









-
P. ginseng is traditionally used to nourish humans by enhancing immune system function, exhibiting anti-tumor properties, and has provided anti-aging benefits for thousands of years in China and Korea[1,2]. The increasing demand gradually makes wild ginseng resources endangered. Currently, commercially available ginseng is primarily derived from cultivated alternatives. As a perennial Chinese herbal medicine of the family Araliaceae, five to six years is commonly needed from sowing to harvest[3]. Rust rot is caused by the soilborne fungus Cylindrocarpon destructans (Zins) Scholten (teleomorph Nectria radicicola), which includes three varieties, i.e. C. destructans var. destructans, C. var. coprosmae, and C. macroconidialis[4]. Rust rot is prevalent in many ginseng (P. ginseng) and American ginseng (P. quinquefolius) production areas annually, and results in a significant decline in yield and quality[5,6]. The infection of ginseng by C. destructans was influenced by environmental factors (temperature, humidity, soil type), plant characteristics (variety, growth stage, resistance), cultural practices (crop rotation, field hygiene, fertilization), and types of pathogens (virulence, composition of species)[7]. It has been reported that pH 5, 20 °C, and a −0.02 MPa moisture soil environment was more suitable for C. destructans, and 2-year-old ginseng roots were more susceptible[5]. Furthermore, even without a suitable host, C. destructans can survive in the soil for over 10 years, which significantly discounts the controlling efficacy of using a crop rotation strategy[8]. Currently, the control of rust rot mainly depends on chemical fungicides, such as carbendazim, triflumizole, tebuconazole, etc.[9,10]. However, this results in severe environmental contamination, multiple fungicide resistance, and excessive fungicide residues in ginseng products[11].
It has been demonstrated that developing and applying environmentally friendly and effective biocontrol agents is an optimal alternative solution to ensure relatively stable yield and quality of P. ginseng[9,12]. Recent studies have extensively explored the beneficial biocontrol microorganisms, such as fungi[13], bacteria[14], and other species[15], to control ginseng diseases. So far, numerous microorganisms in the rhizosphere soil showing antagonistic activity towards phytopathogens have been isolated, presenting considerable potential for disease control and plant growth promotion[16−19]. As an important component of the soil ecosystem, bacteria play a unique role in nutrient cycles, growth-promoting, and broad antagonistic spectra[20]. Bacillus species, as a group, offer advantages over other microorganisms for their ability to form endospores, strong survival ability, and produce numerous active and structurally diverse antagonistic metabolites[16,21−24]. Biocontrol agents have been developed and successfully applied targeting many important plant diseases using Bacillus species including B. subtillis[20,25,26].
Lipopeptides, such as surfactin, fengycin, and iturin, are types of secondary metabolites synthesized by non-ribosomal peptide synthetases via a thiotemplate multienzyme mechanism by the genus of Bacillus[27−31]. So far, hundreds of lipopeptides with different chemical structures have been identified[32,33]. As well as their antagonistic activity against important crop pathogens, these lipopeptides are reportedly involved in inducing systemic resistance in plants[34,35]. In our previous research, a large number of microorganisms, including the genus of Trichoderma, Bacillus, Pseudomonas, etc., were isolated from the rhizosphere soils and interior tissues of medicinal plants. Among them, B. subtilis ge28 isolated from a healthy ginseng root indicates excellent potential in diseases control for its significant antagonistic activity to several ginseng casual pathogens.
In the present study, the antifungal activity of the ge28 strain against C. destructans was confirmed in vitro. However, the composition of biocontrol-related metabolites and their biological synthesis pathways are still unknown. The complete genome of ge28 was sequenced by high throughput sequencing, and lipopeptides were identified by UPLC-ESI-QTOF-MS. Furthermore, the antifungal mechanisms of ge28 were appraised, and the biocontrol mechanisms of lipopeptides involve causing deformities and breakage of C. destructans mycelia, and inhibiting spore production. Finally, the control efficacy of ge28 on rust rot of P. ginseng was evaluated by pot experiments, as a result, the disease indexes in treatments of diluted fermentative liquid and suspensions of ge28 were significantly lower than the control, which implied that B. subtilis ge28 has the potential to develop biocontrol agents for the control of ginseng rust rot.
-
Healthy roots of 3-year-old P. ginseng (6 to 8 g on average) of the 'DaMaYa' cultivar were collected from an agricultural station (127°35′23″ E, 42°14′52″ N) in Fusong County, Jilin province (China). Isolation of endophytes from ginseng plants was conducted according to previous methods[36,37]. After being washed in running water, successively treated with 70% ethanol for 3 min, 2.6% sodium hypochlorite for 5 min, and 70% ethanol for 30 s, then washed with sterilized distilled water five times, the retained water in the roots was dried using sterilized filter paper. The roots were then cut into pieces and placed into a sterile mortar with sterile quartz sand and saline (0.85%). After thoroughly grinding, the tissue homogenate was diluted up to 10,000-fold with sterile water. Diluents of 200 μL were uniformly applied on Luria–Bertan (LB) agar medium (10 g/L tryptone, 5 g/L yeast extract, 10 g/L NaCl, 15 g/L agar powder, pH7) and cultivated at 25 °C for 7 d.
Antifungal assays
-
For the isolation of endophytic bacteria from P. ginseng, each candidate strain was purified by gradient dilution and streak plate techniques. The pathogenic fungus, C. destructans, that was stored in our laboratory, was activated at 25 °C on a potato dextrose agar (PDA) plate (200 g/L potato, 20 g/L dextrose, 15 g/L agar powder, pH7) for 7 d. Antifungal assays were conducted according to previously reported methods[15,36]. A piece of C. destructans plug (5 mm in diameter) from the edge of the colony was inoculated on the center of a PDA plate and incubated at 25 °C for 1 d. Then, the ge28 strain was inoculated 20 mm from the center of the plate, and the control was carried out using a sterile PDA plug. Each treatment was performed in three replicates. After co-culture at 25 °C for 7 d, the colony radiuses of C. destructans were measured, then the inhibition rate was calculated using the formula: inhibition rate (%) = (rc – ri) / rc × 100. 'rc' indicates colony radius of control, 'ri' indicates colony radius of treatment that confrontation culture with ge28 strain.
Samples of C. destructans from the control and the ge28 strain treatments were collected for ultrastructural evaluation. After being fixed with 2.5% glutaraldehyde at 4 °C overnight, the samples were rinsed three times with phosphate-buffered-saline at 10 min intervals. Dehydration was performed using 30%, 50%, 60%, 70%, 80%, 90%, and 100% ethanol for 15 min sequentially, and ethanol was displaced three times by 100% tertiary butanol overnight. Then, the samples were examined using SEM JSM-6510LV after freezing, drying, and gold sputtering in a vacuum.
Biocontrol assays
-
Pot experiments were carried out in the experimental field of the Institute of Medicinal Plant Development, Chinese Academy of Medical Sciences (Beijing, China). Triennial P. ginseng seedlings were transplanted into pots (five seedlings per pot) filled with humus before sprouting. The control efficacy of ge28 against C. destructans was evaluated. In detail, suspensions (108 and 109 cfu/mL) and the fermented liquid (10 times diluted) of ge28 were applied around the roots (100 mL per pot) after the leaves of ginseng plants were fully developed. An equal amount of sterile water was applied in the control. A spore suspension of C. destructans (4.4 × 106 cfu/mL) was inoculated (100 mL per pot) 24 h later. Each treatment was performed in three replicates. The severity of rust rot was examined 20 d post-inoculation (dpi) according to the classification standard (Supplementary Table S1) provided in previous studies[14,38]. Disease index and biocontrol efficacy were calculated according to the method of Principe et al.[39].
Phylogenetic analyses
-
The universal primers fD1 (5'-AGAGTTTGATCCTGGCTCAG-3') and rP1 (5'-ACGGTTACCTTGTTACGACTT-3') were used to amplify the 16S rDNA of ge28[40,41]. Amplification was performed in 25 μL reaction volume, containing 1 μL of template DNA (20 ng/μL), 2.5 μL of 10× buffer (Beijing TransGen Biotech Co., Ltd., Beijing, China), 1 μL of Taq DNA polymerase (2.5 U/μL, Beijing TransGen Biotech Co., Ltd.), 1 μL of dNTPs (2.5 μmol/L, Beijing TransGen Biotech Co., Ltd.) and 1 μL of each primer (2 μmol/L), ddH2O was added to reach to the final volume. Amplification was performed in a Bio-Rad thermal cycler T100 under the following cycling conditions: an initial denaturation at 94 °C for 5 min; 30 cycles of 94 °C for 1 min, 55 °C for 1 min, 72 °C for 1 min; a final extension at 72 °C for 10 min[40,42]. After successful amplification was confirmed by electrophoresis on 1% agarose gel, PCR amplicons were purified using a gel extraction kit (Beijing TransGen Biotech Co., Ltd.) and sequenced by Sangon Biotech (Shanghai) Co., Ltd. (Shanghai, China). Blast analyses were performed in the sequences using the blastx program from the National Center for Biotechnology Information (NCBI) website (
http://blast.ncbi.nlm.nih.gov/Blast.cgi ). The 16S rRNA gene sequences of strains that showed high sequence similarity (Identity > 98%) to ge28 were used to build a phylogenetic tree using the MEGA 6.0 program based on the neighbor-joining (NJ) algorithm[43,44].Genome sequencing and assembly
-
Cells of ge28 were cultivated in liquid LB medium (without agar) at 200 rpm and 30 °C overnight, then harvested through centrifuging at 7,000 g for 10 min. Genomic DNA was extracted using the cetyltrimethylammonium bromide (CTAB) method. The complete genome sequence of ge28 was obtained through Illumina HiSeq4000 and Pacific Biosciences (PacBio) RS II sequencing. The genome sequencing data were assembled using the methods described by Koren et al.[45]. Briefly, raw reads were initially trimmed and filtered. Then, SOAPdenovo v2.04 was used to preliminarily assemble data obtained from Illumina. Celera Assembler v8.0 was used for further assembly after the reads from PacBio were corrected by Illumina data. Finally, gaps were closed using GapCloser v1.12 when all scaffolds were linked. The average sequencing depth was 250- and 310-fold for the Illumina and PacBio data, respectively.
Genome annotation and metabolites related to NRPS prediction
-
Gene sequences were predicted using Glimmer software version 3.02 (
www.cbcb.umd.edu/software/glimmer ), and their functions were annotated by BLASTP against NR, Swiss-Prot, GO, COG, and KEGG databases[46,47]. rRNA and tRNA genes were predicted by RNAmmer version 1.2 and tRNAscan-SE version 1.3.1, respectively[48,49]. Gene clusters associated with the biosynthesis of secondary metabolites related to NRPS and polyketide synthase (PKS) were identified by antiSMASH version 4.0 (http://antismash.secondarymetabolites.org/ )[50].Extraction of lipopeptides
-
Following incubation in liquid LB medium at 200 rpm and 30 °C overnight, a 3% (v/v) inoculum of ge28 was inoculated in modified Landy medium[51,52] (20 g/L glucose, 5 g/L L-glutamate, 1 g/L KH2PO4, 0.5 g/L MgSO4, 0.5 g/L KCl, 5 mg/L MnSO4, 0.15 mg/L FeSO4, 0.16 mg/L CuSO4, pH7), and shake incubated at 200 rpm, 30 °C for 48 h. The cell-free supernatant was collected by centrifugation at 12,000 rpm, 4 °C for 20 min. After being adjusted to pH 2 by 6 mol/L HCl and 1 mol/L NaOH, the supernatant was centrifuged at 12,000 rpm, 4 °C for 20 min, lipopeptides were extracted by 100% methanol twice, and the crude lipopeptides were obtained after the methanol was volatilized thoroughly[53]. Finally, the crude lipopeptides were suspended by a suitable reagent for further analysis.
Antifungal assay of lipopeptides
-
The antifungal activity of lipopeptides was evaluated using the cylinder-plate method[54]. Firstly, 400 μL of the spore suspension (106 cfu/mL) of C. destructans was spread on the PDA plate, and four sterile Oxford cups (inner diameter 8 mm) were placed symmetrically at a distance of 20 mm from the center of the PDA plate. Following suspension by PBS and filtration through a 0.22 μm hydrophilic filter, 200 μL of each lipopeptide (10 mg/mL) was added into the Oxford cup. The same volume of PBS was used instead for the control treatment. Each treatment was performed in three replicates. Then, after incubation at 25 °C for 4 d, morphological characteristics of C. destructans mycelia in different treatments were observed through SEM. The detailed procedure was the same as that described for the antifungal assay above.
Identification of lipopeptides
-
UPLC-ESI-QTOF-MS (Xevo G2-XS QTOF, Waters, USA) was performed to analyze the lipopeptides produced by ge28. The UPLC system was equipped with a photodiode array detector, and chromatographic separation was performed on a reversed-phase column (BEH C18, 1.7 μm, 2.1 mm × 100 mm, Waters), with 3 μL injection volume. The mobile phase consisted of water containing 0.1% formic acid (solvent A) and acetonitrile (solvent B). Separation was performed as follows: 20% A and 80% B holding for 15 min. The UV detector was set at 210 nm wavelength. The flow rate remained at 0.3 mL/min throughout the run, and the temperature of the column and sample was kept at 25 °C. The positive ion acquisition mode at a capillary voltage of 3 kV was implemented in QTOF-MS analysis. The mass spectral scan time was 0.2 s, and the positive mode full scan was performed from m/z 50 to 2000. Data acquisition and collection were performed by Masslynx version 4.1. The molecular formulae of lipopeptides were deduced according to Liao et al.[55].
Statistical analysis
-
Data obtained in the experiments was analyzed by SPSS 22.0 via analysis of variance (ANOVA). The least significant difference test (LSD) was used to estimate the significant differences between treatments.
-
In our previous research, dozens of endophytic bacteria were isolated from healthy ginseng roots. Among them, ge28 exhibited significant antifungal activity against C. destructans through the dual culture assay method. The radius of the C. destructans colony that was exposed to ge28 was 16 ± 1 mm after 8 d of culture, whereas the control colony had a radius of 44 ± 1 mm (Fig. 1a, d). B. subtilis ge28 exhibited significant antifungal activity against C. destructans with an inhibition rate of 63.63% ± 2.27%. Results of scanning electron microscopy (SEM) showed that the hyphae of C. destructans in control were smooth and well-distributed (Fig. 1b, c), while those exposed to ge28 were distorted, malformed, and fractured (Fig. 1e, f).
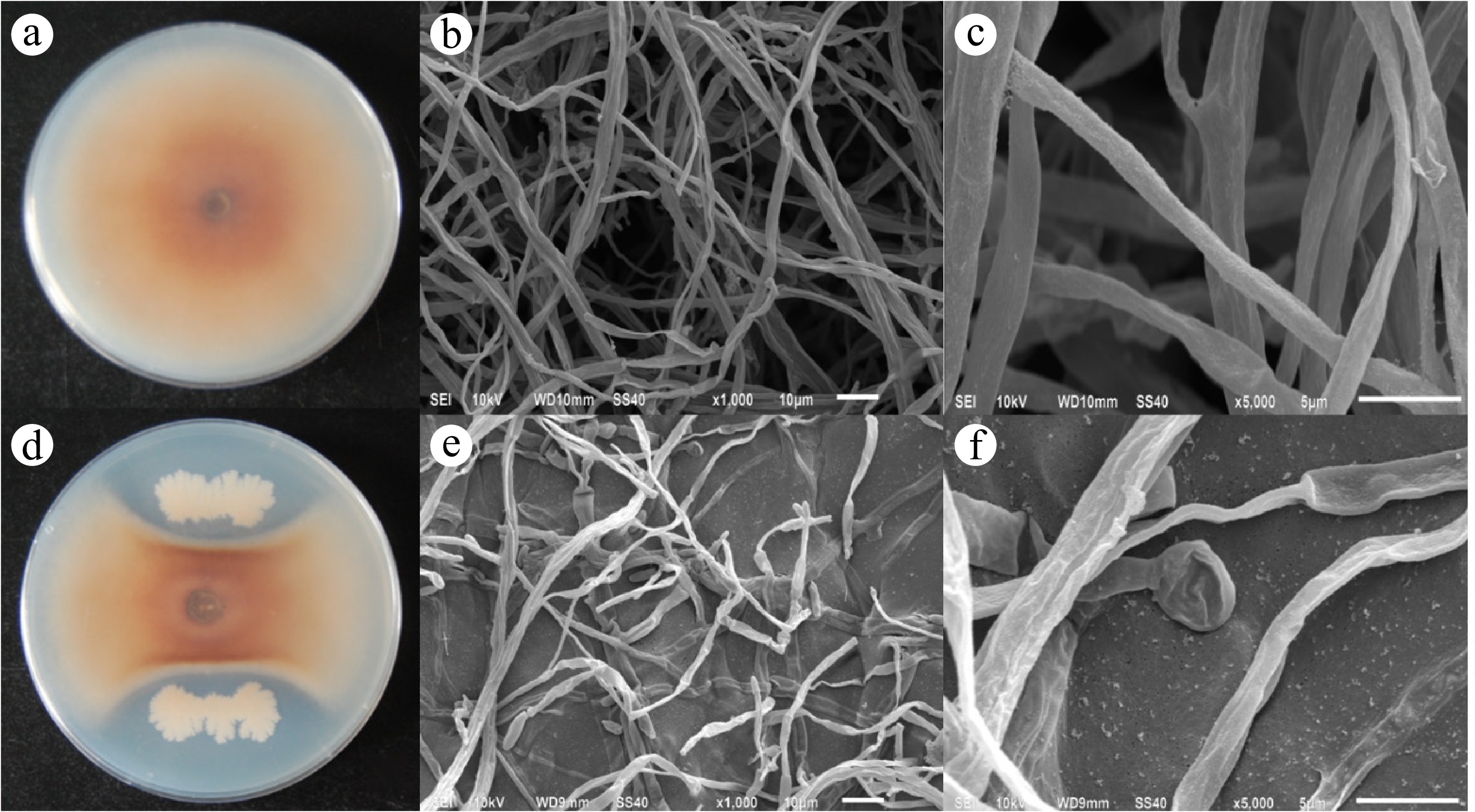
Figure 1.
Antifungal activity of ge28 against C. destructans. (a) Colony of C. destructans after 8 d of culture; (b) C. destructans grown under normal conditions (magnification 1,000×); (c) C. destructans grown under normal conditions exhibiting a smooth hyphae surface and a uniform thickness (magnification 5,000×); (d) the colony of C. destructans treated with ge28 incubated for 8 d; (e) hyphae of the C. destructans, whose growth was inhibited by ge28, exhibiting malformations, distortions, and fractures (magnification 1,000×); (f) Mycelial shrinks significantly and expands into a spherical shape at the end (magnification 5,000×).
Biocontrol efficacy of ge28 on rust rot of P. ginseng
-
Pot experiment results indicated that the number and size of rusty lesions were reduced in all ge28 treatments. Ginseng roots in the control exhibited apparent rusty lesions that were moderately larger, with the lesions occupying more than half of the root area or the roots entirely rotted (Fig. 2a). However, the disease severity in ginseng roots in treatments of both ge28 fermentation (Fig. 2b) and ge28 suspension (Fig. 2c, d) was significantly reduced, and only very few roots developed sporadic lesions.

Figure 2.
Rust severity degree in ginseng plants under different ge28 inoculation solution or ge28 suspension treatments. (a) control; (b) ge28 fermentation solution (diluted 10 times); (c) ge28 suspension of 108 cfu/mL; (d) ge28 suspension of 109 cfu/mL.
Compared with the control, the disease index of rust rot on ginseng roots was reduced significantly in the ge28 fermentation or suspension treatments (Table 1). Among them, the fermentation liquid of ge28 that was 10-times diluted exhibited the highest biocontrol efficacy on rust rot of P. ginseng, followed by the 108 cfu/mL suspension, while the biocontrol efficacy of 109 cfu/mL suspension was the lowest.
Table 1. Control efficacy on rust rot of P. ginseng in a pot experiment.
Treatment Disease index (%) Biocontrol efficacy (%) CK 46.67 ± 1.33a − ge28 fermentative liquid (10-times dilution) 22.67 ± 2.67c 51.42 ge28 suspensions (108 cfu/mL) 24.00 ± 2.31c 48.58 ge28 suspensions (109 cfu/mL) 34.67 ± 2.67b 25.71 Different lower case letters indicate significant differences at the p < 0.05 level between the treatments and control. Phylogenetic tree of ge28 and related strains
-
The amplified length of the 16S rRNA gene of ge28 was 1,453 bp, and its nucleic acid sequence is presented in Supplementary File 1. In total, 17 strains from the Bacillus genus, including four B. subtilis (BSn5, SG6, SZMC 6179J, and 168), four B. methylotrophicus (SH1, PY5, PK9, and ZZB21), four B. velezensis (JS64Y, 9912D, You33, and JS65O), three B. amyloliquefaciens (UMAF6614, CC178, and Y2) and two B. vallismortis (NBIF-001 and 263XY1), together with the sequence of 16S rRNA gene of ge28, were used for blast comparative analyses and the phylogenetic tree construction. The blast results showed that ge28 exhibited the highest (≥ 99%) sequence homology to B. subtilis. As illustrated in the phylogenetic tree, ge28 shows the closest relationship to B. subtilis BSn5, clustered in the same branch with all the four trains of B. subtilis, yet it was distinct from other species of the Bacillus genus (Fig. 3).
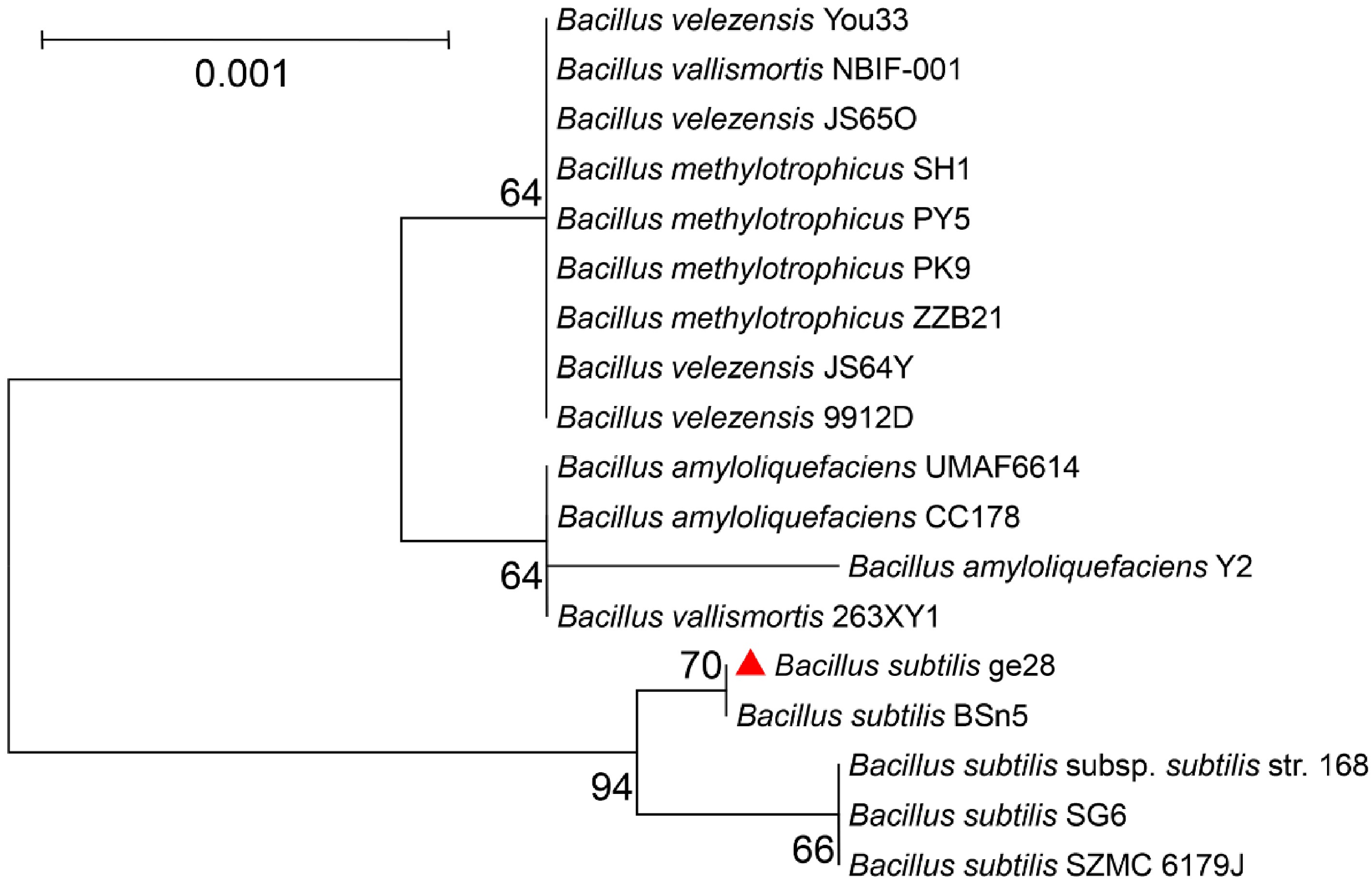
Figure 3.
Neighbor-joining (NJ) phylogenetic tree based on the 16S rRNA gene sequence of species from the genus of Bacillus illustrating their genetic relationships. The bootstrap method (1,000 resamplings) was used to construct the NJ tree. Bootstrap values > 50% are indicated at branch points. The scale bar represents 0.001 substitutions per nucleotide position.
General genome features of ge28
-
The 4,065,429 bp genome of ge28 exhibited a GC content of 43.8%. The chromosome contained 3,879 protein-encoding genes, 30 rRNA genes, and 89 tRNA genes (Fig. 4, Supplementary Table S2). Most of the identified genes were associated with energy production and conversion, amino acid transport and metabolism, nucleotide transport and metabolism, lipid transport and metabolism, carbohydrate transport and metabolism, cell motility, secondary metabolite biosynthesis, transport and catabolism, and defense mechanisms.
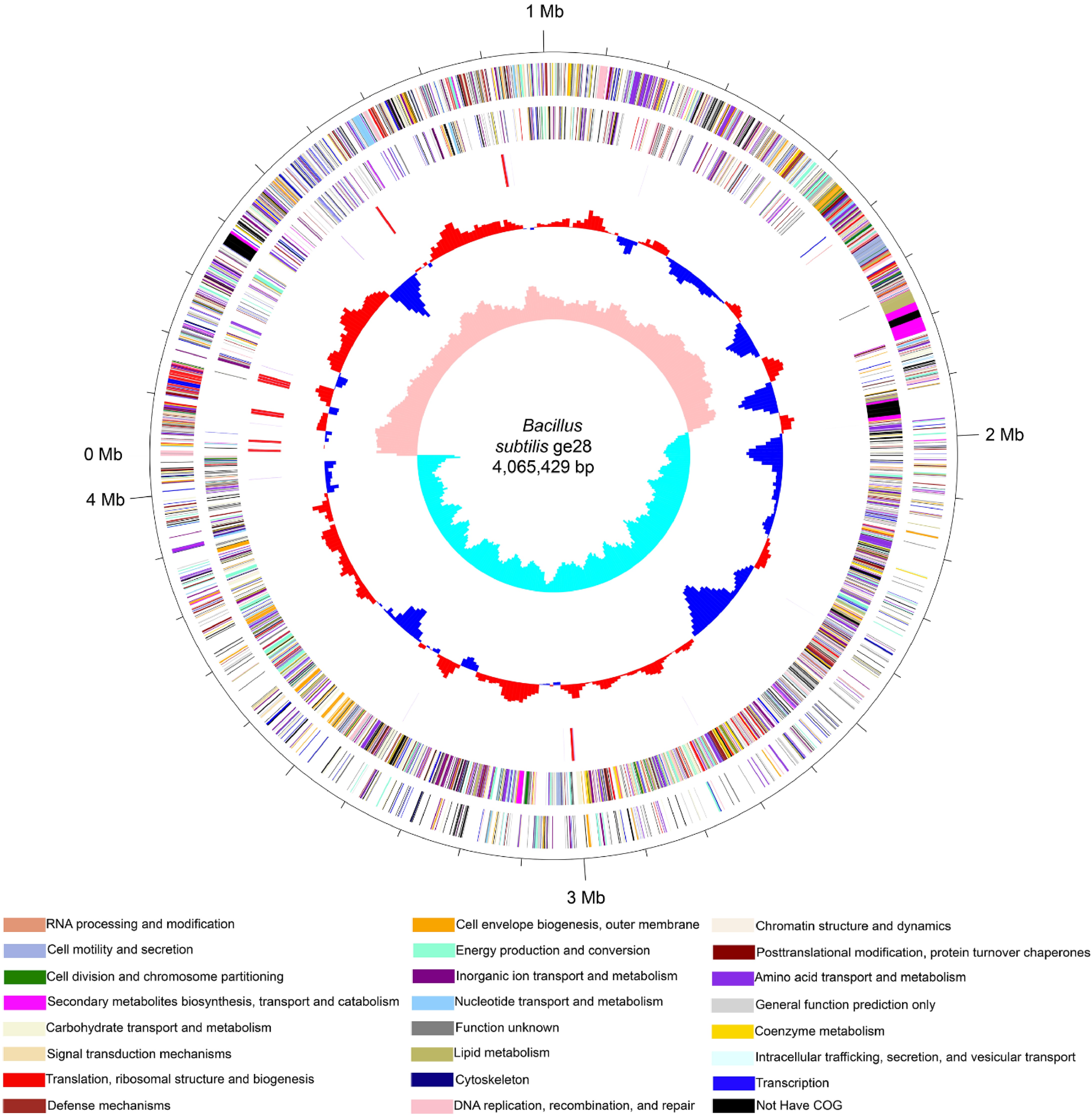
Figure 4.
Circular genome map of B. subtilis ge28. The five circles (outer to inner) represent forward-strand CDSs, reverse-strand CDSs (different colors correspond to different categories in the COG designation of CDSs), rRNA and tRNA, GC content, and GC skew.
Secondary metabolites related to the NRPS gene cluster
-
In total, 13 candidate gene clusters responsible for secondary metabolite biosynthesis were identified using antiSMASH version 4.0. In addition, putative gene clusters involved in subtilin, subtilosin, bacilysin, and bacteriocin biosynthesis were identified. Three gene clusters encoding NRPS modules were predicted to facilitate the assembly of surfactin, fengycin, and bacillibactin (Fig. 5). Furthermore, a few antibiotic peptide synthesis genes were also identified, namely srfAA, srfAB, srfAC, srfAD, fenA, fenB, fenC, fenD, fenE, dhbA, dhbB, dhbC, dhbE, and dhbF. Regarding surfactin, antiSMASH predicted the core structures of surfactin A, X-Glu-Leu-Leu-Val-Asp-Leu-Leu, and fengycin, X-Glu-Orn-Tyr-Thr-Glu-Ala (fengycin A)/Val (fengycin B)-Pro-Gln-Tyr-Ile, where X corresponds to the β-hydroxy acid residue. In addition, bacillaene was unique among the various metabolites as it was synthesized by the NRPS-TransatPKS cluster, a large and complex cluster.

Figure 5.
NRPS gene clusters involved in the synthesis of three cyclic lipopeptides (surfactin, fengycin, and bacillibactin) in B. subtilis ge28. Red-colored genes represent core biosynthetic genes. The srfAA, srfAB, srfAC, srfAD, fenA, fenB, fenC, fenD, fenE and dhbA, dhbC, dhbE, dhbB, dhbF genes were identified in these NRPS gene clusters.
Antifungal activity of the lipopeptides
-
The lipopeptides synthesized by ge28 exhibited antagonistic activity against C. destructans by forming an obvious inhibition zone (Fig. 6d), while the colony grew normally in control media (Fig. 6a). During observations through SEM, we found that hyphae exposed to lipopeptides exhibited obvious malformations, distortions, and fractures (Fig. 6e), and very few spores were observed (Fig. 6f). However, colonies in the control group grew normally, and a large number of spores were observed (Fig. 6b, c).

Figure 6.
Influence of lipopeptides on the growth of C. destructans. (a) No antifungal activity was observed for PBS in Oxford cups; (b) C. destructans grown under normal conditions exhibiting smooth hyphae surface and a uniform thickness (magnification 1,000×); (c) C. destructans grown under normal conditions producing a large number of spores (magnification 5,000×); (d) Lipopeptides exhibiting antifungal activity by forming an inhibition zone; (e) hyphae of the C. destructans, whose growth was inhibited by lipopeptide, exhibiting malformations, distortions, and fractures (magnification 1,000×); (f) twisted C. destructans hyphae and only a small amount of C. destructans spores was produced (magnification 5,000×).
UPLC-ESI-QTOF-MS analysis of the lipopeptide antibiotics
-
The mass spectra showed clear peak clusters of the surfactin and fengycin families. The peaks at m/z 994.6479, 1,008.6641, 1,022.6797, 1,036.6954, and 1,050.7100 observed in the mass spectra were the protonated forms ([M + H] +) of surfactin homologs. In contrast, the peaks at m/z 1,449.7849, 1,463.7986, 1,477.8088, 1,491.8223, and 1,505.8361 were the protonated forms ([M + H] +) of fengycin homologues (Supplementary Fig. S1). The corresponding molecular formulae of surfactin and fengycin families (Fig. 7, Supplementary Table S3) were deduced.

Figure 7.
Chemical structures of surfactin and fengycin families.
-
Soil is a complex and dynamic environment with numerous microorganisms interacting with each other, which makes it difficult to target specific soilborne pathogens without disrupting other microorganisms that are beneficial to soil health. As generations of chemical pesticides have been developed and applied in disease control, it is difficult to avoid environmental pollution, pesticide residue, and pathogen resistance[11]. Biocontrol strategies have been demonstrated to be more sustainable and reasonable for its advantages in being environmentally friendly, edible, and having no risk for resistance, etc.[56]. Microorganisms in the genus of Pseudomonas, Streptomyces, and Trichoderma, have been described as possible biocontrol agents for some diseases with a control effectiveness ranging from 30% to 50%[57−59]. Bacillus species have become attractive biological control agents due to their ability to produce hard, resistant endospores and antibiotics which control a broad range of plant pathogens[60]. Now, more than 142 species within the genus of Bacillus have been identified[56]. In our previous research, though a few of biocontrol strains have shown excellent antagonistic activity in vitro, their successful colonization in the soil is significantly influenced by environmental factors, which made the control efficacy unstable and usually unsatisfactory. Differently, B. subtilis ge28 was isolated from the interior tissue of healthy ginseng root, the endophytic characters made it easily colonized in ginseng plants and provides more stable and enduring protection against the infection of external pathogens[61]. Furthermore, except for C. destructans, ge28 also exhibited substantial antifungal activity against several other ginseng pathogens, such as Fusarium oxysporum causing root rot, Alternaria alternata causing black spot, Phytophthora panax causing blight, etc., which demonstrate its great potential to control diseases prevailing in ginseng producing areas.
The most important feature of Bacillus species is their diverse secondary metabolism pathway and their ability to produce a wide variety of structurally different antagonistic substances[62]. Biocontrol mechanisms of potent amphiphilic and surfactant lipopeptides, such as bacillomycins, iturins fengycins, mycosubtilin, and plipastatin synthesized by Bacillus species include the disintegration of cell wall, cell membranes, and fungal intracellular organs, such as nucleic acid and the mitochondria of pathogenic fungi[63,64]. Strains of B. subtilis have approximately 4-5% of their whole genomes dedicated to the synthesis of secondary metabolites, with the capability to produce more than two dozen structurally diverse antimicrobial compounds[65]. Previous studies have confirmed that B. subtilis and B. amyloliquefaciens strains isolated from forest soils and ginseng leaves showed strong antagonistic activity against C. destructans[66,67]. One of the most important antagonistic mechanisms for the genus of Bacillus is their ability to synthesize lipopeptides. The biocontrol capacity of B. subtilis is mainly associated with the lipopeptides that they synthesize[62]. In this study, the significant antagonistic activity of the bacteria-free fermentation broth of B. subtillis ge28 against C. destructans indicates that lipopeptides were synthesized. In recent years, the biosynthetic pathway of natural antagonistic metabolics, such as non-ribosomal peptides and polyketides, were usually recognized through genome mining of the secondary metabolite clusters[50]. The identification of three lipopeptide synthesis gene clusters in the genome and mass spectrometry peaks of surfactin and fengycin indicates that the speculation of lipopeptides synthesized by these gene clusters was accurate, and lipopeptides should be important antagonistic metabolites[68]. The combination of whole-genome mining and mass spectrometry make the exploitation of new antagonistic metabolites time-saving and efficient[69].
Bacillibactin have been reported to be synthesized by several species of the Bacillus genus, such as B. anthracis and B. subtilis, and which are classified as 1,2-dihy-droxyaryl (pyrocatechol)-centered hexadentate siderophores to complex ferric iron (Fe3+) from the surrounding environment and transferred into the cytoplasm[70−72]. In this study, bacillibactin, was also predicted from ge28 by antiSMASH. Iron, although the fourth most abundant metal in the earth's crust, is usually a growth-limiting nutrient for microorganisms[73]. This siderophore can facilitate the acquisition of ferric iron under low iron concentrations in the environment. Bacillibactin cannot be utilized by pathogens, which usually acts as an inhibitor of plant pathogens. Chen et al.[74] reported that rhizospheric B. amyloliquefaciens FZB42 produces high concentrations of the siderophore bacillibactin, which inhibits the growth of pathogenic competitors by depriving them of essential iron ions. The prediction of the synthesis of bacillibactin by ge28 made the antagonistic mechanisms against C. destructans more complicated.
The efficacy of biocontrol agents on soil-borne diseases can be influenced by many factors, such as host specificity, microbial population in the soil, the ability to colonize the host, etc.[75]. One of the most crucial factors is the massive colonization of biocontrol agents in the plant rhizosphere soil, this may be realized by increasing the concentration and frequency of biocontrol agents' application[14]. This will however, substantially elevate production costs. It was inferred that the microbial community colonizing the soil of the ginseng rhizosphere maintained a dynamic balance, where a certain number of B. subtilis accommodated in a finite habitat niche, play a key role in the growth of healthy ginseng plants[76]. In this study, ge28 exhibits prominent biocontrol potential on rust rot of P. ginseng at a very low concentration, indicating that which could be used to develop biocontrol agents to control soilborne diseases, including rust rot in the process of ginseng production. Further research is needed to determine the effectiveness of large-scale applications of the ge28 strain and lipopeptides in controlling C. destructans, to establish the optimal biological strategy to control ginseng rust rot. All these efforts will establish an empirical foundation for advancing our understanding of the complex plant-microbe interaction mechanism, and enable us to design more precise, targeted management.
-
This study isolated an endophytic B. subtilis strain ge28 that exhibits strong inhibition against C. destructans, the causal pathogen of rust rot in ginseng roots. In vitro and pot experiments demonstrated that fermentation and the supernatant of ge28 show substantial biocontrol potential against rust rot. Three gene clusters coffering the synthesize of lipopeptides were found in the genome of ge28, and two types of lipopeptide substances, surfactin, and fengycin, were identified by UPLC-ESI-QTOF-MS, their molecular formulae and chemical structure were elucidated. These results will accelerate the application of ge28 in ginseng production regions to enhance the yield and quality of P. ginseng through substituting of chemical fungicides with microbial-derived agents.
This work was supported by the National Key R&D Program of China (2022YFC3501504), the CAMS Innovation Fund for Medical Sciences (2021-I2M-1-032), the National Project for Standardization of Traditional Chinese medicine (ZYBZH-C-JL-27), and the Innovation Research Fund of Peking Union Medical College (1007-14).
-
The authors confirm contribution to the paper as follows: study conception and design, methodology, funding acquisition: Gao Y, Li Y; data collection, analysis and interpretation of results, draft manuscript preparation: Gao Y, Wang J; supervision: Wei J, Ding W; manuscript proofreading and editing: Li Y, Zhong S, Wang R. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available in GenBank with the Accession no. CP021903 (www.ncbi.nlm.nih.gov/nuccore/CP021903).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yuan Gao, Junchi Wang
- Supplementary Table S1 Classification standard of disease severity.
- Supplementary Table S2 Genomic features of B. subtilis ge28.
- Supplementary Table S3 Main peaks of lipopeptides synthesized by B. subtilis ge28.
- Supplementary Fig. S Each peak (m/z) of surfactin and fengycin families. (a)−(e) surfactin family; (f)−(h) fengycin family.
- Supplementary Fig. S1
- Supplementary File 1 Bacillus subtilis ge28, 16S rRNA gene.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Gao Y, Wang J, Zhong S, Wang R, Wei J, et al. 2025. Bacillus subtilis ge28, a potential biocontrol agent for controlling rust rot in Panax ginseng caused by Cylindrocarpon destructans. Medicinal Plant Biology 4: e011 doi: 10.48130/mpb-0025-0009 

Bacillus subtilis ge28, a potential biocontrol agent for controlling rust rot in Panax ginseng caused by Cylindrocarpon destructans
- Received: 13 December 2024
- Revised: 27 February 2025
- Accepted: 20 March 2025
- Published online: 16 April 2025
Abstract: Panax ginseng is a traditional herbal medicine used worldwide. Rust rot caused by Cylindrocarpon destructans infects the root of P. ginseng and usually results in severe yield losses and quality deterioration. Quality control of P. ginseng is crucial to ensure its medicinal value. However, the increasing application of chemical fungicides for controlling severe epidemics of diseases including rust rot in ginseng-producing regions every year has led to significant environmental pollution, fungicide residues in harvested ginseng, and an increased risk of phytopathogen resistance. In this study, an endophytic Bacillus subtillis strain, temporarily named ge28, which has shown strong antagonistic activity to C. destructans, was isolated from healthy roots of P. ginseng. In vitro and pot experiments showed fermentation and the supernatant of ge28 exhibited significant biocontrol efficacy on rust rot. The whole genome of ge28 was 4,065,429 bp, containing 4,162 functional genes. Furthermore, three clusters of non-ribosomal peptide synthetase genes conferring the synthesis of lipopeptides, a group of secondary metabolites exhibiting antagonistic properties, including surfactin, fengycin, and bacillibactin, were identified through genome analysis. Surfactin and fengycin were confirmed to be present in the supernatant of ge28 by UPLC-ESI-QTOF-MS analysis. In conclusion, this study demonstrated that the ge28 strain possesses substantial biocontrol potential against the rust rot of P. ginseng, and its antagonistic mechanisms were preliminarily clarified. These results should accelerate the utilization of ge28 to enhance the yield and quality of P. ginseng through reduced fungicide application.
-
Key words:
- Bacillus subtilis /
- Rust rot /
- Panax ginseng /
- Cylindrocarpon destructans /
- Biocontrol