










-
Compared to annual herbaceous plants, extensive secondary growth is a distinctive feature of woody tree species. During secondary growth, stem cells in the vascular cambium undergo periclinal divisions, giving rise to secondary phloem on the outer side, and secondary xylem on the inner side of the stem[1]. Secondary xylem, which is commonly known as wood, primarily consists of fiber and vessel cells. Fibers provide strong mechanical support for the ever-increasing plant biomass, while vessels are essential for water conduction and nutrient transportation from roots to crowns[2]. Therefore, secondary xylem development is fundamentally important for tree growth.
Secondary cell wall (SCW) biosynthesis is a crucial step in secondary xylem development and has three major components, including 40%–50% cellulose, 20%–30% lignin, and 20%–30% hemicellulose[3]. Cellulose is a linear β-1,4–linked glucose polymer synthesized by the plasma membrane-localized family-2 glycosyltransferase (GT) cellulose synthase (CesA)[4,5]. In transmission electron microscopic visualization, the CesAs are assembled into a six-lobed rosette structure complex, termed cellulose synthase complexes (CSCs), which include 18, 24, or 36 CesAs[6]. Genetic and biochemical studies found there are two types of CSC, type I contains CesA4, CesA7, and CesA8, while type II contains CesA1, CesA3, and CesA6[7−9]. Quantitative proteomics analysis showed that the CESA stoichiometry in angiosperm tree aspen and gymnosperm tree Norway spruce is different, furthermore, the CESA stoichiometry of aspen tension wood is different from that of normal wood[10]. Compared to the wild type, the plant growth, SCW thickness, and cellulose content are significantly reduced in PtrCesA4, PtrCesA7, and PtrCesA8 RNAi knockdown or Cas9/gRNA-induced knockout plants[11]. Lignin is a phenolic biopolymer that is hydrophobic and plays a critical role in vascular transport. Lignin is polymerized from three major monolignols: guaiacyl (G), syringyl (S), and 4-hydroxyphenyl (H) monolignol. In dicotyledon woody trees, S and G are the most abundant lignin subunits, and the S/G ratio significantly affects wood properties. The monolignol biosynthetic pathway is well-characterized, including 23 enzymes and 24 metabolites[12−13]. Lignin polymerization is catalyzed by laccases and peroxidase[14,15]. Genetic studies demonstrated that suppression or knockout of monolignol biosynthesis genes significantly reduce lignin content[16−18], and down-regulation of laccases genes also lead to significantly reduced lignin content and SCW structure[19−21].
The biosynthesis of cellulose and lignin is tightly controlled by a hierarchical transcriptional network[22,23]. Many studies have demonstrated that MYB transcription factors are key regulators directly regulating the expression of cellulose and lignin biosynthesis genes[24−28]. ASYMMETRIC LEAVES1 (AS1) is an R2R3 type MYB transcription factor that promotes adaxial identity in cotyledons, leaves, and floral organ development[29,30]. In Arabidopsis, AS1 forms a complex with AS2 and represses the expression of class I KNOX genes during the establishment of adaxial-abaxial polarity in leaf primordia[30]. The AS1–AS2 complex represses abaxial genes through both Polycomb-dependent and independent mechanisms[31−33]. In addition, mutation of AS1 and AS2 also cause abnormal midvein development[34]. A recent study demonstrated that overexpression of AS1 promotes the elongation of abaxial petiole cells through activating the auxin biosynthesis and signaling pathways during leaf hyponasty growth, and as1 mutants display reduced hypocotyl growth under shade conditions in Arabidopsis[35]. These studies suggested that AS1 may also play an important role in regulating vascular system development.
In this study, we functionally characterized the AS1 ortholog gene PagAS1a in Populus. Sequence analysis revealed that the PagAS1a protein has a highly conserved R2R3 type MYB domain structure. Genetic analysis showed that both overexpression and dominant repression of PagAS1a promote xylem development and plant growth. Transcriptome analysis revealed the lignin and cellulose biosynthesis pathways are significantly up-regulated in PagAS1a overexpression plants. Together, our results demonstrate that PagAS1a promotes xylem development in Populus, which may lead to increased plant growth.
-
All experiments used the hybrid poplar (Populus alba × Populus glandulosa) clone 84K. The plants were grown under a 16 h light/8 h dark cycle at 25 °C. All plants were propagated asexually, and 1-month-old seedlings were transplanted into the soil for phenotypic analysis.
Phylogenetic analysis
-
Phytozome (
https://phytozome.jgi.doe.gov/pz/portal.html ) and NCBI (www.nlm.nih.gov ) were used to retrieve the protein sequences of AS1 from different species. Phylogenetic analysis was performed using MEGA 5.0 software[36]. Sequence alignment was conducted with ClustalW, and the phylogenetic tree was constructed using the neighbor-joining method with 1,000 bootstrap replicates.Subcellular localization of PagAS1a
-
The coding sequence (CDS) of PagAS1a was amplified with gene-specific primers (Supplementary Table S1) and cloned into the pROKII vector, which contains a GFP reporter gene. The recombinant plasmid 35S::PagAS1a-GFP and the control plasmid 35S::GFP were transformed into Agrobacterium (GV3101) and injected into Nicotiana benthamiana leaves, respectively. The nuclei were stained with DAPI and observed under an Olympus BX53 microscope.
RNA extraction and Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
-
Tissue samples, including xylem, phloem, cortex, roots, leaves, and petioles were collected from 2-month-old wild-type (WT) plants. Phloem and xylem samples were collected by stripping the bark and scraping off the phloem or xylem flanks. Total RNA was extracted using the cetyltrimethylammonium bromide (CTAB) method[37]. RNA (1 μg) was reverse-transcribed using the HiScript II Q RT Super Mix for RT-qPCR Kit (Vazyme, R233-01). The RT-qPCR experiments were performed using ChamQ Blue Universal SYBR RT-qPCR Master Mix (Vazyme, Q312-02), and normalized gene expression levels were determined using the ΔΔCt method, employing the Actin gene as the internal reference. The primers used for RT-qPCR are listed in Supplementary Table S1. At least three replicates were performed.
Gene cloning and plant transformation
-
The full-length CDS of PagAS1a was amplified from 84K plants by RT-PCR. The construct for 35S::PagAS1a was generated by ligating the coding sequence into the pZP211-35S-PolyA vector. The construct for dominant repression of PagAS1a was generated by ligating the coding sequence into the pBI121-SRDX vector. All constructs were verified by sanger sequencing. Transgenic plants were generated by Agrobacterium (GV3101)-mediated transformation of leaf disks as previously published[38].
Cross-sectioning and histological staining
-
The 7th and 13th internodes from 2-month-old plants were collected for stem cross-sectioning. Sections were stained with 0.1% phloroglucinol solution (10 mL pure ethanol, 1.6 mL HCl, 0.01 g phloroglucinol) for 10 min. Images of the cross-sections were captured using an Olympus BX53 microscope.
Quantification of lignin and hemicellulose
-
The whole 12th−20th stem internodes of 2-month-old WT and PagAS1a transgenic plants were collected for quantitative analyses of lignin and hemicellulose contents. The plant materials were baked at 80 °C for 48 h. 0.01 g of dried material from each sample was used to quantify lignin content. Lignin was extracted using the Lignin Content Detection Kit (Solarbio, BC4205), and the absorbance (A) values of the sample and the blank control were determined with a microplate reader at 280 nm to calculate the lignin content using the following formula: Lignin content = 1.3105 × ΔA/W, where A is the absorbance value and W is the sample weight. 0.02 g of dried material from each sample was used to quantify hemicellulose. Hemicellulose was extracted using the Hemicellulose Content Kit (Geruisi, G0716W48), and the absorbance (A) values of the sample, standard, and blank control was determined with a microplate reader at 460 nm to calculate the hemicellulose content using the following formula: Hemicellulose content = 2.61 × ΔA/(A standard × A blank)/W, where A is the absorbance value and W is the sample weight.
Transient expression assay
-
Each effector and reporter plasmid was independently transformed into Agrobacterium (GV3101). Agrobacterium transformed with the indicated effector and reporter was cultured to an optical density (OD600) of 0.8, mixed at a ratio of 1:1, and then infiltrated into Nicotiana benthamiana (tobacco) leaves. The firefly luciferase (LUC) and Renilla luciferase (REN) activities were measured after 48 h incubation.
RNA sequencing (RNA-seq) and data analysis
-
The 7th to 12th internodes of whole stems were collected from 2-month-old WT and PagAS1a-OE (OE11) transgenic plants for RNA extraction. The Illumina X Ten platform was used for 150 bp paired-end RNA sequencing. The genome assembly version 3.0 of Populus trichocarpa was used in RNA-seq analysis. Clean sequencing reads were mapped to the genome using hisat2 with default parameters[39], the mapped reads were counted using htseq-count[40], and significantly differentially expressed genes (DEGs) were identified using the edgeR package[41] with a Fisher's exact test FDR of less than 0.05. Each sequencing sample included three biological replicates.
Accession number
-
Accession numbers for genes used in this study are as follows: PtrAS1a (Potri.004G102600), PtrAS1b (Potri.017G112300), PtrAS1c (Potri.006G085900), PagAS1a (Pag.A04G001079), PagAS1b (Pag.B17G000462), PagAS1c (Pag.A06G002169), AtAS1 (AT2G37630), OsAS1 (LOC_Os12g38400), ZmAS1 (Zm.01G232800), TaAS1a (Traes_5BS_E635B0A281), TaAS1b (Traes_5DS_8F3BD4450), TaAS1c (Traes_5BS_E635B0A281), BpAS1a (BPChr11G06937), BpAS1c (BPChr14G12721), MdAS1a (MD17G1073900), MdAS1c (MD12G1142500), SpAS1a (SapurV1A.0101s0440), SpAS1c (SapurV1A.0001s1780), TcAS1a (Thecc1EG019507t1), TcAS1c (Thecc1EG021932t1), GmAS1a (Glyma.03G081900), GmAS1c (Glyma.18G181300), SlAS1 (Solyc09g010840).
-
MYB transcription factors have a highly conserved MYB domain, which is composed of multiple incompletely repeated amino acid sequences (R). Based on the number of R repeats, MYB transcription factors are divided into four categories: 1R-MYB, R2R3-MYB, R1R2R3-MYB, and 4R-MYB[42]. The main MYB transcription factors in plants are R2R3-MYB transcription factors[43]. In our previous RNA-seq analysis of differentially expressed genes (DEGs) between the secondary xylem and secondary phloem in Populus, we found that the PagAS1a, an ortholog gene of AS1, expressed significantly higher in the secondary phloem than in the secondary xylem (Supplementary Fig. S1)[44]. Sequence analysis showed that PagAS1a is a R2R3-MYB transcription factor which contains highly conserved R2R3 repeats in the DNA-binding domain in the N-terminal region (Fig. 1a). To further characterize the PagAS1a gene, we evaluated the expression of PagAS1a in xylem, phloem, cortex, root, leaf, petiole, 1st to 4th internodes, and 9th to 12th internodes by RT-qPCR (Fig. 1b). The results showed that PagAS1a was expressed to varying degrees in different tissues and the expression level was highest in the phloem, which was consistent with the RNA-seq results.
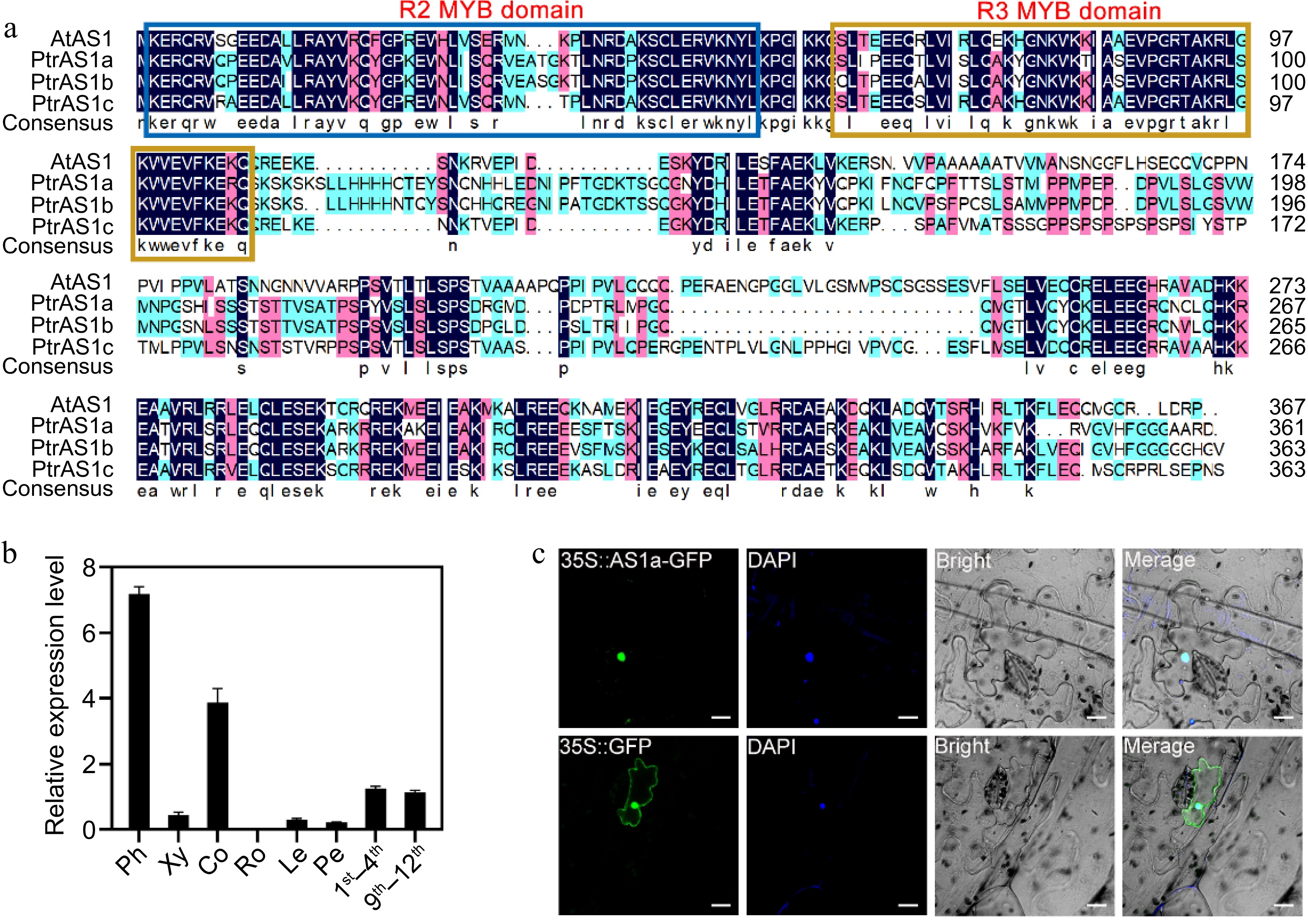
Figure 1.
Characterization of PagAS1a. (a) Multiple sequence alignment of Populus and Arabidopsis AS1 proteins. (b) PagAS1a expression levels in various tissues assayed by RT-qPCR. Ph, phloem; Xy, xylem; Co, cortex; Ro, root; Le, leaf; Pe, petiole; 1st−4th, first to fourth stem internodes; 9th−12th, ninth to twelfth stem internodes. (c) Subcellular localization of PagAS1a. The nucleus was confirmed by DAPI staining. Scale bar: 20 μm.
To examine the subcellular localization of PagAS1a, we constructed a vector expressing the PagAS1a-GFP fusion protein and transiently expressed it in tobacco leaves. Our results showed that the PagAS1a-GFP fusion protein was specifically distributed in the nucleus, while the GFP alone was distributed in both nucleus and cytoplasm (Fig. 1c), indicating that PagAS1a is a nuclear-localized protein.
Phenotype of AS1a overexpression and dominant repression transgenic plants
-
To functionally characterize the PagAS1a gene, we generated its overexpression transgenic plants (PagAS1a-OE) under the control of the cauliflower mosaic virus (CaMV) 35S promoter in Populus. We also employed the chimeric repressor silencing technology (CRES-T) to construct PagAS1a dominant repression lines (PagAS1a-SRDX) by fusing the PagAS1a coding sequence with the well-defined SRDX repressor domain (PagAS1a-SRDX) and transformed it into Populus. Two representative lines of PagAS1a-OE (OE11, OE24) and PagAS1a-SRDX (DR24, DR28) were selected for further analysis (Fig. 2a). The line OE11 exhibited the highest PagAS1a expression level, which was 279.6-fold increase relative to WT, while the line OE24 displayed a 25.1-fold increase relative to WT. The PagAS1a expression level in dominant repression lines DR24 and DR28 was elevated 102-fold and 61-fold relative to WT, respectively (Fig. 2b). Compared to WT, the PagAS1a-OE and PagAS1a-SRDX lines displayed significantly greater plant height (Fig. 2c). The differences were positively correlated with PagAS1a expression levels in different transgenic lines. No significant differences in leaves were observed between WT and PagAS1a transgenic plants (Supplementary Fig. S2). Notably, PagAS1a-SRDX plants exhibited a significantly greater number of internodes than both WT and PagAS1a-OE plants (Fig. 2d). These results suggested that PagAS1a overexpression and dominant repression have both similar and divergent functions in regulating plant growth.
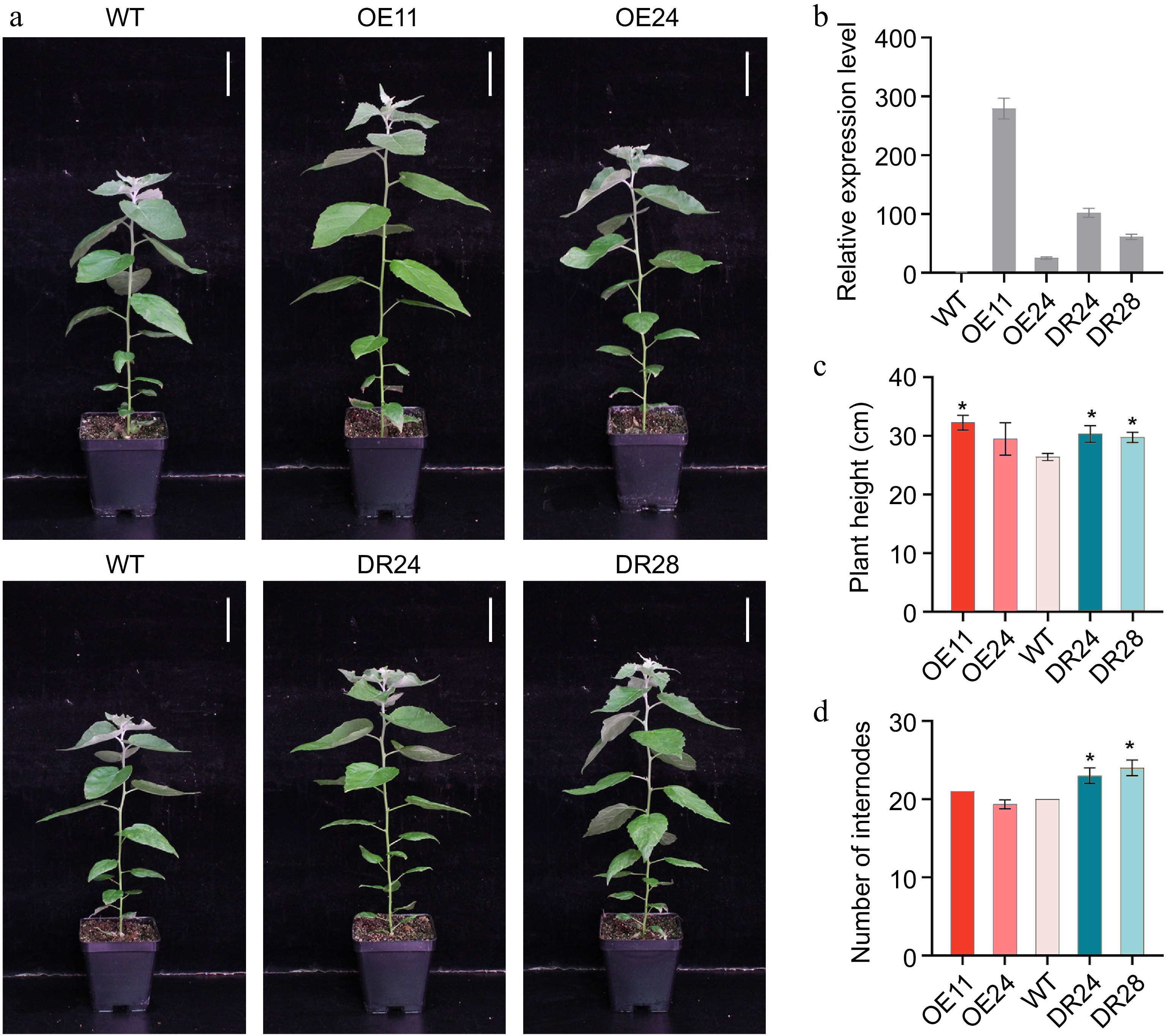
Figure 2.
Phenotype of PagAS1a overexpression and dominant repression transgenic plants. (a) Morphological phenotype of 2-month-old PagAS1a-OE transgenic plants (OE11, OE24) and PagAS1a-SRDX transgenic plants (DR24, DR28) compared with WT. Scale bar: 4 cm. (b) Expression levels of PagAS1a in PagAS1a-OE and PagAS1a-SRDX plants determined by RT-qPCR. (c) Plant height, and (d) number of internodes of transgenic lines and wild type (WT). Error bars represent SD. Asterisks indicate significant differences between WT and transgenic lines (Student's t-test): * p < 0.05.
Both AS1a overexpression and dominant repression plants promote xylem development
-
To further dissect the function of PagAS1a during secondary growth in Populus, we performed cross-section analyses of stems from WT, PagAS1a-OE (OE11, OE24), and PagAS1a-SRDX (DR24, DR28) plants. At the 7th internode, the PagAS1a transgenic plants showed enhanced lignification compared to WT, with the most pronounced increase observed in OE11, the line exhibiting the highest transgene expression level (Fig. 3a). At the 13th internode, the PagAS1a transgenic plants exhibited significantly wider stem diameter and secondary xylem compared to WT (Fig. 3a). Quantitative analyses showed that the stem diameter (Fig. 3b), xylem width (Fig. 3c), and xylem area/stem cross-sectional area (Fig. 3d) of PagAS1a-OE (OE11, OE24) and PagAS1a-SRDX (DR24) plants were significantly increased compared to WT. There were no significant differences in secondary phloem between WT and PagAS1a transgenic plants (Supplementary Fig. S3). We also quantified the content of lignin and hemicellulose, which are two main components of the secondary cell wall (SCW) in the secondary xylem. Consistent with the enhanced secondary xylem development, our results showed that both lignin and hemicellulose levels were significantly elevated in PagAS1a transgenic plants (Supplementary Fig. S4). Notably, the xylem/stem area ratio and lignin content in PagAS1a-SRDX line DR28, which had the low transgene expression level, were similar to WT (Fig. 3b−d). Together, these results suggested that both PagAS1a overexpression and dominant repression function similarly in promoting xylem development.
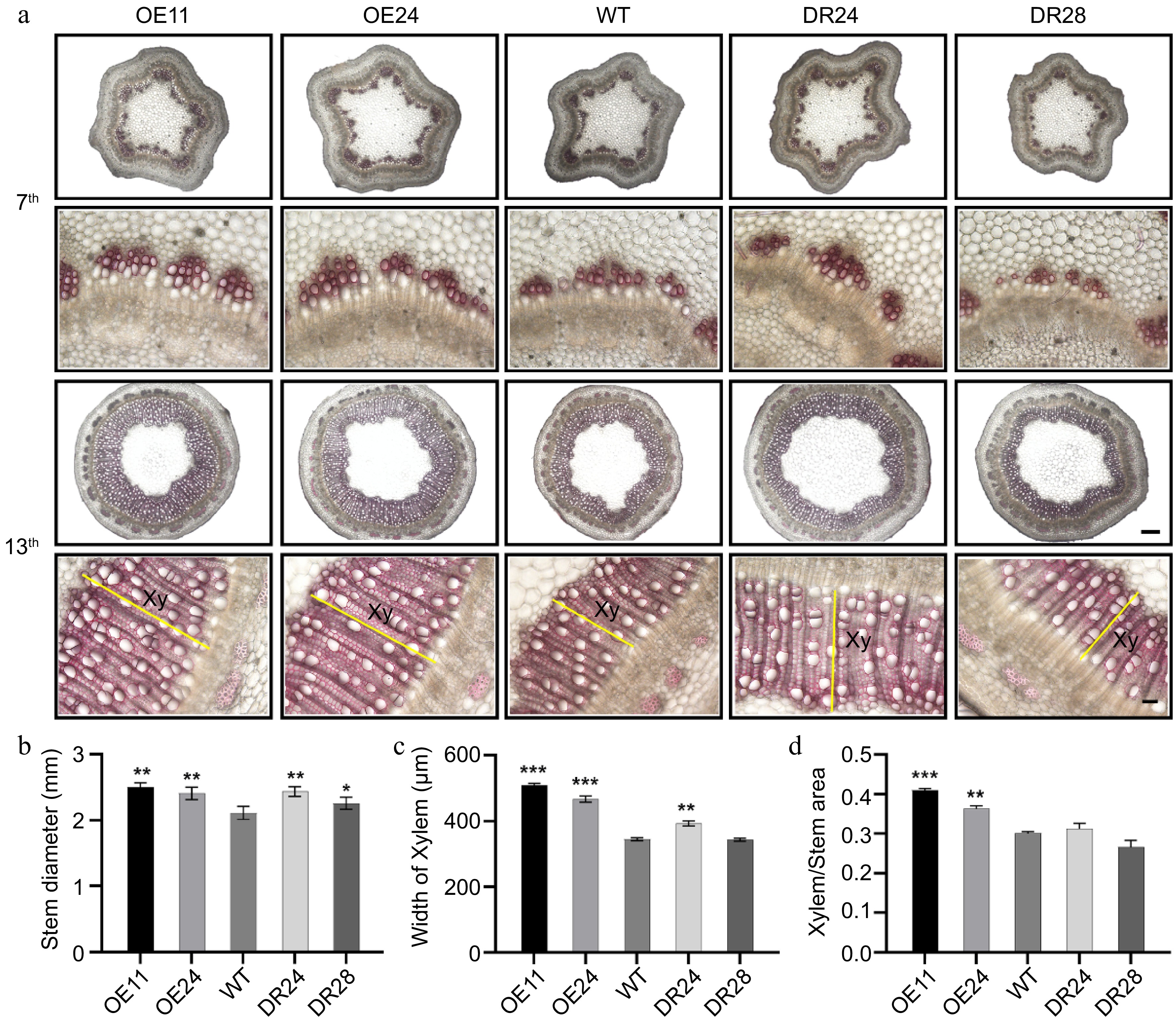
Figure 3.
Both PagAS1a overexpression and dominant repression plants promote xylem development. (a) Cross-section of the 7th and 13th stem internodes of WT, PagAS1a-OE, and PagAS1a-SRDX plants stained with 0.1% phloroglucinol solution, yellow lines label the secondary xylem (Xy) region. Scale bars: 300 μm for upper panels and 50 μm for lower panels. (b) Stem diameter of WT, PagAS1a-OE (OE11, OE24), and PagAS1a-SRDX (DR24, DR28) plants. Quantification of (c) xylem width, and (d) xylem area/stem cross-sectional area shown in (a). Data in (b)−(d) were collected from at least three individual plants for WT and transgenic lines. Asterisks indicate significant differences between WT and transgenic lines (Student's t-test: * p < 0.05; ** p < 0.01; *** p < 0.001.
PagAS1a is a transcriptional repressor
-
Since the PagAS1a overexpression and dominant repression plants exhibited similar phenotypes in plant growth and xylem differentiation, we speculated that PagAS1a functions as a transcriptional repressor. To test this hypothesis, we performed transient expression assays by co-transfection of the reporter and various effector constructs in tobacco leaves (Fig. 4a). As shown in Fig. 4b, the relative luciferase activity in samples transfected 35S::AS1a was significantly lower than in samples transfected the vector control (CK), suggesting that PagAS1a acts as a transcriptional repressor. To further confirm this result, we fused the PagAS1a with VP16 (the activation domain from Herpes simplex virus). The relative luciferase activity in samples transfected 35S::AS1a-VP16 was significantly lower than in samples transfected 35S::VP16, supporting the conclusion that PagAS1a functions as a transcriptional repressor.
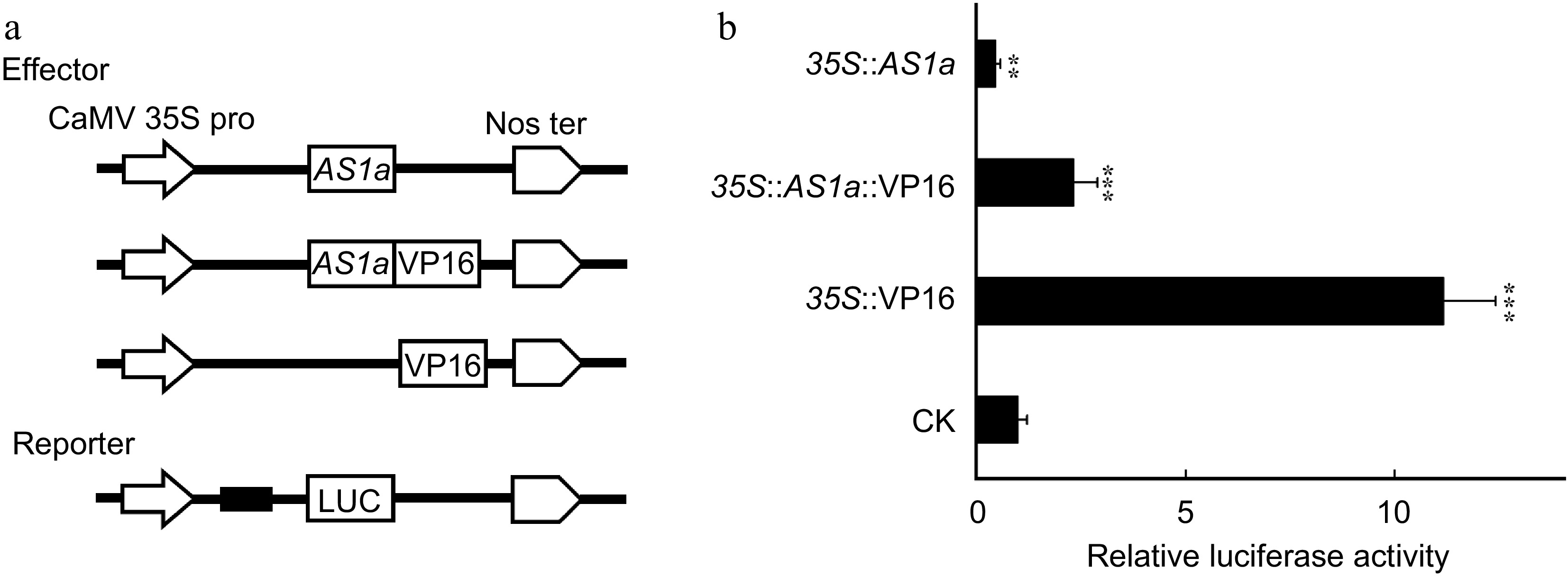
Figure 4.
PagAS1a is a transcriptional repressor. The PagAS1a construct driven by the CaMV 35S promoter was used as the effector. The relative luciferase activity was measured 48 h after injection. The value of the mock group injected with the reporter and corresponding empty effector construct was set to 1. Student's t-test was performed for significance with three biological replications (** p < 0.01; *** p < 0.001).
Transcriptome analysis of PagAS1a-OE plants
-
To identify genes regulated by PagAS1a during secondary growth, we collected whole stems from the 7th to the 12th internodes of PagAS1a-OE (OE11) and WT plants for RNA sequencing (RNA-seq). A total of 2,736 DEGs were identified between OE11 and WT (Supplementary Table S2), of which 1,669 genes were significantly up-regulated and 1,067 genes were significantly down-regulated in OE11. GO analysis revealed that 220 and 163 biological pathways (BP) were significantly enriched in the up-regulated and down-regulated DEGs, respectively (Supplementary Tables S3, S4). Consistent with the increased plant growth and advanced xylem development in PagAS1a-OE plants, the up-regulated DEGs in OE11 were significantly enriched in pathways including carbon fixation (GO:0015977), photosynthesis (GO:0015979, GO:0019685), lignin biosynthetic process (GO:0009809), flavonoid metabolic process (GO:0009812), cellulose biosynthetic process (GO:0030244), and flavonoid biosynthetic process (GO:0009813) (Fig. 5a; Supplementary Table S3). In contrast, the down-regulated DEGs in OE11 were significantly enriched in pathways such as the lignin catabolic process (GO:0046274), phenylpropanoid catabolic process (GO:0046271), and cell cycle-related categories (GO:0044843, GO:0007346, GO:0000079) (Fig. 5b; Supplementary Table S4). Detailed inspection showed that lignin biosynthetic genes, including CAD9, C3H3, LAC11, and LAC17[45−48], were significantly up-regulated in OE11 (Fig. 5c; Supplementary Table S5). Similarly, cellulose biosynthesis genes, including CESA6 and IRX1[7,49], were also significantly up-regulated in OE11 (Fig. 5d; Supplementary Table S5). In contrast, genes involved in lignin catabolic pathways, including LAC3, LAC5, and LAC12[21,50−54], were significantly enriched in down-regulated in OE11 (Fig. 5e; Supplementary Table S5). Interestingly, genes associated with cell fate specification, including YAB1, YAB2, and CPC[55−58], were also significantly enriched in down-regulated in OE11 (Fig. 5f; Supplementary Table S5). We also inspected the expression of genes related to vascular cambium maintenance and secondary phloem/xylem differentiation. The key regulatory genes PtaLBD1, PtWOX4a, CLE41b, and XND1c, were significantly down-regulated in OE11[59−61], while PtrHB8 and XCP1 were significantly up-regulated in OE11[25,62,63] (Supplementary Fig. S5). Together, the transcriptome analysis supported the observation that overexpression of PagAS1a promotes plant growth and secondary xylem development.
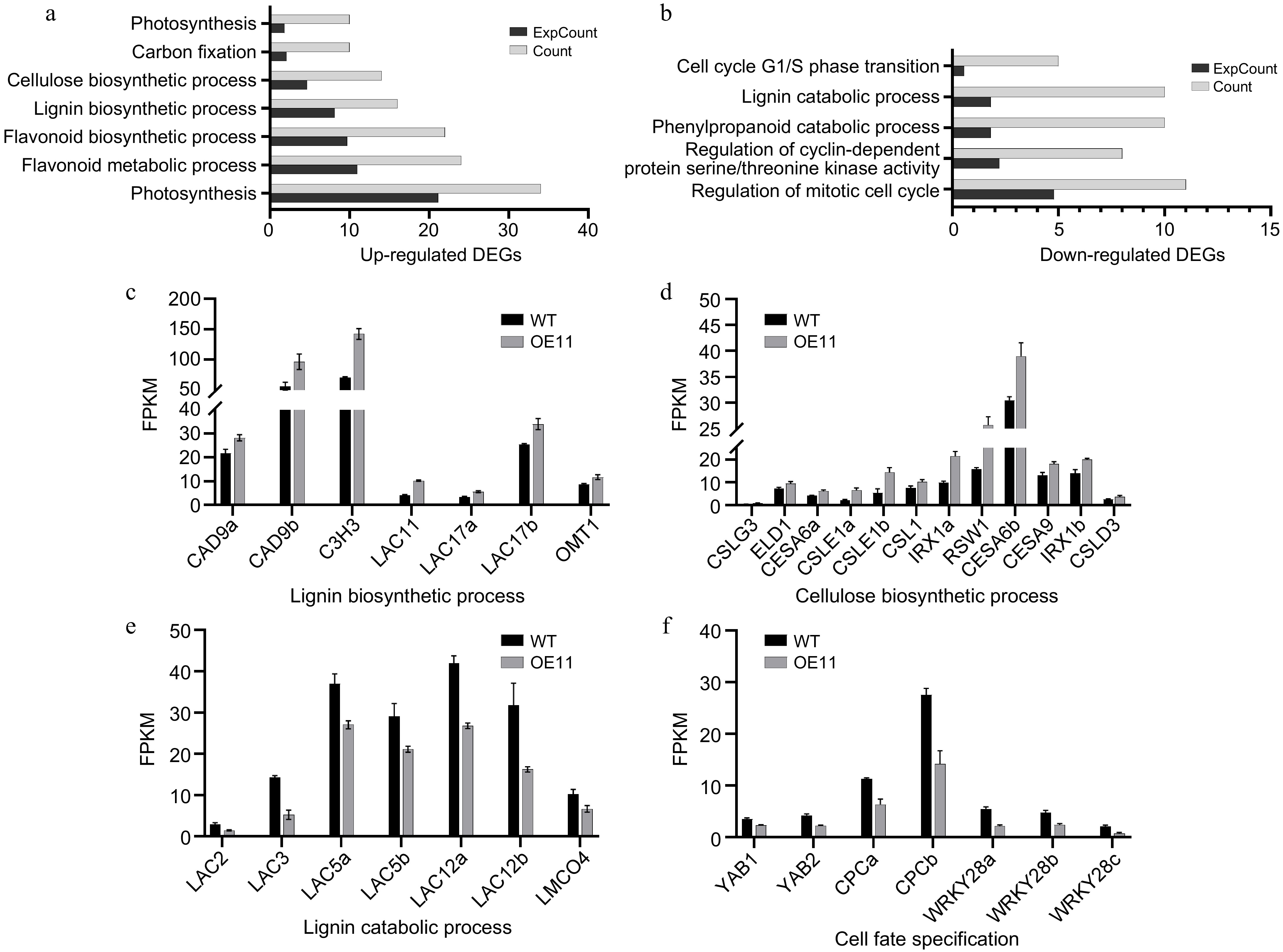
Figure 5.
Transcriptome analysis of PagAS1a overexpression plants. Significantly enriched GO categories in the (a) up-regulated, or (b) down-regulated DEGs in PagAS1a-OE transgenic plants (OE11). Expression of DEGs involved in (c) lignin biosynthetic process, (d) cellulose biosynthetic process, (e) lignin catabolic process, and (f) cell fate specification. The average fragments per kilobase of exon per million fragments mapped (FPKM) for each gene from RNA-seq experiments are shown.
Phylogenetic analysis of AS1 ortholog proteins in plants
-
We identified three AS1 orthologs genes in Populus: PagAS1a (Potri.004G102600), PagAS1b (Potri.017G112300), and PagAS1c (Potri.006G085900). To elucidate the evolutionary relationships among AS1 orthologs, we identified and isolated 28 AS1 ortholog proteins from 12 representative species, including three monocot species (Zea mays, Oryza sativa, and Triticum aestivum) and nine dicot species. The dicot species included three herbaceous species (Glycine max, Solanum lycopersicum, and Arabidopsis thaliana) and six woody species (Betula platyphylla, Malus domestica, Populus alba × Populus glandulosa (84K), Populus trichocarpa, Salix purpurea, and Theobroma cacao). We then constructed a phylogenetic tree using the full-length protein sequences with the neighbor-joining method. The results revealed that these 28 AS1 orthologs were divided into two subgroups (Fig. 6). Interestingly, subgroup I contain AS1 orthologs from all species, while subgroup II only contains AS1 orthologs from woody plants, indicating that there might be functional divergence between these two subgroups. Furthermore, a clear boundary was observed between monocot and dicot species within the subgroup I. These results indicate that the AS1 ancestral gene existed before the divergence of monocot and dicot species.
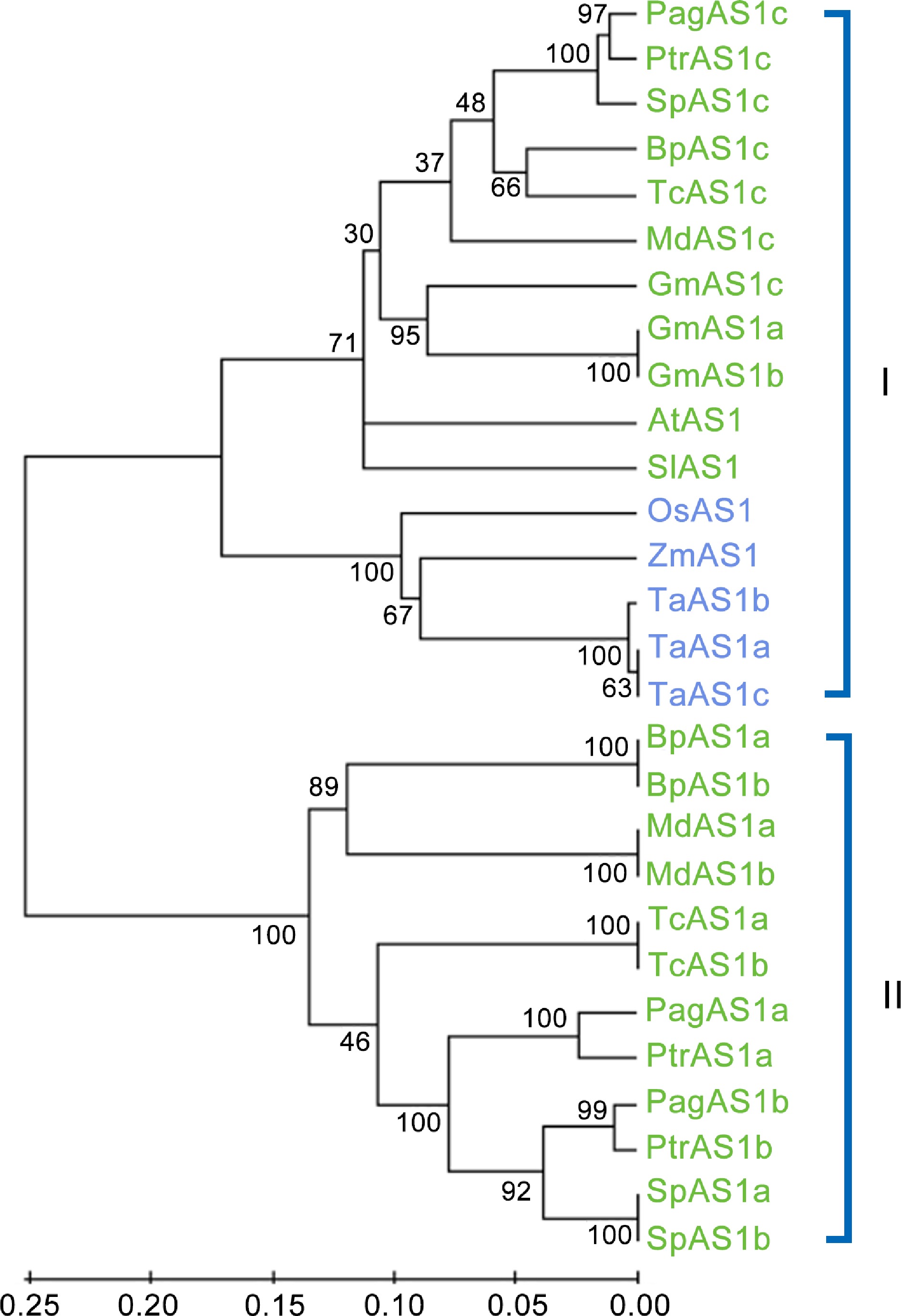
Figure 6.
Phylogenetic analysis of AS1 ortholog genes from 12 representative species. The neighbor-joining (NJ) tree was constructed based on the alignment of full-length protein sequences. MEGA 5.0 was used with 1,000 bootstrap replicates. Dicot species and monocot species are represented in green and blue fonts, respectively.
-
Polarity establishment plays a key role in lateral organ formation. In leaves, AS1 and class III HD-ZIPs serve as essential adaxial regulators for the establishment of leaf adaxial-abaxial polarity[32,64]. Similarly, in stem secondary growth, the differentiation of the secondary phloem (outward) and secondary xylem (inward) from the vascular cambium represents another form of polarity establishment. Previous studies have shown that several class III HD-ZIPs transcription factors are important regulators of vascular cambium cell maintenance and differentiation during secondary growth in Populus[65−68], suggesting that plants may employ a similar set of genes to establish polarity in both leaves and stems.
There are three AS1 orthologs in total, PagAS1a (Potri.004G102600), PagAS1b (Potri.017G112300), and PagAS1c (Potri.006G085900) in Populus, all of which encode proteins with a conserved R2R3 MYB domain (Fig. 1a). Our previous transcriptome analysis of secondary phloem and secondary xylem showed that all these three AS1 ortholog genes expressed significantly higher on the secondary phloem side, and the PagAS1a has relatively higher expression than PagAS1c but lower than PagAS1b (Supplementary Fig. S1). It should be noted that we collected samples by peeling the bark, then scraped the bark side as the secondary phloem (which included most cambium cells), and the stem side as the secondary xylem. Therefore, the secondary phloem samples in our transcriptome data were mixed with cambium samples. Further analysis with transcriptome data from AspWood showed that PagAS1b was highly expressed in the secondary phloem and cambium zone, while PagAS1c was expressed across the whole wood forming zone except the expanding xylem region (Supplementary Fig. S1)[69]. For unknown reasons, PagAS1a was not included in the AspWood dataset. In the future, carrying out RNA in-situ hybridization and native promoter-driven reporter gene analysis would be helpful to characterize the native expression of these genes more precisely.
We selected PagAS1a for further characterization in Populus. As expected, PagAS1a is localized in the nucleus (Fig. 1c). We obtained PagAS1a overexpression and dominant repression plants for functional analysis. As shown in Fig. 2, both PagAS1a overexpression and dominant repression transgenic plants displayed significantly faster growth than WT, suggesting that PagAS1a and PagAS1a-SRDX function similarly in regulating poplar growth. These results suggest that PagAS1 may function as a transcriptional repressor as PagAS1a-SRDX. We also performed cross-sectional analysis to determine whether PagAS1a regulates stem secondary growth. The results showed that secondary xylem development was significantly advanced in PagAS1a transgenic plants compared to WT (Fig. 3). Transcriptome analysis revealed that genes related to lignin and cellulose biosynthesis were significantly enriched in up-regulated DEGs in PagAS1a overexpression plants (Fig. 4), supporting the observed increase in secondary xylem development. Since fiber and vessel elements in the secondary xylem play essential roles in mechanical support and water conduction from roots to crown, the enhanced secondary xylem development may be the primary reason for the increased plant growth in PagAS1a transgenic plants. Notably, all functional analyses in this study were performed with PagAS1a overexpression and dominant repression transgenic plants, both of which were driven by the 35S promoter. The dominant repression of PagAS1a could cause functional interference of its interacting proteins. Therefore, the phenotype of PagAS1a transgenic plants may be partially due to PagAS1a ectopic expression and functional repression of PagAS1a interacting proteins. However, the significantly increased plant growth and secondary xylem development suggest that PagAS1 genes are excellent candidate genes for breeding fast-growth trees with high wood production. In the future, it will be important to generate CRISPR/Cas9-mediated PagAS1a, PagAS1b, and PagAS1c single, double, or triple knockout mutants to further explore the functions of each AS1 ortholog gene and their potential functional redundancy in Populus.
We also performed transcriptome analysis of gene expression changes caused by PagAS1a overexpression. Our results showed that the lignin biosynthetic process (GO:0009809) was significantly enriched in up-regulated DEGs while the lignin catabolic process (GO:0046274) was significantly enriched in down-regulated DEGs (Fig. 5a, b; Supplementary Tables S3, S4), which are consistent with the increased secondary xylem development and lignin content in PagAS1a overexpression plants (Fig. 3; Supplementary Fig. S4). We also carefully inspected the expression of genes related to cambium cell activity, and secondary phloem/xylem differentiation (Supplementary Fig. S5). Our results showed that the PagLBD1, PtWOX4a, CLE41b, and XND1c genes, which participate in regulating vascular cambium cell maintenance and secondary phloem/xylem differentiation, were significantly down-regulated in OE11 (Supplementary Fig. S3; Supplementary Table S6)[59,60,62], while PtrHB8 (promotes xylem development) and XCP1 (specifically expressed in xylem vessel cells) were significantly up-regulated in OE11 (Supplementary Fig. S3; Supplementary Table S6)[25,62,63]. In Arabidopsis, AS1 is a key regulator in the establishment of leaf polarity, and AS1 forms a protein complex with the LOB domain transcription factor AS2 (AS1−AS2) and directly represses the expression of class I KNOX homeobox genes, which is critical for the establishment of leaf primordium[29]. In our transcriptome analysis, the class I KNOX homeobox genes, including ARK1, ARK2, and KNAT2/6b, which are important regulators of cambium cell maintenance and differentiation[61,70,71], were down-regulated in PagAS1a overexpression plants (Supplementary Table S6). These results suggest that AS1 ortholog proteins may employ a similar regulatory module to promote cambium cell differentiation into xylem cells. In the future, it will be important to identify the direct target genes of PagAS1 through further experiments, as this will enhance our understanding of the molecular mechanisms underlying its function. Additionally, since AS1 functions in a protein complex with AS2 and other proteins in regulating leaf development, it will be informative to isolate PagAS1a-interacting proteins to reveal the molecular mechanisms underlying its function.
Phylogenetic analysis of AS1 orthologs from 12 species revealed that they are divided into two distinct subgroups (Fig. 6). Subgroup I contains AS1 orthologs from both monocot and dicot (including herbaceous and woody) species, while subgroup II exclusively contains orthologs from woody dicot species. This result indicates that subgroup I may have more conserved functions during evolution, whereas subgroup II expanded more recently and potentially gained new functions (Fig. 6). As there are no significant differences in PagAS1a transgenic plant leaves (Supplementary Fig. S2), it would be interesting to test whether PagAS1c, which belongs to subgroup I and has a closer distance to Arabidopsis AS1, functions in regulating leaf polarity establishment. In Populus, PagAS1a, and PagAS1b are two closely related orthologs belonging to subgroup II, suggesting that there might be functional redundancy between them. Notably, orthologs from monocots and dicots are divided into two subclusters within subgroup I, indicating that the AS1 ancestral gene likely existed before the divergence of monocots and dicots (Fig. 6).
-
In this study, we identified the AS1 orthologs in Populus. Phylogenetic analysis indicated that the three PagAS1 ortholog genes may have both common and divergent functions in Populus. Functional characterization of PagAS1a demonstrated that increasing its expression significantly promotes secondary xylem development and plant growth, providing a valuable candidate gene for breeding fast-growing trees for wood production. Notably, there were no significant morphological differences in leaves between PagAS1a transgenic plants and WT. It will be interesting to dissect whether PagAS1b or PagAS1c has a more prominent effect on leaf polarity and conduct functional comparison studies between PagAS1a, PagAS1b, and PagAS1c in the future.
This work was supported by the Natural Science Foundation of Shandong (ZR2022MC140), and the National Key R&D Program of China (2021YFD2200800).
-
The authors confirm contribution to the paper as follows: generating the transgenic plants and performing phenotypic analysis: Li YZ, Li YL, Zhang B; data analysis: Li YZ, Liu L, Zhang B; writing the manuscript: Li YZ, Wu A, Liu L. All authors reviewed the results and approved the final version of the manuscript.
-
The original RNA-sequencing data have been deposited into the Genome Sequence Archive at National Genomics Data Center, China National Center for Bioinformation/Beijing Institute of Genomics, Chinese Academy of Sciences (GSA: CRA024667) that are publicly accessible at https://ngdc.cncb.ac.cn/gsa.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primers used for gene cloning, vector construction and qPCR.
- Supplementary Table S2 Summary of DEGs between the wild type and PagAS1aOE.
- Supplementary Table S3 Gene Ontology (GO) analysis of the up-regulated DEGs from RNA-seq (PagAS1aOE).
- Supplementary Table S4 Gene Ontology (GO) analysis of the down-regulated DEGs from RNA-seq (PagAS1aOE).
- Supplementary Table S5 Gene expression levels associated with lignin, cellulose, cell fate specification from RNA-seq (PagAS1aOE).
- Supplementary Table S6 Gene expression levels associated with cambium from RNA-seq (PagAS1aOE).
- Supplementary Fig. S1 Analysis of expression patterns of PtrAS1a/b/c genes.
- Supplementary Fig. S2 Comparative analysis of leaves between WT and PagAS1a transgenic lines. Leaf and cross-sectional pictures of internode 6th of the plants. Scale bars: 0.5 cm for upper panels and 50 µm for lower panels.
- Supplementary Fig. S3 Phloem width identification of PagAS1a transgenic plants and WT.
- Supplementary Fig. S4 Determination of lignin and hemicellulose contents.
- Supplementary Fig. S5 Gene expression levels associated with cambium in WT and PagAS1a overexpression transgenic line (OE11) from RNA-seq.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li Y, Li Y, Zhang B, Wu A, Liu L. 2025. The ASYMMETRIC LEAVES1 ortholog PagAS1a promotes xylem development and plant growth in Populus. Forestry Research 5: e010 doi: 10.48130/forres-0025-0011 

The ASYMMETRIC LEAVES1 ortholog PagAS1a promotes xylem development and plant growth in Populus
- Received: 24 January 2025
- Revised: 15 April 2025
- Accepted: 14 May 2025
- Published online: 23 May 2025
Abstract: ASYMMETRIC LEAVES1 (AS1) is a central regulator of leaf polarity establishment in Arabidopsis. However, the functions of its ortholog genes in Populus are unclear. In this study, we performed gene expression analysis and found that the AS1 ortholog gene PagAS1a exhibited significantly higher expression levels in the secondary phloem than in the secondary xylem within the wood formation zone. Subcellular localization analysis showed that PagAS1a localizes in the nuclei. Compared to the wild type, both PagAS1a overexpression (PagAS1a-OE), and dominant repression (PagAS1a-SRDX) plants exhibited enhanced height and stem diameter growth. Analysis of stem cross-sections revealed that both overexpression and dominant repression of PagAS1 promoted secondary xylem development. We performed transcriptome sequencing (RNA-seq) for gene expression analysis, and identified 2,736 significantly differentially expressed genes (DEGs) in PagAS1a-OE plants. GO analysis revealed that genes associated with photosynthesis, lignin biosynthetic, and cellulose biosynthetic pathways were significantly enriched in up-regulated DEGs in PagAS1a-OE, while genes associated with lignin catabolic, and phenylpropanoid catabolic pathways were significantly enriched in down-regulated DEGs. Together, our results suggest that the AS1 ortholog PagAS1a is an important regulator of secondary xylem development and plant growth in Populus.
-
Key words:
- Populus /
- Xylem development /
- Plant growth /
- PagAS1a /
- MYB domain transcription factor