










-
As sessile organisms, plants frequently experience different types of stress, including salinity, low temperatures, and infections from pathogenic bacteria[1]. Soil salinization is one of the main problems that damage cultivated land, this leads to stunted plant growth and decreased yield[2]. About 20% of irrigated land around the globe suffers from salinization, significantly affecting productivity[3]. Salinity can cause osmotic and toxic stress and affect the growth and metabolism of plants. Simultaneously, halophytes have developed defense mechanisms against salt stress, including salt rejection and dilution[4,5]. The ability of plants to tolerate salt is a multifaceted characteristic shaped by genetic factors, physiological conditions, and environmental elements. Grasping how plants tolerate salt is crucial for enhancing the breeding of plant varieties that can withstand saline-alkali soil.
Long non-coding RNAs, commonly referred to as lncRNAs, represent a significant class of RNA molecules found in eukaryotic organisms. These RNA molecules are distinguished by their relatively large size, typically exceeding 200 nucleotides in length. LncRNAs have no clear potential to encode proteins, which makes them different in cell function[6−8]. Most of them are synthesized by RNA polymerase II, which plays an important role in the transcription process. In addition, RNA polymerase I and RNA polymerase III also have a certain contribution, but to a lesser extent[9]. According to their position in the coding gene, lncRNAs are divided into four categories: sense, antisense, intronic, and intergenic[10]. Initially, lncRNAs were dismissed as mere transcriptional noise[11]. The subsequent demonstration of their involvement in both transcriptional and post-transcriptional regulation was achieved[12]. Improvements in sequencing technologies have resulted in the discovery of many lncRNAs within eukaryotic organisms[13−17].
In plants, lncRNAs have been recognized as essential to play a critical regulatory role in reproductive growth, photomorphogenesis, the timing of flowering, and the plant's reaction to both abiotic and biotic stressors[18−24]. At present, the role of lncRNAs in plants remains relatively underexplored, with only a limited number of these molecules having been thoroughly analyzed. Recent studies have shown that lncRNAs have the ability to react to different stress factors, such as salt stress[25−28]. A relevant example can be seen in lncRNA LAL found in Medicago truncatula, which is instrumental in promoting salinity tolerance. This lncRNA achieves its effect by modulating the expression of a specific gene, MtLHCB1, thereby contributing to the plant's ability to withstand high salinity levels. The findings underscore the vital roles that lncRNAs play in the stress response pathways of plants, highlighting their significance in bolstering resilience against both biotic and abiotic stresses[29]. lncRNA TRABA promotes salt tolerance in cotton by inhibiting the expression of β-glucosidase BGLU24[30]. lncRNA BplncSIR1 enhances the salt tolerance of Betula platyphylla by regulating BpNAC2[28]. MtCIR1, a lncRNA from Medicago truncatula, has been shown to be more sensitive to salt stress by inhibiting CYP707A2 expression and enhancing ABA accumulation[31]. The lncRNA354 of upland cotton can act as a competitive endogenous RNA of miR160b, and then regulate the ARF gene in response to salt stress[32]. In summary, research suggests that lncRNAs significantly contribute to the regulation of salt tolerance in plants.
Mulberry (Morus L.) is a kind of perennial woody plant with high economic value and ecological functions. The mulberry fruit is rich in nutrition. Mulberry is recognized as an important genetic resource for its ability to withstand both drought and salt stress[33]. Earlier studies focused on the mechanism of salt tolerance in mulberry. However, lncRNAs involved in salt stress response in mulberry have not been systematically studied. The advancement of transcriptome sequencing technology, along with the development of related bioinformatics techniques, has facilitated the comprehensive discovery of novel lncRNAs. In this study, the primary objective was to identify genes and lncRNAs that exhibit a response to salt stress in the root tissues of mulberry (Morus notabilis). Through analysis, genes linked to salt stress were pinpointed, along with the functional characterization of some salt-responsive lncRNAs, resulting in the identification of lncRNAs linked to salt tolerance. Notably, a low salt-tolerant lncRNA was specifically identified. These results offer valuable perspectives on the function of lncRNAs in salt tolerance and may enhance our comprehension of their roles within this framework.
-
In this study, the seeds of M. notabilis were soaked in a 10% NaClO solution for 10 min, and then they were left to soak in distilled water until the seeds germinated. Under conditions of 16 h of light and 8 h of darkness, the temperature was maintained at 25 °C during the day, providing optimal warmth for growth and development, while the nighttime temperature was slightly cooler at 22 °C, with an air humidity level of 75%, the mulberry seedlings were transferred to a substrate for one month. Subsequently, they were exposed to 200 mM NaCl for one day to induce salt stress. The Control group was watered normally. Mulberry roots were collected for later analysis experiments.
Identification of lncRNAs
-
RNA extraction was conducted using a Trizol kit (Invitrogen, Carlsbad, CA, USA) according to the instructions given by the manufacturer. Mulberry genome Mnot-SWU downloaded from MorusDB (
https://morus.swu.edu.cn )[34]. Sequencing was conducted using the Illumina HiSeq™ 4000 platform at Gene Denovo Biotech Co. (Guangzhou, China). The software CNCI (version 2)[35], CPC (version 0.9-r2)[36], and FEELNC (version v0.2)[37] were used with default parameters to assess the protein-coding potential of the new transcript. Non-protein-coding intersections are selected as long non-coding RNAs.Determination of physiological changes
-
The total antioxidant capacity was determined by referring to previous methods[38]. A precise amount of 0.1 g of root tissue was carefully weighed out. Following this, 1 mL of a pre-cooled extract was introduced to the root tissue. To ensure thorough mixing, the combination was homogenized while submerged in an ice bath, which is crucial for maintaining sample integrity and preventing the degradation of sensitive compounds. After homogenization, the mixture underwent centrifugation at a high speed of 10,000 revolutions per minute (rpm) for 10 min at a temperature of 4 °C. This centrifugation process facilitated the separation of the sample components, allowing the supernatant to be collected. Finally, the supernatant was retained on ice, preparing it for subsequent measurements. Absorbance was measured at a wavelength of 593 nm.
The level of malondialdehyde (MDA) was assessed using the thiobarbituric acid (TBA) assay[39]. 0.1 g root tissue was weighed, 1 mL extract solution added for grinding, centrifuging at 12,000 rpm for 10 min. The supernatant was added into 2 mL 0.6% TBA, shaken well, and the obtained solution was placed at 100 °C for 60 min, cooled and centrifuged. The measurement of the absorption supernatant was conducted using a spectrophotometer. The analysis involved the use of two distinct wavelengths: one at 532 nm and another at 600 nm.
The proline concentration was assessed through a ninhydrin reaction. About 0.1g of root tissue was weighed and ground, 1 mL of tissue extraction solution was added, and then extracted in boiling water for 10 min, then centrifuged for 10 min, and the supernatant retained for analysis. Then, acidic ninhydrin solution and acetic acid were added to the supernatant in proportion. After the mixing process, the solution was subjected to boiling for 30 min, with shaking occurring every 10 min, and then allowed to cool to room temperature. The absorbance was recorded at a wavelength of 520 nm.
The staining methods using nitroblue tetrazolium (NBT) and 3'-diaminobenzidine (DAB) were carried out as described in previous literature[40]. The activity levels of peroxidase (POD) and ascorbate peroxidase (APX) were evaluated following the manufacturer's instructions through a plant detection kit (Solarbio, Beijing, China).
Differentially expressed transcripts analysis
-
The analysis focused on the differential expression of coding RNA and long non-coding RNA (lncRNA) transcripts. For this purpose, DESeq2[41] software was employed to perform a comprehensive evaluation. In this analysis, genes or transcripts that exhibit a false discovery rate (FDR) of less than 0.05, along with an absolute fold change of a minimum of 2, are categorized as differentially expressed genes or transcripts. Subsequently, an analysis of GO functions and KEGG pathway enrichment was conducted on the coding RNAs that were expressed differentially. The 'clusterProfiler' R package was used to analyze the GO and KEGG pathways to further understand the main biological functions of DEGs. The p < 0.05 was the threshold of significance.
qRT-PCR validation
-
RNA was extracted from the samples employing the TRIzol reagent (Invitrogen, Carlsbad, CA, USA). The synthesis of first-strand cDNA was carried out with the PrimeScript™ RT-PCR Kit (TaKaRa, Kusatsu, Shiga, Japan). The expression levels were measured using the SYBR Green detection system in a StepOnePlus™ Real-Time PCR System (Applied Biosystems, CA, USA). The MnActin gene was used as the internal control of mulberry. The data presented in this paper are based on three biological replicates. More information about the qRT-PCR primers can be found in Supplementary Table S1.
Dual-luciferase assays
-
The Chr1G00048820.1 promoter was cloned into the vector to drive the expression of luciferase (LUC) reporter gene and generate proMnIAA27:LUC vector. To assess the inhibitory effect of lnc15780 on the MnIAA27 gene, we injected Agrobacterium cells containing proMnIAA27:LUC gene into tobacco leaves using a needle-free syringe. The isolated leaves were sprayed with 1 mm of fluorescein. Luciferase activity was measured by live cell imaging.
Chromatin isolation by RNA purification (ChIRP)
-
A collection of probes intended for ChIRP was created following the target sequence, with each probe positioned roughly 100−150 bp apart. Five distinct probes were developed and numbered from 1 to 5. An antisense DNA probe, featuring a Biotin-TEG label at its 3' terminus, was synthesized. The five probes were categorized into two groups based on their respective locations. The 'Odd' group, which corresponds to the forward DNA strand, is made up of probes labeled 1, 3, and 5. In contrast, the 'Even' group, which aligns with the reverse DNA strand, contains probes numbered 2 and 4. To ensure the reliability of the results, three biological replicates of the experiment were performed. Additionally, the LacZ cDNA probe was employed as a negative control for each DNA sample analyzed via qPCR.
Generation of transgenic mulberry
-
The lncRNA and gene coding sequences were first amplified. Following the purification of the PCR products, the next step involved the cloning of these purified fragments into the pLGNL vector. This process utilized the in-fusion cloning system, which facilitated the assembly of a recombinant expression vector designed for further experimental applications[42]. Once the recombinant vector was successfully constructed, it was introduced into the Agrobacterium rhizogenes strain K599. The transformed Agrobacterium was then injected into the junction between the true leaf and cotyledon of four-week-old plants. After this injection, the plants were nurtured in the soil for seven weeks, allowing for adequate growth and development of the modified plant structures.
Histochemical GUS staining
-
For GUS staining, the roots were soaked in 50 mM sodium phosphate buffer (pH 7.0), 5 mM K3Fe(CN)6, 5 mM K4Fe(CN)6, 0.1% Triton X-100, 20% (v/v) MeOH, and 1 mM X-Gluc. A 12-h staining treatment was carried out at 37 °C in the dark, followed by alcohol elution.
Statistical analyses
-
SPSS 26.0 statistical software was used for significance analysis of all data, and student's t-test and one-way ANOVA were used. The results are shown as the average ± standard error. A p-value below 0.05 was considered statistically significant.
-
To verify the effects of salt treatment on the phenotype and physiology of mulberry seedlings, 200 mM NaCl was applied for 24 h, and it was found that mulberry leaves had obvious wilting and drooping, and their color turned pale green, with a slightly shriveled texture (Fig. 1a). In addition, MDA content, total antioxidant capacity, and proline content were also determined (Fig. 1b−d). The results showed that the contents of MDA and proline in the roots of mulberry seedlings treated with salt stress were significantly increased compared with those not subjected to salt stress. At the same time, the total antioxidant capacity in the roots was significantly reduced. The findings indicate that salt induces various important physiological alterations.
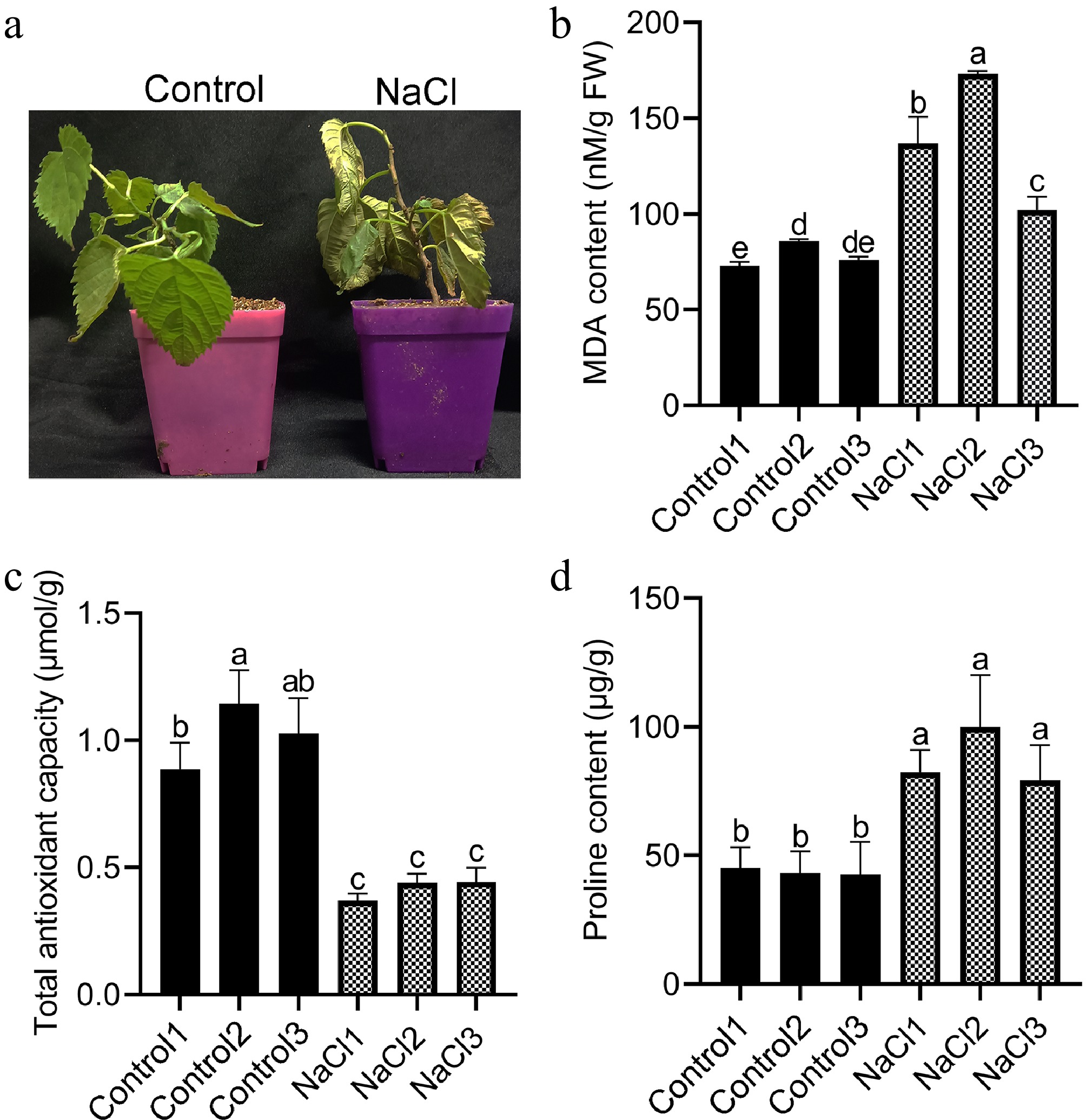
Figure 1.
Phenotype and physiological characteristics of mulberry under control and salt treatment. (a) Phenotypes of mulberry plants under control and salt treatment. (b) MDA content of mulberry roots. (c) Total antioxidant capacity of mulberry roots. (d) Proline content of mulberry roots. Different letters indicated significant differences (p < 0.05).
RNA-seq analysis
-
The root libraries of mulberry were constructed under normal conditions (Control) and salt treatment for 24 h (NaCl). A clean dataset totaling 7.79 billion entries was produced, with every sample achieving a Q30 score of at least 90.89%, which indicates a base call accuracy of 99.9%. The GC content ranged from 49.19% to 49.84%. This detailed dataset underscored that the quality of high-throughput sequencing is adequate to satisfy the stringent requirements for lncRNA analysis, as illustrated in Supplementary Table S2. Furthermore, a principal component analysis (PCA) was conducted on the data pertaining to both lncRNAs and mRNAs, which is visually represented in Supplementary Fig. S1. On the PC1 × PC2 scoring plots, the two groups (Control and NaCl) were significantly separated. In addition, the expression violin plot of the samples showed that the difference in lncRNAs expression abundance was relatively small, while the difference in mRNAs expression abundance was relatively large. Most lncRNAs have 1 or 2 exons (Fig. 2a). Among them, the lncRNAs of exon 1 accounted for 93.83% of the total number of identified lncRNAs, and the lncRNAs of exon 2 accounted for 5.02% of the total number of identified lncRNAs. In contrast, mRNAs generally have more exons, with 34.05% of mRNAs having more than 2 exons. These differences in exon numbers may indicate that different functions between lncRNAs and mRNAs may be associated with them. In addition, the average length of lncRNA transcripts is shorter than that of mRNAs (Fig. 2b). Specifically, 78.75% of lncRNAs were no longer than 1,500 bp, and 40.91% of mRNAs was longer than 1,500 bp.
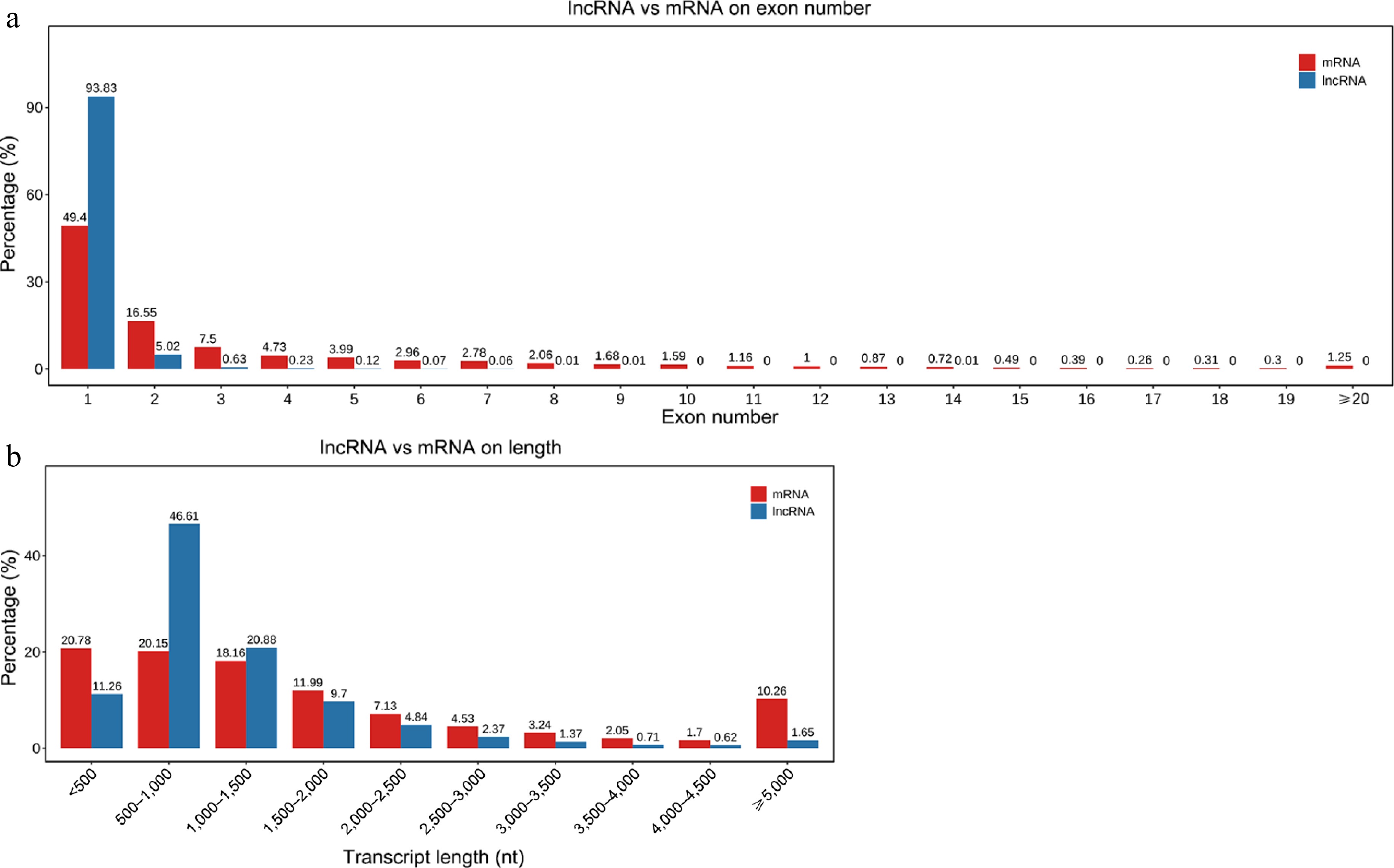
Figure 2.
Comparison of structural characteristics of lncRNAs and mRNAs. (a) Exon distribution in lncRNAs and mRNAs. (b) Length distribution of lncRNAs and mRNAs.
Analysis of differential expression for lncRNAs and mRNAs
-
To examine the transcriptional changes of lncRNAs and mRNAs under salt stress, we conducted a comprehensive analysis. lncRNAs that were differentially expressed (DE-lncRNAs) along with genes that exhibited differential expression (DEGs) were identified. A total of 1,201 lncRNAs with differential expression characteristics and 6,330 genes with differential expression were identified (Fig. 3a, b). In the NaCl group compared to the Mock group, 714 lncRNAs were up-regulated and 487 were down-regulated, while 4,557 genes were up-regulated and 1,773 were down-regulated. In the environment of salt treatment, the expression of genes and lncRNAs in plants showed a significant change trend. To show this change more intuitively and systematically, heat map clustering analysis was carried out (Fig. 3c, d). Plant hormones are signaling compounds that regulate key aspects of environmental stress responses. Additionally, a heat map analysis focusing on plant hormone signal transduction pathway genes within the DEGs was conducted (Supplementary Fig. S2), showing significant up-regulation of most pathway genes under salt stress.
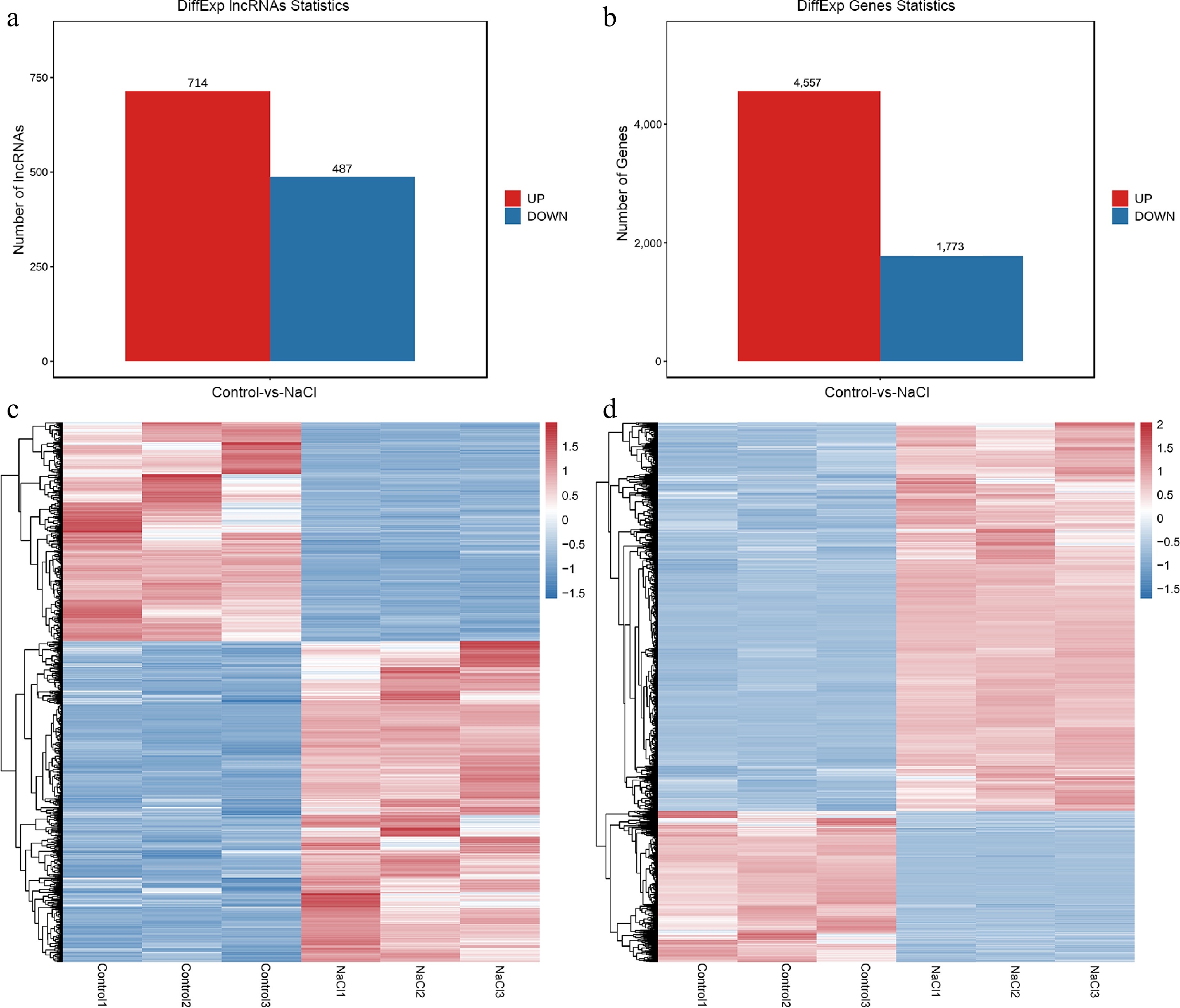
Figure 3.
Overview of DE-lncRNAs and DEGs in mulberry under control and salt treatment. Number of (a) DE-lncRNAs and (b) DEGs. Hierarchical clustering heat maps of all (c) DE-lncRNAs and (d) DEGs.
DE-lncRNAs target gene enrichment analysis
-
To explore the potential biological role of lncRNAs in the salt stress response of mulberry, we used cis-, antisense-, and trans-acting methods to identify DE-lncRNAs fine-tuned target genes. The GO classification showed that the main types of cis-target genes and trans-target genes were responses to stimulus, and the main type of antisense-target gene was alpha-amino acid catabolic process (Fig. 4a−c). These findings indicate that lncRNAs may play a role in salt stress response by promoting the expression of genes related to resistance and metabolism. KEGG pathway anaylsis showed that the main types of cis-, antisense-, and trans-target genes were metabolic pathways (Fig. 4d, e). The findings indicate that lncRNAs may contribute to salt stress response primarily by upregulating metabolism-related gene expression.
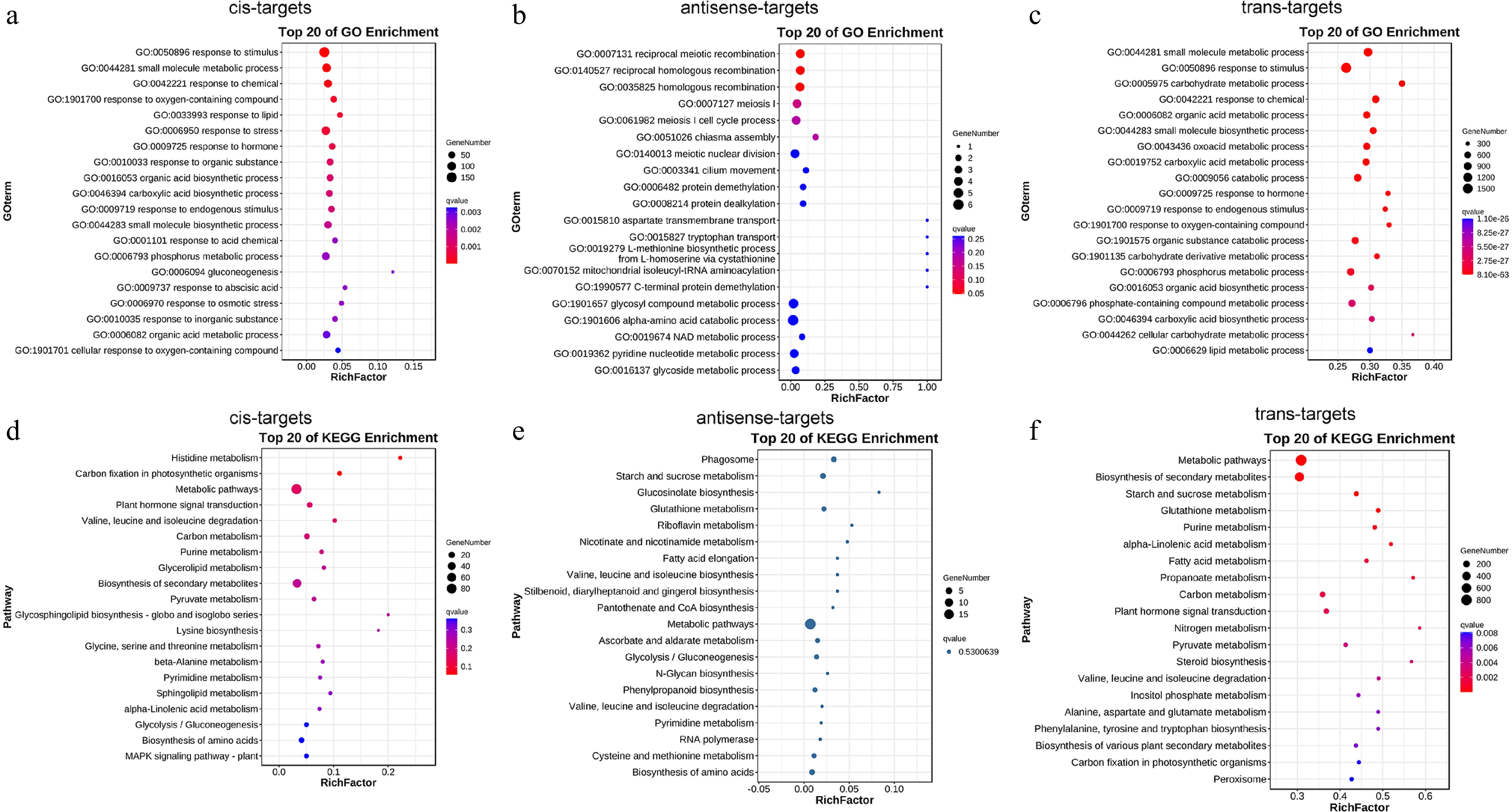
Figure 4.
Functional annotation of DE-lncRNAs target genes in roots by GO classification and KEGG enrichment. The top 20 enriched the GO classification of DE-lncRNAs (a) cis-, (b) antisense-, and (c) trans-target genes. Top 20 enriched KEGG pathways of DE-lncRNAs (d) cis-, (e) antisense-, and (f) trans-target genes.
Validation of salt-reactive lncRNAs and target genes
-
qRT-PCR was employed to measure the expression levels of five lncRNAs and their potential target genes in mulberry seedlings under salt stress (Fig. 5). After salt treatment, the expression levels of all five target genes detected in the study were significantly increased. In addition, the expression levels of various lncRNAs have also experienced significant fluctuations; specifically, some lncRNAs showed marked increases in expression, while others demonstrated significant decreases. This pattern of expression changes implies that lncRNAs potentially play a regulatory role in the modulation of target gene expression, employing varied mechanisms of action to influence these genetic processes. The qRT-PCR results were consistent with the omics data (Supplementary Table S3), suggesting that these differentially expressed lncRNAs and target genes may be associated with salt stress resistance in mulberry. To further explore their role in salt tolerance, lncRNA with inhibitory effects and target genes with potential stress resistance were focused on. Based on this, lnc15780 (MSTRG.15780.1) and its target gene MnIAA27 (Chr1G00048820) were analyzed in depth.
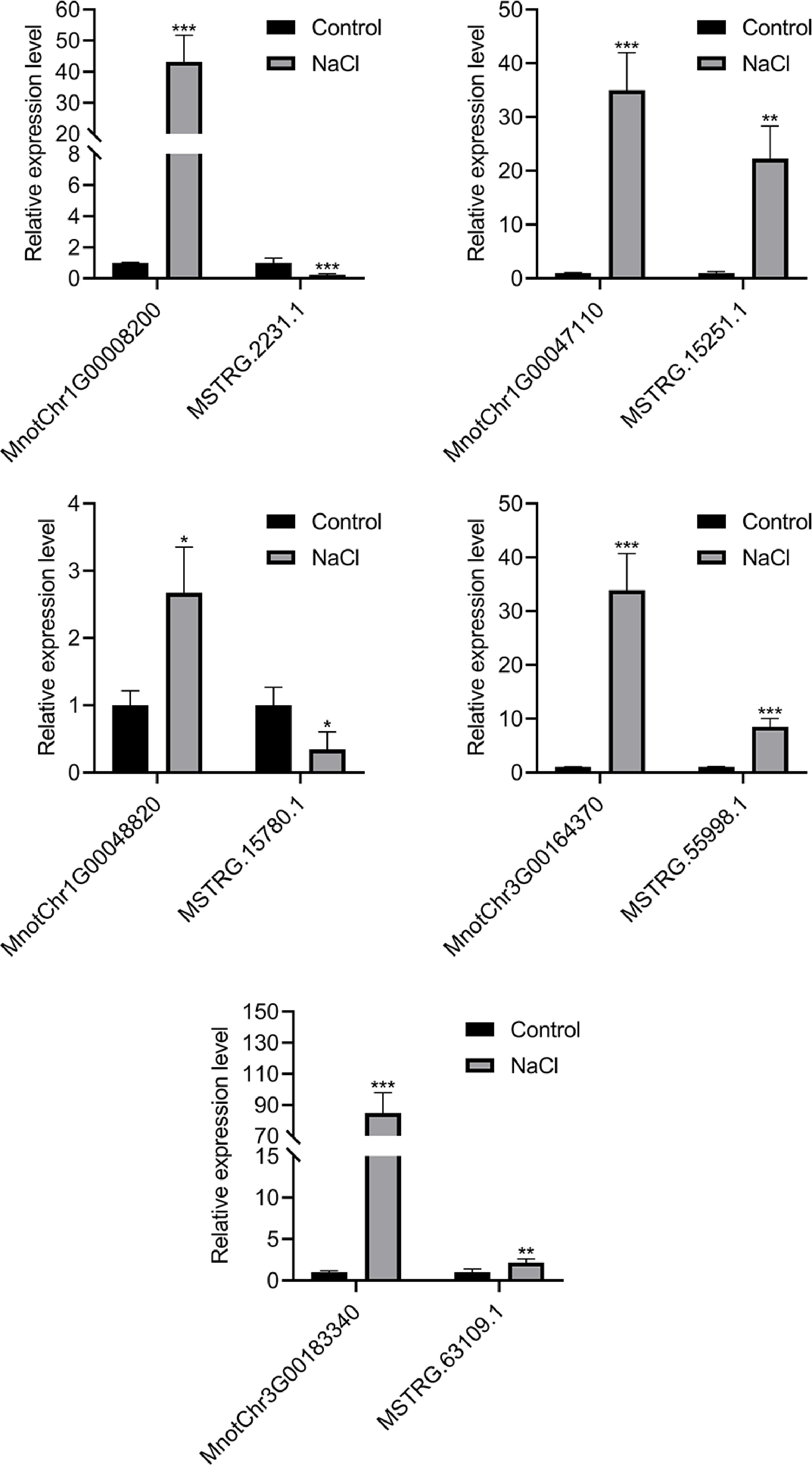
Figure 5.
The relative expression of five pairs of differentially expressed lncRNAs and potential target genes in mulberry roots of control group and salt treatment group. Error bars represent standard deviation, n = 3. The asterisks indicate a statistically significant difference (* p < 0.05; ** p < 0.01; *** p < 0.001; two-tailed t-test).
MnIAA27 is a direct target gene of lnc15780
-
To explore the interaction between lnc15780 and the MnIAA27 promoter, effector (lnc15780-OE), and reporter (LUC driven by the MnIAA27 promoter) vectors were constructed and transiently co-transformed into tobacco leaves. A dual luciferase activity assay was then performed. LUC activity analysis revealed that lnc15780 binds to the MnIAA27 promoter, inhibiting its expression (Fig. 6a, b). These findings suggest that lnc15780 interacts with the MnIAA27 promoter. To further investigate whether lnc15780 directly regulates MnIAA27, ChIRP was performed (Fig. 6c). The promoter region, extending 2,000 bp upstream, was segmented into five equal parts for ChIRP-qPCR amplification. The findings indicated that the truncated promoters of MnIAA27 exhibited significant enrichment through ChIRP, irrespective of the usage of 'Even' or 'Odd' probes. This implies that the regulation of MnIAA27 occurs via its interaction with lnc15780.
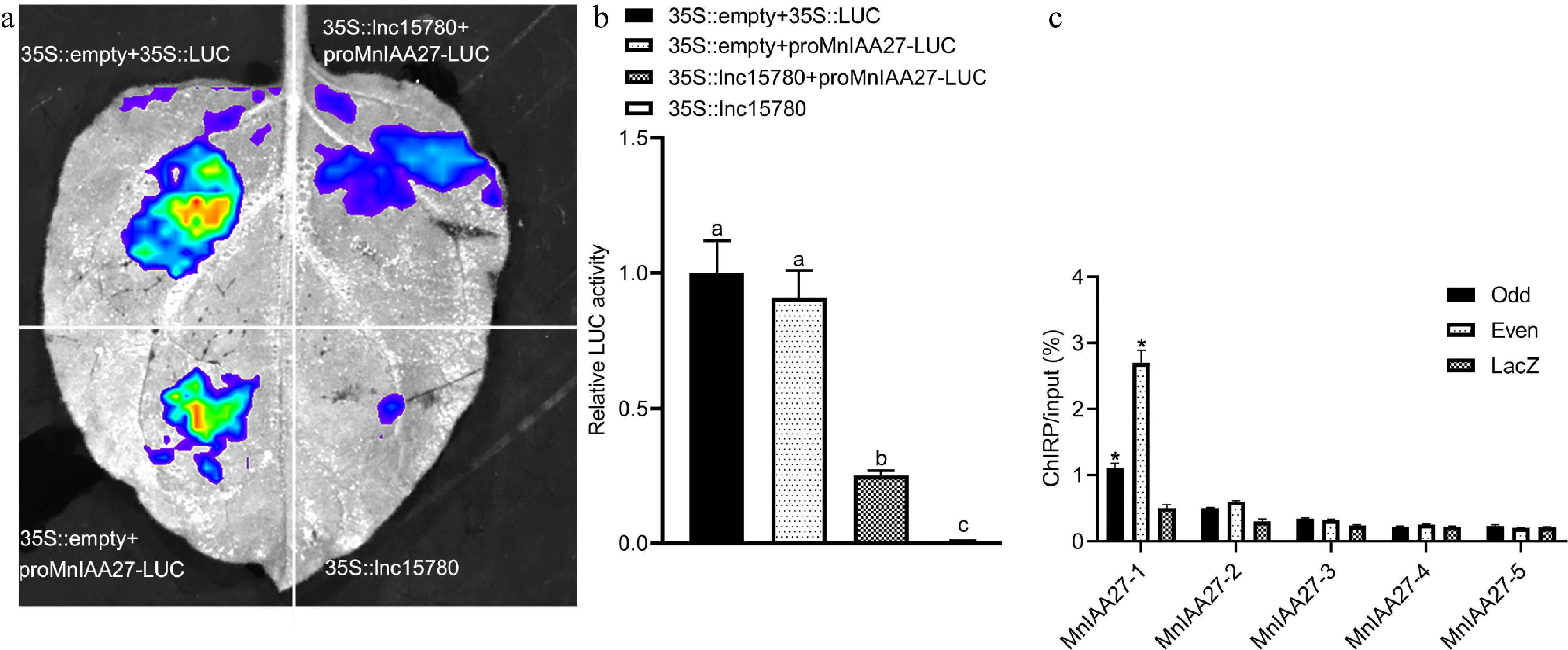
Figure 6.
The mulberry lnc15780 regulates MnIAA27 expression. (a) Transient expression determination of LUC in tobacco leaves. The vector was transformed into tobacco leaves and the fluorescence intensity was observed using the PlantView imaging system. When the transcription factor inhibited promoter activity, the fluorescence value decreased. (b) Quantitative analysis of LUC activity. The error bar represents the standard deviation. According to Tukey's HSD test, different letters indicated significant differences (p < 0.05). (c) ChIRP analysis of lnc15780 binding to MnIAA27 promoter. Odd, samples hybridized with odd probes; Even, samples hybridized with even probes; Lac, LacZ-DNA probes were used as negative controls. The asterisks indicate a statistically significant difference (* p < 0.05).
The function of lnc15780 and MnIAA27 was further studied by using a transgenic hairy root system. The histochemical analysis conducted using GUS revealed a significant level of GUS staining in the roots of mulberry seedlings (Fig. 7a). This observation underscores the effectiveness of the transgenic hairy root expression system utilized in mulberry. Transcriptional levels of lnc15780 or MnIAA27 were evaluated in the control group and the lnc15780 or MnIAA27 overexpression group (Fig. 7b, c). In the lnc15780 overexpression group, we observed significantly increased levels of lnc15780 transcripts. Similarly, in the MnIAA27 overexpression group, the expression level of MnIAA27 was significantly increased. To evaluate the resistance of mulberry trees to salt stress after lnc15780 and MnIAA27 overexpression, mulberry trees in the control group, lnc15780 and MnIAA27 overexpression group were subjected to salt stress for 12 h respectively (Fig. 7d). Under salt stress, the growth of MnIAA27 strain was the best, CK (empty vectors) was the second, and lnc15780 strain was the last. This indicates that MnIAA27 can enhance the salt tolerance of mulberry, while lnc15780 can reduce it. In addition, the expression of MnIAA27 was significantly down-regulated after overexpression of lnc15780 (Supplementary Fig. S3), further indicating that MnIAA27 is a target gene of lnc15780.
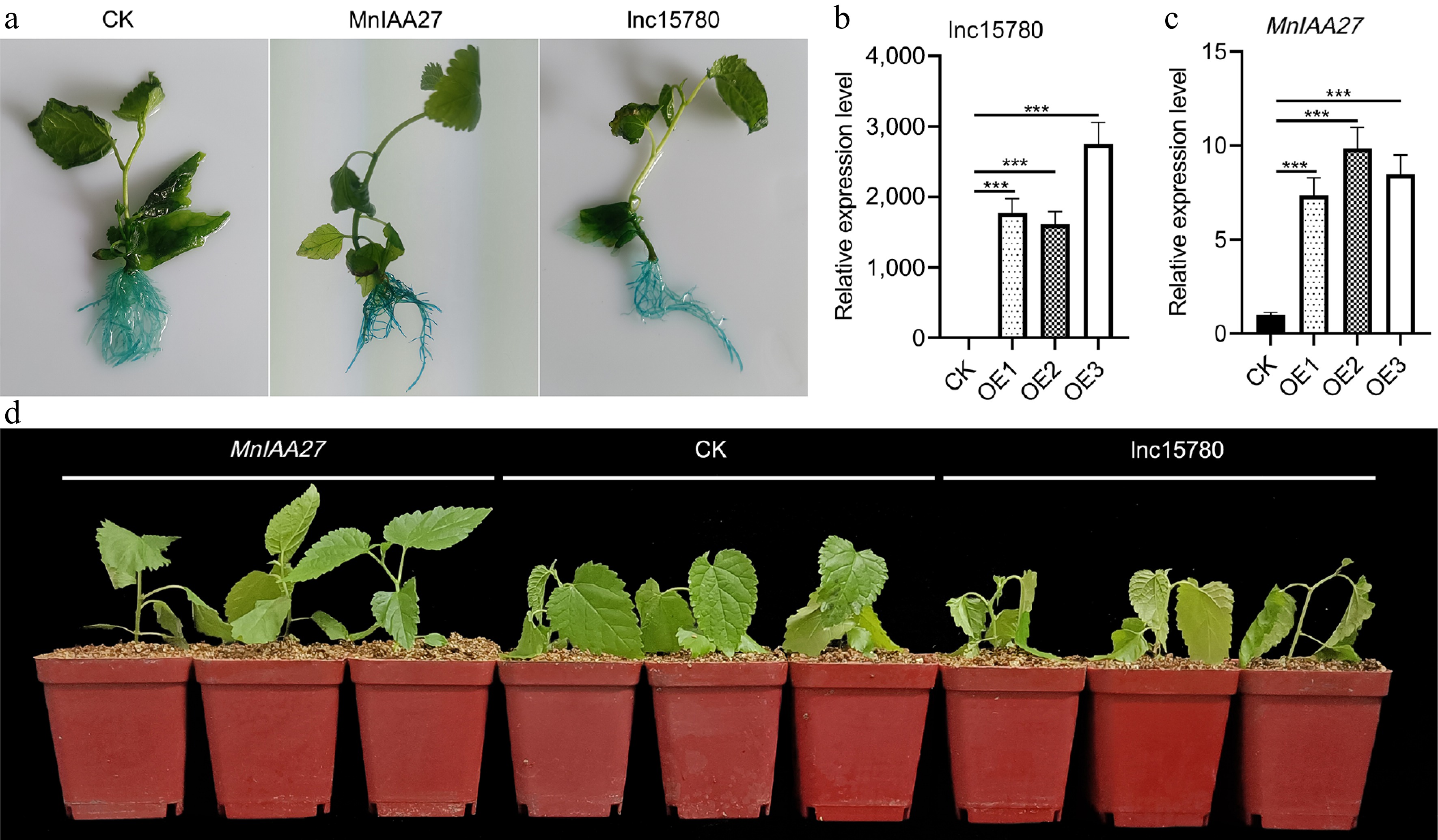
Figure 7.
Analysis of lnc15780 and MnIAA27 salt tolerance of using transgenic hairy roots mulberry. (a) GUS staining was performed on transgenic hairy roots mulberry. The relative expression levels of (b) lnc15780 and (c) MnIAA27 in transgenic hairy roots were determined by RT-qPCR. (d) After salt treatment for 12 h, the transgenic hairy roots mulberry trees were photographed. Error bars represent the SD of three biological replicates (each replicate was a pool of three infected plants) (*** p < 0.001; two-tail t-test). CK, empty vectors.
The levels of hydrogen peroxide (H2O2) and superoxide anion (O2−) were assessed using DAB and NBT staining techniques, respectively (Fig. 8a, b). Under normal conditions, H2O2 and O2− accumulation levels were similar in all plants. Under salt stress, H2O2 and O2− accumulation in MnIAA27-OE lines decreased compared with CK plants, while H2O2 accumulation in lnc15780-OE lines increased. The results indicated that the overexpression of MnIAA27 led to a significant reduction in the accumulation of both H2O2 and O2−. Conversely, the overexpression of lnc15780 was associated with an increase in the levels of both H2O2 and O2−. Therefore, we further studied the ROS clearance ability of overexpression plants. The activities of APX, POD, and CAT were measured. Compared with CK plants, the activities of APX, POD, and CAT in MnIAA27-OE line were significantly increased after 12 h of salt stress, while the activities of APX, POD, and CAT in lnc15780-OE line were significantly decreased (Fig. 8c−e). However, under normal conditions, there was no difference in APX, POD, and CAT activity between study lines. These results suggest that MnIAA27 can improve antioxidant capacity and reduce oxidative damage under salt stress, thereby improving salt tolerance, while lnc15780 has the opposite function.
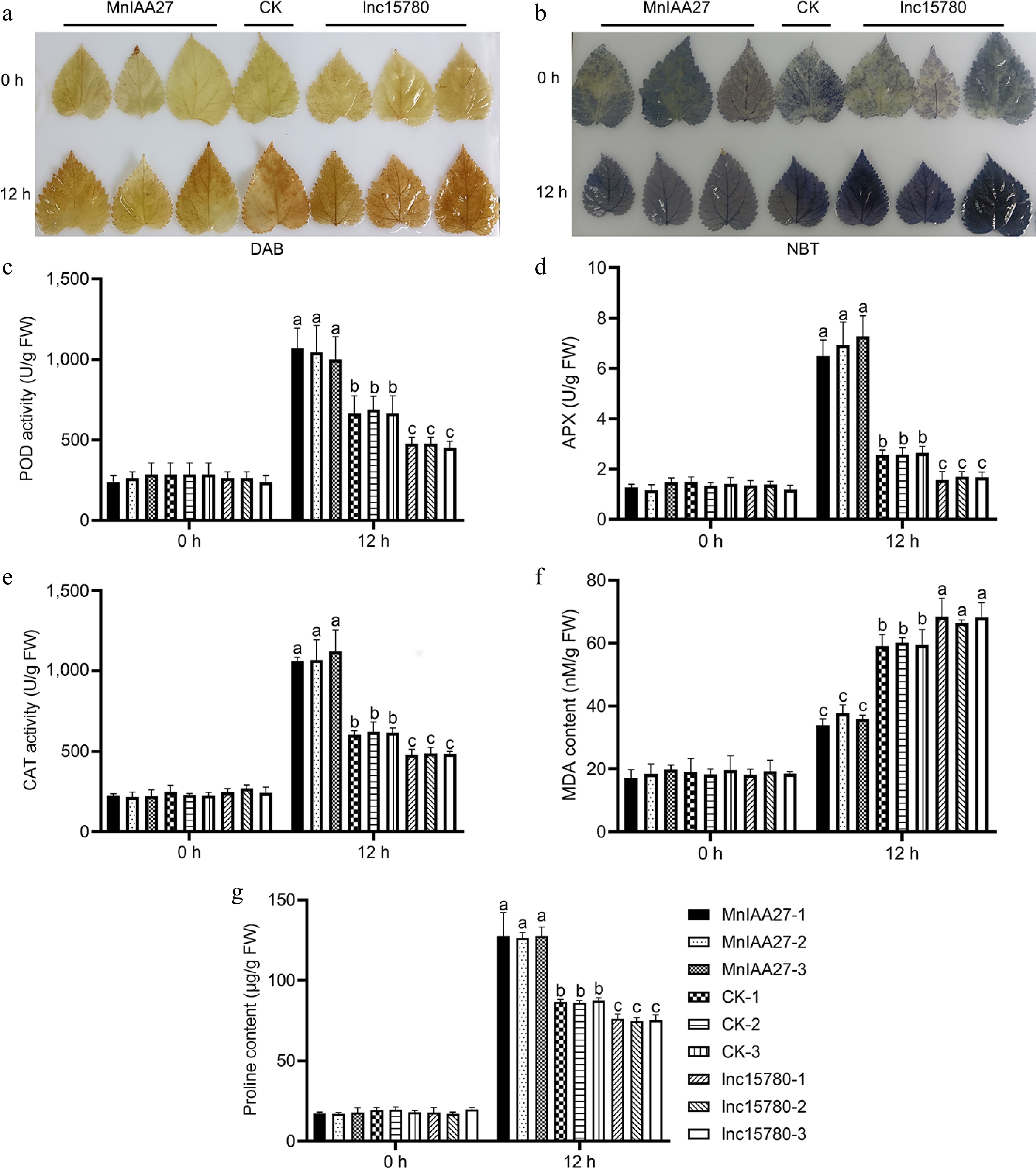
Figure 8.
Determination of salt tolerance by histochemical and physiological analysis. (a) DAB staining showed H2O2 level. (b) NBT staining showed O2− level. (c) POD activity analysis. (d) APX activity analysis. (e) CAT activity analysis. (f) MDA content analysis. (g) Proline content analysis. The value is the average of the three replicates. The error bar represents the standard deviation. According to Tukey's HSD test, different letters indicated significant differences (p < 0.05).
In addition, we evaluated the alterations in MDA levels both in normal environmental conditions and in situations of salt stress. This assessment was conducted as a means to gauge the extent of damage inflicted on cell membranes (Fig. 8f). The results indicated no significant differences in MDA content among all plants under normal growth conditions. Compared with CK plants, the content of MDA in MnIAA27-OE line was significantly decreased after 12 h of salt stress, while the content of MDA in lnc15780-OE line was significantly increased. These results showed that the plasma membrane damage of plant lnc15780-OE was the most obvious, CK was the second, and MnIAA27-OE line was the lightest. Under normal conditions, all plants showed no significant difference in proline content (Fig. 8g). Compared with CK plants, the content of proline in MnIAA27-OE line was significantly increased after 12 h of salt stress, while the content of proline in lnc15780-OE line was significantly decreased.
-
At present, long non-coding RNA (lncRNA) has been recognized more and more as a crucial regulatory molecule. In the realm of botany, a variety of research has shown that lncRNAs are essential in numerous biological processes. One of the significant factors influencing the yield and quality of plants is salt stress. Recently, numerous studies have emphasized the important relationship between the regulatory functions of plants and the responses of lncRNAs to salt stress[43−45]. However, the potential role of lncRNA under salt stress in mulberry has not been studied. Salt-response genes and lncRNAs have been identified in mulberry roots. Through the analysis of genes and lncRNAs, we found 1,201 DE-lncRNAs and 6,330 DEGs in root tissue. Numerous long non-coding RNAs (lncRNAs) exhibiting a range of functions, including cis-acting, trans-acting, and antisense interactions with mRNA, have been identified in recent studies. These findings have provided valuable insights into the diverse roles that lncRNAs play in gene regulation. Our study demonstrated that lncRNA plays a key role in the salt tolerance of mulberry trees through direct means.
Research indicates that exposure to salt stress can hinder the growth and development of mulberry[33,46]. Proline is involved in oxidative processes, offers protection to plants from harm, and serves as a stabilizer for proteins[47]. Lipid peroxidation is a biological process that occurs when lipids, particularly those in cellular membranes, undergo oxidative degradation. This process leads to the formation of various byproducts, one of which is malondialdehyde (MDA). The presence and concentration of MDA within plant cells can be utilized as a crucial indicator of cellular damage[48,49]. After salt treatment, the levels of proline and MDA increased, and the antioxidant capacity decreased (Fig. 1). As expected, the lncRNAs identified in mulberry exhibited a shorter length and had a reduced number of exons when compared to protein-coding transcripts (Fig. 2), which is in agreement with earlier findings in other plant species[50]. A prior investigation demonstrated that lncRNAs may influence gene expression by boosting it at the transcriptional stage. In particular, under all salinity conditions, a large number of DE-lncRNAs upregulation dominated, suggesting that high salinity had a significant impact on lncRNA expression.
KEGG analysis found that deg in the NaCl group was preferably enriched in 'metabolic pathways' compared to the control group (Fig. 4), suggesting that salt treatment enhanced this process in the roots. Plants produce many different metabolites in spatio-temporal and/or environment-dependent ways[51]. To mitigate stress impacts on growth and development, plants regulate metabolic pathways to meet the energy demands of changing environmental conditions[52,53]. A plant's adaptability is determined by its capacity to reorganize metabolic networks, regain active growth, and achieve a new state of homeostasis following a period of stress relief[54]. Following salt treatment, many hormone-related genes were significantly upregulated (Supplementary Fig. S2). A large number of studies indicate that no plant hormone is restricted to a singular biological function in plants; instead, it serves multiple and intricate roles across various stages, tissues, or environmental situations[55,56]. Salinity signals triggered by salt stress are mediated by plant hormones to manage the adaptation of plant growth, while plants establish defense mechanisms through the regulation of synthesis, signaling, and metabolism of different hormones[57]. Genes in GO classification that respond to salt stress are mainly enriched in 'response to stimulus'. One of the intracellular signaling pathways is through the Ca2+ signaling system, which is responsible for further signal transduction[58]. As mentioned earlier, the response to stimuli is a crucial biological process linked to candidate genes. Specifically, external factors, including salinity stress, can trigger reactions in plants. Calcium ions (Ca2+) play a vital role in the signaling pathway, with their concentration varying rapidly within the cell. This variation initiates a cascade of signals that are transmitted internally. Calcium receptors can detect these fluctuations, leading to the activation of downstream processes[1].
In addition, in this research, we discovered a specific lncRNA, lnc15780, that is responsive to salt stress. Predictions for target genes suggested that lnc15780 might directly influence MnIAA27, a coding gene located next to lnc15780 within the genome. At the same time, MnIAA27 can be inhibited by lnc15780, indicating that this gene is a target gene of lnc15780 (Fig. 5). LUC studies have shown that lnc15780 binds to the promoter of MnIAA27 (Fig. 6). In summary, lnc15780 plays a key role in the gene regulation system of plants. It can accurately bind to the promoter region of MnIAA27 gene, and then inhibit the expression of MnIAA27 gene. When plants encounter adverse external conditions such as abiotic stress, the internal environment of cells will undergo dramatic changes, and a large number of reactive oxygen species such as H2O2 and O2− begin to accumulate rapidly. This abnormal accumulation situation breaks the original balance between the generation mechanism and the scavenging mechanism of reactive oxygen species in cells, which leads to the imbalance of redox homeostasis in cells and eventually leads to cytotoxicity[59]. Reactive oxygen species (ROS) are recognized for causing harm to different cellular components, such as lipids, proteins, and nucleic acids. This oxidative stress results in increased levels of MDA, a marker for cellular damage and membrane instability. To counteract these detrimental effects, plants employ various protective strategies. One such strategy involves the accumulation of osmotic substances, with proline being a key factor. Proline helps maintain osmotic balance within the plant cells, enabling them to withstand dehydration and preserve cellular integrity. Furthermore, the enzymatic antioxidant system is crucial for managing the harmful effects of ROS. This system functions by enhancing the activities of specific enzymes that serve important roles in detoxifying these reactive species. Among these key enzymes are peroxidase (POD), catalase (CAT), and ascorbate peroxidase (APX). The upregulation of these enzymes fosters a more efficient reduction of ROS concentrations, which is particularly essential for the survival and health of mulberry plants experiencing salt stress. By improving the functionality of these antioxidant enzymes, plants can bolster their defenses against oxidative damage, ensuring their resilience in challenging environmental conditions. Furthermore, the expression of MnIAA27 improved the functionalities of POD, CAT, and APX, leading to a significant reduction in ROS levels and a decrease in oxidative damage to mulberry plants under salt stress (Fig. 8), thereby bolstering the salt tolerance of the mulberry species. In contrast, the expression of lnc15780 resulted in a reduction of POD, CAT, and APX activities, leading to an increased accumulation of ROS that worsened oxidative damage to mulberry plants subjected to salt stress. In summary, lnc15780 reduced the ROS clearance ability of mulberry by inhibiting MnIAA27, thus making mulberry salt sensitive.
-
In this study, it was found that lncRNAs and mRNAs were highly activated in mulberry roots and played an important role in salt stress response. lncRNAs regulate salt stress tolerance mainly by enhancing 'metabolic pathways'. At the same time, five pairs of lncRNAs and their target genes were identified in response to salt stress. In addition, a working model of salt tolerance mediated by lnc15780 was proposed. The expression of lnc15780 was inhibited by salt stress, and the expression of MnIAA27 was up-regulated. The upregulation of MnIAA27 can lead to significant physiological changes in mulberry, including the decrease of ROS accumulation, the decrease of MDA content, and the increase of proline content, which ultimately improves the salt stress tolerance of mulberry.
The authors acknowledge funding from the Natural Science Foundation of Shandong Province (ZR2022QC136).
-
The authors confirm contribution to the paper as follows: study design: Xin Y, Wang D; data collection: Wang D, Wu H; analysis results: Yu L, Lu Z, Pan L; draft manuscript preparation: Xin Y, Wang D. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files. The raw sequencing data collected from our experiments is publicly available. This data can be accessed through the National Center for Biotechnology Information's Sequence Read Archive (SRA). The specific identification number for these data is noted as Accession no. PRJNA1191174.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primers used in this study.
- Supplementary Table S2 The characteristics of the high throughput sequencing results.
- Supplementary Table S3 Differential expression analysis of lncRNAs and target genes in Control and NaCl.
- Supplementary Fig. S1 Basic statistics about sequencing. Principal component analysis scores of lncRNA (a) and mRNA (b) in control group and salt treatment group. Violin diagram of lncRNA (c) and mRNA (d) expression distribution in control group and salt treatment group.
- Supplementary Fig. S2 Heatmap analysis of differentially expressed plant hormone signal transduction pathway genes.
- Supplementary Fig. S3 Relative expression of MnIAA27 in CK and lnc15780 (OE) in mulberry. Error bars represent the SD of three biological replicates (Each group was treated with three plants) (*** p-value < 0.001; two-tailed t-test).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang D, Wu H, Yu L, Lu Z, Pan L, et al. 2025. Identification and functional characterization of lncRNAs and mRNAs in response to salt stress in mulberry. Forestry Research 5: e006 doi: 10.48130/forres-0025-0006 

Identification and functional characterization of lncRNAs and mRNAs in response to salt stress in mulberry
- Received: 17 February 2025
- Revised: 14 March 2025
- Accepted: 17 March 2025
- Published online: 31 March 2025
Abstract: Long non-coding RNAs (lncRNAs) have been recognized as significant contributors to the mechanisms underlying abiotic stress tolerance in various organisms. Nevertheless, the specific mechanisms through which they contribute to this tolerance remain ambiguous. This study identified salt-response genes and characterized the functions of lncRNAs in mulberry roots. RNA-seq identified 6,330 mRNAs and 1,201 lncRNAs as responsive to salt treatment. We identified the function of mulberry salt in response to lncRNA (lnc15780). Transgenic hairy root plants of mulberry overexpressing lnc15780 were generated. lnc15780 reduces the salt tolerance of plants by inhibiting antioxidant activity. LUC combined with RNA sequencing showed that lnc15780 inhibited the expression of MnIAA27 by binding to its promoter. The hairy root plants of mulberry with MnIAA27 overexpression were produced. The overexpression of MnIAA27 gives it salt tolerance. In addition, MnIAA27 reduces ROS accumulation. In summary, lnc15780 acts as a negative regulator of MnIAA27, leading to a reduction in the expression of lnc15780 during periods of salt stress. This decrease activates the expression of MnIAA27, which subsequently promotes both plant growth and enhances salt tolerance. Consequently, lnc15780 emerges as a promising candidate gene for molecular breeding efforts aimed at developing plant varieties that exhibit higher growth rates alongside improved resilience to salt stress. Our findings indicate that lncRNA is integral to the salt response mechanisms in mulberry, highlighting its significance in the plant's adaptation to saline environments.
-
Key words:
- Mulberry /
- Salt tolerance /
- Long noncoding RNA /
- ROS /
- lnc15780 /
- MnIAA27