









-
Nitrogen (N) is an essential macronutrient for plant growth and development, serving as a structural component of proteins, nucleic acids, and chlorophyll, while also acting as a regulatory signal that coordinates growth, flowering, and fruiting[1]. Although N fertilization has substantially increased crop yields worldwide[2,3], chronic over-application has resulted in low nitrogen use efficiency (NUE) and severe environmental pollution[4,5]. Typically, less than 50% of the applied N is absorbed by crops, with the surplus lost through leaching, denitrification, and volatilization[6]. These problems are further exacerbated in vegetable production systems, where frequent irrigation and heavy fertilization amplify N overuse and losses[7]. In China's intensive greenhouse vegetable systems, annual N surpluses can exceed 3,000 kg hm−2 yr−1, whereas the in-season NUE often averages only about 24%[8]. Therefore, enhancing vegetable NUE under reduced N input represents a critical step toward sustainable production[9].
Cucumber (Cucumis sativus L.) ranks as the third most important vegetable crop worldwide and is a major greenhouse species appreciated for its short growth cycle, high economic return, and nutritional quality[10]. As a nitrate-preferring species with a pronounced N demand[11], cucumber receives an average N input of 1,122.0 kg hm−2, a rate that is 1.27 and 1.88 times higher than the inputs for tomato, and pepper, respectively[7]. However, excessive N application depresses NUE[12] and deteriorates fruit quality by reducing vitamin C, amino acid, and soluble protein contents while elevating nitrate levels beyond food safety thresholds[13−16]. Therefore, given its high N demand, disproportionately high fertilization rate, and documented sensitivity to N-induced quality decline, cucumber is a critical model for developing optimized N management strategies that ensure sustainable and high-quality vegetable production.
NUE is mainly determined by two physiological axes that act sequentially[5,9,17]. Systemic nitrogen signaling (SNS) allows plants to sense heterogeneous soil N and directs root foraging toward patches rich in nitrate[17−20]. Thereafter, nitrate uptake capacity determines how effectively plants acquire external N resources[21]. SNS involves the CEP peptide–CEPR receptor module, which transduces long-distance N-status signals between roots and shoots[22,23]. The nitrate uptake capacity is governed by the activity of high- and low-affinity transport systems such as NRT1.1, and NRT2.1[21]. SNS responsiveness and nitrate uptake capacity are tightly integrated with N remobilization and assimilation processes mediated by nitrate reductase (NR), nitrite reductase (NiR), glutamine synthetase (GS), and glutamate synthase (GOGAT)[24,25]. Despite the fundamental importance of these processes, the extent of their natural variation across cucumber germplasms and their specific contributions to LN tolerance remain largely unexplored.
γ-Aminobutyric acid (GABA) is a stress-responsive metabolite that participates in redox and osmotic homeostasis via the GABA shunt, and functions as a signaling molecule[26]. Accumulating evidence from other species links GABA metabolism to improved tolerance under nutrient deficiency[27,28], but its role in cucumber adaptation to LN stress and its potential association with SNS and nitrate uptake has not been explored.
Here, we developed an integrated pipeline to quantify SNS responsiveness via a hydroponic split- root assay and to measure nitrate uptake capacity through 15N-nitrate tracing across 56 cucumber germplasms. We identified germplasms with contrasting SNS responsiveness and nitrate uptake capacity and evaluated their physiological and molecular responses to low N stress. Comparative transcriptomic and functional analyses revealed that GABA metabolism, particularly the upregulation of CsGS1, contributes to LN tolerance. Our findings uncover a GABA-mediated mechanism that enhances N efficiency, and establish an integrated framework anchored by SNS and nitrate uptake for identifying and developing nutrient-efficient cucumber germplasm.
-
The 56 cucumber germplasm accessions used in this study were obtained from a previously whole-genome re-sequenced panel[29], provided by Dr. Yang Xueyong (Chinese Academy of Agricultural Sciences). Experiments were conducted in a climate chamber (Huazhong Agricultural University) under a 14/10 h (light/dark) photoperiod. Growth conditions were maintained at 28/23 °C and 50%–60%/80%–90% relative humidity (day/night), with a photosynthetic photon flux density (PPFD) of 300 ± 10 µmol m−2 s−1.
Seedling cultivation and hydroponic setup
-
Hydroponic culture was utilized throughout the study. Cucumber seeds were surface sterilized in 0.075% acidic KMnO4 solution for 15 min, followed by thorough rinsing and soaking in water at 55 °C for 6−8 h. The pretreated seeds were then incubated in a growth chamber at 28 °C for 36 h to promote germination. Once radicles reached 2−3 cm, seeds were sown on 2.5 cm × 2.5 cm sponge blocks placed in 20 × 10-hole trays. Upon full expansion of the cotyledons, seedlings were transferred to hydroponic containers (38 cm × 24.5 cm × 10 cm). Each container was covered with a black foam board containing 12 holes, with one seedling per hole. A modified Hoagland nutrient solution with normal nitrate supply (NN, 10 mM KNO3) was used as the basal nutrient solution (Supplementary Table S1a). To maintain adequate dissolved oxygen levels, aeration was provided automatically using small air pumps operating at 1 h intervals.
Split-root experiment design
-
At the stage of full cotyledon expansion, the primary root of each seedling was excised to a length of 2–3 cm to induce lateral root formation. Following excision, seedlings were allowed to recover for 3 d, including an initial 24 h period in darkness with frequent misting to minimize desiccation. After recovery, seedlings were transferred to an ammonium-based Hoagland nutrient solution, in which nitrate was replaced by ammonium nitrogen (Supplementary Table S1b), and grown for an additional 3 d to allow physiological adaptation.
For split-root experiments, the lateral roots of each seedling were evenly divided into two portions and placed into a partitioned hydroponic container. Three nitrate treatments were applied: (1) both compartments supplied with high nitrate (HN, 20 mM KNO3; Supplementary Table S1c); (2) both compartments supplied with low nitrate (LN, 0.1 mM KNO3; Supplementary Table S1d); and (3) a split nitrate treatment, with one compartment supplied with high nitrate (SHN, 20 mM KNO3), and the other with low nitrate (SLN, 0.1 mM KNO3).
After 4 d of treatment, root systems were harvested and scanned using the WinRHIZO Pro 2013 image analysis system. Sensitivity to SNS was evaluated as the ratio of total root length on the SLN side, to that on the SHN side within the same seedling.
Measurement of nitrate (NO3−) uptake rate in cucumber
-
The nitrate (NO3−) uptake rate of 56 cucumber germplasms was determined using a 15N-labeled tracer technique. Two-week-old cucumber seedlings grown hydroponically in the modified Hoagland nutrient solution with normal nitrate supply (NN, 10 mM KNO3; Supplementary Table S1a) were used for the assay. Prior to 15N labelling, roots of each seedling were gently rinsed in 1 mM CaSO4 solution for 1 min to remove surface-adsorbed N. Seedlings were then immersed for 6 min in the modified Hoagland nutrient solution (NN) containing K15NO3 (50 atom% 15N). After labelling, roots were rinsed again in 1 mM CaSO4 solution for 1 min and subsequently washed three times with deionized water. Seedlings were blotted dry with absorbent paper, placed in paper bags, killed at 105 °C for 30 min, and then dried at 72 °C to constant weight.
Total N and 15N contents were determined using an Elementar Vario PYRO cube coupled with an Isoprime100 elemental analyzer–stable isotope ratio mass spectrometer (ELEMENTAR, Germany). The root nitrate uptake rate for each cucumber germplasm was calculated according to the method described by Garnett et al.[30].
The 15N isotopic composition was expressed as δ15N (‰), calculated as:
$ {\delta }^{15}N\left(\text{‰} \right)=\left[\left({R}_{sample}/{R}_{standard}\right)-1\right]\times 1000 $ where, R is the 15N/14N ratio, Rsample is the ratio in the sample, and Rstandard is the ratio of atmospheric N2 (AIR).
Methods for physiological measurements
-
LN stress treatment was initiated after full expansion of the cotyledons, with 0.1 mM KNO3 supplied as the LN condition, while 10 mM KNO3 served as the normal-nitrogen (NN) control. After 8 d of treatment, whole seedlings were harvested for physiological measurements. For dry weight determination, seedlings were oven-dried at 105 °C for 15 min to terminate metabolic activity, followed by drying at 72 °C for 72 h until a constant weight was reached. Dry weight was then determined using an electronic balance.
For the determination of relative electrical conductivity (REC), leaf samples (0.1 g) were excised and cut into approximately 1 cm2 segments, rinsed with deionized water, and incubated in deionized water at 22 °C for 4 h with shaking. The initial electrical conductivity of the bathing solution was recorded as EC1. Samples were then boiled for 30 min, cooled to room temperature, and the final electrical conductivity was recorded again as EC2. REC was calculated as the ratio of EC1 to EC2.
For the determination of malondialdehyde (MDA) content, 0.1 g of leaf tissue was homogenized in an extraction buffer consisting of 1/15 M phosphate buffer (pH 7.8) supplemented with 0.2 mM EDTA and 2% (w/v) polyvinylpyrrolidone. The homogenate was centrifuged at 12,000 rpm for 20 min, and the supernatant was mixed with a reaction solution containing 0.65% (w/v) thiobarbituric acid (TBA) and 10% trichloroacetic acid (TCA). The mixture was heated in a boiling water bath at 100 °C for 15 min, rapidly cooled, and centrifuged at 10,000 rpm for 10 min. The absorbance of the supernatant was measured at 532, 600, and 450 nm, and the MDA content was calculated accordingly.
RNA-Seq analysis
-
For RNA-Seq analysis, seedlings of cucumber accessions 9,930, G23, and G47 were subjected to LN (0.1 mM KNO3), and NN (10 mM KNO3) treatments for 2 d. Roots and leaves were harvested separately from each accession, with three independent biological replicates per tissue per treatment, resulting in a total of 36 samples. Samples were immediately frozen in liquid nitrogen and stored at −80 °C until use. Total RNA extraction, library construction, and Illumina sequencing were performed by Anno Biotechnology (Beijing, China).
Raw reads were processed to remove adaptor sequences and low-quality reads to obtain clean reads, which were then aligned to the cucumber (Cucumis sativus cv. Chinese Long) reference genome (version 3.0). Differentially expressed genes (DEGs) were identified using DESeq2 with thresholds of at p < 0.05 and |log2 (fold change)| > 1. Gene Ontology (GO) enrichment analysis of DEGs was conducted using the Gene Ontology database (
www.geneontology.org ).GABA and 3-MP application
-
Cucumber seedlings were grown hydroponically in a modified Hoagland nutrient solution (NN, 10 mM KNO3) until the one-true-leaf stage and then transferred to LN (0.1 mM KNO3) or maintained under NN (10 mM KNO3) conditions. To evaluate the effects of exogenous γ-aminobutyric acid (GABA) and its biosynthesis inhibitor 3-mercaptopropionic acid (3-MP), three concentrations of GABA (50, 100, and 200 mM), and 3-MP (50, 100, and 200 μM) were applied. Treatment solutions were freshly prepared and applied by foliar spraying, with each seedling receiving 10 mL of the corresponding solution. Control plants were sprayed with an equal volume of distilled water. A second application was performed 5 d after the initial treatment. Phenotypic and physiological parameters were assessed after 10 d of LN and NN treatment.
Statistical analysis
-
Experimental data are presented as the mean ± standard deviation (SD). Differences among treatments were analyzed using two-way ANOVA with the Data Processing System (DPS) software. Statistical significance was determined at p < 0.05.
-
Soil nitrogen availability is spatially heterogeneous. SNS directs root growth towards N-rich patches, a behavior referred to as nitrogen foraging, and is therefore critical for NUE in crops. To evaluate SNS in cucumber, we adapted a split-root system, a well-established approach that mimics spatial heterogeneity of N availability in natural soils[20]. Specifically, after cotyledon expansion, the primary root was excised to induce lateral root (LR) formation. Once the LR system had fully developed, the LR of each seedling was evenly divided into two portions and placed in a partitioned hydroponic container, with one side supplied with SHN (20 mM KNO3), and the other side supplied with SLN (0.1 mM KNO3), thereby simulating heterogeneous nitrogen conditions (Fig. 1a–c). Control seedlings were grown under homogeneous N conditions, with their entire LR systems exposed to either HN (20 mM KNO3), or LN (0.1 mM KNO3).
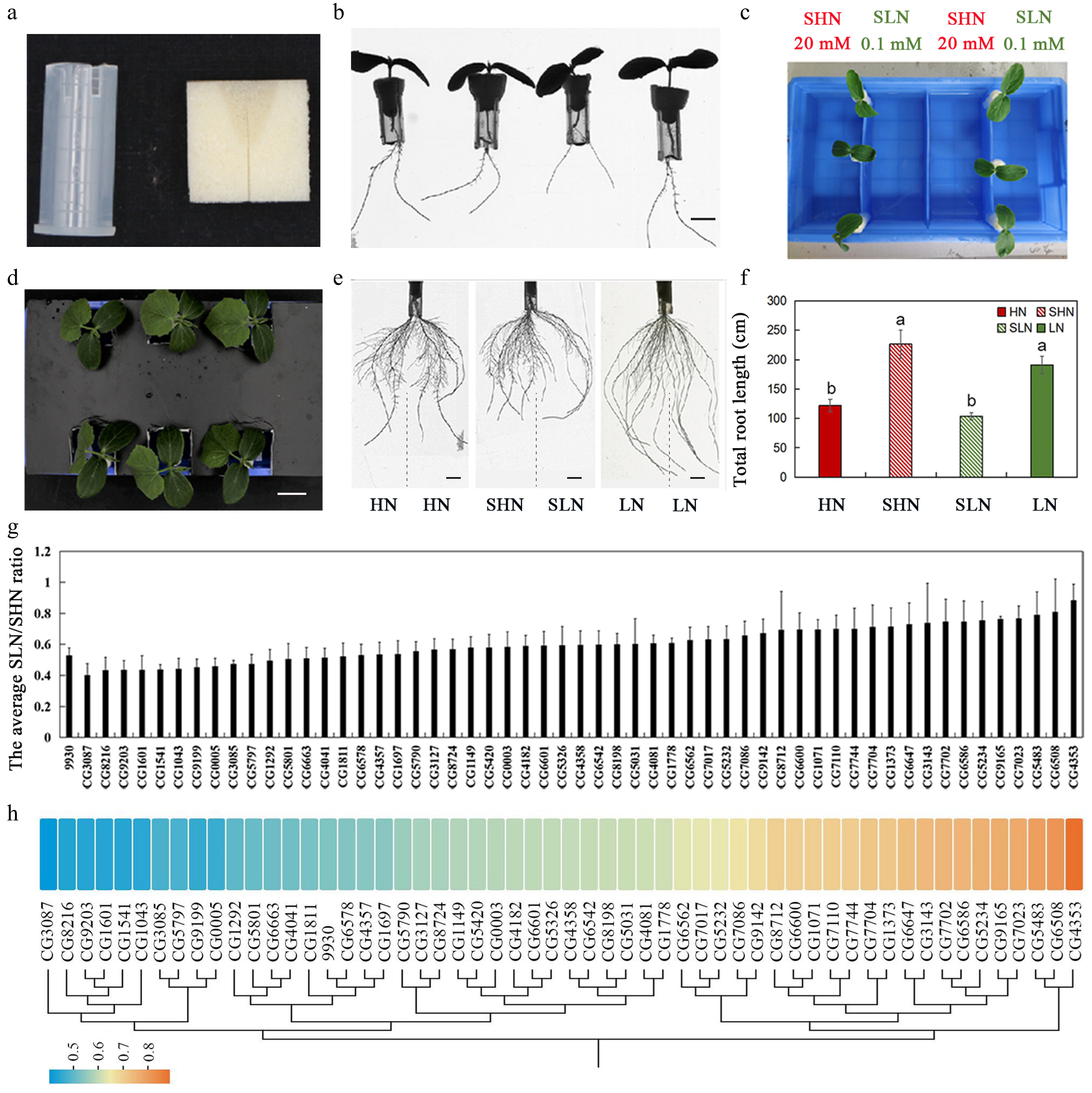
Figure 1.
Evaluation of systemic N signaling sensitivity in different cucumber germplasms using a split-root system. (a) Materials used to assemble the split-root system, including sponge blocks and modified conical (centrifuge) tubes. (b) Cucumber seedlings prepared for split-root experiments, the scale is 1 cm. (c) Configuration of the split-root hydroponic system. Lateral roots from the same seedling were evenly divided between two compartments of a partitioned container, with one side supplied with SHN (20 mM KNO3), and the other with SLN (0.1 mM KNO3). (d) Representative seedlings grown under homogeneous (HN or LN), and heterogeneous (SHN/SLN) conditions, the scale is 2 cm. (e) Representative root system phenotypes under homogeneous HN and LN conditions and under heterogeneous SHN/SLN conditions. Vertical dashed lines illustrate the physical separation of root systems into two compartments of a single plant in the split-root assay. Scale bar equals 1 cm. (f) Total lateral root length under different N concentrations. Different letters represent statistically significant differences. (g) Natural variation in SNS responsiveness among 56 cucumber germplasms, quantified as the SLN/SHN ratio of total LR length (mean ± SD). (h) Heat map of the sensitivity analysis of 56 cucumber germplasms to N signals. Color scale indicates SLN/SHN ratio. The figure also includes a cluster analysis based on the SLN/SHN ratio.
LR length was measured after 4 d of treatment. As expected, homogeneous HN conditions significantly suppressed LR elongation compared with LN, consistent with the well-established inhibitory effect of high nitrate and the promotive effect of low nitrate on LR growth. However, a contrasting response was observed under heterogeneous conditions, with LR growth enhanced on the SHN side but reduced on the SLN side (Fig. 1d–f). Because SHN and HN (20 mM KNO3), as well as SLN and LN (0.1 mM KNO3), had identical local nitrate concentrations, the contrasting responses under heterogeneous conditions cannot be explained by local effects alone, but instead implicate a role for SNS, which transmits signals between the root and shoot to coordinate preferential root growth toward N-rich zones.
To quantify SNS strength, the SLN/SHN ratio of LR length was used as a phenotypic indicator, following previously established methods[20]. A total of 56 cucumber germplasms from a previously whole-genome re-sequenced panel[29] were evaluated (Fig. 1g, h; Supplementary Table S2). Substantial natural variation in SNS responsiveness was observed among germplasms, with SLN/SHN values ranging from 0.40 (CG3087) to 0.88 (CG4353), highlighting pronounced genotype-dependent differences in SNS sensitivity.
Assessing NO3− uptake capacity in cucumber germplasms using 15N-labeled nitrate tracing
-
Nitrate (NO3−) uptake capacity is a key factor governing plant N acquisition, particularly under N limitation, and is therefore a key determinant of NUE in crops. To assess natural variation among cucumber germplasms, we evaluated nitrate uptake capacity using a 15N-labeled nitrate tracing assay, with root δ15N (‰) serving as an uptake indicator, following published protocols with minor modification[30−34]. Across the same panel of 56 germplasms analyzed above, nitrate uptake capacity exhibited striking variation, spanning approximately 15-fold, as reflected by root δ15N (‰) values ranging from 3.30 in the low-uptake accession CG5232, to 50.27 in the high-uptake accession CG3087 (Fig. 2a, b; Supplementary Table S3). These results highlight substantial genetic diversity in nitrate acquisition capacity among cucumber germplasms.
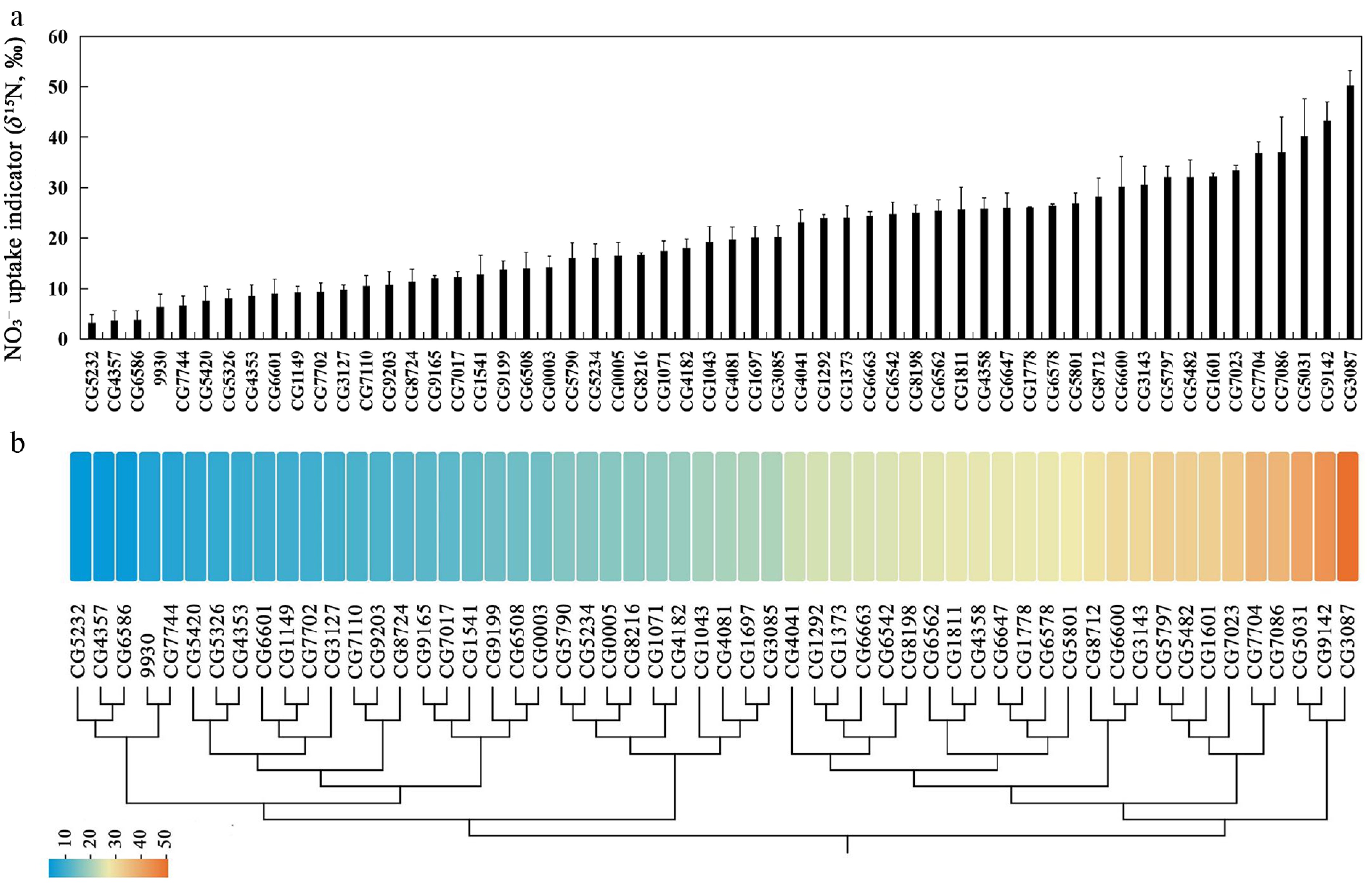
Figure 2.
Natural variation in NO3− uptake capacity among cucumber germplasms revealed by a 15NO3− tracer assay. (a) NO3− uptake capacity assessed by root δ15N (‰) after 15NO3− labelling across 56 cucumber germplasms. (b) Heat map of NO3− uptake capacity (root δ15N, ‰) in 56 cucumber germplasms. Colors bars from blue to red represent low to high NO3− uptake capacity. The dendrogram shows hierarchical clustering based on the NO3− uptake capacity.
The LN tolerance evaluation of germplasm 9930, G23 (CG4353), and G47 (CG3087)
-
Building on the above findings, CG3087 emerged as the strongest SNS responder (lowest SLN/SHN ratio) with the highest NO3− uptake capacity, contrasted sharply with CG4353, which showed weaker SNS responsiveness (higher SLN/SHN ratio) and lower uptake. To examine whether these differences translate into whole-plant performance under N limitation, we selected CG3087 (high SNS and high uptake), CG4353 (low SNS and low uptake), and the sequenced cultivar 9930 (high SNS but lower uptake) as a reference for comparative LN tolerance assays (Figs. 1g, 2a). For readability and consistency with internal sample tracking, we hereafter refer to CG3087 as germplasm 47 (G47), and CG4353 as germplasm 23 (G23).
Seedlings of cucumber germplasms 9930, G47, and G23 were subjected to LN (0.1 mM KNO3) or NN (10 mM KNO3) for 8 d. Relative to NN, LN resulted in visually evident growth inhibition and leaf chlorosis (Fig. 3a). Quantitative analyses showed that LN significantly decreased whole-plant and shoot biomass across all three genotypes, with the magnitude of reduction following G47 < G23 < 9930 (Fig. 3b, c). Root responses varied among germplasms: LN significantly reduced root dry weight (Fig. 3d), total root volume (Fig. 3e), and root tip number (Fig. 3f) in 9930 and G47—more strongly in 9930—whereas these metrics increased in G23 (Fig. 3d–f). Thus, LN inhibited root growth more in 9930 than in G47, whereas G23 maintained or enhanced root growth under LN.
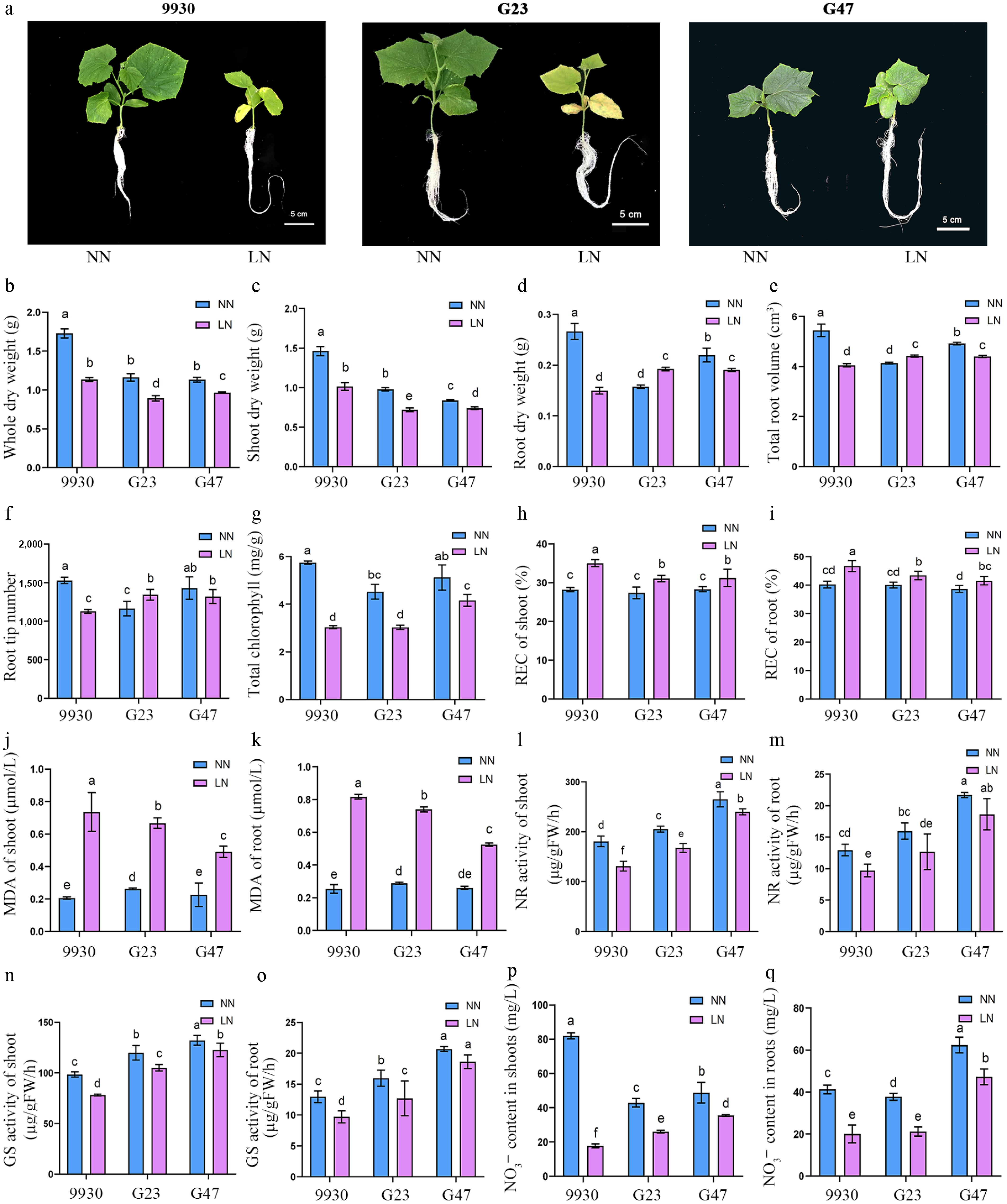
Figure 3.
Growth, physiological and biochemical responses of cucumber accessions 9930, G23 and G47 to LN stress. (a) Growth phenotypes, (b) whole-plant dry weight, (c) shoot dry weight, (d) root dry weight, (e) total root volume, (f) root tip number, (g) total chlorophyll content, (h) REC of shoots, (i) REC of roots. (j) MDA of shoots, (k) MDA of roots, (l) NR activity in shoots, (m) NR activity in roots, (n) GS activity in shoots, (o) GS activity in roots, (p) NO3− content in shoots, (q) NO3− content in roots, of cucumber germplasm 9930, G23, and G47 grown under LN and NN treatment for 8 d.
We next evaluated physiological indicators to elucidate LN stress responses. Total chlorophyll, in line with the visual chlorosis in Fig. 3a, was reduced across all three germplasms under LN (Fig. 3g), with reductions largest in 9930, intermediate in G23, and smallest in G47, indicating the least photosynthetic impairment in G47. Consistent with this protective profile, relative electrolyte conductivity (REC) and MDA—proxies for membrane injury and lipid peroxidation—increased least in G47, intermediate in G23, and most in 9930 (Fig. 3h−k), indicating better membrane integrity and redox homeostasis in G47.
Finally, we examined N assimilation and nitrate status. Activities of nitrate reductase (NR) and glutamine synthetase (GS), key enzymes in nitrate reduction and assimilation, were significantly reduced by LN in all germplasms, with inhibition least in G47 and most in 9930 (G47 < G23 < 9930) (Fig. 3l−o). LN also decreased tissue NO3− content, with leaf declines of 78.35% (9930), 39.15% (G23), and 27.31% (G47), and root declines of 51.55%, 43.86%, and 24.17%, respectively, indicating suppressed nitrate uptake and retention, but to a lesser extent in G47 and G23 (Fig. 3p, q).
Together, morphological, physiological, and biochemical readouts converge on a consistent ranking of LN tolerance: G47 > G23 > 9930. Notably, G47 exhibits strong SNS responsiveness and high nitrate uptake that translate into superior LN tolerance, whereas G23 appears to partially offset lower SNS/uptake by reallocating resources to root elongation and branching under LN.
RNA-seq analysis of G47, G23, and 9930 germplasm under LN
-
To investigate the transcriptional basis of the contrasting LN responses among 9930, G23, and G47 to low N stress, we performed RNA-seq on leaf and root tissues after 2 d of LN and NN treatment. Firstly, principal component analysis (PCA) revealed that the three replicate samples of each treated leaf and root were within the 95% confidence ellipse range, indicating that the transcriptome data were accurate and reliable. Additionally, PCA also revealed that the germplasms G23 and 9930 clustered closer together under both LN and NN, while the G47 was distinctly separated (Fig. 4a, b). Furthermore, the DEGs were identified in leaves and roots under LN treatment. Interestingly, we found that LN-sensitive germplasms 9930 had the least number of DEGs (462 and 1804), both in roots and shoots (Fig. 4c, d). In summary, the tolerant and sensitive germplasms exhibited distinct gene expression patterns, and the germplasms G47 and G23 with better performance under LN stress likely achieve enhanced adaptation to N deficiency through more dynamic gene expression reprogramming.
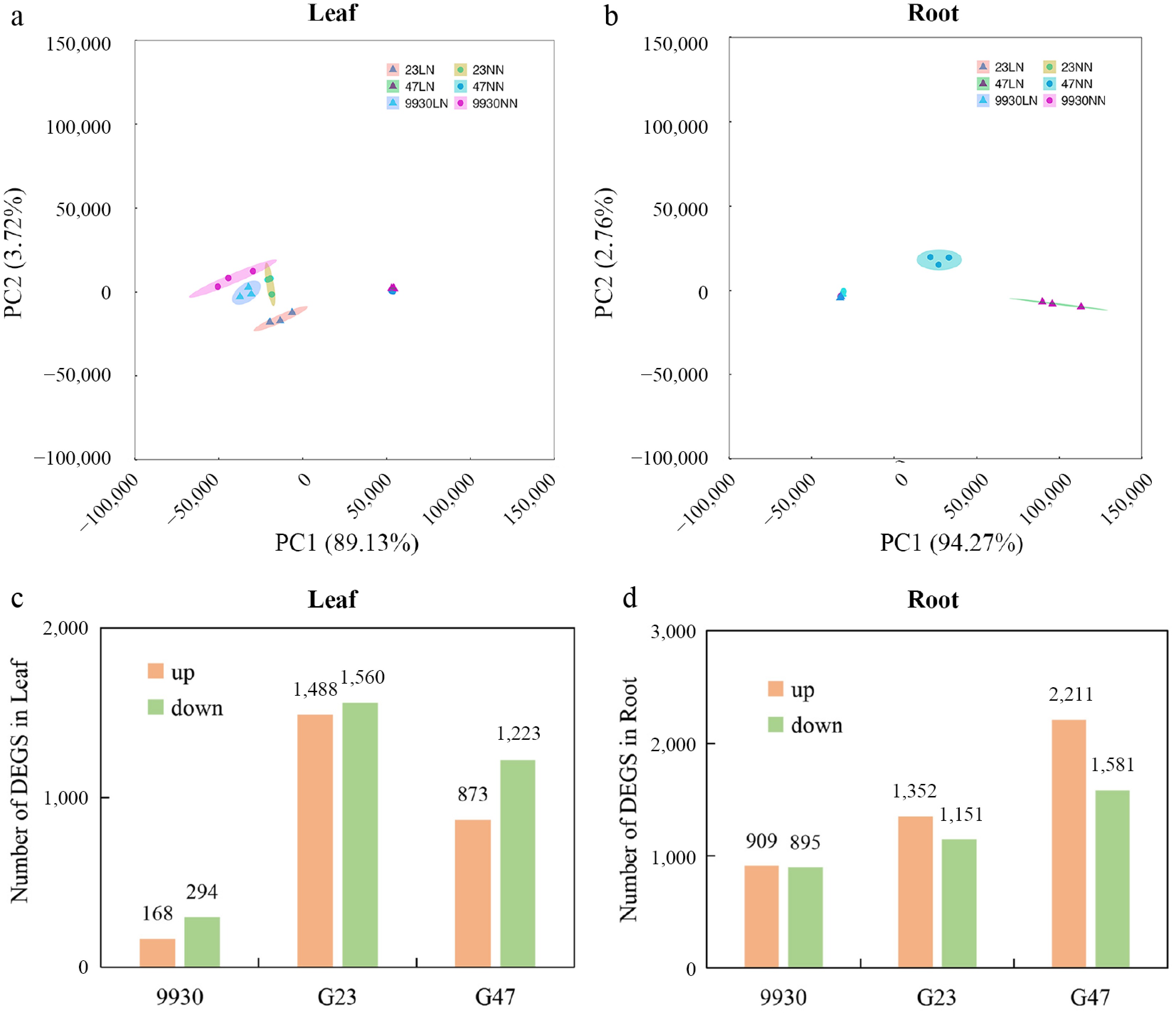
Figure 4.
RNA-seq analysis of leaf and root transcriptomes under LN stress. Principal component analysis (PCA) score plots (PC1 vs. PC2) of (a) leaf, and (b) root samples under LN and NN treatments. Numbers of DEGs that were upregulated (up), and downregulated (down) in (c) leaf, and (d) root of different genotypes after 2 d of LN treatment (relative to NN).
The pathway enrichment analysis of DEGs
-
To further explore the molecular mechanisms underlying cucumber responses to LN stress, Gene Ontology (GO) and KEGG enrichment analyses were performed using the shared DEGs between G23 and G47, the two germplasms exhibiting higher LN tolerance than 9930. A total of 541 shared DEGs in leaves and 619 shared DEGs in roots were identified (Fig. 5a, b). In leaves, shared DEGs were primarily enriched in GO terms related to antioxidant activity, oxidoreductase activity, and calcium ion binding, and were significantly associated with KEGG pathways including N metabolism, alanine, aspartate and glutamate metabolism, and galactose metabolism (Fig. 5c, d). In roots, shared DEGs were enriched in GO terms associated with protein heterodimerization activity, transferase activity, and catalytic activity, and in KEGG pathways related to N metabolism, alanine, aspartate and glutamate metabolism, and fatty acid degradation (Fig. 5e, f). Notably, alanine, aspartate, and glutamate metabolism are closely linked to GABA biosynthesis, which has been implicated in antioxidant regulation and stress responses under LN conditions. Consistent with this, genes involved in GABA biosynthesis and metabolism were more strongly upregulated in G47 and G23 than in 9930 (Fig. 5g), suggesting that enhanced GABA-associated metabolic reprogramming may contribute to enhanced LN tolerance in cucumber.
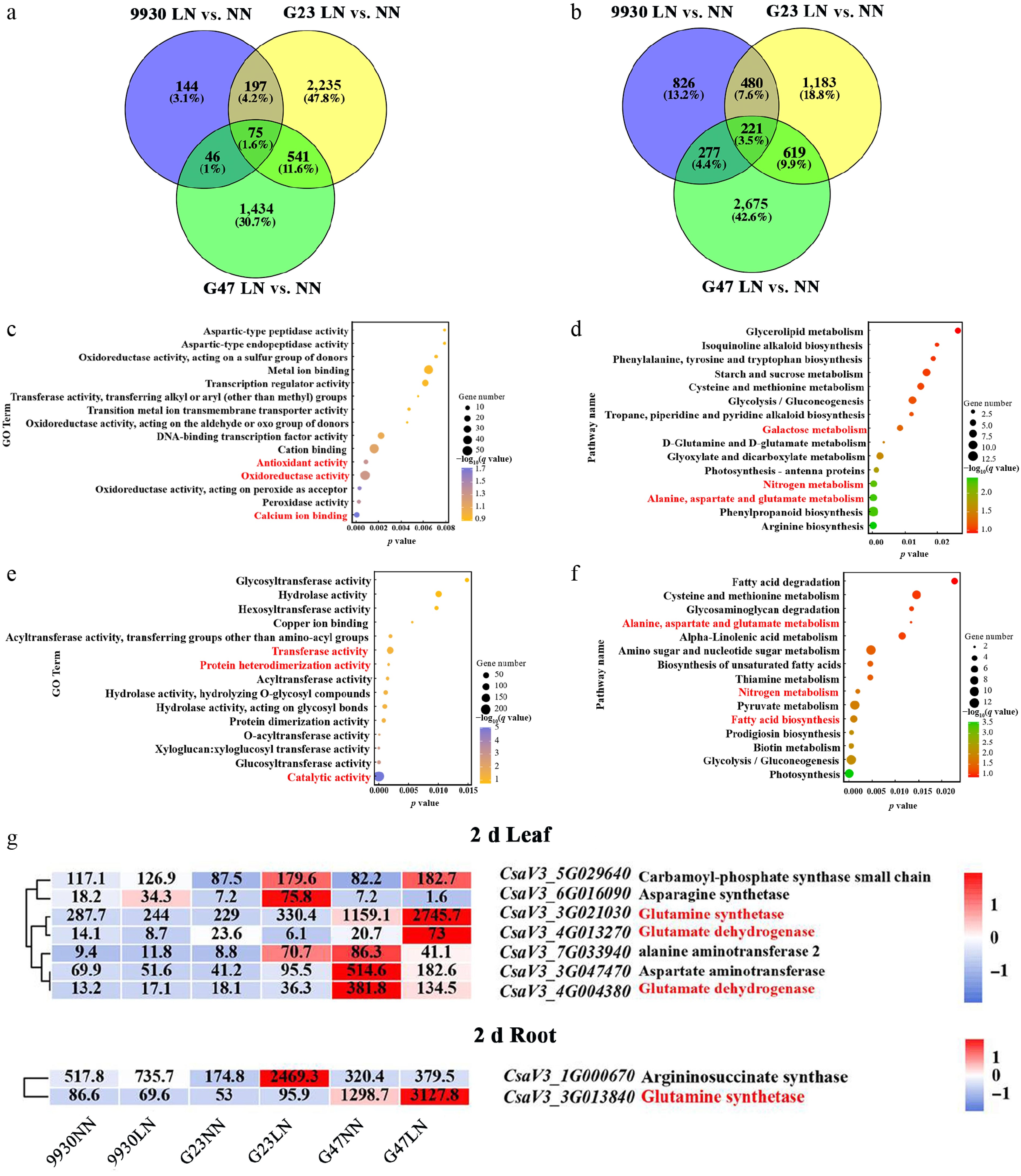
Figure 5.
DEG profiling of cucumber accessions 9930, G23, and G47 in leaves and roots after 2 d of LN treatment. Venn diagram showing DEGs identified for LN vs. NN in (a) leaves, and (b) roots. (c), (e) GO enrichment analysis, and (d), (f) KEGG pathway enrichment analysis of DEGs shared by G23 and G47 under LN in (c), (d) leaves, and (e), (f) roots. (g) Heatmaps show the expression of genes involved in GABA biosynthesis and metabolism in leaves and roots of 9930, G23 and G47 under LN for 2 d.
Exogenous GABA application improves LN tolerance in cucumber germplasm 9930
-
To further validate the transcriptome-inferred role of GABA in mediating LN stress tolerance, we first conducted dose-response screening assays with exogenous GABA and its biosynthesis inhibitor 3-mercaptopropionic acid (3-MP), which identified 50 mM GABA and 200 μM 3-MP as the most effective concentrations for subsequent experiments (Supplementary Fig. S1). Further analysis showed that foliar application of 200 μM 3-MP or 50 mM GABA to cucumber germplasm 9930 had no significant effect on growth under NN conditions (Fig. 6a, b). Under LN conditions, however, treatment with 200 μM 3-MP markedly suppressed growth, resulting in significant reductions in both shoot and root dry weights (Fig. 6c, d) and exacerbating oxidative stress, as evidenced by elevated MDA and REC in shoots and roots (Fig. 6e–h). In contrast, foliar application of 50 mM GABA significantly enhanced LN tolerance, promoting shoot and root growth and alleviating oxidative damage, as indicated by reduced MDA and REC levels under LN conditions (Fig. 6c–h). Collectively, this data show that exogenous GABA application confers improved growth performance under LN in cucumber accession 9930.
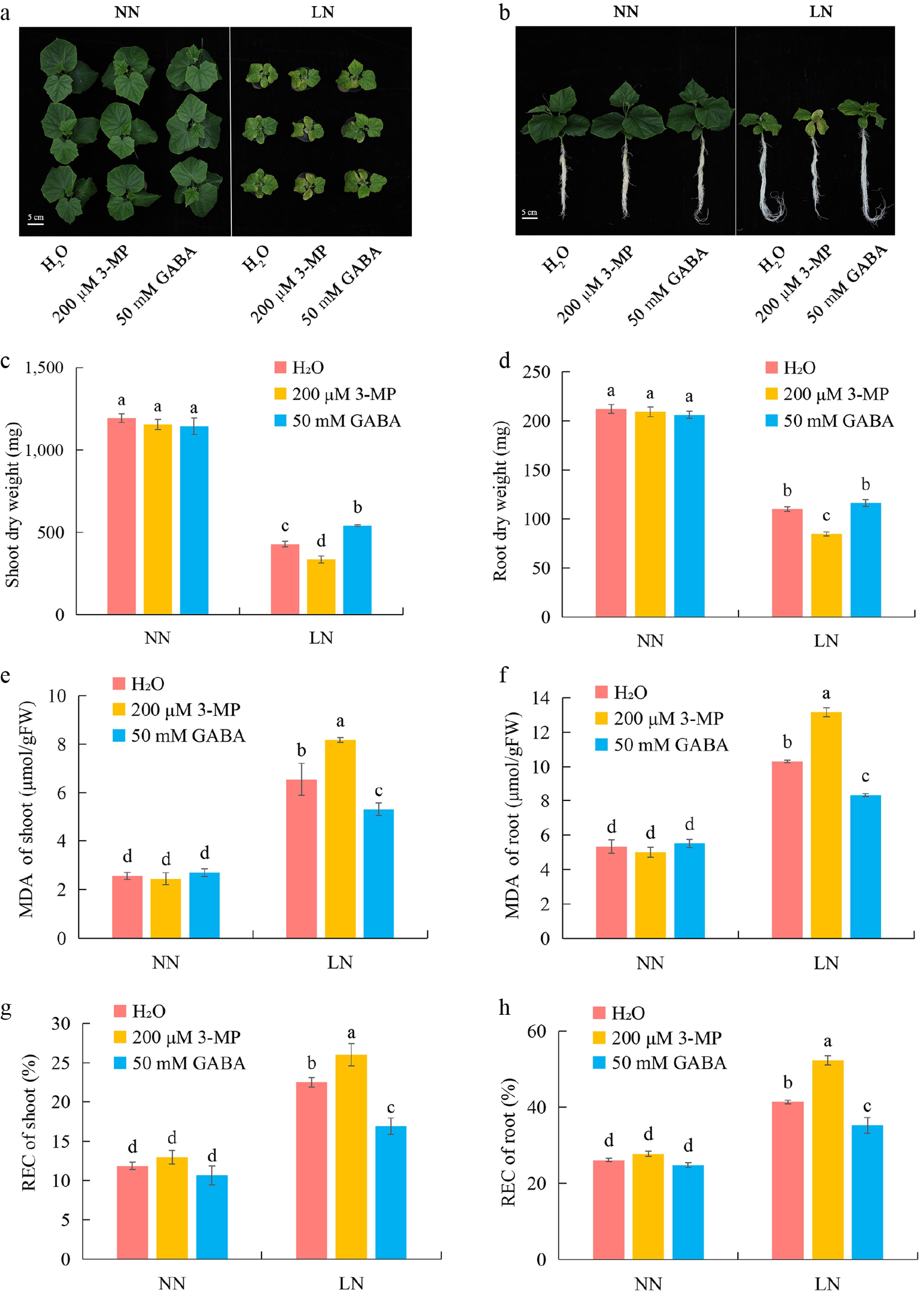
Figure 6.
Exogenous GABA application enhances LN tolerance of cucumber accession 9930. (a), (b) Representative shoot and root phenotypes of accession 9930 treated with water (H2O), 200 μM 3-MP, or 50 mM GABA under NN and LN conditions. (c), (d) Shoot and root dry weights under NN and LN conditions after the indicated treatments. (e), (f) MDA content in shoots and roots under NN and LN conditions. (g), (h) REC in shoots and roots under NN and LN conditions. Data are presented as mean ± SD. Different letters indicate significant differences (p < 0.05).
-
Nitrogen is a fundamental macronutrient governing plant growth, development, and yield formation. Since the 1960s, intensive N fertilization has substantially boosted global crop productivity[5,9], but has also resulted in excessive N dependence, limited availability of high-NUE cultivars with strong LN tolerance, and serious environmental contamination of soil and water systems[35]. Improving NUE in crops is therefore a central goal of sustainable agriculture. Current studies indicated NUE is determined by the coordination of multiple physiological processes, including root system architecture[18,19], N uptake and transport[36], N remobilization[24], and N assimilation[25]. However, most existing evaluation strategies focus on individual traits, making it difficult to capture integrated whole-plant responses to heterogeneous N availability. In natural soils, N distribution is spatially uneven, which strongly influences root developmental plasticity and N acquisition strategies, giving rise to N foraging behavior[19]. Plants perceive this heterogeneity through SNS, which integrates local N status with long-distance shoot–root communication to preferentially promote root growth and N uptake in N-rich zones while restraining growth in N-poor regions[22,37]. Despite the recognized importance of SNS, phenotypic indicators directly reflecting SNS strength have rarely been exploited to evaluate crop LN adaptation or NUE. In this study, we established a split-root system in cucumber and quantified SNS responsiveness using the SLN/SHN root length ratio. Notably, SNS responsiveness displayed a broad and continuous distribution across the germplasm panel rather than forming discrete categories (Fig. 1g), and clustering analysis further resolved multiple SNS response groups (Fig. 1h), underscoring extensive natural diversity among cucumber accessions. Within this population framework, G47 shows the strongest SNS responsiveness and exhibits superior LN tolerance (Figs. 1, 3), supporting the use of SNS-related phenotypes as indicators of LN adaptation and NUE-related performance in cucumber.
Coupling of SNS responsiveness and nitrate uptake as a germplasm-level indicator of LN adaptation and NUE
-
Intensive N fertilization is widely used in cucumber production, yet it is often associated with poor NUE and substantial environmental burdens, highlighting the need to develop germplasm with improved LN tolerance and high NUE. Because SNS biases root growth towards NO3−-rich zones, and nitrate uptake capacity determines the rate of N capture, these two traits together provide a practical framework for prioritizing germplasm with potential NUE relevance. Under this framework, identifying accessions combining strong SNS responsiveness with high nitrate uptake capacity represents a logical route for selecting germplasm with improved LN adaptation and NUE potential. Accordingly, in this study, we integrated SNS phenotyping with 15N-labelled nitrate uptake assays and identified G47 as a genotype exhibiting both strong SNS responsiveness and high nitrate uptake capacity (Figs. 1, 2). Importantly, subsequent LN stress assays demonstrated that G47, characterized by strong SNS responsiveness and high nitrate uptake capacity, displayed superior LN tolerance relative to G23 (CG4353; weak SNS responsiveness and low nitrate uptake capacity) and the reference cultivar 9930 (strong SNS responsiveness but lower nitrate uptake capacity, Fig. 3).
Together, these results support the use of combined SNS responsiveness and nitrate uptake capacity as germplasm-level indicators for prioritizing cucumber accessions with enhanced LN adaptation and NUE potential.
Notably, this dual-trait profile was not unique to G47. Several accessions displayed similarly strong SNS responsiveness, including CG1601, CG5797, CG5801, and CG6578 (Fig. 1g, h; Supplementary Table S2) and also ranked among the higher-uptake accessions in the 15N tracer assay (Fig. 2a, b; Supplementary Table S3). These accessions, therefore, represent valuable genetic resources for future work to test how SNS phenotypes and nitrate uptake capacity jointly contribute to LN adaptation and, ultimately, NUE improvement in cucumber.
GABA enhances cucumber tolerance to low-nitrogen stress
-
γ-Aminobutyric acid (GABA) is a ubiquitous non-protein amino acid that accumulates in plants under diverse abiotic stresses and is widely implicated in stress mitigation through the modulation of redox homeostasis and antioxidant capacity[38,39]. Consistent with emerging links between GABA metabolism and LN stress[40], our transcriptome data provide genotype-dependent evidence that LN-tolerant germplasm preferentially activates GABA-associated N metabolism. In particular, a glutamate dehydrogenase gene (CsaV3_4G013270) was induced in G47, but repressed in the LN-sensitive cultivar 9930, and CsGS1 (CsaV3_3G021030), which supports glutamate supply for downstream GABA biosynthesis, showed strong LN induction in G47 but minimal induction in G23 and 9930 (Fig. 5g).
Importantly, this transcriptional signature is complemented by functional pharmacology: exogenous GABA alleviated LN-induced growth inhibition and stress-associated physiological changes in 9930, whereas the GABA biosynthesis inhibitor 3-MP exacerbated LN stress symptoms (Fig. 6). The negligible effects of either treatment under normal N conditions further suggest that GABA primarily modulates LN stress responsiveness rather than basal growth. Collectively, these lines of evidence support GABA as a stress-responsive metabolic modulator associated with LN tolerance, and are consistent with the hypothesis that enhanced GABA-associated metabolic capacity contributes to improved resilience under nitrogen limitation.
Nevertheless, the current evidence does not resolve the causal and mechanistic basis of this association. Although CsGS1 displays genotype-specific LN induction, expression differences alone cannot determine whether cis-regulatory or coding variation contributes to the observed transcriptional divergence or to LN tolerance. Future work integrating endogenous metabolite profiling, genetic co-segregation analysis in segregating populations (e.g., G47 × G23), promoter–reporter assays, and ultimately, transgenic or genome-editing validation will be required to dissect how GABA metabolism is genetically controlled and how it interfaces with LN adaptation pathways in cucumber. It will also be valuable to determine whether GABA acts solely downstream as a stress-mitigating metabolite or whether it feeds back on SNS and NO3− uptake capacity. Overall, our study provides complementary physiological and transcriptomic evidence implicating GABA as a metabolic modulator associated with LN adaptation, while the underlying molecular mechanisms remain to be established.
-
In this study, we established a hydroponic split-root system combined with 15N labeling to systematically evaluate the SNS responsiveness and nitrate uptake capacity across 56 cucumber germplasm materials. Substantial natural variation was observed, and two contrasting accessions—G47 (high SNS and uptake) and G23 (low SNS and uptake)—were selected together with the reference cultivar 9930. Subsequent LN stress tests also revealed that signaling-sensitive germplasm G47 exhibited superior LN tolerance. Further transcriptome analysis indicated significant activation of the γ-aminobutyric acid (GABA) metabolic pathway in G47, with marked upregulation of the key gene CsGS1. Exogenous application of GABA enhanced plant tolerance to LN stress, indicating a crucial regulatory role for GABA in N use efficiency in cucumber. In conclusion, these findings identify GABA as a metabolic modulator of LN adaptation in cucumber and establish an integrated framework—driven by SNS responsiveness and nitrate uptake capacity—for the strategic discovery of high nitrogen use efficiency germplasm and the identification of molecular targets for nutrient efficient breeding.
This work was financially supported by the National Natural Science Foundation of China (Grant Nos 32502693 and 32272704), and the Guangdong Provincial Special Fund for Modern Agriculture Industry Technology Innovation Teams (2024CXTD01). We would like to express our sincere gratitude to Dr. Yang Xueyong from the Chinese Academy of Agricultural Sciences for his strong support. The experimental materials he provided have played an important role in our research work. We are grateful to Dr. Kong Qiusheng from the Huazhong Agricultural University for his guidance on our article.
-
The authors confirm their contributions to the paper as follows: study conception and design, experiment performance, data analysis: Wang W, Ye R; manuscript preparation: Sun C, Wang Y; supervision, resources, funding acquisition: Yang L, Cao H; experimental and writing participation guidance: Ma M, Geng S, Luo C, Zhen S, Yan X, Bie Z. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/vegres-0026-0008.
-
# Authors contributed equally: Chenyu Sun, Yaqi Wang, Runwen Ye, Wenbo Wang
- Supplementary Table S1 Composition of nutrient solutions used in hydroponic experiments.
- Supplementary Table S2 The SLN/SHN ratio of 56 cucumber germplasm.
- Supplementary Table S3 Statistical results of NO3− uptake capacity in 56 cucumber germplasms.
- Supplementary Fig. S1 Effects of different concentrations of GABA and its inhibitor 3-mercaptopropionic acid (3-MP) on cucumber accession 9930 under low-nitrogen (LN) conditions.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Sun C, Wang Y, Ye R, Wang W, Luo C, et al. 2026. Comparative analysis of cucumber germplasm identifies GABA as a modulator of low-nitrogen tolerance. Vegetable Research 6: e015 doi: 10.48130/vegres-0026-0008 

Comparative analysis of cucumber germplasm identifies GABA as a modulator of low-nitrogen tolerance
- Received: 10 November 2025
- Revised: 01 February 2026
- Accepted: 11 February 2026
- Published online: 30 April 2026
Abstract: Excessive nitrogen (N) fertilization in cucumber cultivation reduces nitrogen-use efficiency (NUE) and imposes severe environmental costs. Improving NUE requires coordinated regulation of systemic nitrogen signaling (SNS), which directs root foraging toward nitrate (NO3−)-rich zones, and nitrate uptake capacity, which defines the rate of N acquisition. Here, we established a hydroponic split-root assay and a 15N-nitrate tracing system to quantify SNS responsiveness and nitrate uptake capacity across 56 cucumber (Cucumis sativus L.) germplasms. Significant variation was identified across the germplasm, resulting in the selection of G47 (high SNS responsiveness and nitrate uptake capacity), and G23 (low SNS responsiveness and nitrate uptake capacity), as contrasting accessions for evaluation alongside the reference cultivar 9930. Comparative analysis of these three genotypes revealed that G47 exhibited superior low-nitrogen (LN) stress tolerance, showing the smallest declines in biomass and chlorophyll, incurring the lowest oxidative and membrane damage, and demonstrating greater osmolyte and antioxidant accumulation alongside sustained nitrate assimilation. Comparative transcriptomic analysis revealed strong induction of γ-aminobutyric acid (GABA)-associated metabolism in G47, with the GABA pathway gene CsGS1 markedly upregulated. Functionally, foliar GABA application enhanced, whereas the GABA-biosynthesis inhibitor 3-mercaptopropionic acid (3-MP) diminished, LN tolerance. These findings identify GABA as a metabolic modulator of LN adaptation in cucumber and establish an integrated framework based on SNS responsiveness and nitrate uptake capacity for discovering high-NUE germplasm and molecular targets for nutrient-efficient breeding.
-
Key words:
- Cucumber /
- Systemic nitrogen signaling /
- Nitrate uptake capacity /
- GABA /
- Low-nitrogen tolerance