









-
Cultured meat presents a promising alternative for meat consumption. The production of cultured meat involves the isolation and expansion of animal cells, followed by myogenic or adipogenic differentiation, and subsequent food processing[1,2]. This innovative technology offers consumers a favorable perception regarding animal welfare, food safety, and sustainable development[3,4].
Satellite cells (SCs) are myogenic stem cells found between the basement membrane and sarcolemma of muscle fibers in vivo[5]. Once isolated and cultivated under well-controlled conditions, SCs can proliferate and differentiate into multi-nucleated myotubes[6], which have been widely used in cultured meat research and production[7−10]. However, the dependence on animal-derived components in cell expansion and differentiation makes cultured meat hardly commercially viable[11,12]. Traditional protocols for cell culture require fetal bovine serum (FBS), which provides ample nutrition for cells and exhibits extensive versatility[13]. However, the utilization of serum raises numerous concerns, including high cost, poorly chemically defined components, batch instability, ethical concerns, and the risk of carrying pathogens[12]. Thus, it is imperative to establish a serum-free system for cultured meat production.
The culture process of SCs can be divided into two stages: proliferation and differentiation, each requiring specific media. During the last decades, multiple serum-replacements or serum-free media for different cell types have been developed, including induced pluripotent stem cells[14], embryonic stem cells[15], and haematopoietic stem cells[16]. These findings provide valuable insights for understanding the nutritional requirements of livestock stem cells and motivate the development of serum-free media for cultured meat. Recently, several serum-free proliferation media for bovine SCs or myoblasts have been reported[17−20]. However, a specific serum-free medium may not adequately support the stable expansion of cell lines from different species. Meanwhile, the understanding of specific factors in serum that stimulate and maintain cell growth remains largely incomplete. Investigations should be committed to seek the appropriate factors and environments to meet the physiological needs of SCs. Specifically, optimize growth factor combinations and concentrations for growth stimulation[21−24], supply carbon sources, amino acids, and lipids for nutritional maintenance[25], provide antioxidants for redox homoestasis[2,2627], and retain pH stabilization[12].
In terms of myogenic differentiation of SCs, the standard protocol involves a process known as serum starvation[28], indicating a lesser dependency on serum during differentiation. Chemically defined media have been reported to support the differentiation of muscle cell lines including C2C12 and bovine SCs[29–30]. However, the myogenic induction of serum-free media exhibits a lesser extent than the serum starvation approach, especially the myogenic differentiation in three-dimensional (3D) scaffolds to form meat-like constructs[29]. Furthermore, the entire serum-free system maintaining long-term proliferation and supporting efficient differentiation of SCs is still unavailable.
Like most primary cells, SCs undergo replicative senescence during the culture process, which severely impairs their normal functions[31]. This is regulated by various biological mechanisms, including cell cycle regulatory factors[32]. The CDKN2A gene encodes two splicing-isoform products, namely p16INK4A, and p14ARF, which are cell cycle repressors[33]. p16INK4A inhibits the activity of the CDK4-CDK6-cyclin D complex to suppress Rb1 phosphorylation[34]. The unphosphorylated RB1 interacts with transcription factor E2F, thereby inhibiting E2F-mediated translation and causing cell cycle arrest at the G1/S transition[35]. On the other hand, p14ARF blocks the cell cycle in both the G1 and G2 phases and inhibits the proliferation of cells by indirectly activating the p53 pathway[33]. p16INK4A is also a general marker that is expressed highly in senescent cells, and life-long removal of p16INK4A-expressing cells delayed the onset of age-related disorders, especially in adipose tissue, eye, and skeletal muscle[36]. Moreover, p16INK4A expression is specifically induced in geriatric SCs and drives geroconversion at the expense of proliferation both in humans and mice[37–38]. Therefore, targeting the CDKN2A gene locus is essential for preventing ageing or inducing cell immortalization[39]. Thus, CDKN2A gene might act as an important target for regulating cell expansion and functional maintenance in livestock SCs.
In this study, we developed a serum-free proliferation medium and establised CDKN2A−/− SCs via CRISPR/Cas-mediated genetic engineering. The serum-free medium allowed for the robust and stable expansion of CDKN2A−/− cells. Further, through optimizing the serum-free differentiation media, we established a continuous serum-free proliferation and serum-free myogenic differentiation system for cultured meat production.
-
Purified porcine SCs were isolated via cell surface markers CD31−/CD45−/CD56+/CD29+ as described previously[8]. Then SCs were cultured on collagen-coated dishes in DMEM/F12 medium (Gibco, Cat# 11320033) supplemented with 15% FBS (Sigma, Cat# F8318), 5 ng/mL bFGF (R&D, Cat# 233-FB-025) and 1% Pen-Strep (Gibco, Cat# 15140122). For serial expansion, cells were passaged every three days to maintain a density of < 60% confluence. During cell passaging, 0.25% trypsin-EDTA (Gibco, Cat# 25200072) was used to detach the cells from the culture dishes, and the digestion was terminated by adding an equal volume of culture medium. Cell number, viability, and expanding fold were measured by CountStar (Shanghai Ruiyu Biotech Co., Ltd, PML# Countstar Mira FL, USA) through Trypan Blue staining. The cell suspension and trypan blue were mixed in a 1:1 volume ratio and then added to the device, where the cell number and viability were obtained through imaging. For cell differentiation, SCs were plated onto 1% matrigel (Corning, Cat# 356234)-coated dishes in growth medium. When confluence, the medium was changed to DMEM (Gibco, Cat# C11995500BT) with 2% horse serum (Hyclone, Cat# SH30074.02) and 1% penicillin-streptomycin. Cell differentiation was induced for 5 d and committed to further experiments.
Development of serum-free proliferation medium
-
SCs were trypsinized and suspended in serum-containing growth medium to 5 × 103 cells/mL. Two hundred μL medium with 1,000 cells per well were plated onto collagen-coated 96-well plates (Coring, 3599). After 12 h, the growth medium was changed to a different medium formula with varied concentrations of lipids, non-essential amino acids (NEAA), growth factors, antioxidants, molecular inhibitors, and other supplements. Subsequently, the medium was changed every 2 d, and cells were grown for 4 d. Then cell growth was tested by high content analyzer (Molecular Devices, PML# ImageXpress Pico). Prior to the assay, the medium was replaced with fresh medium with 10 μg/mL Hoechst (Sigma, Cat# B2261) and incubated at 37 °C for 10 min. The appropriate concentrations and combinations of different constituents were determined mainly via single-factor tests.
Generation of CDKN2A knockout SCs
-
CDKN2A−/− SCs were generated using the CRISPR/Cas9 system. Single guide RNAs (sgRNAs) targeting exon 2 of the porcine CDKN2A gene (shared by both p14 and p16 proteins) were designed using the online design tool available at
http://crispr.genome-engineering.org . SgRNAs were cloned into the lentiCRISPR v2 vector (Addgene, Cat# 52961) and transfected into SCs with LipofectamineTM Transfection Reagent (Invitrogen, Cat# L3000001), according to the manufacturer's recommendation. The editing efficiency of sgRNAs was analyzed by T7 Endonuclease I (Vazyme, Cat# EN303). The sgRNA with the highest cutting efficiency was chosen for further experiments: 5'-GTCGTGTACCGGTCGGGTGAGGG-3'. The single-cell-derived MuSC clones were established via the limiting dilution method. Then, knockout clones were identified by Sanger sequencing.Gene expression analysis
-
According to the manufacturer's instruction, the RNAprep Pure Cell/Bacteria Kit (Tiangen, Cat# DP430) was used to extract RNA from cells, and cDNA was synthesized by PrimeScript™ RT Master Mix (Takara, Cat# RR036A). Quantitative RT-PCR was performed in triplicate using TB green® premix ex taq™ ii (Takara, Cat# RR820A) on an ABI QuantStudio6. The primers used in these assays were as follows:
PAX7-F, 5′-GTGCCCTCAGTGAGTTCGATT-3′,
PAX7-R, 5′-TCCAGACGGTTCCCTTTGTC-3′;
MYOD-F, 5′-GCTCCGCGACGTAGATTTGA-3′,
MYOD-R, 5′-GGAGTCGAAACACGGGTCAT-3′;
MYOG-F, 5′-AACCCCACTTCTATGACGGG-3′,
MYOG-R, 5′-TTATCTTCCAGGGGCACTCG-3′;
GAPDH-F, 5′-TGAGATCCAGGGAGCCATCA-3′,
GAPDH-R, 5′-ATGGTCAGGGGTCCGATGTA-3′.
Immunofluorescence analysis
-
Cells were washed twice with PBS after being fixed in 4% paraformaldehyde (PFA) at room temperature for 20 min or overnight at 4 °C. Samples in the 3D culture were fixed in PFA, washed by PBS and frozen with liquid nitrogen and then sliced into a thickness of 2 nm under freezing conditions. Subsequently, samples were permeabilized in 0.5% Triton X-100 in PBS for 20 min and blocked with 10% goat serum in PBS for 1 h, and then incubated with primary antibody anti-mouse myosin heavy chain (MyHC, 1:800, Merck Millipore, Cat# 05-716) at 4 °C overnight. After three consecutive washes with PBS, samples were incubated for 1 h with secondary antibody (1:1000, Alexa Fluor 594 goat-anti-mouse, Invitrogen, Cat# A11005). After washing, samples were covered with VECTASHIELD mounting medium (with DAPI, Vector Labs, Cat# H-1200) and imaged with Leica DMi8 confocal microscope. After immunofluorescence staining, the fusion index during myogenic differentiation was assessed through Adobe Photoshop (v14.0), which represents the percentage of nuclei in myotubes relative to the total number of nuclei in the sample.
Serum-free myogenic differentiation
-
For myogenic differentiation, SCs were plated onto 1% matrigel-coated dishes in serum-free growth medium. When confluence, cells were induced with different differentiation media. The regular serum starvation medium was DMEM containing 2% horse serum and 1% penicillin-streptomycin. Serum-free myogenic differentiation medium (Version 1.0) was DMEM supplemented with 0.1 × ITS-X (Gibco, Cat# 51500056), 25 ng/mL IGF-1 (Genscript, Cat# Z03017), 25 ng/mL IGF-2 (Genscript, Cat# Z03147), 0.01% BSA (Gibco, Cat# A1933) and 0.1 × lipids concentration (Gibco, Cat# 11905031). Serum-free adipogenic differentiation medium (Version 2.0) was DMEM/F12 supplemented with 1 × ITS-X, 1 ng/mL bFGF, 0.4% BSA, 50 ng/mL IGF-1, 50 μM Asc-2P (Sigma, Cat# 49752), 10 μM Y-27632 2HCl (Selleck, Cat# S1049), 50 ng/mL EGF (Genscript, Cat# Z02691), 1 μM dexamethasone (Selleck, Cat# S1322), 2 μM SB-431542 (Selleck, Cat# S1067), 100 μM 3-isobutyl-1-methylxanthine (Abmole, Cat# M6804), 10 μg/mL Insulin (Abmole, Cat# M9336), 2 μM rosiglitazone (Abmole, Cat# M1894). The modified serum-free myogenic differentiation medium (Version 3.0) was supplemented with 10 μg/mL insulin. The modified serum-free adipogenic differentiation medium (Version 4.0) refers to the removal of dexamethasone, 3-isobutyl-1-methylxanthine, and rosiglitazone from Version 2.0.
3D cultivation of CDKN2A−/− SCs using peanut wire-drawing protein as scaffold
-
Peanut wire-drawing protein scaffolds were soaked in sterile water at room temperature for 5 min to completely expand. Absorbent paper was used to remove excess water from the surface of scaffolds. The peanut wire-drawing protein was cut into 1 cm × 1 cm × 1 mm pieces and soaked in 75% ethanol solution for 2 h to keep sterile. Then, the scaffolds were washed three times with PBS and put into serum-free proliferation medium (A19) for use. For cell seeding, scaffolds were transferred to six-well plates, and 10 μl cell suspension with a cell density of 5 × 105 cells/μl was evenly inoculated on scaffolds. Then, scaffolds were placed in a 37 °C incubator for 2 h to facilitate cell adhesion. After that, A19 medium was added into wells to begin the 6-d proliferation process and the medium was changed every 2 days. After the proliferation process, the A19 medium was replaced with serum-free differentiation medium (Version 4.0). The medium was changed every 2 days. 3D cell culture was finished after 4 days of differentiation. Cells in the 3D culture were collected on day 0 and day 6 of proliferation and day 4 of differentiation for immunofluorescence analysis, gene expression analysis, and western blot analysis.
Texture analysis of CDKN2A−/− SCs-derived cultured meat
-
Samples of 3D culture from proliferation day 6 and differentiation day 4 were collected for Texture Profile Analysis (TPA) tests. Blank peanut wire-drawing protein was used as negative control and pork as positive control. Texture Analyzer TA-XT2i (Stable Micro Systems Ltd., Godalming, UK) with the application program provided with the apparatus (Texture Expert for Windows, Version 1.0) was used. Parameters are as follows: a P/50 probe (diameter 5 cm), pretest speed 5.0 mm/s, test speed 1.0 mm/s, posttest speed 5.0 mm/s, compression ratio 75%, trigger force 5 g, and data acquisition rate 200 pulses per second.
Western blot analysis
-
Cells in the 3D culture were shredded by a tissue grinder and lysed with RIPA buffer. Protein concentration was detected by a BCA protein assay kit (Thermo, CAT# 23225). Protein lysates were subjected to the 4%-20% SDS-PAGE gel (GenScript, CAT# M00657), and pre-stained protein standard (GenScript, CAT# M00624) was used as a marker. Then, protein lysates were electro-transferred to a PVDF membrane. After blocking with 5% skimmed milk powder in TBST (Solarbio, CAT# T1085), the membrane was incubated with appropriate dilutions of primary antibody for 16 h at 4 °C and secondary antibodies for 2 h at room temperature. Primary antibodies: anti-MYHC (1:1,000) and MYOG (1:1,000, BD pharmingen, Cat# 556358). Then imaging was performed using ImageQuant LAS 4000 (GE).
Statistical analysis
-
Statistical analyses were performed using one-way ANOVA, two-way ANOVA, or two-tailed Student's t-test as appropriate, and the p value was calculated. Statistical significance was defined as * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001. If not otherwise stated, error bars indicate standard deviation (SD).
-
Based on prior research evidence (Supplementary Table S1) and empirical iterative strategies, we designed and optimized the serum-free proliferation media step by step (Fig. 1a). To determine essential factors with optimal combinations and concentrations, purified porcine SCs were seeded onto 96-well plates and analyzed via high content analyzer (Fig. 1a). In Formula 1, basal media DMEM/F12 was supplemented with factors that are demonstrated to be essential for SCs survival and proliferation: nutritional and supporting factors insulin-transferrin-selenium-ethanolamine (ITS-X) and bovine serum albumin (BSA), ROCK inhibitor Y-27632, growth factors basic fibroblast growth factor (bFGF), epidermal growth factor (EGF), insulin-like growth factor-1 (IGF-1), and leukemia inhibitory factor (LIF)[17−19,40]. All factors were used at their recommended concentrations (Supplementary Table S1, Table 1). SCs could survive in Formula 1 with minimal vitality (Fig. 1e, f). We then developed Formula 2 by adding lipids, non-essential amino acid (NEAA), and antioxidants[2,9]. After optimizing, the optimal concentrations of components in Formula 2 were: lipids concentration 1×, NEAA 1×, L-Ascorbic acid 100 μM, L-Ascorbic acid 2-phosphate (Asc-2P) 100 μM, Trolox 50 μM (Fig. 1b, Table 1). Subsequently, we examined the effects of hydrocortisone, forskolin, hepatocyte growth factor (HGF), dexamethasone, and lysophosphatidic acid (LPA) on SCs proliferation, which have been reported to regulate myogenic cell functions[24,41,42]. Our single-factor experiment showed that 100 nM hydrocortisone, 10 μM forskolin, 50 ng/mL HGF, 5 nM dexamethasone, and 10 μM LPA improved the short-term proliferation of SCs (Fig. 1c). The modified medium was termed as Formula 3 (Table 1). SCs in Formula 3 displayed higher proliferative capacity compared with cells in Formula 1 and Formula 2 (Fig. 1e, f). Finally, the optimal concentrations of ITS-X, bFGF, BSA, Y-27632, IGF-1, EGF, and LIF in Formula 3 were tested. We found that Y-27632, EGF, and LIF at their initial concentrations showed the best effects (Fig. 1d). Nevertheless, ITS-X (2×), bFGF (60 ng/mL), BSA (0.2%), and IGF-1 (100 ng/mL) required increased concentrations to promote cell growth in serum-free conditions (Fig. 1d, Table 1, Formula 4). Formula 4 showed the strongest effect in promoting SC proliferation (Fig 1e, f). Medium Formula 4 was then termed as A19 as it contained 19 constituents. The formulation of the A19 medium has been granted proprietary rights[43], and was used in subsequent experiments.
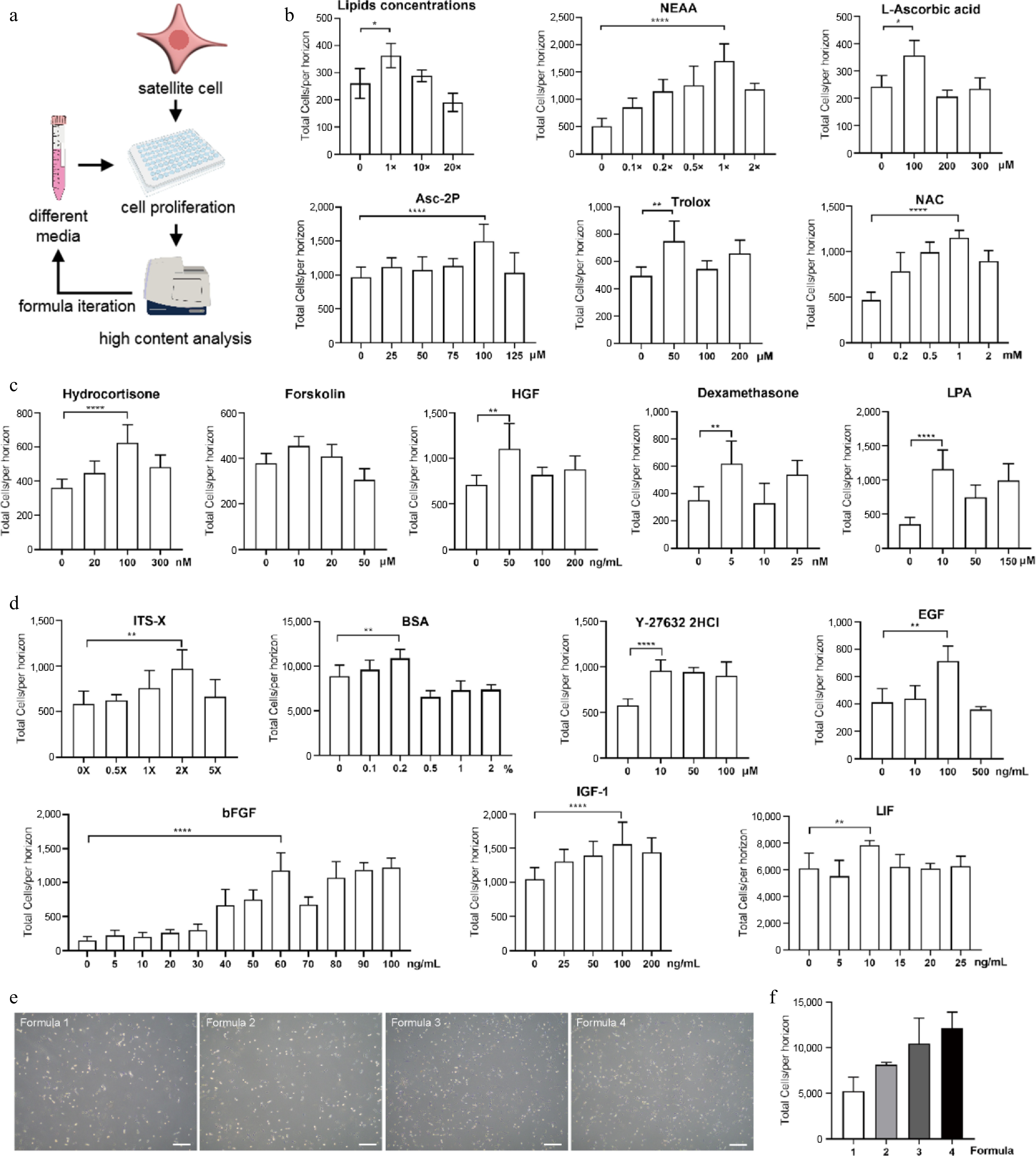
Figure 1.
Construction of serum-free proliferation medium for porcine SCs. (a) Schematic overview of medium formula iteration. (b) Effects of different supplements on cell growth with varied concentrations in Formula 2. (c) Effects of different factors on cell growth with varied concentrations in Formula 3. (d) Optimization of concentrations of factors used in Formula 1 to yield Formula 4. (e) Bright-field images of SCs cultivated by different media at day 3. Scale bars, 500 μm. (f) The effect on cell growth of different media. Mean ± SD from at least three independent experiments. Significance was analyzed by one-way ANOVA. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, ns, not significant (p > 0.05).
Table 1. The development of serum-free proliferation medium.
Formula 4 (A19) Formula 3 Formula 2 Formula 1 Basal media DMEM/F12 Basal media DMEM/F12 ITS-X 1 × Lipids concentration 1 × Hydrocortisone 100 nM ITS-X 2 × BFGF 10 ng/mL NEAA 1 × Forskolin 10 μM BFGF 60 ng/mL BSA 0.1% L-Ascorbic acid 100 μM HGF 50 ng/mL BSA 0.2% Y-27632 2HCl 10 μM Asc-2P 100 μM Dexamethasone 5 nM Y-27632 2HCl 10 μM EGF 100 ng/mL Trolox 50 μM LPA 10 μM EGF 100 ng/mL IGF-1 25 ng/mL NAC 1 mM IGF-1 100 ng/mL LIF 10 ng/mL LIF 10 ng/mL The primary porcine SCs were then cultivated using the A19 medium for three passages. Notably, SCs exhibited continuous proliferation with > 90% viable cell rate (Fig. 2a). Expression of key myogenic genes was also measured. PAX7, the master regulator of skeletal muscle stem cells[44], was significantly up-regulated in A19-cultivated SCs compared to cells maintained in serum-containing medium in all three passages tested (Fig. 2b). Consistent results were observed in MYOD and MYOG, the factor characterizing myogenic precursors and regulating myogenic commitment, respectively[45] (Fig. 2c, d). These data suggest that the A19 medium is superior to serum-containing medium in maintaining the stemness of porcine SCs. Nevertheless, as the passages increased, the decline of myogenic gene expression could not be fully prevented by the A19 medium (Fig. 2b−d).
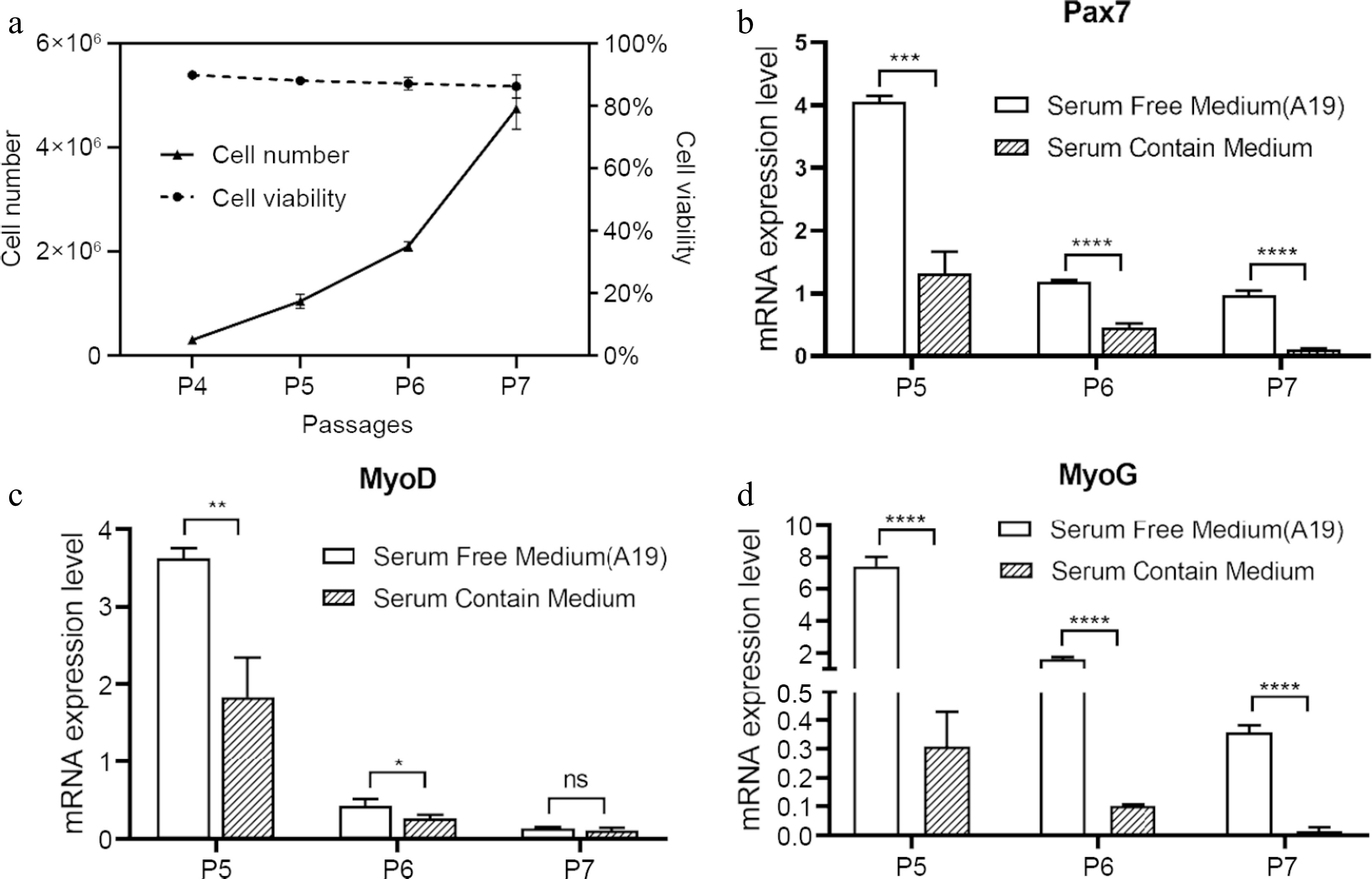
Figure 2.
Proliferation and gene expression of SCs in serum-free proliferation medium. (a) Cell number and viability of SCs cultured in serum-free medium (A19) during passage 4 (P4) to P7. (b)−(d) Real-time PCR quantification of PAX7, MYOD, and MYOG for SCs. Mean ± SD from at least three independent experiments. Significance was analyzed by two-tailed Student's t-test. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, ns, not significant (p > 0.05).
Construction and characterization of CDKN2A−/− SCs under serum conditions
-
As the expression of myogenic factors failed to maintain in serum-free A19-cultivated primary SCs (Fig. 2b−d), we next attempted to modify SCs through targeted genomic engineering approaches. We selected the CDKN2A gene locus as the target, which encodes p16INK4A and p14ARF, two factors known to drive cell cycle arrest and cellular senescence[33,34]. To knock out CDKN2A in porcine SCs, we designed single guide RNAs (sgCDKN2A) for Cas9 targeting exon 2 of CDKN2A gene, which provides genetic information for both p16INK4A and p14ARF (Fig. 3a). SCs were transduced with vectors expressing Cas/sgCDKN2A, and knockout clones were screened via single cell cloning and gene typing methods (Fig. 3a). Two CDKN2A−/− SC clones, KO-9 (−2 bp) and KO-15 (-2 bp) were successfully selected (Fig. 3b, c). We then cultured CDKN2A+/+ (WT) and CDKN2A−/− (KO) SCs for long-term periods. As shown in Fig. 3d, the proliferation rate of KO-9 and KO-15 SCs was significantly higher than that of WT cells during at least 18 passages. Moreover, expression level of PAX7 was significantly upregulated to ~194 folds in KO-9 SCs and ~52 folds in KO-15 SCs at passage 20 (P20) when compared to WT controls (Fig. 3e). Consistent results were observed at P30 (Fig. 3e). CDKN2A depletion also improved the expression of MYOD in two CDKN2A−/− clones at both P20 and P30 (Fig. 3e). MYOG mRNA level in KO-9 SCs was significantly up-regulated at P20 while downregulated at P30 compared with WT cells (Fig. 3e). Nonetheless, expression of MYOG was comparable in WT and KO-15 SCs at both passages (Fig. 3e). These results suggest that CDKN2A depletion significantly promote the proliferation capacity and maintenance of myogenic factors in porcine SCs.
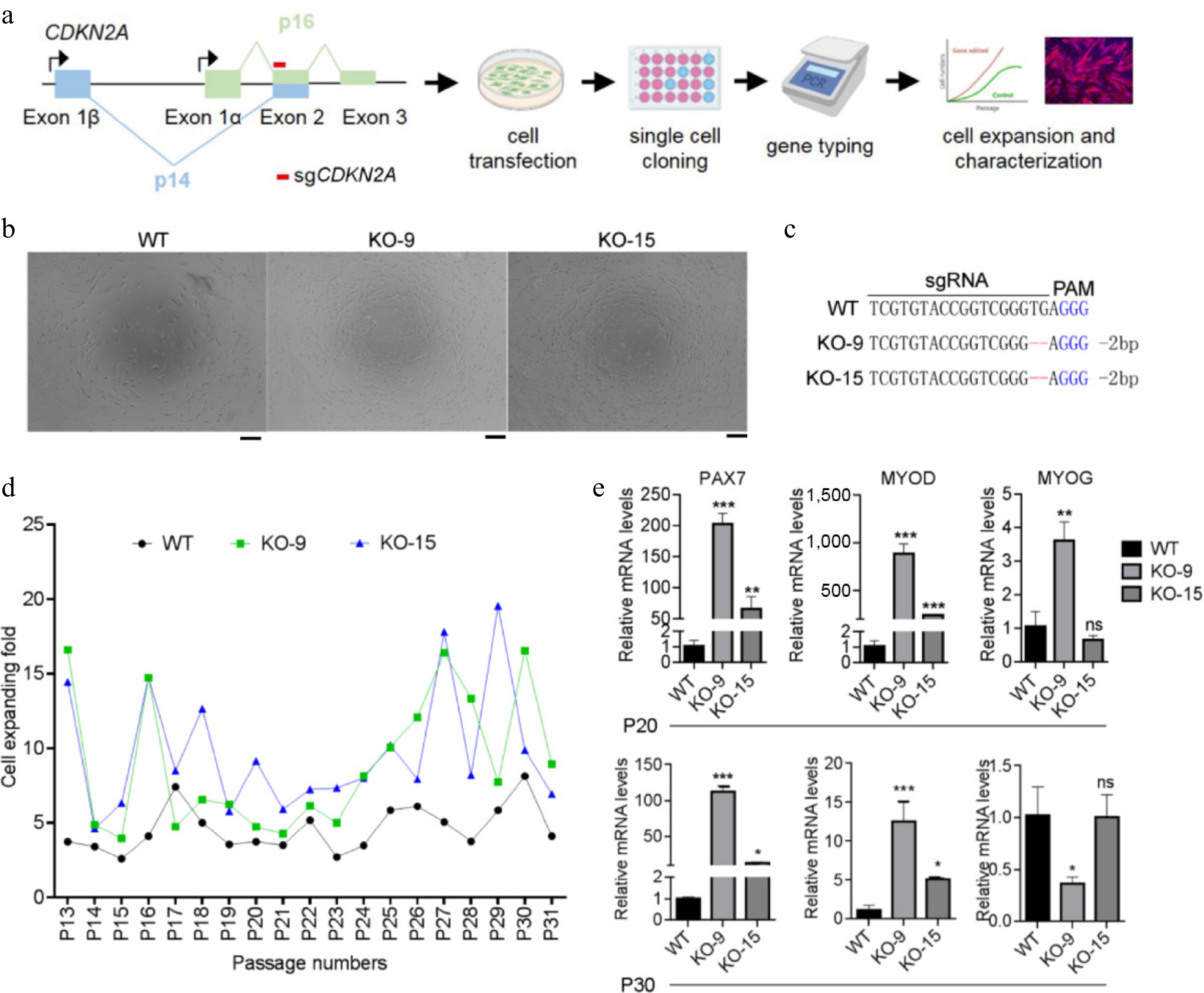
Figure 3.
Generating CDKN2A−/− SCs under serum condition. (a) Flowchart of experimental design for CDKN2A knockout, cell genotyping, and characterization of porcine SCs. (b) Microscopic images of CDKN2A+/+ (WT) and CDKN2A−/− (KO-9 and KO-15) SCs under bright field. Scale bar, 100 μm. (c) Sequencing data of CDKN2A mutations induced by CRISPR/Cas9 in SC clones. Deletions are indicated by dashes, and PAM is shown in blue. (d) Cell expanding folds of SCs during long-term expansion. (e) Expression by qPCR of PAX7, MYOD, and MYOG of SCs at P20 and P30. Mean ± SD from three independent experiments. Significance was analyzed by one-way ANOVA. * p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant (p > 0.05), compared with WT groups.
Next, we assessed the myogenic differentiation of CDKN2A+/+ and CDKN2A−/− SCs using the serum starvation method. As characterized by the minimal expression of myosin heavy chain (MyHC) and no fused myotubes, CDKN2A+/+ SCs at P10 almost lost their myogenic properties (Fig 4a, b). This is consistent with previous reports in porcine[2] and bovine SCs[18]. In contrast, CDKN2A−/− SCs around P10 successfully differentiated into mature myotubes with fusion index (16.3 ± 1.5)% in KO-9 cells and (56.3 ± 1.9)% in KO-15 cells (Fig 4a, b). Nevertheless, CDKN2A−/− SCs after long-term expansion (around P30) failed to differentiate into MyHC-positive myotubes, which was comparable to control SCs (Supplementary Fig. S1). These data suggest that, under serum conditions, CDKN2A deficiency maintains the myogenic potential of SCs at relatively early passages.
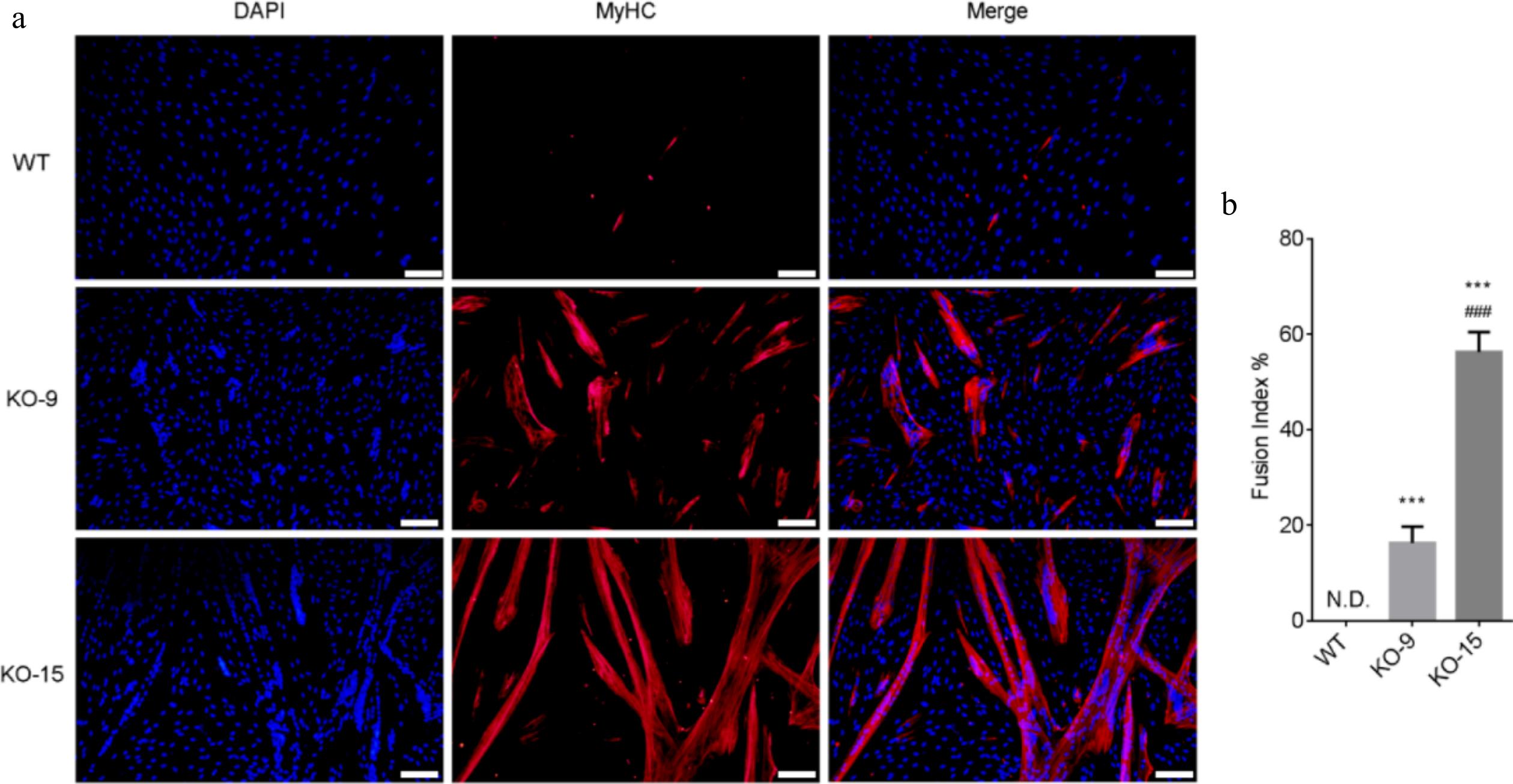
Figure 4.
Differentiation capacity of CDKN2A−/− SCs under serum condition. (a) Immunofluorescence staining of MyHC (red) and nucleus (blue) in differentiated WT and KO cells. Scale bar, 100 μm. (b) The fusion index was calculated as the percentage of myotubes with more than three nuclei. Mean ± SD from three independent experiments. Significance was analyzed by one-way ANOVA. *** p < 0.001, compared with WT group. ### p <0.001, compared with KO-9 group.
Long-term and stable expansion of CDKN2A−/− SCs in serum-free conditions
-
Subsequently, CDKN2A+/+ and CDKN2A−/− SCs were cultivated in serum-free medium to test their proliferation and differentiation capacities. After adapting in serum-free A19 medium for three passages (from P15), WT SCs displayed typical senescent features: flattening and enlarging morphology, the presence of granular substances, and minimal proliferation (Fig. 5a, b). In contrast, KO SCs exhibited robust viability in A19 medium and were more spherical in morphology in comparison with spindle-like SCs in serum conditions (Fig. 5a, b). The spherical morphology always characterizes myogenic stem cells[46,47], suggesting well-maintained stemness of CDKN2A−/− SCs. The WT and KO SCs were then cultured in serum and serum-free medium for long-term periods. WT SCs in A19 medium gradually lost the proliferation ability after three to four passages (Fig. 5b). Strikingly, KO SCs proliferated stably and efficiently for at least 15 passages in A19 medium and were superior to serum controls (Fig. 5b). We then examined the myogenic genes expression in SCs. When cultured in A19 medium for short passages (P15 + 2), expressions of PAX7 and MYOD were significantly upregulated in KO-serum and KO-serum-free SCs compared to WT-serum and WT-serum-free SCs, respectively (Fig. 5c). Serum-free cultivation also elevated the expression of MYOG in both WT and KO SCs (Fig. 5c). At P15 + 6, as WT-serum-free group failed to yield sufficient cells, we compared the remaining three groups. Expressions of PAX7, MYOD, and MOYG were all significantly upregulated in KO-serum and KO-serum-free SCs compared to WT-serum SCs, except MYOG in KO-serum SCs (Fig. 5d). Collectively, these results suggest that CDKN2A−/− SCs are compatible with our developed serum-free A19 medium and exhibit high proliferative activity and stemness maintenance during long-term culture.
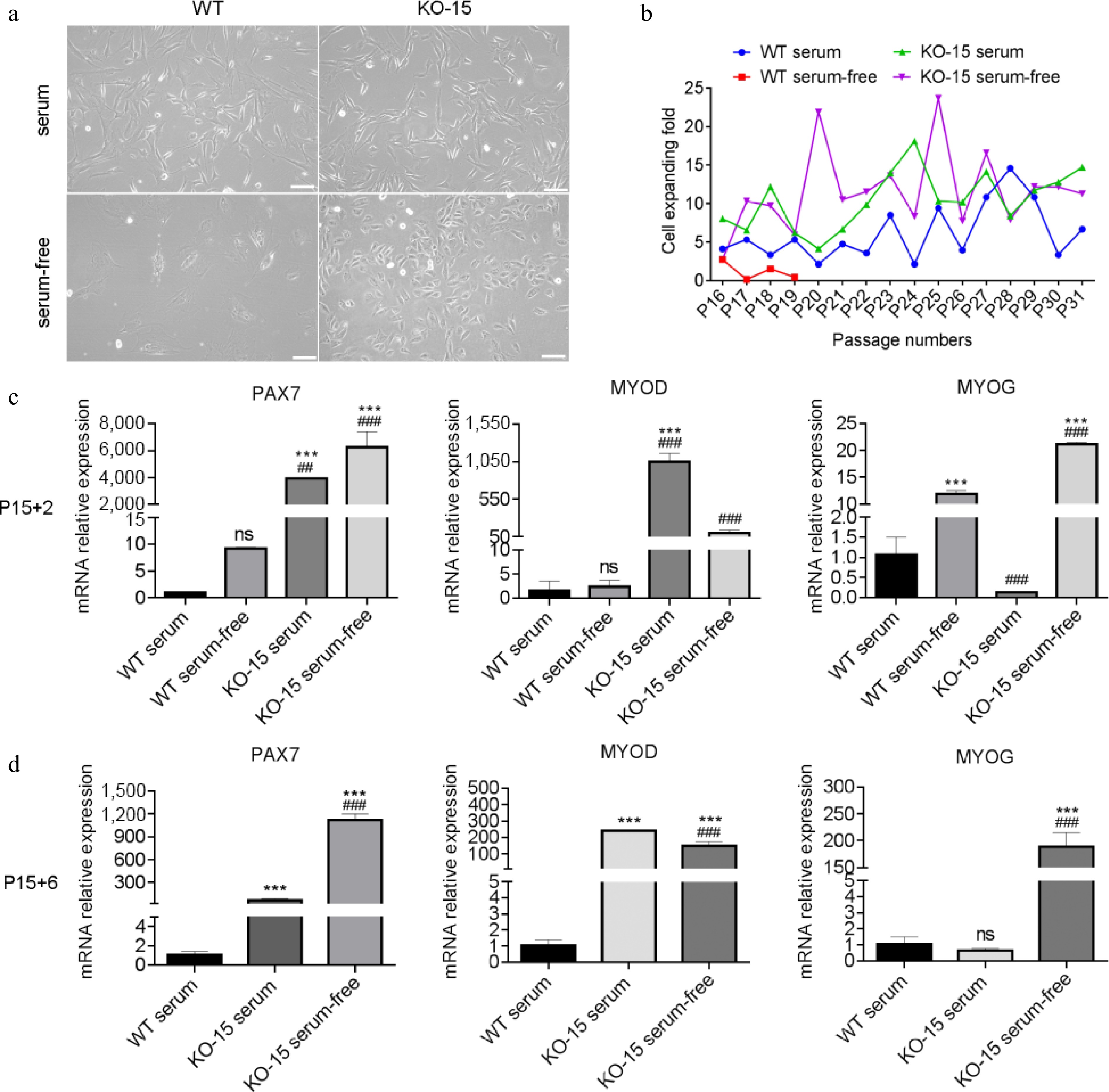
Figure 5.
Characterization of CDKN2A−/− SCs in serum-free condition. (a) Microscopic images of WT and KO-15 SCs under bright field. Scale bar, 100 μm. (b) Cell expanding folds of SCs under serum and serum-free conditions. (c), (d) Expression by qPCR of PAX7, MYOD, and MYOG of SCs at P15 + 2 and P15 + 6. '+X' indicates the cultivated passages in serum-free condition. Mean ± SD from three independent experiments. Significance was analyzed by (c) two-way ANOVA, and (d) one-way ANOVA . *** p < 0.001, compared with WT group. ## p <0.01, ### p <0.001, compared with KO-9 group, ns, not significant (p > 0.05).
Efficient myogenic differentiation of CDKN2A−/− SCs in serum-free conditions
-
Next, we examined the myogenic differentiation of CDKN2A−/− SCs after long-term expansion in serum-free proliferation medium. At first, we committed differentiation assay via serum starvation method and found that KO SCs at late passages (around P15 + 15) failed to form MyHC+ myotubes (Supplementary Fig. S2). We then tested the effect of several serum-free differentiation media. Our team had established a serum-free myogenic differentiation medium (Version 1.0), which consisted of DMEM, 0.1 × ITS-X (final concentration of insulin, 1 μg/mL), 25 ng/mL IGF-1, 25 ng/mL IGF-2, 0.01% BSA, and 0.1 × lipids concentration[48]. This medium improved the myogenic differentiation of porcine primary SCs at early passages compared to serum-containing differentiation medium (Supplementary Fig. S3). However, when differentiating in Version 1.0 medium, only minor MyHC+ and multi-nucleated myotubes emerged (Fig. 6a, b). This indicates that Version 1.0 medium is not optimal for CDKN2A−/− SCs differentiation. Previously, we constructed a serum-free adipogenic differentiation medium (Version 2.0), which contained DMEM/F12 (1:1), 1× ITS-X (insulin, 10 μg/mL), 1 ng/mL BFGF, 0.4% BSA, 50 ng/mL IGF-1, 50 μM Asc-2P, 10 μM Y-27632, 50 ng/mL EGF, 1 μM dexamethasone, 2 μM SB-431542, 100 μM 3-isobutyl-1-methylxanthine, 2 μM rosiglitazone and additional 10 μg/mL insulin[49]. This medium supported the efficient adipo genic differentiation of mesenchymal stem cells (MSCs) (Supplementary Fig. S4). Unexpectedly, during adipogenic differentiation assessment of CDKN2A−/− SCs, we observed that the Version 2.0 medium promoted the formation of aligned and elongated myotubes (Fig. 6c, d). In this medium, KO SCs could not form lipid droplets (Supplementary Fig. S5), further strengthening its myogenic properties. The above unexpected finding prompted us to further analyze and optimize the components in Version 2.0 medium. When comparing the media formula of Version 1.0 and Version 2.0, we found that the concentration of insulin was one of the differences (1.0 μg/mL in Version 1.0 and 20 μg/mL in Version 2.0). As evidenced by previous reports, a higher concentration of insulin was essential for myogenic differentiation[29]. Accordingly, we raised the insulin level to a final concentration of 11 μg/mL in Version 1.0 medium to yield Version 3.0 medium. In this medium, some MyHC+, elongated myotubes were formed (Fig. 6e, f). However, the efficiency was still not high enough. Subsequently, we focused on Version 2.0 medium, which might contain essential factors for high-efficient differentiation. As dexamethasone, 3-isobutyl-1-methylxanthine, and rosiglitazone were specific for inducing adipogenic differentiation[50], we removed them from Version 2.0 and generated Version 4.0 medium. Strikingly, in this medium, elongated and thicker myotubes with expression of MyHC were formed from CDKN2A−/− SCs cultivated in A19 medium for over 15 passages (Fig. 6g, h). The differentiation efficiency matched that observed in earlier-passage CDKN2A−/− SCs (Fig. 4a, b), suggesting the powerful role of serum-free Version 4.0 medium for myogenic differentiation.
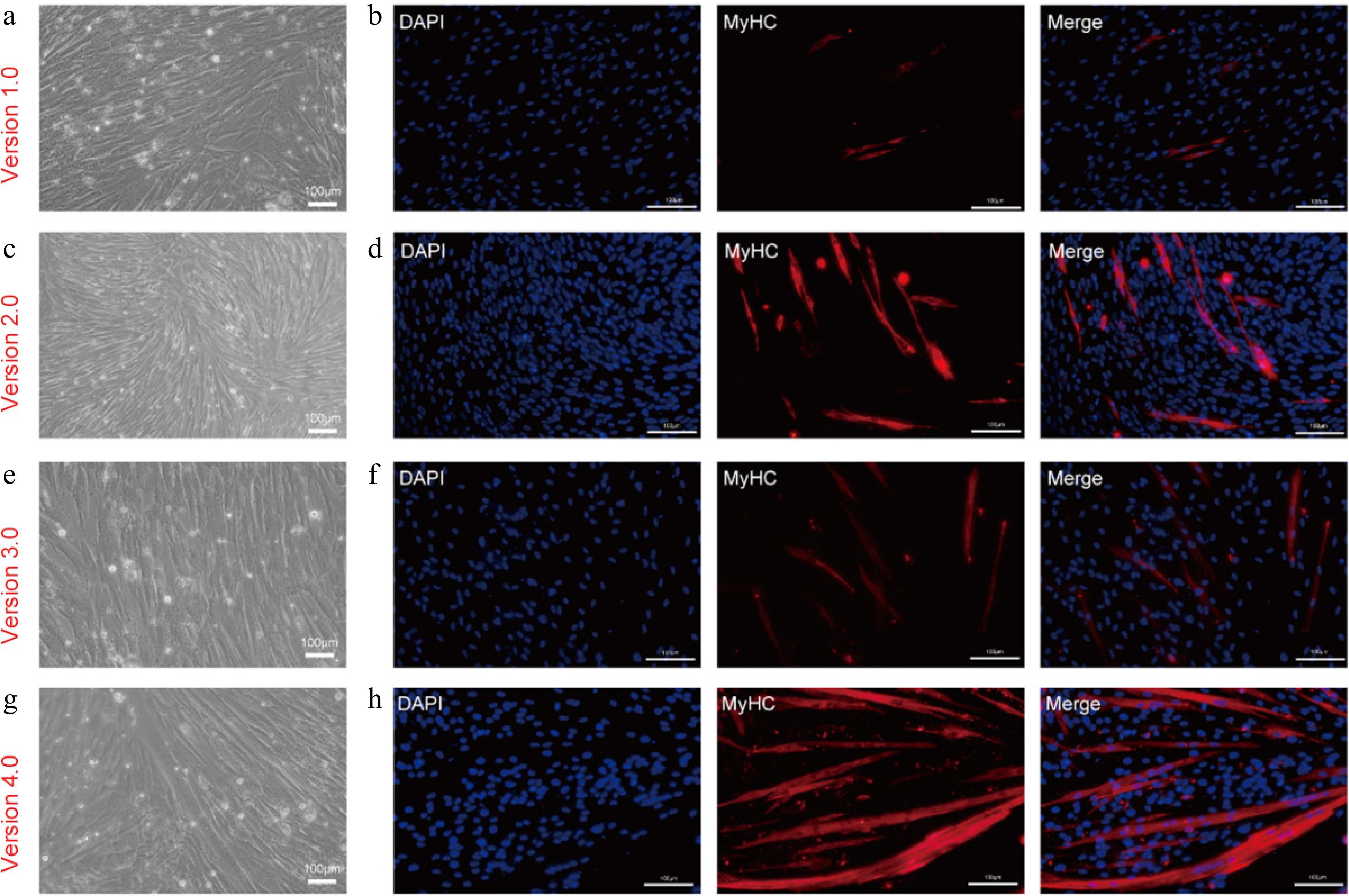
Figure 6.
Myogenic differentiation of CDKN2A−/− SCs in serum-free condition. (a), (c), (e), (g) Bright-field images, and (b), (d), (f), (h) immunofluorescent staining of KO-15 MuSCs induced by different serum-free differentiation media at day 4: (a), (b) serum-free myogenic differentiation medium (Version 1.0). (c), (d) Serum-free adipogenic differentiation medium (Version 2.0). (e), (f) Modified serum-free myogenic differentiation medium with 10 ng/mL insulin (Version 3.0). (g), (h) Modified serum-free adipogenic differentiation medium without dexamethasone, 3-isobutyl-1-methylxanthine and rosiglitazone (Version 4.0). Scale bars, 100 μm.
Serum-free media and CDKN2A−/− SCs enable the creation of meat-like construct
-
Production of cultured meat requires cell growth and maturation in 3D scaffolds. We therefore assessed the potential of serum-free proliferation medium (A19) and serum-free myogenic differentiation medium (Version 4.0) for supporting the formation of CDKN2A−/− SCs-derived cultured meat. The peanut wire-drawing protein scaffold was used here, which is a plant-based 3D edible biomaterial without animal-derived components and allows porcine cells to adhere and proliferate[51−53]. When inoculated into the 3D scaffold, CDKN2A−/− SCs could proliferate and contribute to the formation of viable meat-like constructs, as evidenced by the darker appearance (Supplementary Fig. S6a) and the emergence of MyHC+ signals after six days of culture (Fig. 7a). After four days' myogenic differentiation in Version 4.0 medium, expression of MYOD, MYOG, and MYHC in SCs were extensively upregulated compared to the proliferative condition (Fig. 7a−c), indicating the maturation of cells in 3D culture.
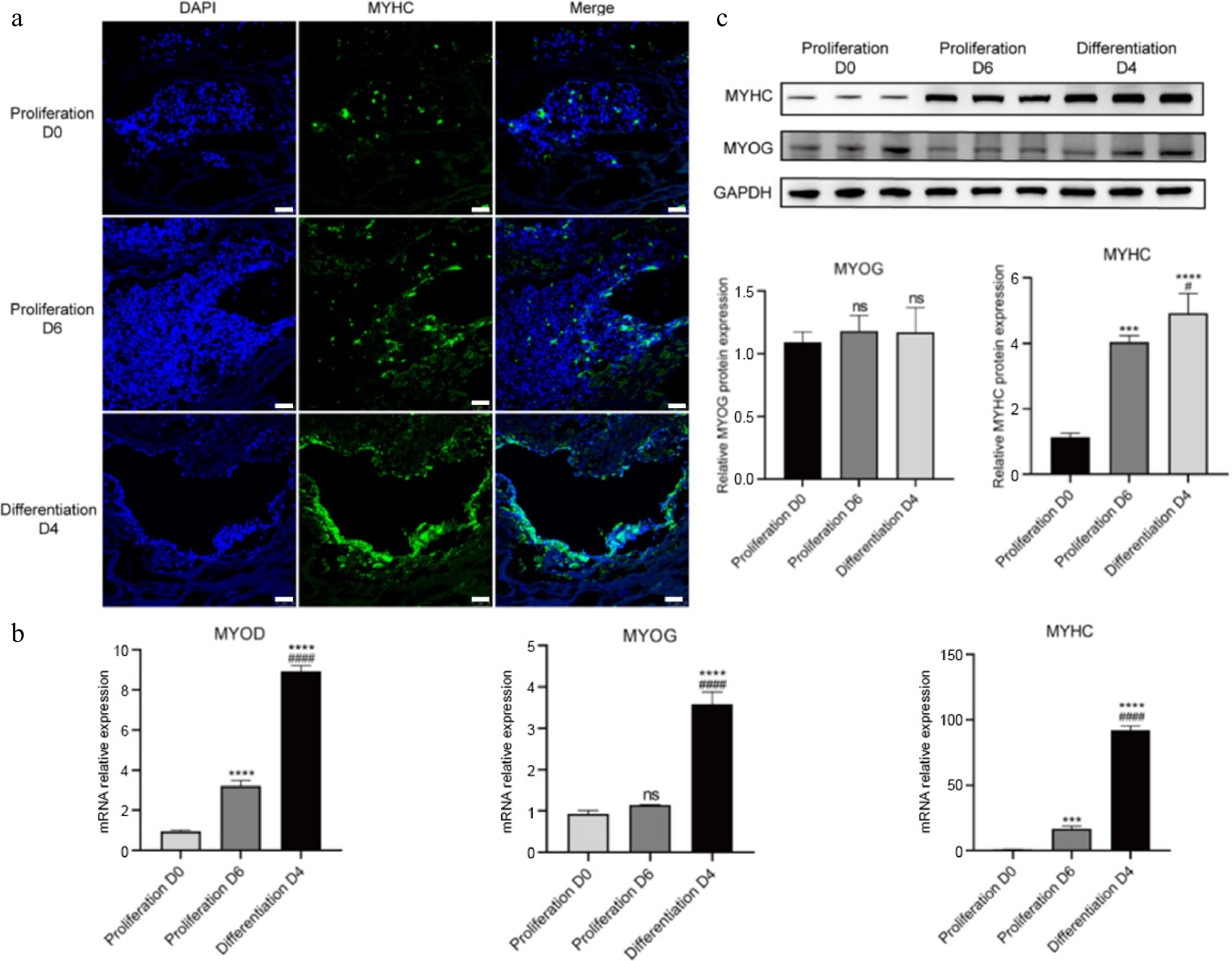
Figure 7.
3D culture of CDKN2A−/− SCs in peanut wire-drawing protein scaffolds. (a) Immunofluorescence staining of MYHC (green) and nucleus (blue) in proliferation and differentiation process. Scale bar, 25 μm. (b) Real-time qPCR quantification of MYOD, MYOG, and MYHC. (c) Protein expression of MYOG and MYHC examined by western blot and quantitative results analyzed by ImageJ. Mean ± SD from at least three independent experiments. Significance was analyzed by one-way ANOVA. *** p < 0.001, **** p < 0.0001, compared with proliferation D0 group; # p < 0.05, #### p < 0.0001, compared with proliferation D6 group, ns, not significant (p > 0.05).
We then determined the textural properties of SC-derived cultured meat constructs. The results indicated that there was no significant difference in terms of hardness among the blank scaffold, scaffold with proliferative cells and scaffold with differentiated cells, all of which were significantly lower than traditional pork (Supplementary Fig. S6b). For chewiness and springness, meat-like constrcut with mature cells was significantly upregulated in comparison with the pork group (Supplementary Fig. S6b). Notably, myogenic differentiation significantly enhanced gumminess parameters of cultured meat (Supplementary Fig. S6b). Overall, with the plant-based 3D scaffold, the serum-free system enables the preliminary generation of SCs-derived cultured meat.
-
Cost-effective production of cultured meat necessitates serum-free culture media. Herein, we established a defined serum-free medium (A19) that sustains porcine SC proliferation. Single-factor tests were used to optimize essential components for SC survival and proliferation. This strategy is simple and effective and is feasible to iterate. However, the formulation of cell culture media is a complex system, and the interrelationship between different factors may affect the outcomes of serum-free media[40]. Advanced experimental design methodologies including factorial, fractional factorial, Plackett-Burman, central composite, and response surface designs[18,19,54] can enhance optimization efficiency for serum-free medium development. For formulations requiring the evaluation of multiple variables (> 15), stochastic optimization approaches such as genetic algorithms offer valuable solutions[55]. The integration of machine learning and artificial intelligence has further revolutionized formulation development efficiency through predictive modeling[56]. Concurrently, systems biology approaches incorporating genome-scale metabolic modeling, isotope-assisted flux analysis, and spent media profiling have emerged as powerful tools for optimizing cost-effectiveness and manufacturing scalability of serum-free media[12].
Cell lines achieving long-term proliferation and maintaining cellular characteristics are key resources for cultured meat production. Several approaches have been established to induce the infinite proliferation of cultured cells, namely cell immortalization[57]. The immortalized cell lines can reduce the reliance on biopsies while being able to proliferate efficiently on a large-scale[58]. Currently, there are two main approaches for establishing immortalized cell lines, namely spontaneous immortalization and induced immortalization. The probability of spontaneous immortalization is extremely low, typically requiring long-term passaging to obtain suitable cells[59]. Induced immortalization methods primarily include viral infection or viral gene delivery[60,61], modulation of oncogenes or tumor suppressor genes[62], and activation of telomerase reverse transcriptase (TERT)[24,39]. The aforementioned methods often involve viral manipulation and gene integration, raising food safety concerns. Herein, using CRISPR/Cas9-mediated genetic engineering, we ablated the CDKN2A gene in porcine SCs. The CDKN2A−/− clones proliferated stably for over 18 passages with a maintained expression of myogenic factors and efficient differentiation (Figs 3 & 4), which could be recognized as immortalized cell lines.
Herein, we intriguingly found that CDKN2A depletion significantly promoted the stemness of SCs in serum-free conditions. CDKN2A gene functions as a cell cycle brake through S-phase entry inhibition[34]. Thus, CDKN2A depletion allows porcine SCs to bypass the Hayflick limit and acquire the ability for unlimited proliferation (Fig. 3d & b). Apart from this, CDKN2A might directly or indirectly regulate the expression of myogenic factors, such as MYOD, MRF5, and MYOG[63]. It is intriguing to elucidate the molecular mechanisms underlying CDKN2A-mediated regulation of myogenic stem cells, especially poultry and livestock stem cells for cultured meat production.
While CDKN2A inactivation enhances proliferation, its food safety implications warrant rigorous evaluation. As demonstrated previously, by indirectly repressing the activity of RB1 and p53, other properties of CDKN2A−/− cells including DNA damage response and genomic integrity would be preserved[39]. Further, comprehensive safety assessments are required to characterize the genomic integrity of engineered cell lines, including chromosomal karyotyping to monitor chromosomal stability, soft agar colony formation assays to evaluate anchorage-independent growth, and comet assays to quantify DNA strand breaks[58]. These approaches would provide critical safety validation for CDKN2A-edited cell lines in cultured meat production.
The texture analysis of 3D constructs indicates that the culture of SCs on the scaffold alters the originally soft and loose texture of the peanut scaffold, making it closer to the texture of pork. However, SCs-derived cultured meat has a light yellow color instead of a bright red one compared to pork. This is due to the absence of endogenous or exogenous myoglobin addition during the production process[64]. Moreover, exploring how to utilize co-cultivation or other methods to produce diversified products from cultured meat, including various nutrients such as muscle, fat, and collagen, is one of the tasks that need to be accomplished in future investigations. Overall, though the serum-free media enable the production of SC-derived cultured meat, the nutritional value, appearance, and other characteristics of the product require further optimization.
-
Development of cultured meat requires low-cost, efficient serum-free media and immortalized, stable cell lines for industrial-scale food production. Herein, we developed a serum-free proliferation medium (A19), which maintained the cell proliferation of porcine SCs at early passages. Subsequently, we constructed CDKN2A−/− SC lines using CRISPR/Cas-mediated genetic editing and found that CDKN2A ablation significantly enhanced proliferation and differentiation of SCs in serum conditions. Moreover, the serum-free A19 medium allowed for the robust and long-term expansion of CDKN2A−/− SCs for over 15 passages, while maintaining the expression of key myogenic factors. Further, we optimized the formulation of serum-free differentiation medium, which induced efficient myogenic differentiation of CDKN2A−/− SCs cultivated in A19 medium for long-term periods. Moreover, the serum-free proliferation and differentiation media enabled the preliminary production of SC-derived cultured meat in 3D food-grade plant scaffolds. Taken together, our study establishes a comprehensive serum-free system supporting long-term expansion and functional maintenance of genetically engineered SCs.
This study was supported by the National Natural Science Foundation of China (32272468), Nanjing Major Science and Technology Special (Comprehensive Category) Project (202309014), the Fundamental Research Funds for the Central Universities (KYT2024003), and the Jiangsu Agricultural Science and Technology Innovation Fund (CX(24)1019).
-
The authors confirm contribution to the paper as follows: Guo R and Wu Z conducted major experiments and analyzed the data. Lu M, Li J, Li W, and Yue B conducted part of experiments. Guo R and Wu Z designed experiments, prepared and revised the manuscript. Ding S designed experiments and revised the manuscript. Zhou G and Li C conceived the project, discussed the experiments and revised the manuscript. All authors reviewed the results and approved the final version of the manuscript.
-
Data will be made available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Renpeng Guo, Zhongyuan Wu
- Supplementary Table S1 Functions and recommended concentrations of various components.
- Supplementary Fig. S1 Differentiation capacity of SCs at later passages under serum conditions.
- Supplementary Fig. S2 Differentiation capacity of CDKN2A−/− SCs (KO-15) expanded in serum-free condition and differentiated with low-serum medium.
- Supplementary Fig. S3 Serum-free myogenic differentiation medium supports the differentiation of primary SCs.
- Supplementary Fig. S4 Serum-free adipogenic differentiation medium supports the differentiation of MSCs.
- Supplementary Fig. S5 Differentiation capacity of CDKN2A−/− SCs (KO-15) expanded in serum-free condition and differentiated with serum-free adipogenic culture medium.
- Supplementary Fig. S6 Photography and texture analysis of SCs-derived cultured meat.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Nanjing Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Guo R, Wu Z, Lu M, Li J, Li W, et al. 2025. Serum-free long-term expansion and functional maintenance of pig satellite cells via targeted genetic engineering for cultured meat. Food Materials Research 5: e007 doi: 10.48130/fmr-0025-0006 

Serum-free long-term expansion and functional maintenance of pig satellite cells via targeted genetic engineering for cultured meat
- Received: 30 November 2024
- Revised: 20 March 2025
- Accepted: 09 May 2025
- Published online: 25 June 2025
Abstract: Cultured meat represents a promising alternative for future meat consumption. However, the progress of research and industrialization has been hindered by the absence of serum-free media capable of supporting the long-term expansion of specific cells, such as satellite cells (SCs). In this study, we developed a serum-free proliferation medium (A19), which supported cell expansion of porcine SCs at early passages. Subsequently, using CRISPR/Cas-mediated genetic editing, we established CDKN2A−/− SC lines that exhibited enhanced proliferation and differentiation capacity in serum conditions compared to CDKN2A+/+ SCs. Notably, the serum-free medium allowed for the vigorous expansion of CDKN2A−/− cells over 15 passages, while maintaining the expression of key myogenic factors. Further, we optimized the formula of serum-free differentiation medium, which effectively induced myogenic differentiation of serum-free cultivated CDKN2A−/− SCs. Moreover, the serum-free media supported the generation of CDKN2A−/− SCs-derived cultured meat constructs using three-dimensional plant-based edible scaffold. Collectively, our study establishes a comprehensive serum-free system for SCs-based cultured meat production through targeted genetic engineering.
-
Key words:
- Cultured meat /
- Serum-free medium /
- Genetic engineering /
- CDKN2A