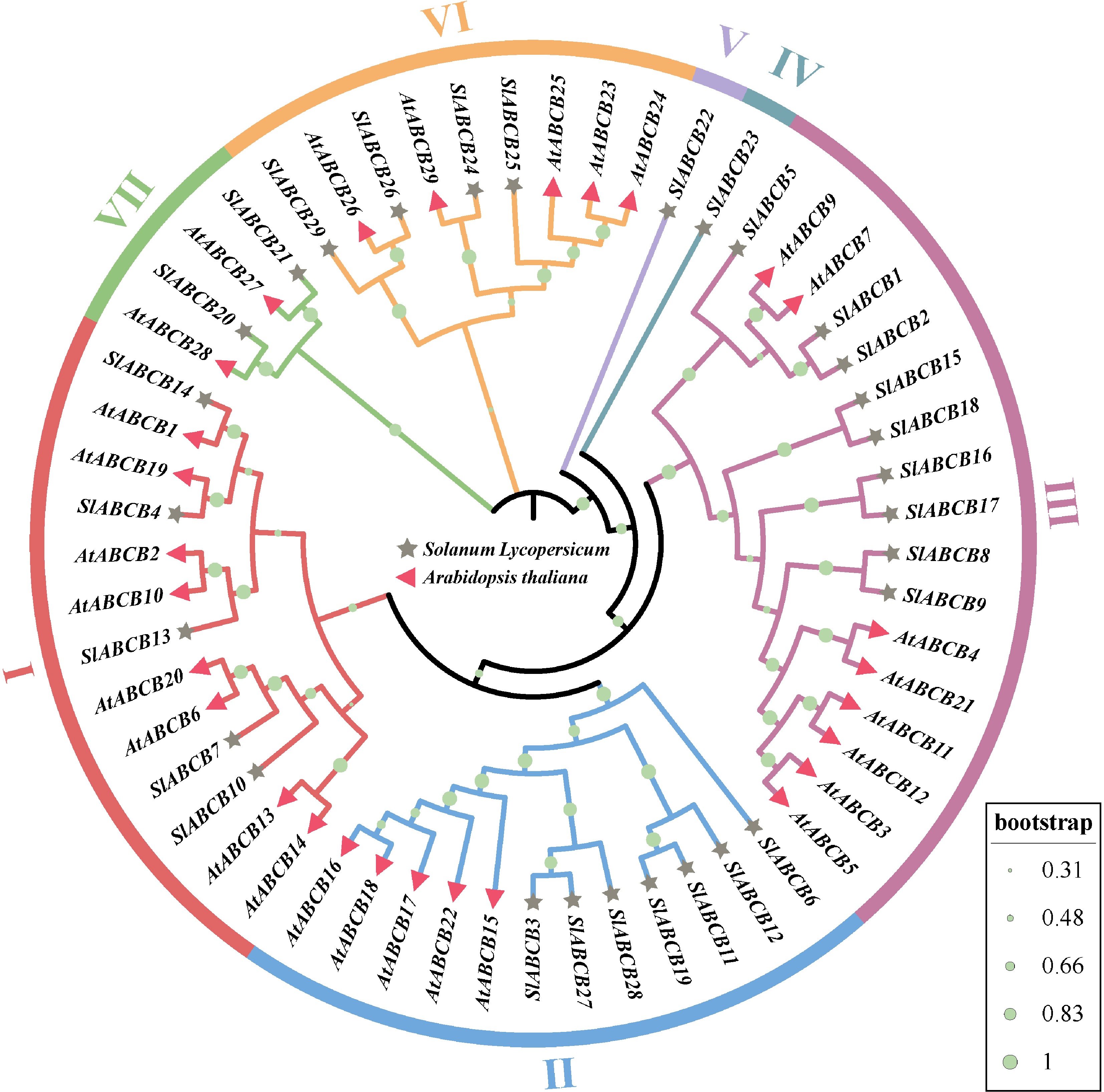
Figure 5.
Phylogenetic tree constructed using MEGA 6.0 by NJ the method based on a total of 58 ABCB proteins comprising 29 S. lycopersicum and 29 Arabidopsis ABCB proteins. The phylogenetic tree reveals seven main clusters (1–7).
Transcriptomic analysis of ABCB genes across all organs and all stages
-
The transcriptome data of ABCB genes were retrieved and examined to confirm their functions in the tissues and organs involved in reproduction. SlABCB9, SlABCB14, and SlABCB21 exhibited relatively high expression levels in S. lycopersicum roots, followed by SlABCB4, SlABCB7, and SlABCB25.
SlABCB14, SlABCB20, and SlABCB21 had higher mRNA expression levels than the other genes at all stages. It is worth noting that SlABCB21 displayed the highest expression in flower buds, followed by SlABCB14 and SlABCB20. SlABCB20 reached a maximum in the fruit breaker and fruit breaker groups, followed by SlABCB25 (Supplementary Fig. S4).
RNA-Seq data and qRT-PCR validation
-
Seven-day-old S. lycopersicum plants were exposed to 10 μM Cd2+ for 2 hours to measure the expression of SlABCB genes under cadmium stress. The RNA-Seq data were processed to determine the expression levels of the SlABCB genes (Fig. 6). The results indicated that under cadmium stress, SlABCB8, SlABCB11, SlABCB15, SlABCB17, SlABCB22, SlABCB25, SlABCB26, and SlABCB27 were upregulated.
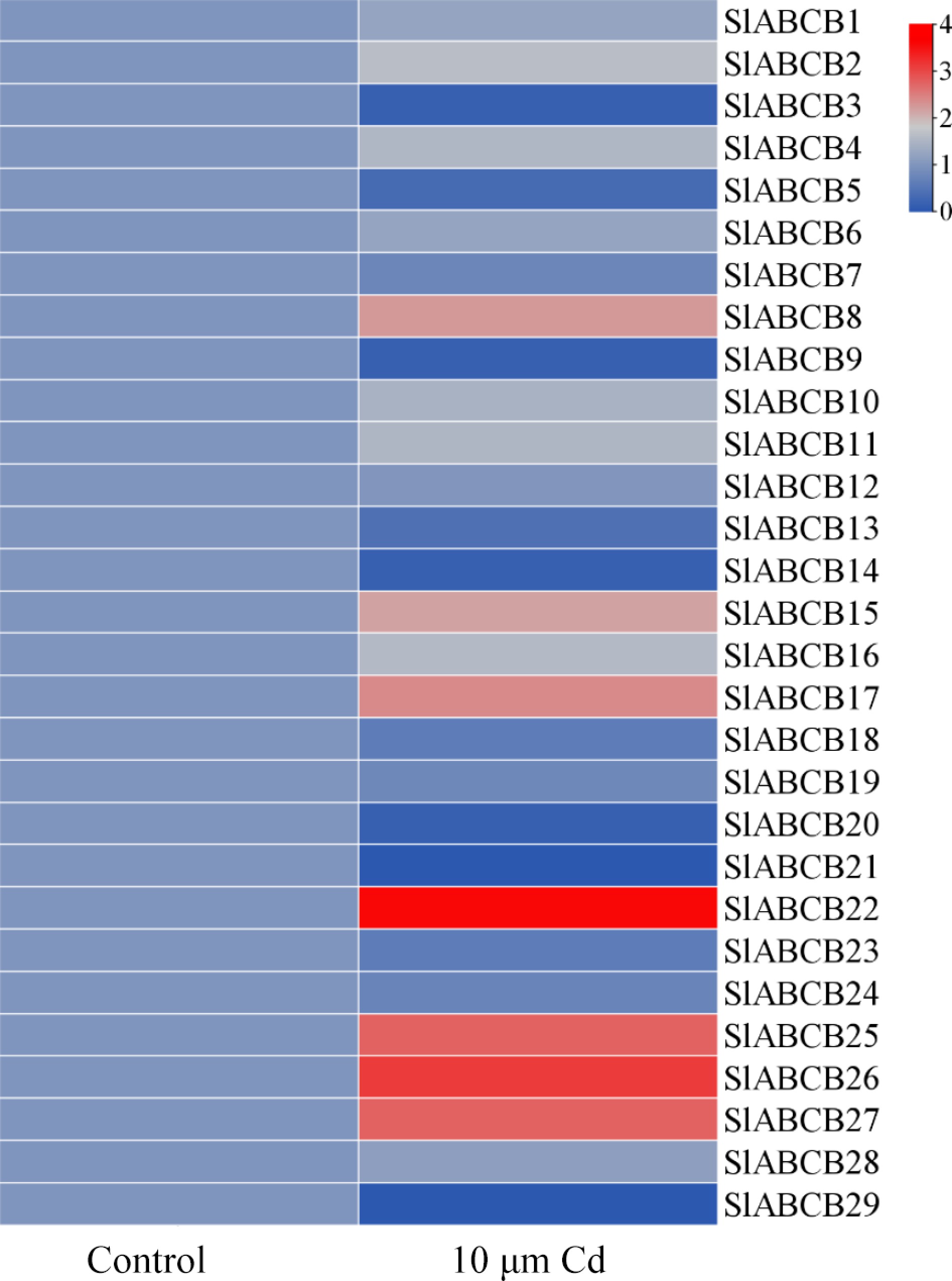
Figure 6.
Heatmap of SlABCB genes determined by microarray analysis and RNA-Seq data. The heatmap was generated using TBtools software using the RNA-Seq data.
Six candidate genes that were upregulated in response to Cd stress were selected for qRT-PCR detection to confirm the accuracy of the ABCB subfamily of genes in the RNA-Seq data. All six genes (ABCB4, ABCB8, ABCB11, ABCB15, ABCB22, and ABCB26) were induced by Cd exposure (Fig. 7). The trend of variations in the expression of the six ABC transporters, as observed by qRT-PCR, was mostly consistent with transcriptome data. These genes might be related to the heavy metal detoxification mechanisms of tomatoes.
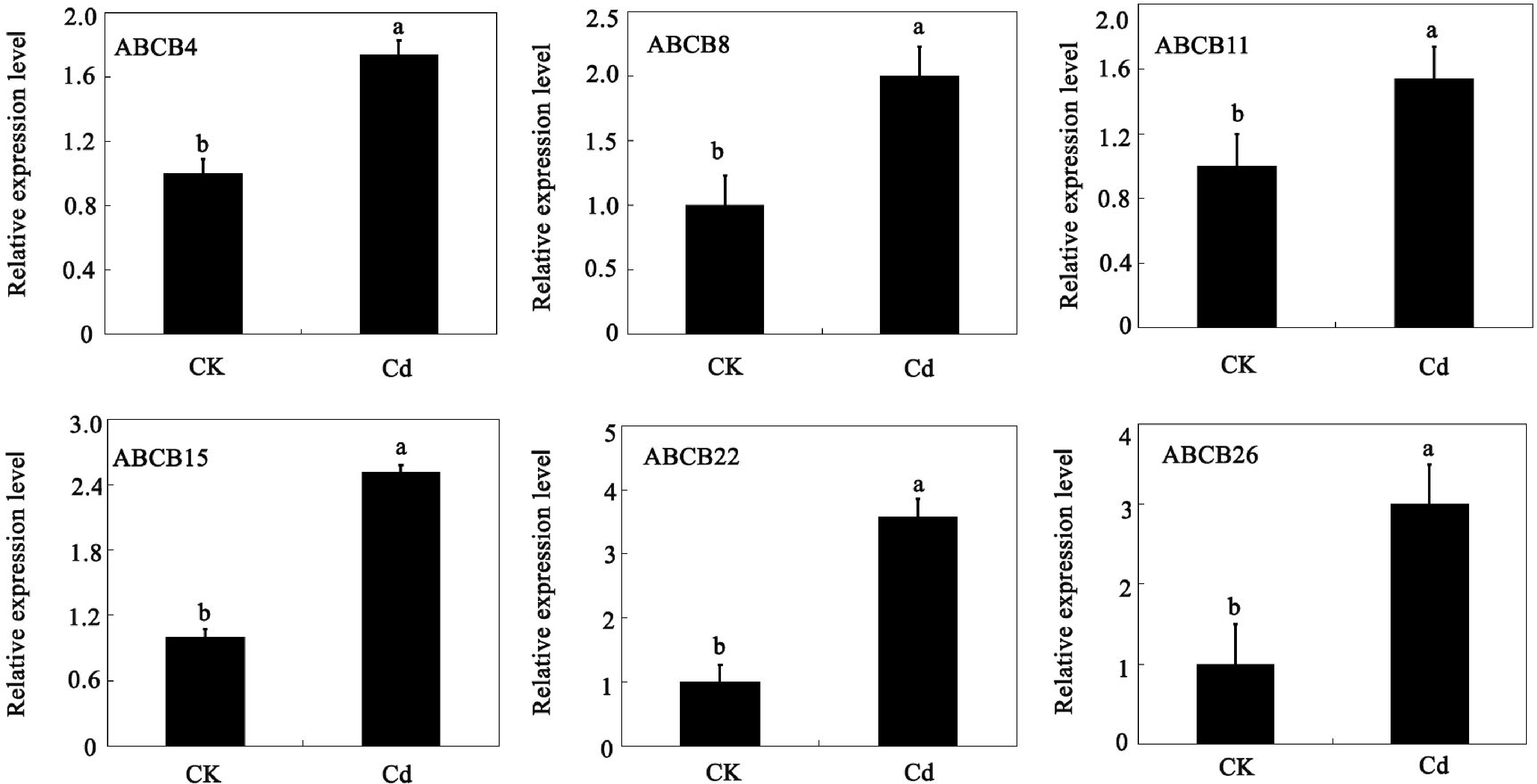
Figure 7.
Expression of ABCB genes in tomato under cadmium stress, determined by qRT-PCR. The data in the figure were obtained from three biological replicates (n = 3), presented as the means + standard error (SE). Significance differences are denoted by different lowercase letters (p < 0.05), according to Duncan's multiple range test.
Several SlABCB genes mediate yeast's tolerance of Cd
-
Family members of SlABCBs with Cd tolerance were induced and screened via expression in yeast. The metal sensitivity assay was conducted using the wild-type yeast strain Y252 and the yeast Cd-sensitive mutant strain Δyap1. The research results demonstrated that the Δyap1 mutant strains carrying the pYES2-SlABCB (SlABCB8 and SlABCB22) plasmids expressed under the control of the GAL1 promoter displayed stronger tolerance to 30 μmol·L−1 CdCl2 compared with the control and those with other SlABCB genes (Fig. 8). In conclusion, in a yeast-based metal sensitivity test assay, the expression of SlABCB8 and SlABCB22 compensated the sensitive phenotypes of the mutant strains grown under excess cadmium.

Figure 8.
Functional verification of SlABCB genes in yeast. SlABCB8, SlABCB15, SlABCB22, and SlABCB26 were transformed into Cd-sensitive strains of Saccharomyces cerevisiae Δyap1 to test their function of mediating tolerance to excess Cd. P/Y, wild-type Y252 transformed with the empty pYES2 vector; P/Δ, the cadmium-sensitive yeast mutant Δyap1 transformed with the empty pYES2 vector.
-
In recent years, ABC transporters have become a major focus for research in plants. In such research,154 ABC transporter proteins in total have been appraised, and their members have been divided into eight groups: ABCA–ABCH[31]. Full-size ABCBs are referred as P-glycoprotein (PGP) or multidrug resistance (MDR) proteins. However, the half-size ABCBs have characteristics labeled with names such as the transporter associated with lipid A-like exporter – putative (LLP), the transporter of antigen processing (TAP), and the ABC transporter of mitochondria (ATM)[3]. In tomato, the ABCB subfamily, with 29 members, is the second-largest subfamily, including 8 half-size, 18 full-size, and 3 quarter-size members[6]. The ABCB subfamily of Arabidopsis contains at least six full-sized ABCBs, encoding auxin transporters[6]. In our research, genes of the ABCB subfamily with 29 members were identified, including 10 half-size, 16 full-size, and 3 quarter-size members (Fig. 2, Table 2), which is highly consistent with the published data.
Several common and unique motifs in the tomato ABC transporters have been identified through MEME motif analysis. Genes with common motifs were grouped into uniform groups, suggesting that they had similar functions in general[5]. In O. sativa, the ABCB subfamily shares four motifs (Motif 1, Motif 2, Motif 4, and Motif 9)[7]. Similar to earlier research, our study revealed that having common motifs often occurs within the same cluster (Fig. 2).
Collinearity analysis revealed a high degree of synteny between tomato and eggplant ABCB genes, with 16 collinear pairs identified, suggesting strong evolutionary conservation and potential functional redundancy within the Solanaceae family[22]. Notably, several of the cadmium-responsive SlABCB genes identified in our study, such as SlABCB8, SlABCB15, SlABCB22, SlABCB25, SlABCB26, and SlABCB27, have putative orthologs in the eggplant genome. For instance, according to the collinearity map, SlABCB22 likely shares a common ancestor with its eggplant counterpart. This high sequence homology implies that these eggplant orthologs may also play roles in heavy metal detoxification, as functional conservation among orthologs across closely related species is frequently observed[6].
An analysis of the member structure of the ABCB subfamily of tomatoes may contribute to functional research. The evolutionary tree revealed that intron–exon arrangements shaped the evolution of this gene family[32,33]. The expression levels of genes with no and few introns are low in plant genomes[34]. Furthermore, the compact gene structure may enable genes to respond rapidly to exogenous or endogenous stimuli in terms of expression[35]. Genes with fewer introns are often associated with rapid transcriptional induction in response to environmental stimuli[35]. This structure allows for quicker mRNA processing and export, which could be crucial for an immediate early response to acute cadmium exposure. Conversely, genes with complex intron–exon architectures (SIABCB20, SIABCB21, and SIABCB25) may be subject to more intricate layers of regulation (Supplementary Fig. S2), including alternative splicing and post-transcriptional modulation[36]. Our genetic structure analysis also indicated that the gene sequences of the ABCB subfamily in tomatoes have the same number of exons and introns with similar functional characteristics, which may have emerged during the evolutionary process of repetitive events.
Duplication, genome size, and gene distribution are the main factors influencing the genetic diversity of land plants. The distribution of ABC transporter genes is not uniform throughout the chromosomes, with chromosome 1 and chromosome 4 hosting the largest proportion of ABC transporter members, accounting for 22.43% and 11.21%, respectively[7]. We discovered that chromosome 2 had four ABCB transporter genes and that chromosome 3 had six ABCB family members (Supplementary Fig. S3). The gene distribution pattern indicated that these genes performed homologous functions in responding to environmental pressure, development, and plant growth[5, 37]. The distribution and mapping of the ABCB subfamily members of tomato at the chromosome level will help tomato breeders develop new tomato varieties with ideal traits.
Studies of cis-elements can provide a necessary foundation for further functional dissection of the ABCB subfamily of genes in tomato. Several hormone-related response elements, including those related to ABA, gibberellin, salicylic acid, MeJA, zein, and auxin responses, were found in the 2,000 bp sequence upstream of the promoter region of the SlABCB genes in the prediction analyses. The link between heavy metal stress and hormone signaling is increasingly recognized as a key mechanism in plant adaptation. Our promoter analysis found that many cadmium-responsive SlABCB genes contain cis-elements for auxin, ABA, jasmonic acid, and salicylic acid (Fig. 2), pointing to likely hormonal regulation. This is particularly relevant for ABCB transporters, many of which are established auxin transporters. Recent work has shown that cadmium stress directly disrupts auxin homeostasis, leading to inhibited root growth[38]. Furthermore, we also found that most of tomato ABCB subfamily promoters contain one or more defense and stress response elements (Fig. 2). The identified Cd-responsive SlABCB genes, including SlABCB8 and SlABCB22, may also contribute to the detoxification of other heavy metals like lead or zinc. Evidence from model plants shows that ABC transporters often confer multi-metal resistance. In Arabidopsis, AtABCB25 provides tolerance to both Cd and Zn[39], and numerous rice OsABC genes are regulated by diverse heavy metals[37]. The upstream regions of these SlABCB genes contain abundant general stress-responsive cis-elements alongside metal-specific ones, suggesting that their expression could be coordinated by broad-acting transcription factors responding to various abiotic stresses. Therefore, to functionally validate the role of these predicted cis-elements in response to cadmium, future studies could employ luciferase reporter assays with progressive promoter deletions and site-directed mutagenesis of key elements.
As a result of exposure to a wide range of environmental stresses, ABC transporters have undergone positive selective pressures during evolution. According to the phylogenetic tree constructed using the ABCB genomes, these genes seem to have evolved from the same ancestor. A few branches exclusively contained homologous tomato or Arabidopsis transporters (Fig. 4). The ABC transporter family members in these gene clusters may perform specific functions in diverse species[39].
Notably, AmABCB1 was highly expressed in the seeds but not in the leaves. Its expression levels were relatively lower in the pericarps and roots[40]. On the 14th day after pollination (DAP), the expression of SlABCB4 reached its peak[6]. In the present study, SlABCB1 showed relatively low expression in leaves and roots, and almost no expression was observed in other tissues and organs. Similarly, the expression level of SlABCB4 was also at a moderate level in young leaves, flowers, and stems, and its expression peaked at the 3-cm fruit stage (Supplementary Fig. S4). These findings were largely consistent with previous research. According to gene expression data based on the eFP Browser, SlABCB7, SlABCB13, SlABCB14, SlABCB18, SlABCB20, SlABCB21, SlABCB24, SlABCB25, and SlABCB29 are widely expressed in all tissues and organs, indicating that they are responsible for basic cell maintenance[6].
Microarray and RNA-Seq data from an earlier study demonstrated that ABC transporter family members are upregulated in response to environmental stress[7]. Following heavy metal (Cd) stress treatment, rice OsABCC9 was upregulated and entered the root vacuoles by chelating Cd, mediating tolerance to and accumulation of Cd[41]. AtABCB25 participates in the biosynthesis of molybdenum cofactors and heavy metal tolerance, probably through its function as a glutathione disulfide (GSSG) transporter[42]. The transcriptional level of ABCG36 in poplar was upregulated under Cd stress. Overexpression of ABCG36 in transgenic Arabidopsis enhanced plants' resistance to Cd stress by expelling it from the plants[43]. In this study, SlABCB22, SlABCB26, and SlABCB27 were identified as potential cadmium-resistant candidate materials, and were worthy of further study (Fig. 6). Whereas our transcriptome and qRT-PCR analyses identified key SlABCB genes that were responsive to cadmium stress at the whole-seedling level, the current study did not separately analyze root versus leaf tissues in detail. Investigating this tissue-specific regulation represents a crucial direction for our future research.
Among the four Cd-tolerant SIABCB genes identified in the yeast assay, SIABCB8 and SIABCB22 conferred a more pronounced tolerance phenotype compared with SIABCB15 and SIABCB26 (Fig. 8). This functional divergence among subfamily members, despite their structural similarities, is a common phenomenon in ABC transporters[9]. Primarily, substrate specificity and transport efficiency are largely determined by the composition of the transmembrane domains (TMDs)[9]. It is plausible that subtle differences in the TMD sequences of SIABCB8 and SIABCB22 confer higher binding affinity or transport kinetics for cadmium ions or their complexes compared with SIABCB15 and SIABCB26. This is supported by studies in Arabidopsis, where different ABC transporters exhibit distinct roles in cadmium detoxification; for instance, AtABCC3 is specifically involved in sequestering Cd–phytochelatin complexes into the vacuoles[44].
The evolutionary history of these transporters across the Solanaceae family warrants further investigation. The recent availability of genomic data for multiple species enables comparative analyses with taxa such as Capsicum annuum (bell pepper) and Solamum tuberosum (potato)[45]. This study provides a foundational analysis of the ABCB subfamily within Solanum lycopersicum. The research offers a very useful reference for future research on the functional divergence and evolution of the ABCB subfamily of genes, especially cadmium transport-related genes.
In conclusion, analyses of gene structure, chromosomal locations, gene duplication, comparative phylogenetics, and cis-regulatory elements were conducted on 29 identified tomato ABCB subfamily genes. Meanwhile, the research offered proof for the biological functions of ABCB subfamily members as transporters in regulating plants' Cd resistance. In addition, it is hoped that this study will have certain reference value for further research on ABC transporter proteins in plants.
This work was funded by the Hunan Provincial Department of Education Key Project (23A0349), Hunan Province College Students' Innovation Training Program (S202310534072), the Hunan Provincial Natural Science Foundation of China (2025JJ70108), and the Graduate Scientific Research Innovation Project of Hunan Province (CX20231038).
-
The authors confirm contributions to the paper as follows: study design: He G, Liang Q, Yang P, Song G; writing the manuscript: Zhang J; reviewing the manuscript: Gao Y. All the authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Fig. S1 Phylogenetic analysis of ABCs gene family members among tomato.
- Supplementary Fig. S2 Exon-intron structure analysis of ABCBs gene family.
- Supplementary Fig. S3 The location of ABCBs gene on the peach chromosome. Chromosomes 2-12 are shown in long bars with yellow font. The approximate distribution of each ABCBs gene is marked in red font on a long bar.
- Supplementary Fig. S4 Expression profiles of SlABCB genes in the root, leaf, flower, fruit and fruit breaker of S. lycopersicum. Data were retrieved from TOM express (http://tomexpress.toulouse.inra.fr/).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yang P, He G, Liang Q, Song G, Li Y, et al. 2025. Genome-wide analysis of the ATP-binding cassette B subfamily in tomato and its response to cadmium stress. Vegetable Research 5: e049 doi: 10.48130/vegres-0025-0042 

Genome-wide analysis of the ATP-binding cassette B subfamily in tomato and its response to cadmium stress
- Received: 12 June 2025
- Revised: 12 October 2025
- Accepted: 15 October 2025
- Published online: 22 December 2025
Abstract: The ATP binding cassette transporter (ABC) superfamilies consist of many membrane proteins and play a very important role in transmembrane transport of substances such as secondary metabolites, phytohormones, and heavy metals. However, it is still unknown what role the B subfamily of the ABC family (ABCB) plays in tomatoes. In the present study, we conducted a genome-wide analysis of the ABCB subfamily in tomato (Solanum lycopersicum), which is the largest group in the ABC family with 29 members, followed by the C subfamily. The predicted ABCB genes were distributed across 10 chromosomes, with up to six SlABCB genes emerging on chromosome 3 and one SlABCB gene detected on chromosome 4. The quantity and structure of exons and introns varied by gene. Analysis of the transcriptome data revealed increased transcript levels of ABCB subfamily genes following cadmium stress. Similarly, these results of quantitative real-time polymerase chain reaction (qRT-PCR) were also confirmed. A yeast experiment showed that the genes SlABCB8, SlABCB15, SlABCB22, and SlABCB26 can mediate Cd tolerance. Overall, our research offers new insight into the molecular function of ABCB subfamily transporter genes in S. lycopersicum, which is highly valuable and important because of the effect of long-term application of ABCB transporter genes on regulation under heavy metal stress.
-
Key words:
- Cadmium stress /
- Bioinformatics /
- Tomato /
- ABC transporter protein /
- Phylogenetic analysis /
- qRT-PCR