









-
Watermelon (Citrullus lanatus) is one of the important horticultural crops in the genus Citrullus of the family Cucurbitaceae. Its fruit is rich in nutrients that are necessary for maintaining human health, such as carotenoids, vitamins, glucose, amino acids, and so on[1]. Among them, carotenoids and their derivatives have important roles in preventing cancer, reducing the risk of cardiovascular disease, alleviating eye diseases, and regulating the immune system[2].
In higher plants, carotenoids are crucial for photosynthesis, serve as precursors for ABA biosynthesis, and are the main dietary source of vitamin A precursors in human diets[3]. More than 750 carotenoids have been identified and are widely distributed in colorful flowers, fruits, leaves, and roots[4]. Carotenoids such as β-carotene[3], lycopene[2], and lutein[5] play important roles in the food and pharmaceutical industries because of their antioxidant activity. Carotenoids are important nutrients and color-presenting substances in the flesh of watermelon[6]. The regulatory mechanism is still not completely clear; therefore, further exploration will be helpful to comprehensively understand carotenoid metabolism and improve the fruit nutrition in watermelon.
Even though the metabolic pathway of carotenoids has been explored in many important plants[7], the metabolic process is still very complicated. Under the interaction of genotype and environment, it has been noticed that the carotenoid metabolic process is mainly regulated by transcriptional level[8], transcription factor[9], post-transcriptional level[10,11], post-translational level[12,13], and epigenetic modification[14,15]. Post-transcriptional regulation plays an essential role in regulating the levels and activities of enzymes related to carotenoid synthesis in plants, and especially during fruit development and ripening, such as the direct interaction of SlSGR1 protein with SlPSY1 protein in tomato, which regulates carotenoid accumulation in fruits[10]. Nudix hydrolase 23 (NUDX23), a Nudix domain-containing protein, regulates carotenoid biosynthesis by directly interacting with PSY and GGPPS in chloroplasts, thereby enhancing the stability of PSY and GGPPS proteins in the large PSY-GGPPS enzyme complex[16]. A CmOr nonsense mutation (Cmor-lowβ) was identified in the EMS-induced low-β mutant in melon, which significantly reduced PSY protein levels and enzyme activity, leading to a decrease in carotenoid metabolic flux and accumulation[17]. The interaction between CmOR family proteins and light-harvesting chlorophyll a-b binding proteins pointed to a previously unrecognized role of CmOR in maintaining chloroplast function in melon fruit[13]. CmOR protein interacts with the carotenoid sequestration protein CmFBN1 protein, which stimulates plastoglobule proliferation and promotes the accumulation of β-carotene in the chloroplasts of melon fruit[18]. PSY, serving as the rate-limiting enzyme in catalyzing carotenoid metabolism, influenced the content of carotenoids[7]. According to previous reports, the expression of ClPSY1 in the red-fleshed watermelon was significantly higher than that in other flesh-colored watermelons, and it regulated carotenoid accumulation and chromoplast differentiation[19].
The Or gene, as one of the important genes that can regulate the differentiation of chromoplasts, was a significant factor in regulating the metabolism and accumulation of carotenoids in plants[20,21]. The Or gene was initially found in cauliflower mutants, and mutations of the Or gene led to the accumulation of large quantities of β-carotene in the weakly or unpigmented tissues, such as floral bulbs, the pith, the base of the leaves, the phloem, and the bud meristems, which turned into an orange color[22]. The Or gene encodes a class of DnaJ-like zinc finger protein-containing structural domains whose function is related to triggering the formation of chromoplasts from proplastids or other noncolored plastids to accumulate carotenoids[20]. Except in melon, ectopic expression of the alfalfa (Medicago sativa L.) Or gene also increased carotenoid accumulation and plant tolerance to drought stress[23]. Furthermore, AtOr (At5g61670) drove the conversion of proplastids or other noncolored plastids into chromoplasts, thereby enhancing carotenoid storage[24]. High expression of AtOrWT and AtOrHis in tomato promoted overall early flowering and more fruit and seed number in tomato[25]. Overexpression of the Or gene in sweet potato (Ipomoea batatas [L.] Lam) promoted carotenoid accumulation and abiotic stress tolerance in storage roots[26]. The OR homolog BrGOLDEN likely enhanced carotenoid accumulation by modulating the expression of key carotenoid biosynthesis genes, including ZEP, DXS, CYP97C1, and PSY in Brassica rapa[27]. Overexpression of either potato StOR or StORHis regulated carotenoid homeostasis and improved plant adaptation to environmental stress[28]. An integration analysis of carotenoids and transcriptome during the development of five different flesh-colored watermelon fruits indicated that the Cla018406 gene (a chaperone protein dnaJ-like protein) may be a candidate gene for β-carotene accumulation in a previous study, Cla018406 was homologous to the BoOr[20] and CmOr[21], and its gene expression pattern was related to the β-carotene content abundant in orange flesh[6]. However, the Or gene function in regulating carotenoid synthesis and accumulation during fruit development and ripening is still unknown and needs further verification in watermelon.
Herein, this research aimed to decipher the Or gene family and functionally verify the key gene regulating carotenoid synthesis and accumulation during fruit development and ripening in watermelon. The carotenoid metabolism analysis, gene expression, genetic transformation, virus-induced gene silencing (VIGS), yeast two-hybrid assay (Y2H), and bimolecular fluorescent complimentary assay (BiFC) were used to determine the gene function and molecular mechanism of ClOr. The retrieved results will provide an insight into the verification of the ClOr gene that regulates carotenoid synthesis and accumulation during development and ripening in watermelon fruits, with the hope of promoting the genetic improvement of watermelon fruit flesh based on the ClOr gene in the future.
-
The two watermelon varieties (LSW-177 [Citrullus lanatus] and W1-60 [Citrullus amarus]) were selected as experimental materials. The mature flesh of the LSW-177 watermelon was red, while the mature flesh of the W1-60 watermelon was white. They were cultivated at the Horticultural Teaching and Experimental Base of Jiangxi Agricultural University. All plants were grown under standardized field conditions with uniform agronomic practices.
The tobacco (Nicotiana benthamiana) was used for subcellular localization assays; its seedlings were grown in autoclaved potting mix within a well-protected climate-controlled condition of greenhouse (24/20 °C day/night cycle, 16 h photoperiod supplemented with artificial lighting) for 30 d post-germination to inject.
The seeds of tomato (Solanum lycopersicum L. cv. Micro-Tom) were obtained from Wuhan Boyuan, China. After complete expansion of the cotyledons, they were excised and used for Agrobacterium-mediated transformation. The seeds of LSW-177 for VIGS were cultured under controlled greenhouse conditions prior to infiltration.
Carotenoid extraction and quantification
-
The fruit flesh samples were collected from five developmental stages at days after pollination (DAP) of red- and white-fleshed watermelons (e.g., 10, 18, 26, 34, and 42 DAP) and freeze-dried. Then, samples were finely ground using a freeze-mixer grinder (30 Hz, 1 min); 50 mg of fruit flesh powder was sampled, and 0.5 mL of a hexane/acetone/ethanol mixture (1:1:1) containing 0.01% BHT (Butylated Hydroxytoluene, g/mL) was added in equal volume ratios, which was fully extracted by vortexing at 2,800 rpm in a vortex mixer (30 s) and aided with an ultrasonic cleaner. The carotenoids were fully extracted by vortex mixer at 2,800 rpm (30 s) and ultrasonic cleaner-assisted dissolution (40 kHz, 20 min); the obtained extract was centrifuged for 5 min (12,000 rpm, 4 °C) to obtain the supernatant, which was repeated once and combined; the supernatant was concentrated by vacuum centrifugal concentrator (1,400 rpm, 6 h), then reconstituted with 100 µL of an equal-volume ratio of methanol/methyl tert-butyl ether, and filtered by a 0.22 µm filter membrane. The solution was filtered through a 0.22 µm membrane into a brown vial and analyzed by Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS) to detect the compositions and contents of carotenoids in the flesh.
The flesh samples of watermelon and tomato were finely ground using a freeze-mixer grinder (30 Hz, 1 min); carotenoid measurements were done by following the method described by Rodriguez et al.[29]. In brief, 4 mL of 95% ethanol (FW, 0.2 g) was extracted under light for 20 min at 4 °C, and at 12,000 rpm for 5 min to obtain the supernatant; the extraction was repeated for the precipitate; the supernatant was mixed and concentrated using a vacuum centrifugal concentrator and re-dissolved with 1 mL of 95% ethanol; and the absorbance was measured using a microplate reader at 665, 649, and 470 nm. The formula for carotenoid content was calculated using the method described by Arnon[30].
Microscopic observation of watermelon flesh
-
The freshly harvested fruits were brought back to the laboratory, and the longitudinal sections of the fruits were photographed to record the fruit development process; 2–3 cm2 of the center flesh was taken and submerged in FAA fixative (Formaldehyde-acetic acid-ethanol fixative, containing 38% formaldehyde 5 mL, acetic acid 5 mL, 70% ethanol 90 mL, and 5 mL of glycerol) for 1 h, and then the flesh was transferred to 0.1 M Na2-EDTA solution (pH = 9) in a 60 °C hot water bath for 150 min to promote intercellular segregation. After ending the flesh treatment, a small amount of flesh tissue was scraped with a razor blade to observe the chromoplast in the flesh cells under the optical microscope. The submicroscopic structure in the flesh cells was observed using a transmission electron microscope (TEM) according to a previous study[19].
Identification of the Or gene family in watermelon
-
The protein database (v2.5) of watermelon genome (97103) from the CuGenDBv2 (
http://cucurbitgenomics.org/v2 ) was selected as the subject database, the amino acid sequence of the AtOr gene (At5g61670) was obtained from NCBI (www.ncbi.nlm.nih.gov ), the amino acid sequence of the AtOr gene was blasted, and the homolog of the Or gene family members of watermelon were obtained (E-value < 1e-10, Identity > 60%). Then, the nucleotide sequence and amino acid sequences of the ClOr were obtained from CuGenDBv2, and their gene structures were analyzed using CSDS (v2.0) (http://gsds.gao-lab.org/ ).For multiple sequence alignment and phylogenetic tree analysis, the amino acid sequences of the Or proteins of 14 other species were downloaded from the NCBI database, and their accessions are listed in Supplementary Table S1. Multiple sequence alignment of the 16 Or proteins was performed by MEGA software (v11), and the results were demonstrated using DNAMAN software (v9.0). The phylogenetic tree was constructed by the ML method (Maximum likelihood) with 1,000 bootsteps and visualized using Chiplot (
www.chiplot.online/ ).Gene cloning and dCAPS molecular marker development
-
Total RNA was extracted from 34 DAP flesh samples of red- and white-fleshed watermelon using the MolPure® Plant RNA Kit (Yeasen, Shanghai, China). The purified total RNA was reverse-transcribed into cDNA using the PrimeScriptTM RT reagent Kit with gDNA Eraser (Takara, Shiga, Japan), and amplification of full-length ClOr1 cDNA with gene-specific primers was performed by polymerase chain reaction (PCR). All the PCR products were then inserted into pClone007 Versatile Simple Vector (Tsingle, Beijing, China) and transformed into TreliefTM 5α Chemically Competent Cell (Tsingle, Beijing, China). In addition, a dCAPS molecular marker based on the SNP in ClOr1 was developed to distinguish between white-fleshed and non-white-fleshed watermelons. The total DNA was extracted from 34 DAP leaves of red- and white-fleshed watermelon using the MolPure® Plant Plus DNA Kit (Yeasen, Shanghai, China). For molecular genotyping, the PCR amplifications were performed using 2 × Hieff Canace® Plus PCR Master Mix (Yeasen, Shanghai, China), then the PCR products were digested with suitable restriction enzymes (Hpy166II, NEBiolabs, Beijing, China), and recognizing dCAPS sites. The information of developed primers is shown in Supplementary Table S2. The web server program SOPMA was used to analyze the secondary structure of proteins predicted by ClOr1Trp and ClOr1Cys, and the Swiss Model was used for online prediction of their tertiary structure.
Subcellular location
-
To confirm where ClOr1 is expressed in cells. Homologous recombination and the Golden Gate seamless cloning method (Bsa I/Eco 31I) were used to construct the pBWA(V)HS-Osgfp-ClOr1 fusion vector. The primers used for the fusion vector are listed in Supplementary Table S2, together with the cytosolic Marker plasmid to transform the GV3101 Competent cell, respectively. Successfully transformed Agrobacterium was cultured in the bacterial fluid OD600 = 0.6, centrifuged at 5,000 rpm (4 min) to obtain the bacterium and resuspended, and injected pBWA(V)HS-Osgfp-ClOr1 into the tobacco leaves by mixing with the Agrobacterium plasmid in the ratio of 1:1 with the cytosolic Marker plasmid. Two days later, Fluorescence images of GFP were observed under a confocal microscope and photographed for documentation.
Genetic transformation of ClOr1 in tomato
-
The pBWA(V)KS-ClOr1-Osgfp fusion vector was constructed by the homologous recombination and Golden Gate seamless cloning method using restriction endonuclease (Bsa I/Eco 31I). The primers used for vector construction are shown in Supplementary Table S2. Subsequently, the fusion vector was converted to the Agrobacterium GV3101 strain using the freeze-thaw method. For tomato transformation, tomato cotyledons were infiltrated with Agrobacterium resuspension with OD600 = 0.2 for 10 min, and kanamycin resistance was used to screen the healing wounds. The screened healing wounds were inoculated into the differentiation medium for 30 d. After the differentiated seedlings grew to about 2–3 cm, they were cut off from the healing wounds and inoculated into the rooting medium for 10 d. After the development of a proper root system, the seedlings were transferred and cultivated in sterilized substrate (24/20 °C, 16 h light/8 h dark). The insertion of ClOr1 in the OE tomatoes was verified using the MolPure® Plant Plus DNA Kit (Yeasen, Shanghai, China).
VIGS assay for silencing the ClOr1 gene
-
To obtain a specific sequence of the ClOr1 gene for VIGS, SsClOr1 was designed using an online tool (
https://vigs.solgenomics.net ) and inserted into plasmid pTRV2. Gene-specific primers containing restriction sites (Xba I and BamH I) were designed for amplification (Supplementary Table S2). The recombinant vector and empty vector were transformed into GV3101, respectively. According to the method reported in a previous study[31], cultivate Agrobacterium GV3101 to an OD600 of 1.0, then thoroughly mix the pTRV1 with the pTRV2 and pTRV2-SsClOr1 in a 1:1 ratio for the control group and experimental group, respectively. After incubating in the dark for three hours, select three fruits (18 DAP) from the potted watermelons of the LSW-177 in the greenhouse. A total of 1 mL of the mixed infestation solution of the control group and experimental group was injected into two opposite sides of the middle fruit using a 1 mL sterile syringe. Infection was repeated on the third and sixth days after the first injection, and fruit flesh tissue near the needle insertion site was collected 3 d after the third injection.Validation of interaction between ClOr1 and ClPSY1
-
The Y2H and BiFC assays were used to validate the interaction between ClOr1 and ClPSY1. For the Y2H assay, the full-length ClOr1 CDS was fused into the pGBKT7 vector (BD), forming the BD-ClOr1 fusion vector, whereas the full-length ClPSY1 (Cla97C01G008760) CDS was fused into the pGADT7 vector (AD), forming the AD-ClPSY1 fusion vector. Five combinations were set up in the following way: BD-ClOr1 + AD-ClPSY1, BD-ClOr1 + AD, BD + AD-ClPSY1, BD-53 + AD-T, and BD-Lam + AD-T. The five combinations were transformed into yeast strain AH109 (Coolaber, Beijing, China). Each combination was cultured on SD/-Leu/-Trp (SD/-LW) medium at 29 °C for 2 d, which was then transferred and streaked onto SD/-Leu/-Trp/-Ade/-His (SD/-LWAH) medium containing 20 mg/L X-α-gal at 29 °C for 2–3 d to test for protein interactions. The primers used are shown in Supplementary Table S2.
For the BiFC assay, ClOr1 CDS was constructed into the vector pCAMBIA1300-35S-YC155 (YC), and the ClPSY1 CDS was constructed into the pCAMBIA1300-35S-NY173 (NY) vector, respectively. Then, Agrobacterium strain (GV3101) carrying the binary constructs was transformed together into leaves of five-week-old plants of N. benthamiana. After 72 h, the fluorescence was observed under a confocal microscope as described previously[32]. The information of the primers used is shown in Supplementary Table S2.
RNA extraction and gene expression analysis
-
Total RNA was extracted using the MolPure® Plant RNA Kit (Yeasen, Shanghai, China). Total RNA was reverse transcribed into cDNA using the PrimeScriptTM RT reagent Kit with gDNA Eraser (Takara, Shiga, Japan). To explore the expression patterns of the ClOr1, ClOr2, and ClPSY1 genes in the various tissues and flesh of red-fleshed and white-fleshed watermelons at different developmental stages, qRT-PCR analysis was conducted using the Hieff® qPCR SYBR® Green Master Mix from Yeasen (Shanghai, China), and relative expression of the genes was determined by the 2−ΔΔCᴛ method using a relative quantitative method. There were three biological replicates for each tissue and three mechanical replicates for each biological replicate. Gene-specific primers are listed in Supplementary Table S2, and the CIYLS8 gene was used as the internal reference gene[33]. For transcriptomic analysis, RNA-Seq data of the W1-60 and LSW-177 (SRX2037303) were utilized to further analyze the expression patterns of ClOr1 and ClOr2 genes, respectively. All experimental data were statistically analyzed using SPSS 22 and Origin 2024.
-
After analysis of carotenoid metabolism, a total of 17 carotenoids were detected in two watermelons, such as lycopene, (E/Z)-phytoene, β-carotene, and lutein, and the standard curves of MRM ion-pair information and relative carotenoid content are shown in Supplementary Tables S3, S4, respectively. In the results of the principal component analysis of carotenoids (Fig. 1a), the sum of PC1 and PC2 was 87.13%. The cluster analysis revealed that the 17 types of carotenoid contents varied among watermelons of different flesh colors. In red-fleshed watermelon of the LSW-177, the content of violaxanthin decreased as the fruit developed, while the content of lycopene, β-carotene, β-cryptoxanthin, γ-carotene, α-carotene, lutein, and others generally increased during fruit development. Moreover, lycopene accumulated rapidly at the early developmental stage of red-fleshed watermelon, and its content accounted for 84.8% of the total carotenoid content, and at the ripening period, lycopene, (E/Z)-phytoene, and β-carotene were the major carotenoid constituents, and the sum of the three contents accounted for 94.2% of the total carotenoids (Fig. 1b, c, Supplemental Table S5). In white-fleshed watermelon of W1-60, the content of violaxanthin, neoxanthin, capsorubin, zeaxanthin, and lutein decreased with fruit development, while the content of canthaxanthin increased with fruit development. During the development of white-fleshed watermelon, lutein was the main carotenoid component. The lutein content of 18 and 34 DAP accounted for 76.6% and 67.5% of the total carotenoids, respectively (Fig. 1b, c, Supplemental Table S5).
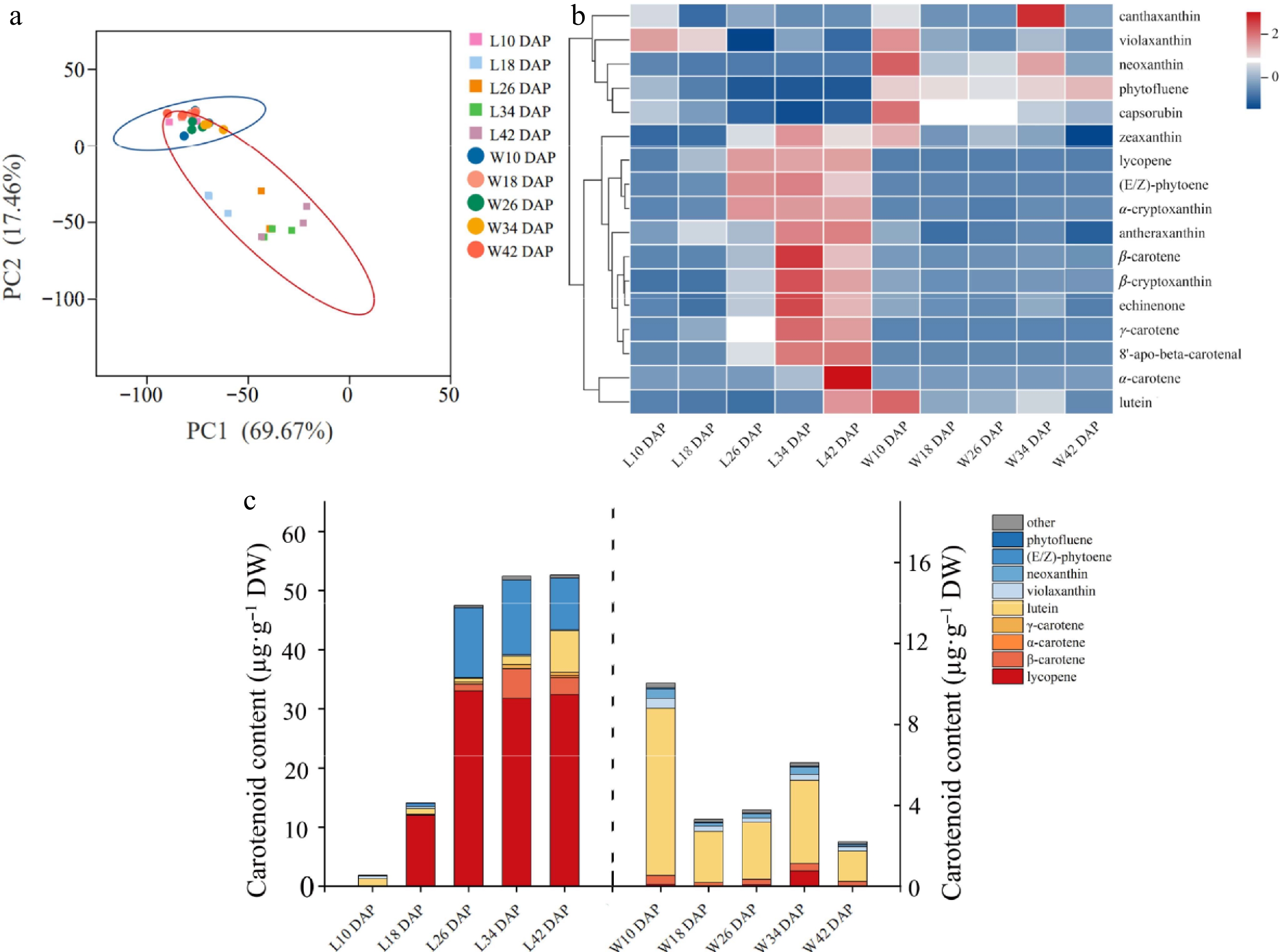
Figure 1.
Carotenoid accumulation and developmental patterns of chromoplast in red- and white-fleshed watermelons. (a) Main component analysis of carotene. The cube with five colors indicates the flesh of LSW-177 at 10, 18, 26, 34, and 42 d after pollination, covered by a red oval. The five colored circles indicate the flesh of W1-60 at 10, 18, 26, 34, and 42 d after pollination, covered by a blue oval. The data was processed log2 (N). (b) Cluster analysis of carotene: the data have been standardized by Z-score processing. (c) Carotenoid composition analysis: the content of carotenoid in fruit of L10, L18, L26, L34, and L42 DAP is shown on the left Y axis, and the content of carotenoid in fruit of W10, W18, W26, W34, and W42 DAP is shown on the right Y axis.
Different patterns were found in the microscopic and submicroscopic structures of the LSW-177 and W1-60 watermelon fruits after observation. There was no visible pigment accumulation in the longitudinal section of 10 DAP fruits of two watermelons (Fig. 2a1, d1), and no chromoplast was in the flesh cells (Fig. 2b1, e1), but the red-fleshed watermelon contained more proplastid (Fig. 2c1, f1). In the longitudinal section of 18 DAP fruits of red-fleshed watermelon, carotenoids were deposited around the seeds (Fig. 2a2), globular chromoplasts appeared in the cells, and some chromoplasts formed endogenous plastoglobules (Fig. 2b2, c2), while the white-fleshed watermelon 18 DAP flesh was still white (Fig. 2d2, e2, f2). The 26 DAP flesh of red-fleshed watermelon was uniformly colored (Fig. 2a3) with lots of red carotenoid crystals and more plastoglobules (Fig. 2b3, c3), while the 26 DAP flesh of white-fleshed watermelon had globular chromoplasts appearing in the cells with fewer plastoglobules (Fig. 2d3, e3, f3). From 34 to 42 DAP in red-fleshed watermelon, flesh color gradually continued to deep red, more carotenoid crystals were accumulated in the cells, the membrane structure of the chromoplast gradually disappeared, and plastoglobules were dispersed into the cytoplasm (Fig. 2a4, a5, b4, b5, c4, c5). In white-fleshed watermelon, there was a pale yellow pigmentation around the seeds, and the number of chromoplasts and plastoglobules increased (Fig. 2d4, d5, e4, e5, f4, f5).
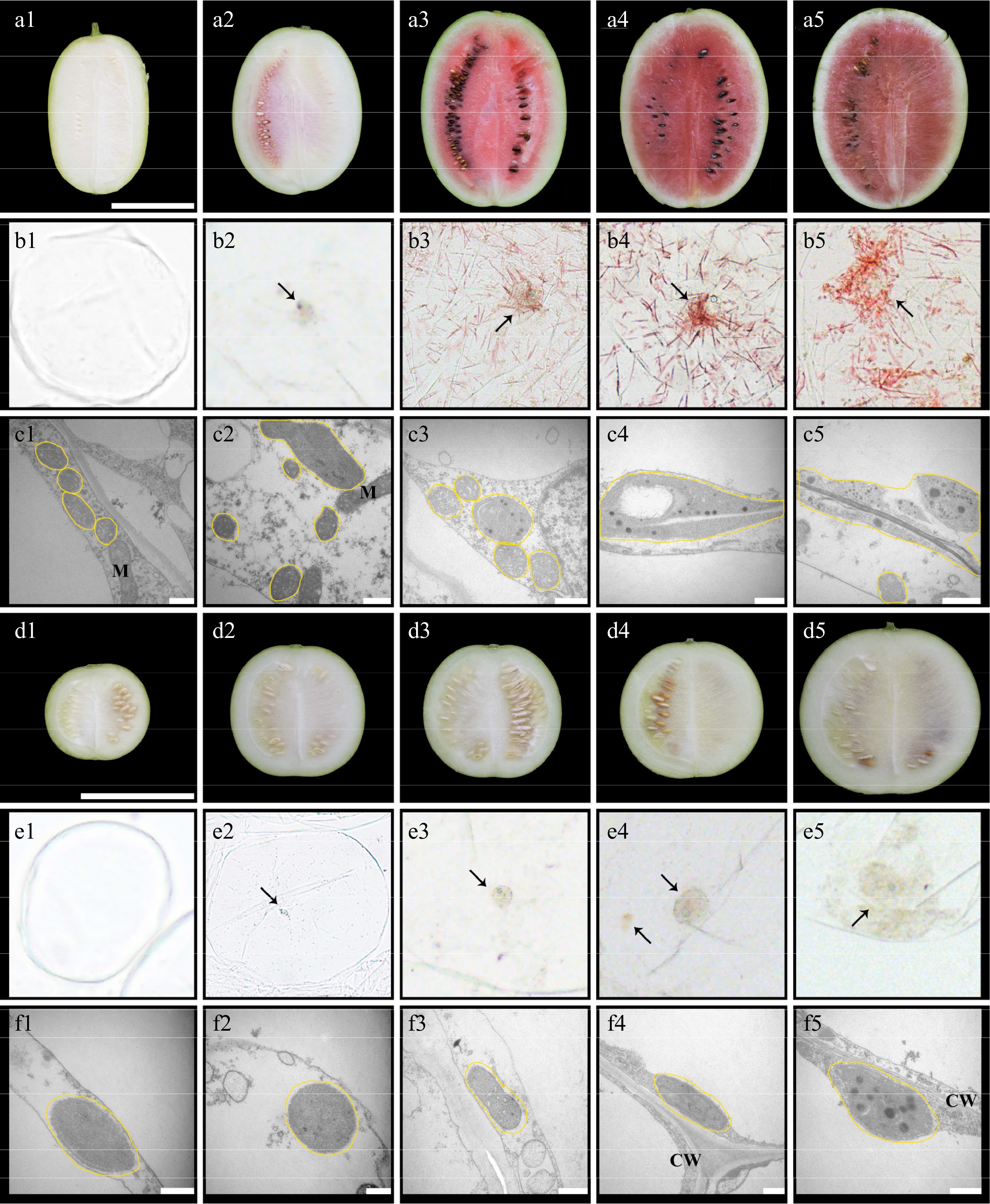
Figure 2.
Cytological observation of chromoplast in the flesh of watermelons (LSW-177 and W1-60). The letters (a)–(c) are the longitudinal section, cell microstructure, and cell sub-microstructure of the fruit of LSW-177, respectively; the letters (d)–(f) are the longitudinal section, cell microstructure, and cell sub-microstructure of the fruit of W1-60, respectively. The scale of (a) and (d) is 10 cm, and the scale of (c) and (f) is 0.5 µm. The numbers (1), (2), (3), (4), and (5) after the letters represent 10, 18, 26, 34, and 42 DAP, respectively. The black arrow points to the chromoplast, the yellow line represents the plast. M: mitochondria; CW: cell wall.
Identification of key Or genes regulating carotenoid accumulation in watermelon fruit
-
To identify the key Or genes regulating carotenoid accumulation in watermelon fruit, the amino acid sequence of the AtOr gene in Arabidopsis was used as the reference sequence for pairwise comparison and screening in the protein database of watermelon genome (97103), and the Cla97C01G005050 and Cla97C04G076970 genes were identified as members of the Or gene family in watermelon genome and named the ClOr1 and ClOr2 genes, respectively. The ClOr1 gene is located on chromosome 1, with a total length of 6,621 bp, and the ClOr2 gene is located on chromosome 4, with a total length of 12,579 bp (Supplementary Table S6). The structures of the ClOr1 and ClOr2 genes analyzed by the CSDS (v2.0) (Fig. 3a), the ClOr1 gene contains eight exons and seven introns, and the ClOr2 gene contains 16 exons and 15 introns; their full-length CDS were 978 bp and 4,230 bp, respectively. The protein sequences of ClOr1 and ClOr2 harbor the cysteine-rich DnaJ and HSP40 zinc-finger structural domains (two CxxCxGxG motifs and two CxxCxxxG motifs) in the multiple sequence comparison and conserved structural domain analysis (Fig. 3b). In the phylogenetic tree (Fig. 3c), the Or genes were mainly divided into two clusters. the ClOr1 was closest to bitter gourd (XP_022155475.1), cucumber (XP_004143670.1), winter melon (XP_038876057.1), and melon (XP_008467325.1). The ClOr2 was clustered into a clade with carrot (XP_017243074.1) and sweet orange (XP_006481741.1).
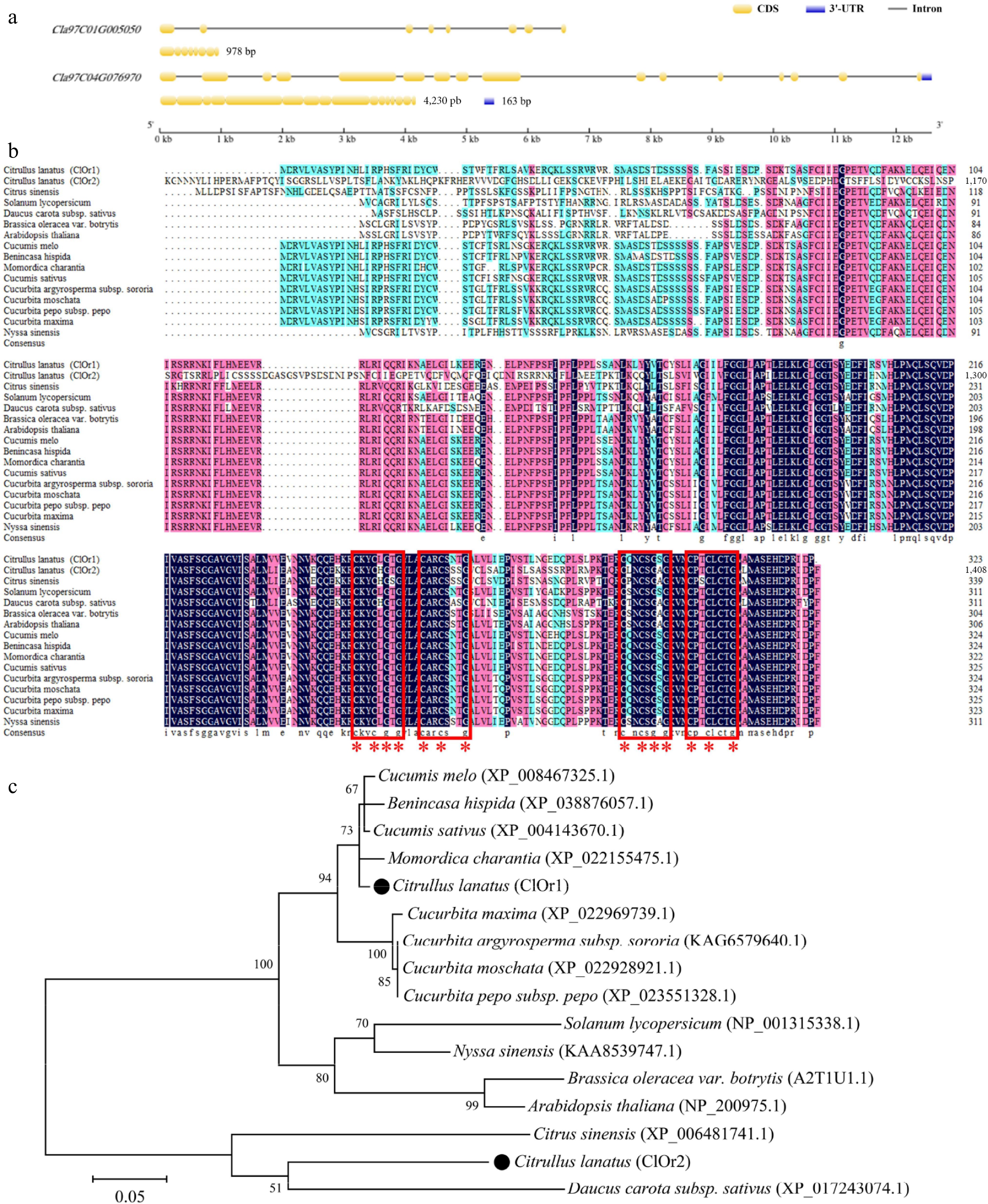
Figure 3.
Sequence alignment and phylogenetic analysis of ClOr. (a) Structure of the ClOr1 and ClOr2 genes. (b) Sequence analysis of the Or protein. The red box marks two (CxxCxGxG) motifs and two (CxxCxxxG) motifs, and the red '*' marks eight cysteine and six glycine residues that are highly conserved. (c) Phylogenetic tree of the Or protein. The bar indicates an evolutionary distance of 0.05%.
The expression levels of the ClOr1 and ClOr2 genes were increasing in the flesh of red- and white-fleshed watermelons during fruit development (Fig. 4a), with expression peaking at 34 DAP. Moreover, the expression level of ClOr1 in the flesh of red-fleshed watermelon was higher than that in white-fleshed watermelon at the same developmental stage. However, there was no significant difference in the expression level of ClOr2 between the flesh of red- and white-fleshed watermelons. Furthermore, the expression profiles of the ClOr1 and ClOr2 genes were examined in various tissues of red- and white-fleshed watermelons at 34 DAP. The ClOr1 gene was most highly expressed in the flesh of red-fleshed watermelon and the leaves of white-fleshed watermelon; however, the ClOr2 gene exhibited the highest expression in the female flower of red-fleshed watermelon and in the male flower of white-fleshed watermelon. Notably, compared to ClOr2, ClOr1 could play a more important role in the red flesh of watermelon based on the above expression level analysis.
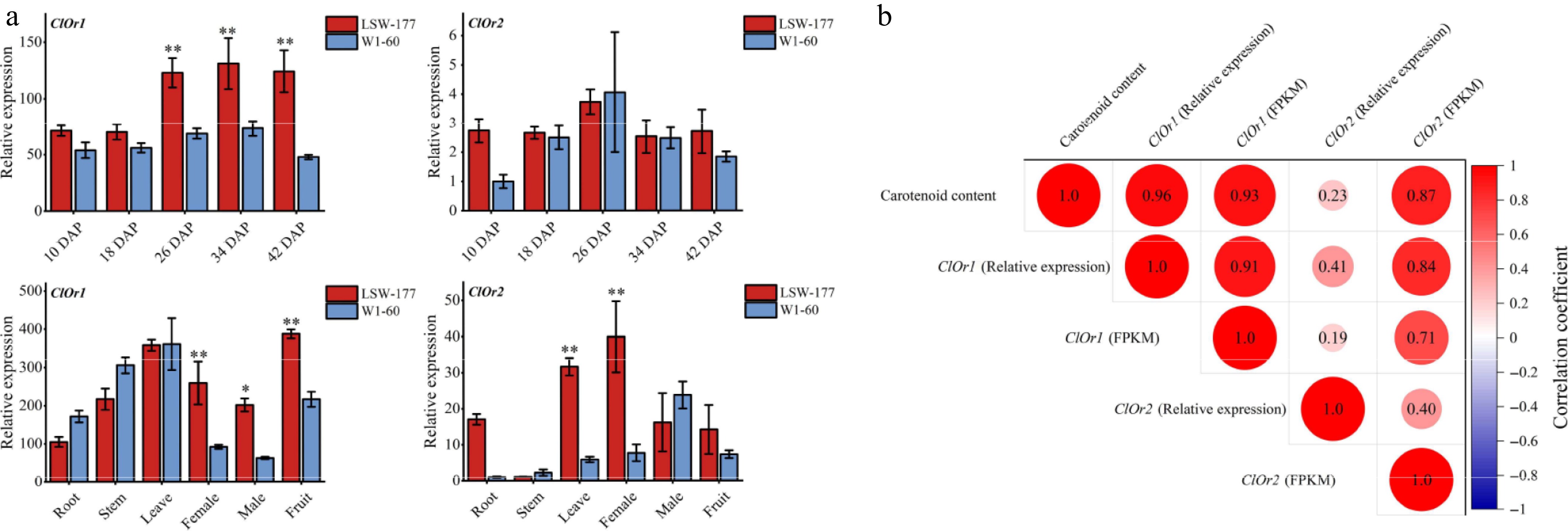
Figure 4.
Expression analysis of the ClOr gene in watermelons. (a) Expression analysis of the ClOr gene in red- and white-fleshed watermelon. Each data point represents the mean of three biological replicates, the error bar represents the mean ± standard error value, * significant difference, p < 0.05; ** p < 0.01 (two-tailed). (b) Correlation analysis between the ClOr gene expression and carotenoid content. Relative expression refers to the fluorescence quantitative data of the ClOr gene in LSW-177 and W1-60 materials at different developmental stages, and FPKM refers to the RNA-Seq data of the ClOr gene in LSW-177 (PRJNA338036) and W1-60 at different developmental stages. The number in the circle pattern is the Pearson correlation coefficient (two-tailed), respectively.
In the correlation analysis between ClOr gene expression and carotenoid content (Fig. 4b), the expression levels of the ClOr1 gene in the flesh of red-fleshed watermelon (LSW-177) and white-fleshed watermelon (W1-60) were significantly correlated with the total carotenoid content at different developmental stages; the correlation coefficients were 0.96 (relative expression and total carotenoid content) and 0.93 (FPKM and total carotenoid content), while the correlation coefficients of the ClOr2 gene were 0.23 and 0.87, respectively. The obtained results suggest that the ClOr1 gene had a higher correlation with carotenoid content in the flesh of watermelon at different developmental periods than that of the ClOr2 gene. Therefore, it is speculated that the ClOr1 gene was involved in carotenoid biosynthesis in watermelon flesh.
Cloning and sequence analysis of ClOr1
-
After cloning the ClOr1 gene successfully (Supplementary Fig. S1a, S1b), the full-length CDS of the ClOr1 gene was 978 bp in red- and white-fleshed watermelons, encoding 325 amino acids. The 87th base of the ClOr1 gene cloned from the flesh of red-fleshed watermelon (LSW-177) was G, encoding the 29th amino acid, tryptophan (Trp), which was the same as that of Watermelon (97103) v2.5 Genome, while the 87th base of the ClOr1 gene cloned from the flesh of white-fleshed watermelon (W1-60) was T, which encodes a cysteine (Cys) here. A dCAPS marker was designed to distinguish the ClOr1 gene in the LSW-177 and W1-60. PCR and restriction enzyme digestion were used to distinguish between G/G, T/T, and G/T. The results confirmed the SNP variation could play an important role in distinguishing the red- and white-fleshed watermelon (Supplementary Fig. S1c, S1d). Secondary and tertiary structure prediction of ClOr1Trp and ClOr1Cys proteins (Supplementary Fig. S2) showed: ClOr1Trp protein consisted of 26.32% α-helix, 10.84% extended strand, 1.24% β-turn, and 61.61% random coil; ClOr1Cys protein consisted of 23.53% α-helix, 9.91% extended strand, 0.93% β-turn, and 65.63% random coil. According to the suggestion, the protein stability could be changed by their structural difference; proteins with a higher proportion of α-helix structure exhibited greater stability. Therefore, the SNP (Cys/Trp) caused a change in ClOr1 protein stability; the stability of ClOr1Trp protein was higher than that of ClOr1Cys protein.
SNP genotype analysis in the ClOr1 gene of watermelon germplasm resources
-
The non-single nucleotide from guanine (G/G) in the LSW-177 to thymine (T/T) in the W1-60 may not only change the gene sequence, but also change the protein sequence of ClOr1 (Fig. 5a). To further confirm whether the SNP (87 bp, T/G, and Cys/Trp) in the ClOr1 gene affects the flesh color of watermelon, the genomic data of 414 publicly available watermelon germplasm resources provided by Guo et al[34], together with 14 germplasm resources of watermelon saved in the laboratory, were utilized to analyze the genotypes of this SNP in the ClOr1 gene. Further, integrating the genotyping and phenotyping results among these 428 watermelon germplasm resources (Supplementary Table S7) showed that, 188 out of 197 red-fleshed watermelons had the G/G genotype, 17 out of 18 pink-fleshed watermelons had the G/G genotype, and 15 out of 16 orange-fleshed watermelons and 20 out of 21 yellow-fleshed watermelons had the G/G genotype, respectively. In contrast, 14 out of 18 pale yellow-fleshed watermelons and 34 out of 49 white-fleshed watermelons had the T/T genotype (Supplementary Table S8, Fig. 5b); these results suggested that this SNP variant in 428 watermelon germplasm resources plays a significant role in controlling the flesh color of watermelons.
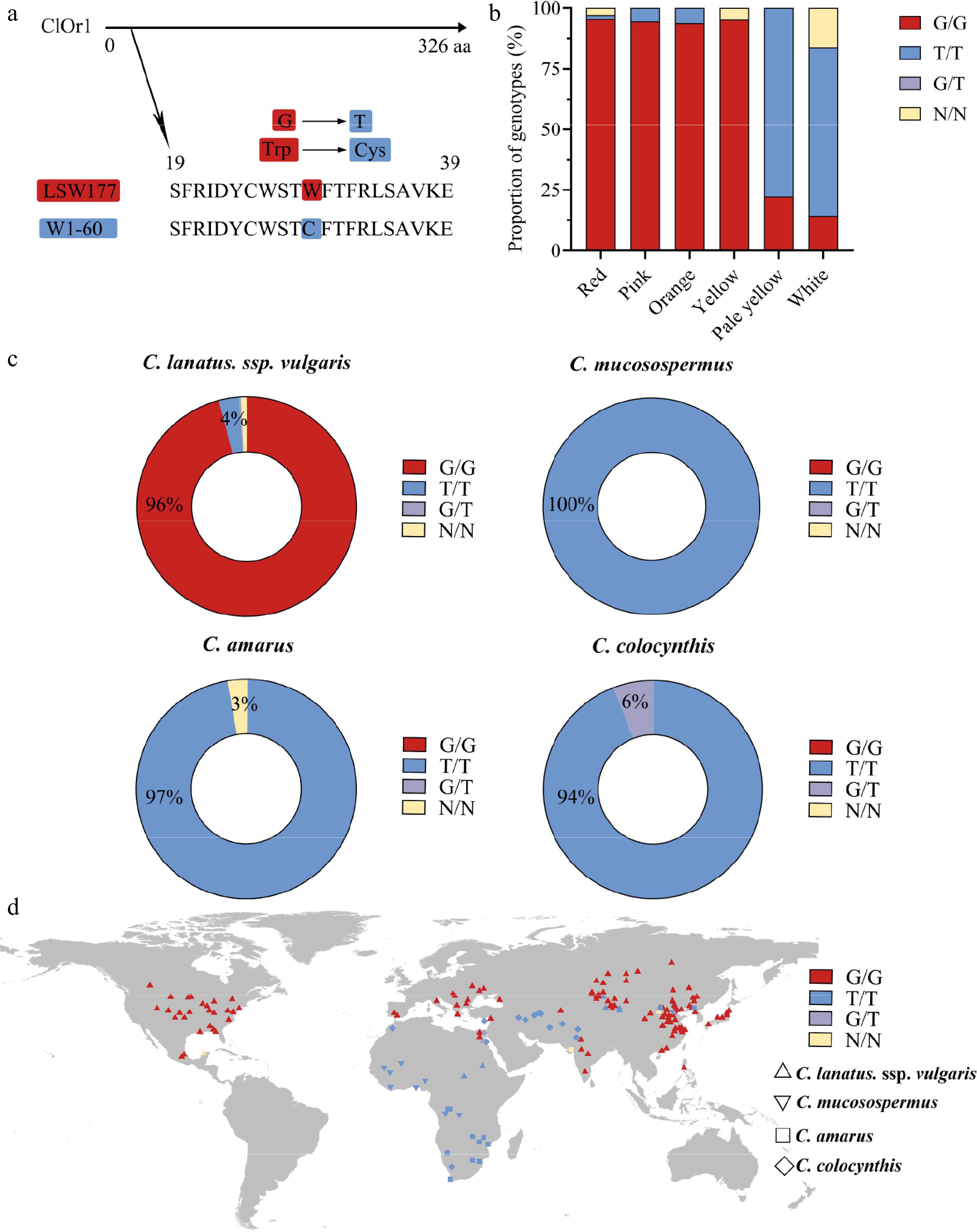
Figure 5.
A single SNP variation in the ClOr1 gene can affect the formation of watermelon flesh color. (a) Sequence variation of ClOr1 protein between LSW-177 and W1-60. A non-synonymous mutation from guanine (G) to thymine (T) at the 29th amino acid of ClOr1 resulted in an amino acid variation from tryptophan (Trp) to cysteine (Cys). (b) Genotype proportions of watermelons with different flesh colors (red, pink, orange, yellow, pale yellow, white). The red sections represented the G/G genotype identical to LSW-177, the blue sections represented the T/T genotype identical to W1-60, the purple sections represented the G/T genotype, and the orange sections represented unknown. (c) Genotype of 406 watermelon accessions using a doughnut chart. (d) Geographical distribution of genomic variations in the ClOr1 gene of 428 watermelon accessions. C. lanatus, ssp. vulgaris was represented by an upright triangle, C. mucosospermus was represented by an inverted triangle, C. amarus was represented by a square, and C. colocynthis was represented by a rhombus.
Moreover, because the LSW-177 belongs to C. lanatus, ssp. vulgaris and the W1-60 belongs to C. amarus, the geographic origin of 428 watermelon germplasm resources were also re-analyzed for insight into the function of the ClOr1 gene. The results showed that there were 341 C. lanatus, ssp. vulgaris accessions, 96% of which were G/G genotypes; 13 C. mucosospermus accessions, all of which were T/T genotypes; 97% of 34 C. amarus; and 94% of 17 C. colocynthis accessions were T/T genotypes (Fig. 5c). G/G genotypes are mainly found in humid and rainy East Asia, North America, and the Mediterranean region, while T/T genotypes are mainly found in arid Africa and West Asia (Fig. 5d). The high concordance between genotypes and ecotypes indicated that this SNP could well distinguish white-fleshed watermelon from non-white-fleshed watermelon, and combined with the expression level of the ClOr1 gene, it was significantly and positively correlated with total carotenoid content (Fig. 4). These results further suggested that the ClOr1 gene might be a key gene affecting the flesh color formation in watermelon.
Subcellular localization of ClOr1 protein
-
The results of subcellular localization of ClOr1 protein exhibited that the green fluorescence expressed by the pBWA(V)HS-Osgfp-ClOr1 fusion vector completely overlapped with the red fluorescence of the cell nucleus Marker (ClOr1-GFP), which significantly indicated that the ClOr1 protein was expressed in the nucleus (Supplementary Fig. S3). Meanwhile, the pBWA(V)HS-Osgfp empty vector and cytosolic Marker showed normal green fluorescence and red fluorescence (Free-GFP), respectively, indicating that the fluorescent proteins of both BWA(V)HS-Osgfp and cytosolic Marker could be expressed normally, which depicted that the experimental results were highly credible.
Total carotenoid content in OE-ClOr1 tomato fruits at different developmental stages
-
To elucidate the role of the ClOr1 gene in carotenoid accumulation, tomato plants with ClOr1 overexpression were generated. The positive transgenic tomato plants (OE-1, OE-2, OE-3) flowered earlier than the WT tomato (Fig. 6a), and the fruits ripened earlier than the WT tomato (Fig. 6b). The fruits of OE tomato were at the mature green stage for 31 ± 1 DPA; the breaker stage and red ripe stage were at 34 ± 2 DPA and 36 ± 2 DPA respectively (DPA: Days post anthesis); while WT tomato fruits reached the mature green and breaker stages at 36 ± 3 DPA and 40 ± 5 DPA, respectively (Fig. 6c).
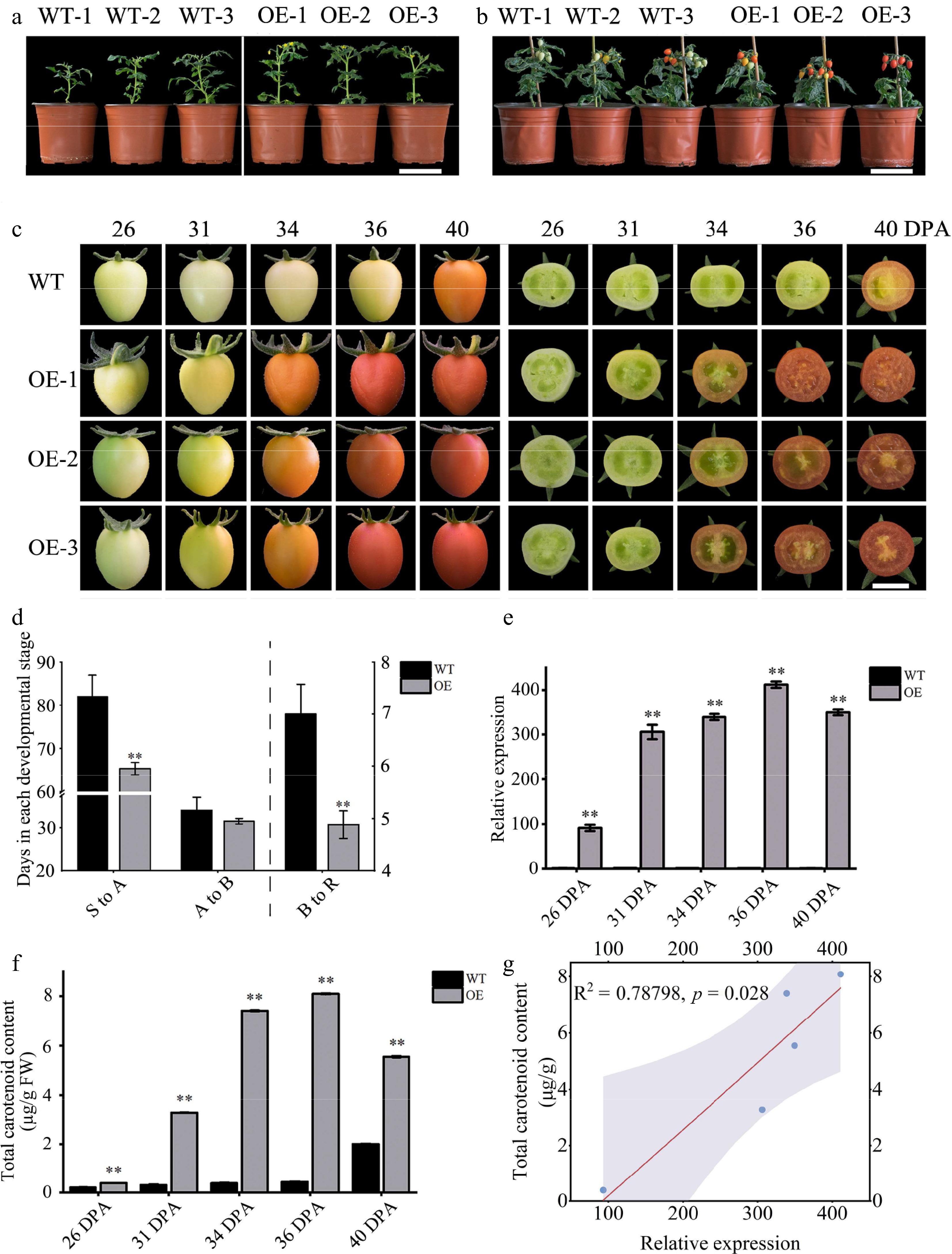
Figure 6.
Phenotypic and gene expression analysis of WT and OE tomatoes. (a) First flowering state of tomato plants, scale = 10 cm. (b) Fruit development status of tomato plants, scale = 10 cm. (c) Tomato fruit phenotype, scale = 1 cm. (d) Number of days required for each developmental period in tomato fruit; see the left Y axis for (S to A) and (A to B), and the right Y axes for (B to R). (e) The expression level of the ClOr1 gene in fruits at different ripening stages, in which the expression level of the 26 DPA fruit of WT tomato was used for calibration. (f) Total carotenoid content in fruits at different ripening stages. In (d)–(f), each data point represents the mean of three biological replicates, the error bar represents the mean ± standard error value, ** significant difference between the WT and OE tomatoes, p < 0.01 (two-tailed). (g) Correlation analysis between the ClOr1 gene expression and total carotenoid content.
After statistical analysis, OE tomato took significantly less time than WT tomato from sowing to anthesis (S to A) and fruit breaker to red (B to R) (Fig. 6d), which showed that overexpression of the ClOr1 gene in tomato can promote flowering and ripening. Compared with the WT tomato, the expression level of the ClOr1 gene was significantly increased in OE tomato fruits at different developmental stages, ranging from 91.4 to 440.1-fold, and reached a peak at the red ripe stage of tomato (36 DPA) (Fig. 6e). Meanwhile, the total carotenoid content in the fruits of OE tomatoes at different developmental stages was significantly increased compared to WT tomatoes (Fig. 6f), with the increase ranging from 1.8 to 19.2-fold. In addition, the correlation analysis showed that the gene expression of ClOr1 in the fruits of OE tomatoes was significantly and positively correlated with the total carotenoid content (Fig. 6g).
TRV-mediated ClOr1 gene silencing decreased carotenoid accumulation in the watermelon fruit flesh
-
Significant phenotypic changes were observed in watermelon fruits after TRV-mediated ClOr1 gene silencing. It was found that the pTRV2-injected fruit portion was normally transformed to red, whereas the pTRV2-ClOr1-silenced fruit could not be smoothly transformed to red (Fig. 7a). In the qRT-PCR analysis and carotenoid content assay (Fig. 7b), compared to the control group pTRV2, the expression level of the ClOr1 gene was down-regulated by 23.6%, and the total carotenoid content was significantly down-regulated in the experimental group pTRV2-ClOr1. The results contributed to the fact that silencing the ClOr1 gene resulted in a significant reduction of total carotenoid content in watermelon fruit flesh.
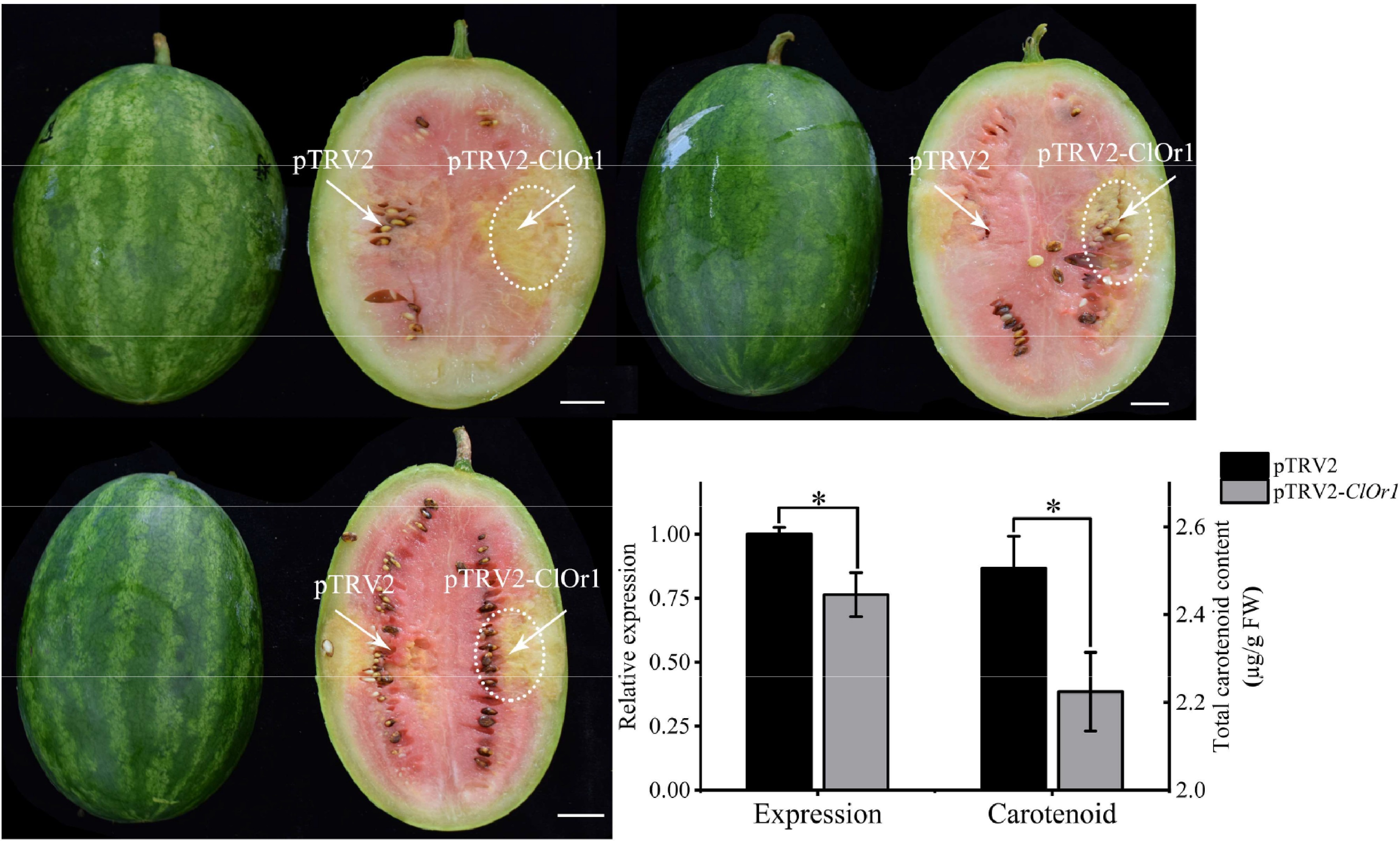
Figure 7.
Analysis of watermelon flesh transformation effect. (a) VIGS effect diagram of watermelon flesh, scale = 2 cm. (b) qRT-PCR analysis and total carotenoid detection, and the expression level of pTRV2 in the control group was used for calibration. * Significant difference between pTRV2 of the control group, and pTRV2-ClOr1 of the experimental group, p < 0.05 (two-tailed). Each data point represents the mean of three biological repeats, and the error bar represents the mean ± standard error value.
The interaction between ClOr1 and ClPSY1 prompted the synthesis and accumulation of carotenoids in watermelon flesh
-
The Y2H assay was used to identify the interaction between ClPSY1 and ClOr1; the ClOr1 and CIPSY1 were constructed into the DNA-binding domain (BD) and the transcription activation domain (AD) of the GAL4 system, respectively. Compared to the control group, only yeast cells harboring AD-ClOr1 + BD-ClPSY1 grew normally and exhibited blue coloration on SD/-Leu/-Trp/-Ade/-His medium containing 20 mg/L X-α-gal, confirming a strong interaction between ClOr1 and CIPSY1 (Fig. 8a). Higher α-GAL activity than other groups also revealed a strong interaction between ClOr1 and ClPSY1 proteins (Fig. 8b). To verify whether ClOr1 and ClPSY1 interact in vivo, bimolecular fluorescence complementation (BiFC) assays were performed. YC-ClOr1 + NY-ClPSY1 showed a strong yellow fluorescence signal in the YFP fluorescence channel that was much higher than the background, and the yellow fluorescence overlapped with the spontaneous fluorescence of chloroplasts. These results indicated that the ClOr1 protein and ClPSY1 protein exhibited direct interaction in tobacco leaf epidermal cells, and their interaction is located in the chloroplast (Fig. 8c).
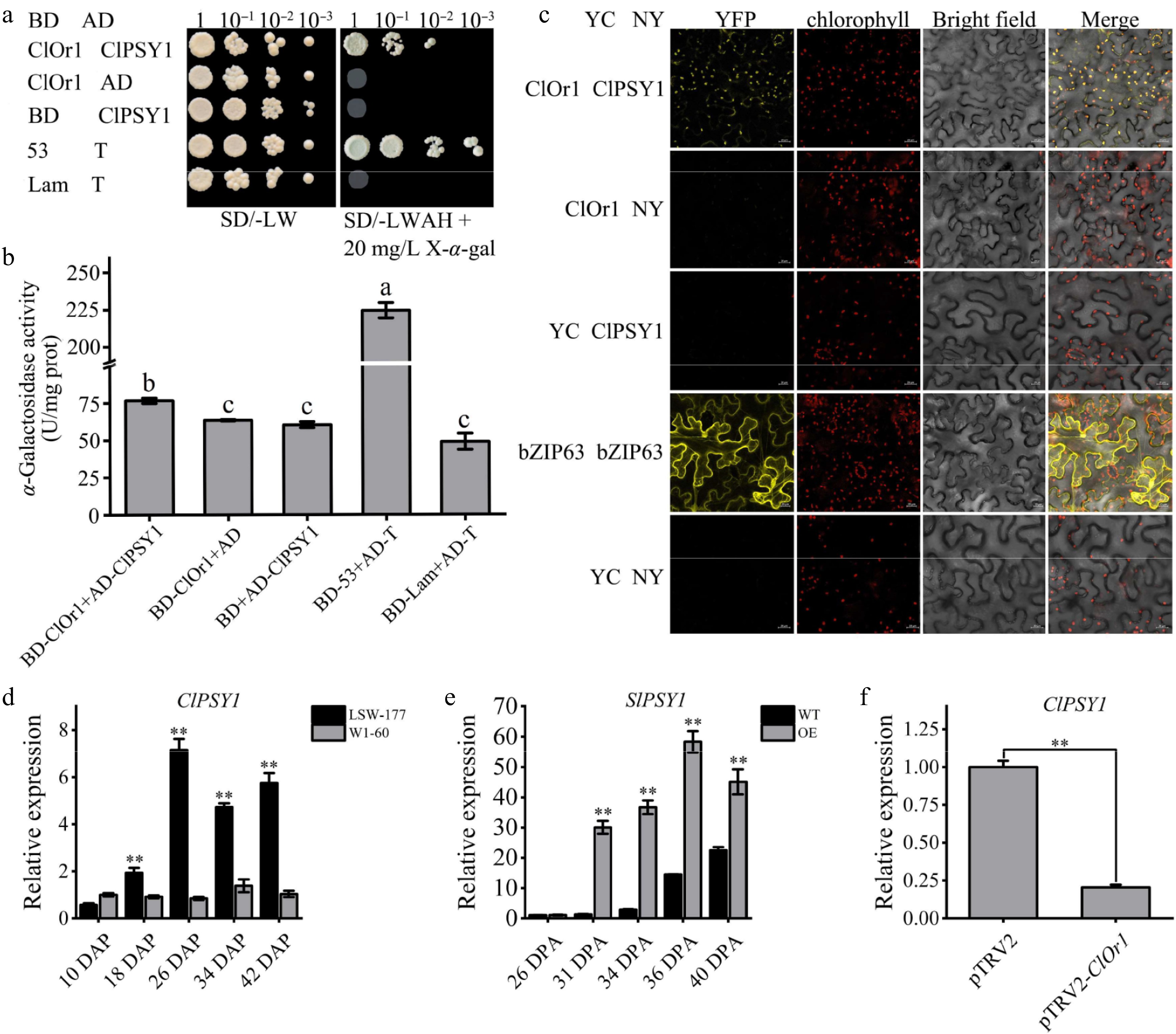
Figure 8.
ClOr1 interacted with ClPSY1. (a) Y2H assays. Growth of co-transformed yeast on non-selective SD/-LW medium and selective SD/-LWAH medium. Positive control BD-53 + AD-T (pGBKT7-53 + pGADT7-T), negative control BD-Lam + AD-T (pGBKT7-Lam + pGADT7-T). (b) α-Galactosidase activity detection. U/mg prot, unit enzyme activity, is 1 nmol p-nitrophenol per mg histone per hour. The results of the one-way analysis of variance were indicated by different lowercase letters (p < 0.05). (c) BiFC assays. YFP: yellow fluorescent channels; Bright field: chloroplast spontaneous fluorescent channels; Merge: superposition field. YC-bZIP-63 + NY-bZIP-63 and YC + NY were used as positive control and negative control, respectively. (d), (f) Expression analysis of the ClPSY1 gene in watermelons. (e) Expression analysis of the SlPSY1 gene in tomato fruits at different ripening stages. Bar data are the means of three biological replicates ± standard error value, * p < 0.05; ** p < 0.01 (two-tailed).
Then, it was found that the expression level of the ClPSY1 gene in the LSW-177 material gradually increased with fruit development and reached its peak at 26 DAP. Except for 10 DAP, the expression levels at all other stages were significantly higher than those in W1-60 (Fig. 8d). The expression level of the SlPSY1 gene rose sharply during the mature green stage (31 DPA) of transgenic tomato fruit and peaked at the red ripe stage (36 DPA), proving that the upregulation of the ClOr1 gene expression level can promote the increase in the SlPSY1 gene expression level (Fig. 8e). In VIGS, the expression level of the ClPSY1 gene decreased significantly, indicating that the downregulation of the ClOr1 expression level can inhibit the expression of the ClPSY1 gene (Fig. 8f). Correlation analysis showed that the expression of ClPSY1 and SlPSY1 genes was significantly correlated with the expression of the ClOr1 gene and the total carotenoid content (Supplementary Fig. S4a, S4b).
-
The flesh color of watermelon is an important trait closely associated with nutritional and economic value. The reason for the different flesh colors of watermelon is the difference in the composition and accumulation of carotenoid content[35]. In this study, the results of carotenoid detection at different stages of development in the flesh of red-fleshed watermelon (LSW-177) and white-fleshed watermelon (W1-60) indicated that mature fruit flesh of the LSW-177 primarily contained lycopene, (E/Z)-phytoene, and β-carotene, while the primary carotenoid in W1-60 fruit flesh was lutein (Fig. 1, Supplementary Tables S3, S4, S5). It has been reported that red-fleshed watermelons contained high levels of lycopene, yellow-fleshed watermelons contained traces of lutein and β-carotene or a small amount of pre-lycopene, and orange-fleshed watermelons contained orange pre-lycopene and β-carotene[36,37]. The white-fleshed watermelon variety (ZXG507) contained only small amounts of lutein and violaxanthin[38]. This study further verified that the composition and amount of carotenoid content in watermelons affected the formation of flesh color.
Chromoplast is the key organelle for carotenoid biosynthesis and accumulation[39]. The interior of chromoplasts produces carotenoid-lipoprotein substructures to sequester and stably store large amounts of carotenoids[40]. The results of chromoplast observation in the fruit of red-fleshed watermelon (LSW-177) and white-fleshed watermelons (W1-60) at different developmental stages showed that the LSW-177 was rich in carotenoid crystals, and the number and size of plastids also continued to increase, while the fruits of W1-60 contained only globular chromoplasts and developed slowly (Fig. 2). The crystalline chromoplasts formed in the mature fruit cells of red-fleshed watermelons were related to the accumulation of lycopene and β-carotene, while the only globular chromoplasts in the fruit cells of white-fleshed watermelons were related to the accumulation of carotenoids such as lutein. The compositions and contents of carotenoids can affect the differentiation of chromoplast, which has been reported in red-fleshed watermelon, red tomato, and red-fleshed papaya[19,41,42]. In studies of colorless or weakly colored tissues such as young watermelon fruit[19], citrus fruit[43], white carrot root system[44], white potato tuber[45], and flower bulb[20], the chromoplasts could be transformed from proplastids. Therefore, it is hypothesized that the chromoplasts in the fruit cells of red- and white-fleshed watermelon are transformed from proplastids.
It is well established that the Or gene can indirectly regulate plant carotenoid accumulation by regulating attributes such as differentiation, number, and size of chromoplasts[7,46]. The function of acquired mutations in the Or gene has been shown to be related to the regulation of the differentiation of chromoplasts to promote carotenoid accumulation in studies of the cauliflower BoOr natural mutant[20], the melon CmOr natural mutant[21], and the Arabidopsis thaliana AtOrHis gene[47]. Two genes of the Or family in watermelons were characterized, ClOr1 and ClOr2 (Fig. 3, Supplementary Table S6). The expression of ClOr1 and ClOr2 in the fruit at different developmental stages and 34 DAP root, stem, leaf, and female and male flower tissues of LSW-177 and W1-60 watermelons was determined, and it was found that the expression level of the ClOr1 gene was higher in the mature fruit of red-fleshed watermelons than in other tissues. The expression level of the ClOr1 gene was higher in all tissues of watermelon compared with that of the ClOr2 gene, and at the same time, the expression of the ClOr1 gene was significantly correlated with the carotenoid content of the flesh of red- and white-fleshed watermelons at different developmental stages (Fig. 4). RNA-Seq data from watermelon (97103, v2.5) with different flesh colors were used to further analyze the expression patterns of the ClOr1 and ClOr2 genes, and the transcript levels of the ClOr1 genes in red-, pink-, yellow-, and white-fleshed watermelons increased with fruit maturity, but the ClOr2 gene did not show any significant changes[6,48,49]. A single-base missense mutation SNP (87 bp, T/G, Cys/Trp) was found in the ClOr1 gene, after cloning the ClOr1 gene and comparing the sequences of the LSW-177 and W1-60 (Supplementary Fig. S1). A dCAPS tool was developed based on this SNP and combined with 428 germplasm resources of watermelon. It was confirmed that the SNP in the ClOr1 gene could well discriminate between white-fleshed watermelon germplasm resources and non-white-fleshed watermelon germplasm resources (Fig. 5, Supplementary Tables S7, S8), indicating that the ClOr1Trp gene could promote carotenoid accumulation in watermelon flesh. Therefore, the ClOr1Trp in the watermelon of LSW-177 was identified as a key candidate for promoting carotenoid accumulation in the fruit flesh.
Or has been shown as an important genetic tool for carotenoid enhancement in major plants[17,28]. Overexpression of the OsOR gene modified thylakoid development in rice and reduced carotenoid content[50]. Tomato SlOR protein regulated the accumulation of chlorophyll and carotenoid, and affected tomato adaptation to high light intensity and disease resistance[51]. The Or gene isolated from a cauliflower mutant increases carotenoid content in potato tubers[52]. The Arabidopsis Or gene increased the carotenoid content of endosperm significantly in transgenic rice and maize lines[24,53]. In this study, tomato was transformed with the ClOr1, resulting in fruit coloration and much higher carotenoid accumulation in the fruit (Fig. 6). In addition, the down-regulation of the expression level of the ClOr1 gene in the virus-induced ClOr1 gene silencing assay resulted in a significant down-regulation of carotenoid content (Fig. 7). Thus, the ClOr1 gene functions to regulate carotenoid accumulation. Most studies have shown that Or proteins interact with phytoene synthase PSY[47,54]. So far, the ClPSY1 has been confirmed to regulate the formation of watermelon flesh color and is a key component of the carotenoid metabolic pathway[55,56]. The results of the Y2H and α-galactosidase activity difference in this study indicated that there was a strong interaction between the ClOr1 protein and the ClPSY1 protein; the results of the BiFC assays indicated that the interaction between the ClOr1 protein and ClPSY1 protein was located in the plastid, while the ClOr1 protein was located in the nucleus. To the best of current knowledge, the subcellular localization of the Or gene may be dynamic and change according to the physiological state or developmental stage of the cell. For example, in Arabidopsis, AtOr had a dual localization pattern of nucleus and plastid[57]. Meanwhile, in BoOr transgenic Arabidopsis, the cauliflower BoOr protein was expressed in the plastid of the leaf epidermal cells and the developing seeds[20], while the BoOR protein was also expressed in the nucleus in the young bud cells of transgenic Arabidopsis[58]. This study found that ClOr1 could affect the ClPSY1 expression level, similar phenomena have also been reported in Arabidopsis[47], which may be because there was a positive regulatory relationship between ClOr1 and ClPSY1: the upregulation of ClOr1 expression promoted the differentiation of chromoplast, which was equivalent to an increase in the storehouse of carotenoids accumulation, leading to the synthesis of more carotenoids. Therefore, the ClPSY1 expression level would increase due to the increase in carotenoids. These results suggested that the ClOr1 gene can positively influence the expression level of the ClPSY1 gene in chromoplasts of watermelon flesh, thereby post-transcriptionally regulating the synthesis and accumulation of carotenoids. (Fig. 8, Supplementary Fig. S4).
-
This study provides new evidence for the role of the ClOr1 gene in the regulation of carotenoid accumulation, chromoplast differentiation, and fruit coloration in plants. It was demonstrated that the ClOr1 gene was highly expressed only in the ripe flesh of red-fleshed watermelon of the LSW-177, and was significantly correlated with carotenoid content. Overexpression of the ClOr1 gene resulted in fruit coloration in transgenic tomato plants. TRV-mediated ClOr1 gene silencing decreased carotenoid accumulation in the watermelon fruit flesh. Furthermore, the ClOr1 combined with the ClPSY1 promotes carotenoid accumulation. These results will provide insight into the gene function and molecular mechanism of the ClOr1 gene to regulate carotenoid synthesis and accumulation during fruit development and ripening in watermelon, with the hope of promoting the genetic improvement of watermelon fruit flesh based on the ClOr1 gene editing approaches in the future.
This work was supported by the National Natural Science Foundation of China (Grant No. 31960607).
-
The authors confirm their contributions as follows: study design and manuscript revision: Zeng C, Wang P; investigation: Wang P, Zhou C; writing-original draft preparation: Zeng C, Zhou C; data curation: Tang Y, Zeng P; writing−review and editing: Amanullah S, Zhou Q, Zhu Q; project administration, supervision: Zhou Q, Zhu Q. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Chenghong Zeng, Putao Wang
- Supplementary Table S1 Or proteins of 14 other species downloaded from NCBI database.
- Supplementary Table S2 The information of primers used in this study.
- Supplementary Table S3 The information of MRM ion pair.
- Supplementary Table S4 A linear regression equation of relative carotenoid content.
- Supplementary Table S5 Analysis of carotenoids in 18 DAP and 34 DAP fruits of red and white fleshed watermelons.
- Supplementary Table S6 Description of the Or gene family members in watermelon.
- Supplementary Table S7 SNP genotypes in the ClOr1 gene of 428 germplasm resources of watermelon.
- Supplementary Table S8 Classification of SNP genotypes in the ClOr1 gene of 428 germplasm resources of watermelon.
- Supplementary Fig. S1 Characterization and structural analysis of ClOr1.
- Supplementary Fig. S2 Secondary and tertiary structure prediction of ClOr1Trp and ClOr1Cys proteins.
- Supplementary Fig. S3 Subcellular localization results of ClOr1 protein.
- Supplementary Fig. S4 Correlation analysis of ClPSY1 (SlPSY1).
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zeng C, Wang P, Zhou C, Amanullah S, Zeng P, et al. 2026. Deciphering the functional characterization of Orange (Or) genes during development and ripening of watermelon (Citrullus lanatus) fruit. Vegetable Research 6: e007 doi: 10.48130/vegres-0025-0049 

Deciphering the functional characterization of Orange (Or) genes during development and ripening of watermelon (Citrullus lanatus) fruit
- Received: 03 September 2025
- Revised: 05 November 2025
- Accepted: 18 December 2025
- Published online: 09 February 2026
Abstract: Carotenoids are the main nutrients and coloring matter in the flesh of watermelon. However, the significant role of Orange gene in carotenoid accumulation and chromoplast differentiation remains uncharacterized in watermelon. In this study, the ClOr1 gene was obtained through homologous alignment, identified to be highly expressed in the red-fleshed watermelon LSW-177, and significantly associated with carotenoid content. A single-nucleotide polymorphism in the ClOr1 gene was detected and confirmed to distinguish well between white-fleshed and non-white-fleshed watermelon after cloning the ClOr1 from LSW-177 and W1-60, and aligning 428 germplasm resources of watermelon, which suggested that the ClOr1 could be taken as a key candidate gene to promote carotenoid accumulation in watermelon fruit flesh. Transient expression assay similarly revealed that ClOr1 is localized to the nucleus, and overexpression of ClOr1 accelerated fruit coloration by contributing a 1.8- to 19.2-fold increase of total carotenoid content in transgenic tomato lines. Furthermore, the ClOr1 gene silencing resulted in a prominent reduction in carotenoid accumulation in the fruits of LSW-177 using virus-induced gene silencing. A direct protein-protein interaction was identified between ClOr1 and ClPSY1 by Y2H and BiFC assays, and ClPSY1 was exhibited as a key gene based on its significant expression related to carotenoid content in the experimental materials. Hence, these results suggested that the ClOr1 plays a positive role in regulating carotenoid accumulation during development and ripening in watermelon fruit by increasing the activities of ClPSY1 protein.
-
Key words:
- Watermelon /
- Carotenoid /
- Orange gene /
- ClOr1 /
- Functional characterization