








-
Railway engineering waste slag refers to the excavation of stone slopes, which generates many natural mixtures such as natural gravel, abandoned soil, and plant debris[1]. These waste products are characterized by complex ground constituent materials with large variations, uneven physical structure and particle size, poor soil fertility or imbalanced nutrients, and unstable pH[2]. As a result, only a small portion of engineering waste slag is utilized on sites, such as landscapes, convenience facilities, and filling of embankments, with most of the waste slag remaining in situ on both sides of the railways[3]. However, these practices can cause soil erosion and landslides, and also lead to the pollution of water resources and the atmosphere[4].
Engineering waste slag can also be used to fill mines[5] and basic water conservancy projects[6], to prepare aggregates[7], and can be applied in road engineering[8]. The waste slag soil can also be mixed with soil conditioner as guest soil for vegetation restoration. Phytoremediation has emerged as a promising eco-remediation technology[9]. Qin et al.[10] used the mixed ratio of screened engineering waste residue with red clay and corn straw biochar to plant alfalfa (Medicago sativa L.) and Amorpha fruticose L., and found that the ratio of slag-soil > 3 was the most favorable ratio for vegetation restoration.
However, research on soil improvement focuses on plant growth[11,12]. There are fewer studies on the soil nutrients and microorganisms that contribute to the improvement of engineering waste slag soil. Moreover, soil nutrients[13] and microbes[14] are key indicators of soil quality and health status, and microbiomes are essential for nutrient cycling, organic matter decomposition, and soil structure. In high-altitude construction areas, traditional methods such as topsoil detachment followed by backfilling or soil cover are employed to manage the waste produced. Given the high expense of thin topsoil and the delicate nature of high-altitude ecosystems, utilizing improved engineering slag for ecological restoration in these areas presents a cost-effective resource utilization strategy. Therefore, in this study, the waste slag soils generated during the railway construction in Xinduqiao Town, Kangding City, China, were mixed with subgrade soil, reclaimed soil, and guest soil after sieving. Relevant plant and soil microbial indices were assessed to determine the best formulations for improved soils in high-altitude areas. These findings will also provide a reference for the use of soil mixing and improvement technology for vegetation restoration in the ecological restoration area of railway projects in similar high-altitude areas.
-
The experimental site was located in Xinduqiao Town, Kangding City, Ganzi Tibetan Autonomous Prefecture, Sichuan Province, China with an average altitude of 3,460 m above sea level, and average annual, summer, and winter temperatures of 7.1, 15.7, and 2.5 °C, respectively. The province has a plateau-type continental monsoon climate, with an annual precipitation of 800−950 mm concentrated between May and September, and experiences mild temperatures. The sunshine duration is sufficient, averaging 1,738 h annually, while the frost-free period is between 150 and 250 d.
Experimental design
-
The waste slag soil was sieved using a 5 mm sieve and mixed with subgrade soils, guest, and reclamation soils in different volume ratios (Table 1) to form a 40 cm thick planting soil layer. The waste slag soil consisted of the soil obtained after crushing and sieving disposal residues such as stones, soil, and plant residues generated in railway projects, while the subgrade soil included the raw soil below 40 cm on the surface of the road surface, and guest and reclamation soils were the topsoil stripped from the railway construction area. A total of 15 soil mix formulations with different ratios were designed, with L1 and L15 as the controls (Table 1). Rotted cow dung, sheep dung, or commercial organic fertilizer was applied as the bottom fertilizer at 300 g·m−2 in all formulations[15]. The drooping wild ryegrass (Elymus nutans) was planted with a sowing rate of 9 g·m−2 in 3 m × 2 m plots in triplicate in May 2022; each treatment was repeated three times, with a total of 45 plots in a 270 m2 total planting area. The aisles were 0.5 m and filled with formulated soil, covered with non-woven fabric in the year of sowing, uncovered after emergence, and not watered to simulate the natural environment. The physical and chemical properties of different soils are shown in Table 2. The waste slag soil had the lowest bulk density and water content, while the guest soil and subgrade soil had higher water content and organic matter content.
Table 1. Soil formulations and mixture proportions used in the 15 treatments (L1–L15) evaluated in this study.
Serial number Soil ratios Waste slag
soil (%)Subgrade
soil (%)Guest soil +
Reclamation soil (%)L1 0 0 100 L2 25 75 L3 50 50 L4 75 25 L5 100 0 L6 25 0 75 L7 25 50 L8 50 25 L9 75 0 L10 50 0 50 L11 25 25 L12 50 0 L13 75 0 25 L14 25 0 L15 100 0 0 Table 2. Partial physical and chemical properties of the four soils.
Soil type Soil bulk weight (g·cm−3) Soil moisture content (%) Soil organic matter content (g·kg−1) Waste slag soil 1.54 ± 0.02 5.61 ± 0.42 15.18 ± 0.23 Subgrade soil 1.30 ± 0.03 16.35 ± 1.06 18.69 ± 0.12 Reclamation 1.41 ± 0.04 9.83 ± 1.27 13.44 ± 0.61 Guest soil 1.31 ± 0.03 17.82 ± 0.45 11.36 ± 0.73 Soil sampling
-
The soil samples were collected in triplicate from each plot with a spade from 0−10 cm on the 13th of August 2023, using a 5-point sampling method, and then mixed to form a sample[16]. The collected soil samples were used to determine the composition of the soil nutrients, microorganisms, and other related indicators. The soil samples were air-dried and sieved using soil sieves of different particle sizes (1 and 0.25 mm) and used to determine the total nitrogen (TN), available phosphorus (AP), potassium (AK), and organic matter (SOM). Soil for microbiological determinations was packed in sterile bags and transported quickly back to the laboratory in ice boxes for total DNA extraction, PCR amplification, and sequence determination. Total microbial DNA extraction, PCR amplification, and sequencing were performed by Shanghai Majorbio Bio-pharm Technology Co., Ltd. (Shanghai, China).
Vegetation cover and aboveground biomass determination
-
The vegetation cover was measured by visual inspection and determined as the ratio of the vertical projected area within the sample plot to the area of the sample plot. The aboveground biomass was measured by a sampling method using 50 cm × 50 cm quadrats. Plants within a quadrat in each plot were cut off from the ground and collected into envelopes. Plants were then heated at 110 °C for 20 min and then dried to constant weight at 65 °C and used to calculate the aboveground biomass per unit area.
Soil physicochemical properties
-
Soil physicochemical properties were determined according to Soil Agrochemical Analysis[17]. The particle size composition was determined from samples taken before planting. Soil samples were mixed evenly and weighed according to < 0.25 mm, 0.25−1 mm, and 1−2 mm particle size sieves, and the proportion of particles of different sizes was calculated. Soil bulk density was measured before sowing by ring knife method, the total N was determined by the semi-micro Kjeldahl method, available P was determined by 0.03 mol·L−1 NH4F−, 0.025 mol·L−1 HCl leaching-molybdenum antimony colorimetric method, available potassium was determined by 1 mol·L−1 neutral NH4OAC leaching-flame photometric method, while the organic matter was determined using the potassium dichromate volumetric method-external heating method.
High-throughput sequencing analysis of the soil microbial community
-
The genomic DNA of the microbial community from the soil was extracted using the E.Z.N.A.® soil DNA Kit (Omega Bio-Tek, Norcross, GA, USA) following the manufacturer's instructions. The quality and concentration of DNA were determined by 1.0% agarose gel electrophoresis and a NanoDrop2000 spectrophotometer (Thermo Scientific, USA). The hypervariable region V3−V4 of the bacterial 16S rDNA gene were amplified with primer pairs 338F (5'-ACTCCTACGGGAGGCAGCAG-3') and 806R (5'-GGACTACHVGGGTWTCTAAT-3')[18], while the fungal ITS region was amplified using primers (ITS1F: CTTGGTCATTTAGAGGAAGTAA, ITS2R: GCTGCGTTCTTCATCGATGC)[19] in triplicates, and the PCR products of the same sample were mixed. The PCR product was extracted from 2% agarose gel and purified using the PCR Clean-Up Kit (YuHua, Shanghai, China) according to the manufacturer's instructions and quantified using Qubit 4.0 (Thermo Fisher Scientific, USA). Purified amplicons were pooled in equimolar amounts and paired-end sequenced on an Illumina Nextseq2000 platform (Illumina, San Diego, USA) according to the standard protocols by Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China).
Statistical analysis
-
Data analysis of the microbial properties was based on the Majorbio cloud platform (
www.majorbio.com ) provided by Majorbio Bio-Pharm Technology Co. Ltd. Raw FASTQ files were de-multiplexed using an in-house Perl script, and then quality-filtered by fastp version 0.19.6[20] and merged by FLASH version 1.2.7[21]. Then the optimized sequences were clustered into operational taxonomic units (OTUs) using UPARSE 7.1[22−23] which had a 97% sequence similarity level. Silva database (Release 138)[24], and the Unite database (Unite, Release 8.0)[25] were used for bacterial and fungal community composition and taxonomy classification based on the OTUs. Chao1 and Shannon indices were used to quantify the α-diversity of the microbial community, and principal coordinate analysis (PCoA) was used to illustrate the β-diversity of the microbial community. The PCoA was also used to assess the effects of treatments on the microbial community. The vegan package (version 2.4.3) in R[26] was used for redundancy analyses. One-way analysis of variance and Duncan's multiple-range test were used to calculate the significant differences among the treatments using SPSS v. 23.0 (IBM, Inc., Armonk, NY, USA)[27]. The significance threshold was set at 5% (p < 0.05). Figures illustrating plant and soil index data were generated using Origin Pro 2021 (OriginLab Corporation, Northampton, MA, USA)[28]. -
The L9 treatment resulted in the highest vegetation cover, followed by the L4 treatment, with the L13 treatment having the lowest cover (Fig. 1a). The vegetation cover for L13-treated plants was 62.50%, lower than the L9 treatment but not significantly different from the L14 treatment (p > 0.05). When the percentage of waste slag used was 0% (L1−L5), the L1 treatment had the lowest vegetation cover, and the L4 treatment had the highest, indicating that the plant establishment capacity in 100% guest and reclaimed soils was worse compared to that in the added subgrade soil. When the percentage of waste slag was 25% (L6−L9), the L9 treatment had the highest cover, 11.11% higher than that of the L6 treatment. However, when the percentage of waste slag was 50%, the cover of Elymus nutans increased when the percentage of subgrade soil was increased (L10 < L11 < L12). The L15 treatment is not shown on the vegetation cover and aboveground biomass histograms because of its poor establishment of Elymus nutans after the second year of planting. These results indicate that the germination of Elymus nutans may be adversely affected by the low volume and moisture content of purified waste slag.
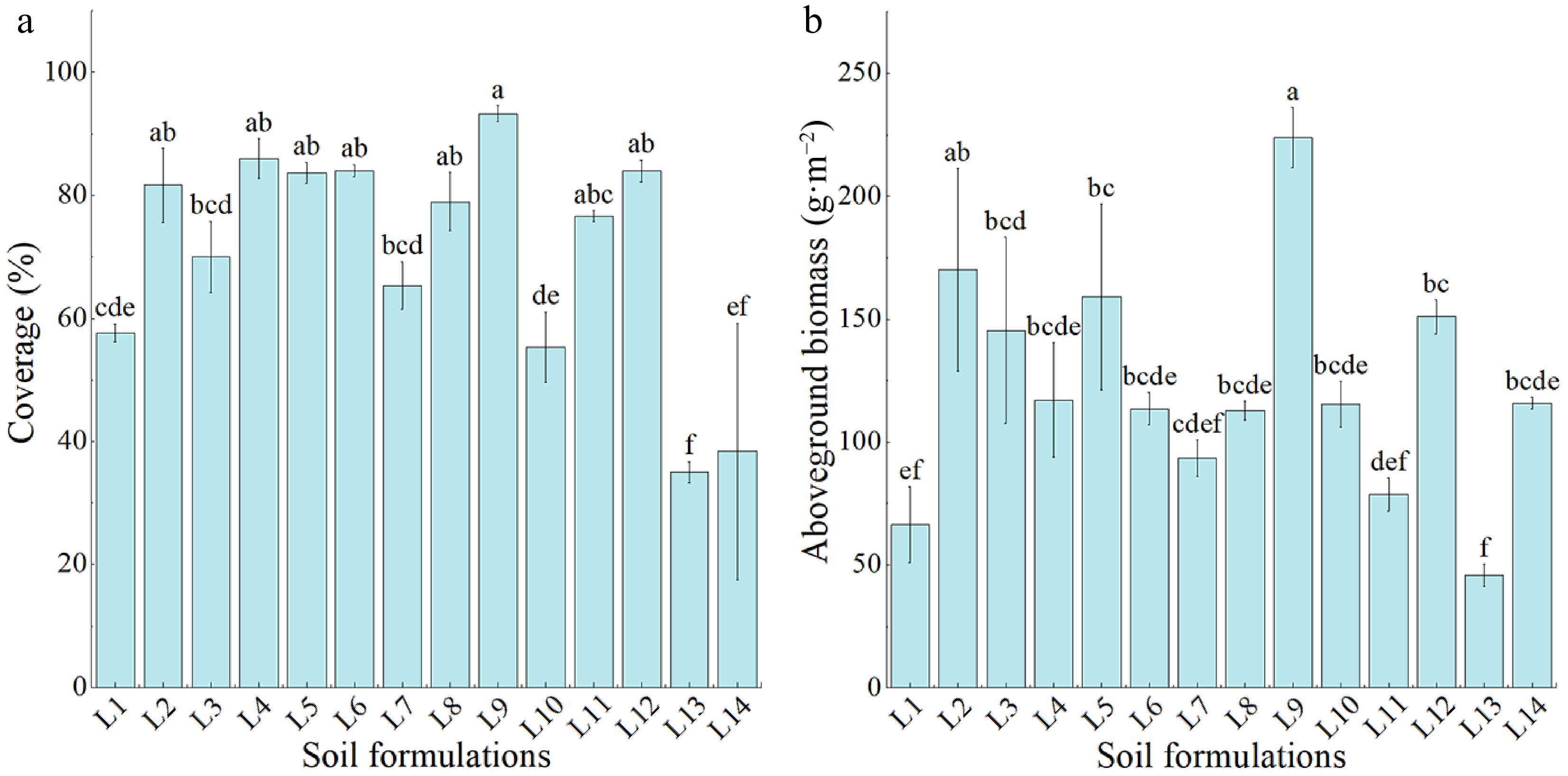
Figure 1.
The effects of different ratios of treated-waste slag soil on Elymus nutans vegetation coverage and aboveground biomass. (a) Vegetation coverage. (b) Aboveground biomass. Different lowercase letters indicate significant differences among different treatments (p < 0.05).
Effects on plant aboveground biomass
-
The analysis showed that Elymus nutans grown in L9 mixed soil had the highest aboveground biomass (Fig. 1b), being significantly different from those in all the other treated mixed soils (p < 0.05) except for the L2 treatment. The aboveground biomass of L13-treated Elymus nutans was the lowest and was comparable to L1 and L11 treatments (p > 0.05; Fig. 1b). The data showed that when the proportion of waste slag soil was 0%, the effect of 100% guest and reclaimed soils was worse than 100% subgrade soil. When the proportion of waste slag soil was 25%, the aboveground biomass of the L9-treated Elymus nutans was the highest, exceeding that of the L6 treatment without subgrade soil by 96.20%. However, when the proportion of waste slag soil was increased to 25%, 50%, or 75%, the treatments with the greatest proportions of subgrade soils exhibited the highest aboveground biomass.
The effect of different ratios of waste slag soil on soil physical and chemical properties
Effects on soil particle size composition and bulk density
-
According to the Chinese particle size classification standard (1987)[29], the particles of 1−3 mm are fine gravel, the particles of 0.25−1 mm are coarse sand, and the particles of < 0.25 mm are fine sand, silt, and clay. As shown in Fig. 2a, among the 15 soil formulations evaluated, the L1 treatment had the highest 1−2 mm particle size composition, and the L6 treatment had the lowest. The L6 treatment contained the highest proportion of 0.25−1 mm particles, and the L9 treatment had the lowest. The L12 treatment contained the most particles with a size of less than 0.25 mm, while the L15 treatment contained the least. The content of fine sand, silt, and clay in the soil increased as the proportion of waste slag soil was reduced from 100% to 0%. This increase was characterized by a trend of initial increase and subsequent decrease, suggesting that the mechanical composition of waste slag soil could be adjusted by the addition of different types of soil. This adjustment could potentially impact the water content, air permeability, and water permeability of the soil to a certain extent. The soil bulk density of the L6 and L9 treatments was lower than that of the L1 treatment (Fig. 2b) due to the former having the lowest proportion of 1−2 mm particle size and the latter having the lowest proportion of 0.25−1 mm particle size.
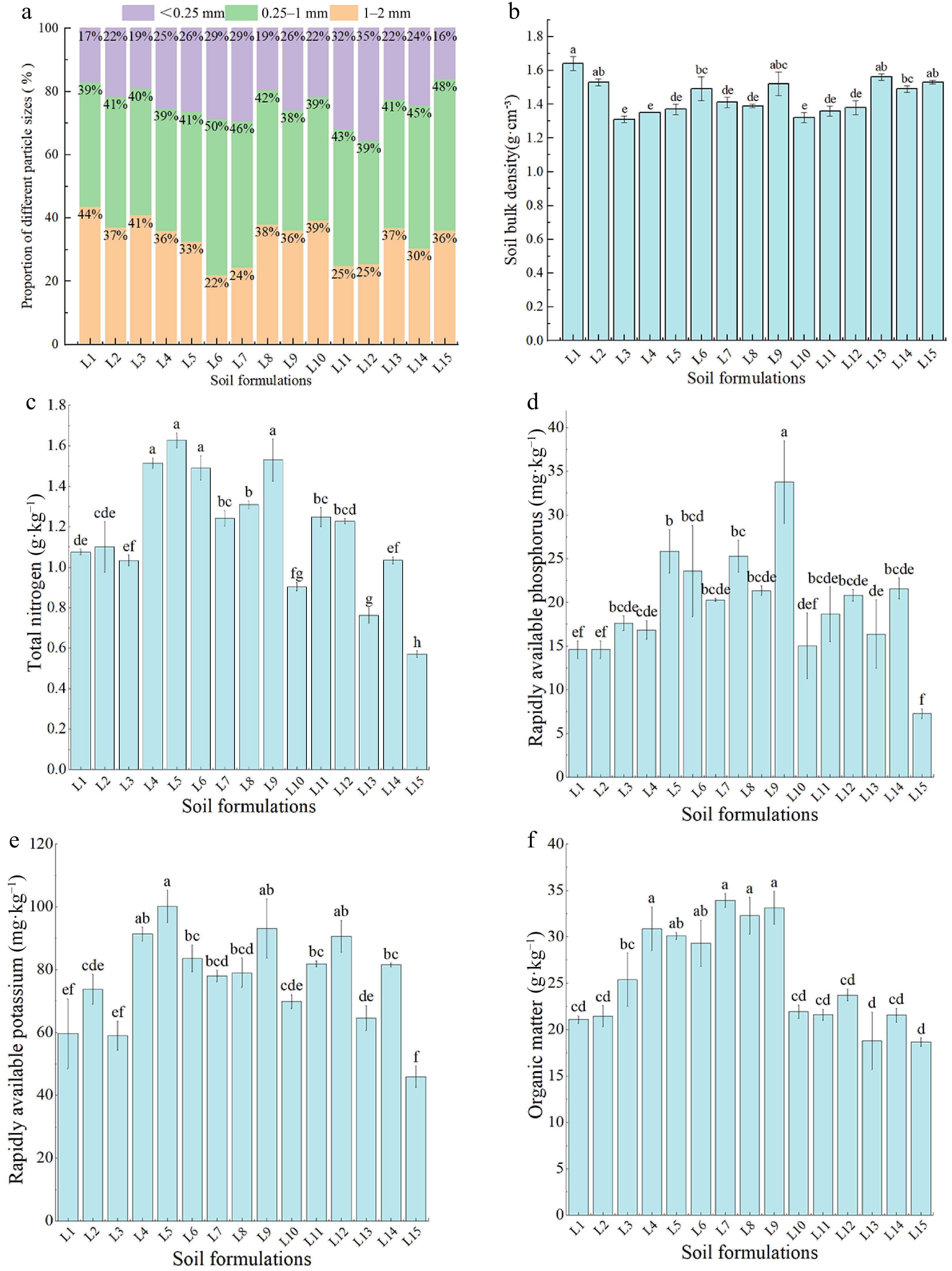
Figure 2.
Effects of different ratios of treated-waste slag soil on soil physical and chemical indexes. (a) Particle size composition. (b) Soil bulk density. (c) Total nitrogen. (d) Available phosphorus. (e) Available potassium. (f) Organic matter.
Effects on total N
-
The L5 mixed soil had the highest total soil N content, followed by L9, L4, and L1 soils, with the least N in L15 mixed soils (Fig. 2c). The L5 mixed soil with the highest percentage of subgrade soil had the highest total N content when the percentage of waste slag was 0%, while the L9 mixed soil had the highest total N content when the percentage of waste slag was 25%. The total N content was highest in L11 mixed soil when it had 50% waste slag. It increased as the percentage of subgrade soil increased, particularly when the percentage of waste slag was 75%.
Effects on soil available P
-
The available P content was highest in L9 waste-mixed soil, followed by L4, L7, and L5 mixed soils, and the lowest in L15 mixed soil (Fig. 2d). The amount of available P in the L9 treatment was higher than that in the L4 and L15 treatments by 30.80% and 365.50%, respectively. The L4-treated mixed waste soils had the highest available P content when the percentage of waste slag was 0% (L1−L5), while the L9 treatment had the highest available P content when the percentage of waste slag was 25% (L6−L9). The available P content increased with the increase in subgrade soil percentage when the percentage of waste slag was 50% (L10−L12) and 75% (L13−L14).
Effects on soil available K
-
The soil available K content was highest in the L5 treatment, followed by the L9 treatment, with the lowest in L15-treated waste mixed soils (Fig. 2e). The contents of soil available K in the L5 and L9 treatments were 117.85% and 102.61% higher than those in the L15 treatment, respectively, while L1 and L3 treatments had a K content comparable to that in the L15 treatment (p > 0.05). The L5 treatment with the highest percentage of subgrade soil had the highest available K content when the percentage of waste slag was 0%. The L9 treatment had the highest available K content when the percentage of waste slag was 25%, but the available K content increased with the percentage of subgrade soil when the percentage of waste slag was increased to 50% or 75%.
Effects on soil organic matter
-
The highest soil organic matter content in waste slag soil formulations was found in the L7 treatment, followed by the L9, L8, and L4 treatments, and the lowest in the L15 treatment. The organic content in the L7 and L9 treatments was 81.88% and 77.63% higher than that in the L15 treatment (Fig. 2f). The L4-treated soils containing no waste slag soil had the highest organic matter content, 46.30% higher than that of the L1 treatment. However, as the proportion of waste slag soil was increased to 25%, the L7 treatment had the highest organic matter content, 15.73% higher than that of the L6 treatment. The findings of this study indicated that when the proportion of waste slag soil was increased to 50% or 75%, the corresponding treatments with the highest proportions of subgrade soils had the highest contents of soil organic matter.
Effects on the taxonomic composition of microbial communities
-
The bacterial soil microbial community analysis revealed 2,923 OTUs, which were assigned to 36 phyla, 95 classes, 223 orders, 274 families, 647 genera, and 1,311 species. Among these, 681 OTUs were shared across all 15 treatments (Fig. 3a). The bacterial OTUs varied greatly across the treatments, with the L5 treatment containing the highest number of specific bacteria, followed by the L4 treatment, whereas the L15 treatment exhibited the lowest count (Fig. 3a). The fungal soil microbial community analysis revealed 4,004 OTUs, which were assigned to 16 phyla, 64 classes, 142 orders, 312 families, 695 genera, and 1,191 species; among these, 81 OTUs were shared across all 15 treatments (Fig. 3b). Fungal OTUs varied greatly across the treatments, with the L5 treatment containing the highest number of specific fungi, followed by the L6 treatment, with the least observed in L14.
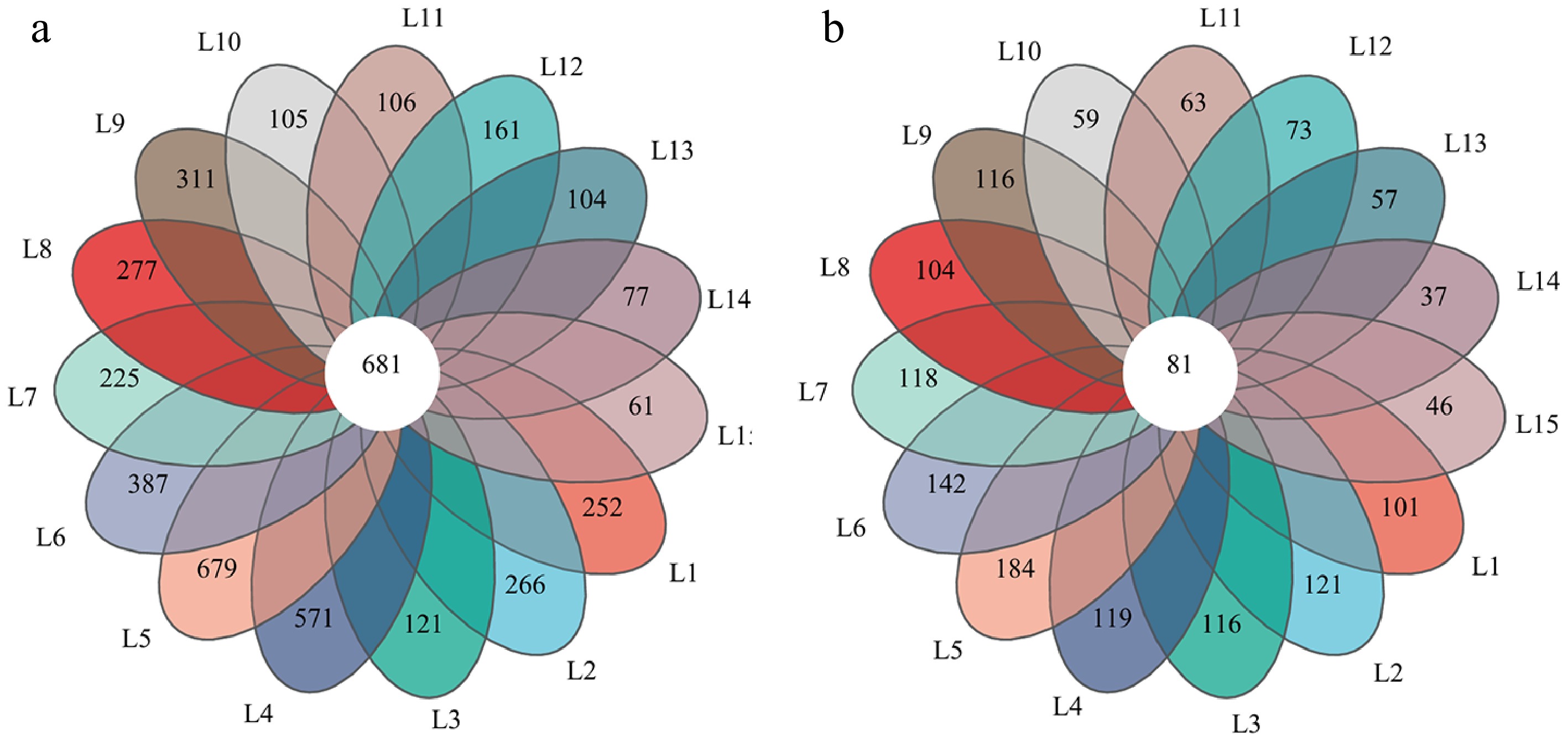
Figure 3.
Venn diagram showing the distribution of microbial OTUs in different waste-mixed soils. (a) Bacteria, (b) Fungi.
The predominant bacterial communities were Proteobacteria, Actinobacteriota, Acidobacteriota, Chloroflexi, and Firmicutes, accounting for 76.20%−83.96% of the total. Other phyla with an abundance greater than 1% were Bacteroidota, Gemmatimonadota, Myxococcota, Verrucomicrobiota, Patescibacteria, Bdellovibrionota, Cyanobacteria, and Planctomycetota (Fig. 4a). The proportion of Actinobacteriota in the treatment groups with waste slag soils was lower than that in the treatment groups without waste slag soil, while the 100% waste slag soil L15 treatment group had the lowest proportion. The proportion of Chloroflexi and Firmicutes in the 100% waste slag soil treatment group was the lowest. Five fungal phyla were present in all treatment groups in a greater than 1% abundance: Ascomycota, Basidiomycota, Mortierellomycota, Unclassified fungi, and Chytridiomycota (Fig. 4b). The number of fungi with abundance greater than 1% was less than that of bacteria, with a decrease of 160%.
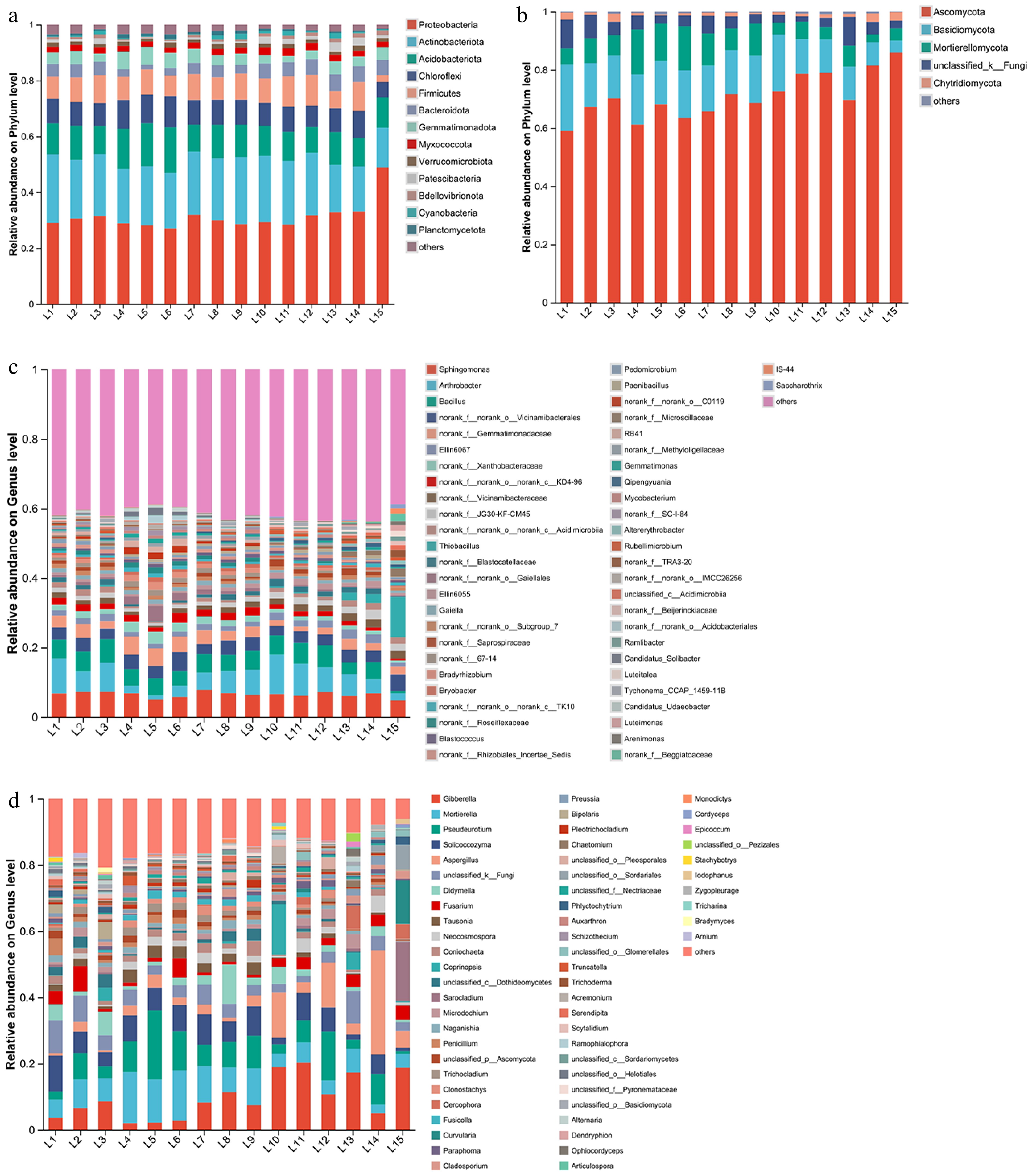
Figure 4.
The community composition and relative abundance of soil microorganisms at the phylum level and genus level under different treatments. The data represent the community composition and relative abundance of (a) bacteria, (b) fungi at the phylum level, and the community composition and relative abundance of (c) bacteria, and (d) fungi at the genus level.
Among the 15 treatments, 52 bacterial genera had an abundance of greater than 1%. Sphingomonas, Arthrobacter, Bacillus, Norank_f__norank_o__Vicinamibacterales, and Norank_f__Gemmatimonadaceae were the top 5 most abundant genera (Fig. 4c), with a relative abundance of 14.50% to 29.20%. The L7 treatment had the highest abundance of Sphingomonas, while the L10 treatment contained the highest number of Arthrobacter. The relative abundance of the top five dominant bacterial genera in the L15 treatment group (100% waste slag) was lower than all the other treatments, indicating that mixing waste slag soils with subgrade soil, guest soil, and reclaimed soils led to an increase in the abundance of dominant bacterial communities.
Among the 15 treatments, 60 fungal genera were identified with an abundance greater than 1%. The top five genera with the highest percentage of abundance were Gibberella, Mortierella, Pseudeurotium, Solicoccozyma, and Aspergillus (Fig. 4d). Gibberella was most abundant in L11, L10, and L15 (Fig. 4d), while Mortierella increased from L5 and L6 to L4. L5 treatment had a higher number of Pseudeurotium, while L14, L10, and L12 had the highest abundance of Aspergillus. Generally, these top five dominant fungal genera were the most abundant in the L14 treatment group, followed by L12 and L5, with the least abundance in the L1 treatment group. The number of fungi with abundance greater than 1% increased by 15.38% compared with bacteria.
Alpha diversity of the microbial community
-
The Chao1 indices of bacteria (Fig. 5a) and fungi (Fig. 5c) were higher in the treatment groups without added wastes and in the treatment groups with 25%, 50%, and 75% of waste slag added, respectively, than in the treatment group with only waste slag. The L4 and L6 treatments exhibited the highest Chao1 index, suggesting that the soil bacteria and fungi species were most abundant in these respective treatments. As evidenced by the Shannon index (Fig. 5b, d), the soil microbial diversity was improved after adding other types of soil.
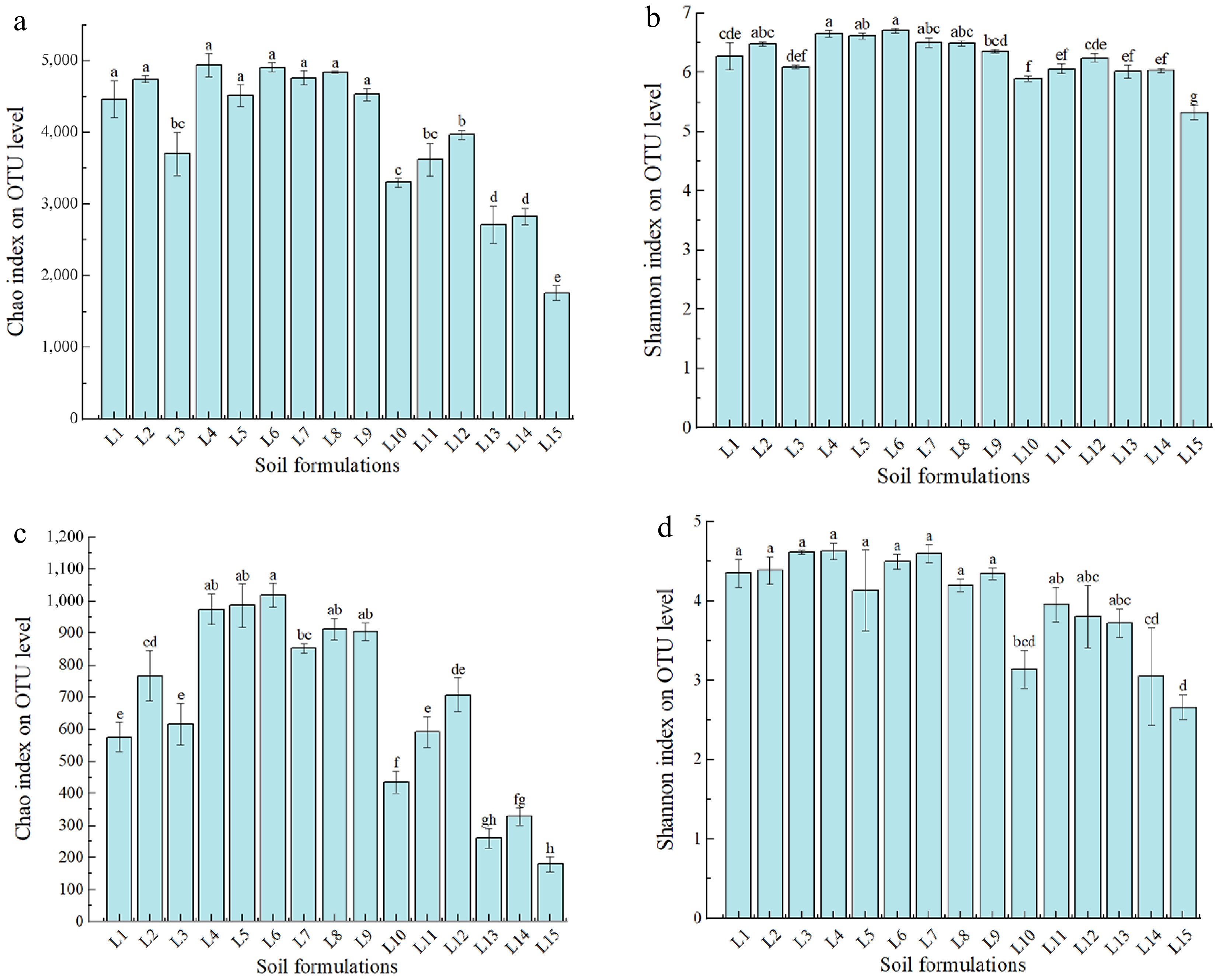
Figure 5.
Estimates of alpha diversity of the bacterial and fungal communities at the level of operational taxonomic units (OTUs) in different waste-mixed soils. (a) Chao index of bacteria. (b) Shannon index of bacteria. (c) Chao index of fungi. (d) Shannon index of fungi.
Beta diversity of microbial communities
-
The contribution rates of the first principal coordinate PC1, and the second principal coordinate, PC2, regarding the bacterial community variation in the treatments were 24.37% and 8.16%, respectively (Fig. 6a). The treatment groups with different proportions of waste slag soil were distributed in different quadrants, and the degree of mutual dispersion was large. The L4, L5, L6, and L9 treatments were clustered in the first quadrant, L2, L7, and L8 in the second quadrant, L1, L3, and L10 in the third quadrant, and L14 and L15 in the fourth quadrant. The scatter points of L2 and L7, L8, L1, and L3, L4, and L6 were relatively close, indicating a high composition similarity among samples. Therefore, results showed significant differences in soil bacterial communities under different treatments, while replicates of the same treatment had similar values.
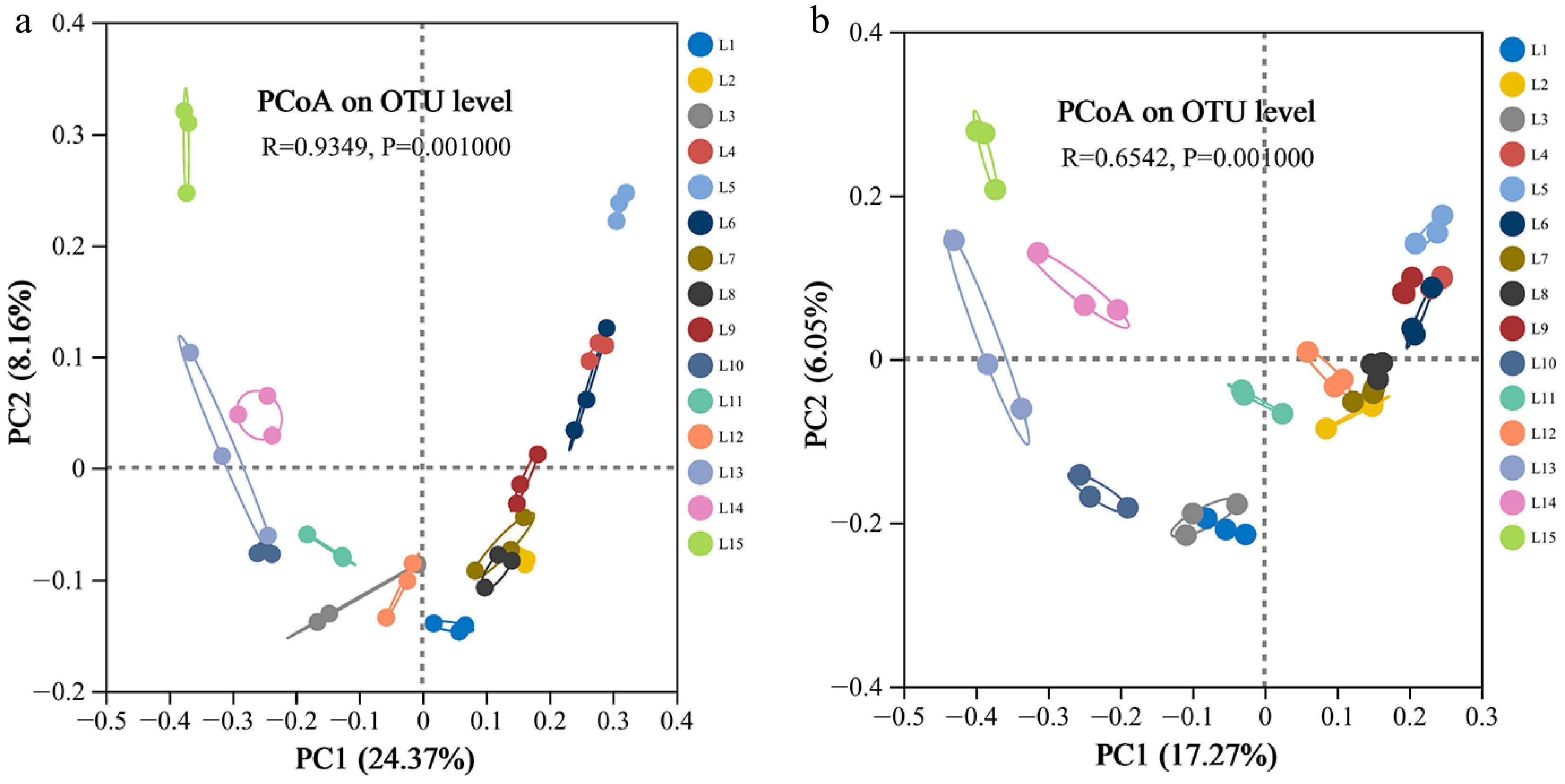
Figure 6.
Principal coordinates analysis (PCoA) of different treatments based on (a) bacterial, and (b) fungal operational taxonomic units (OTUs) abundance based on the Bray-Curtis heterogeneity.
The contribution rates of the PC1 and PC2 for the fungal community were 17.27% and 6.05%, respectively (Fig. 6b). Except for the L9 and L13 treatment groups, all the remaining treatments were consistently clustered in the same quadrant. L4, L5, and L6 were clustered in the first quadrant, L1, L2, L7, and L8 were clustered in the second quadrant, L3, L10, L11, and L12 were clustered in the third quadrant, and L14 and L15 were clustered in the fourth quadrant. The scatter points of L2, L7, and L8 were relatively close. These results indicated that there were significant differences in soil fungal communities under different treatments, while the soil fungal community structure of different replicates of the same treatment was relatively similar.
Redundancy analysis between microbial communities of soil samples and microbial communities of environmental samples
-
The first and second axes explained 35.71% and 6.53% of the variation in soil bacterial community structure, respectively (Fig. 7a). There was an acute angle between TN, AP, AK, and SOM, and an acute angle between coverage and aboveground biomass. Chloroflexi, Firmicutes, and Actinobacteriota were positively correlated with soil nutrient and plant growth indices, while Acidobacteriota was positively correlated with soil nutrient index and coverage, and Proteobacteria was negatively correlated with soil nutrient and plant growth indices.
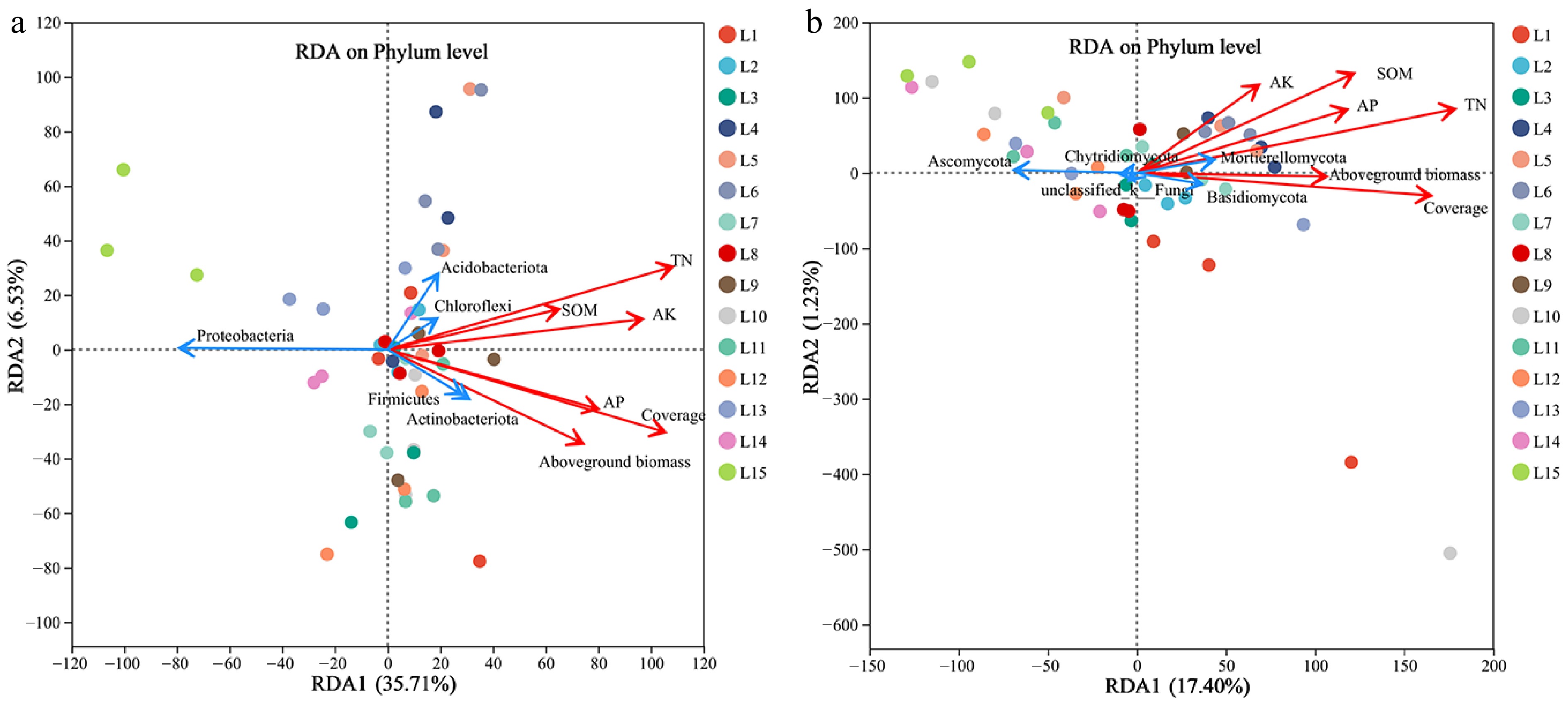
Figure 7.
Redundancy analysis of the relative abundance of soil bacterial (a) and fungal (b) phyla (top five), soil nutrients and plant growth indices under different treatments.
The first and second principal axes explained 17.40% and 1.23% of the variation in soil fungal community structure, respectively (Fig. 7b), with an acute angle observed between soil nutrient and plant growth indicators. Mortierellomycota and Basidiomycota were positively correlated with soil nutrient and plant growth indexes, while unclassified fungi, Chytridiomycota, and Ascomycota were negatively correlated with soil nutrient and plant growth indexes.
-
The particle size distribution of the waste slag soil is inappropriate, characterized by a high proportion of large particles and a markedly low nutrient content[30]. This hinders plant growth, explaining the poor results of Elymus nutans cultivation and growth in 100% waste residue soil in this experiment. The waste slag soil was mixed with other types of soil in different proportions to improve resource utilization. And the addition of soil to the waste slag increased the humidity of the substrate[31], which impacts plant growth. For example, using 60% slag + 20% peat soil + 20% loess matrix seed Ricinus communis L. seeds, seed germination rate and germination vigor, seedling survival rate, and growth vigor are better[32].
In the current study, the plants established in 100% guest and reclamation soils exhibited poorer growth than those established in 100% subgrade soil, which may be attributed to the high total N and available K contents in subgrade soil that favored plant growth. Adding an appropriate amount of waste slag soil has a certain promotional effect on plant growth. For example, tall fescue grown using a waste slag stone mixture with topsoil was taller, with greater root length, water content, and total aboveground biomass than those grown using topsoil only[33]. Covering a 30 cm soil layer with slag soil promoted crop growth and litter accumulation[34]. These results are similar to our findings since the L9-treated Elymus nutans had larger cover and aboveground biomass than the plants grown in the 100% guest and reclaimed soils. However, the use of a greater proportion of waste slag soil, such as 50% and 75%, affected the plant cover and aboveground biomass to a certain extent. In this study, Elymus nutans grown in the L13 and L14 soil mixtures exhibited cover below 50%, primarily attributable to the substantial presence of waste slag soil, which results in suboptimal soil structure, diminished water retention capacity, and nutrient deficiencies, as has been observed previously[35].
Effects of different slag ratios on soil nutrients
-
The waste slag exhibits lower amounts of soil organic matter, total N, total P, and available K, which is unfavorable to the growth and reproduction of plants. However, the soil can be improved by applying additional spent mushroom substrate[36] or biochar into sandy soils to increase nutrient contents[37], and promote plant growth[38] and microbial activity[39]. The nutrient contents of single subgrade soils were generally higher before soil blending, whereas waste slag soils had the lowest contents of total N, available P, and K and a higher content of organic matter. This study demonstrated that treatments with the greatest percentage of subgrade soil at a similar waste slag ratio exhibited the highest total N and available K content, suggesting that the incorporation of subgrade soil can effectively enhance these nutrients. Though the L5 treatment with a high percentage of subgrade soil had a higher soil total N content than the L9 treatment, the L9-treated plants exhibited higher growth than L5-treated plants, indicating that the plants absorbed N from the soil, resulting in a decrease in the total soil N content, similar to the previous results[40]. The change in soil available K content was equivalent to that of the total N and soil available P. In addition, the soil organic matter content was enhanced by the addition of a specific percentage of waste slag. This is likely due to the fact that waste slag contains coarser grains, which are difficult to erode by natural weathering in a short period and are loosely piled up with large gaps[2]. Additionally, waste slag may also improve the soil and water permeability, which is conducive to the growth of plant roots and microbial activities[41], while the decomposition of plant litter can provide nutrients to the soil, as has been previously observed[42].
Effects of different waste slag ratios on soil microbial community composition
-
Soil microorganisms make a major contribution to the preservation and regulation of the nutrient cycles of the soil ecosystem and function, but they are highly sensitive to environmental changes, resulting in shifts in the abundance of the soil[43]. The bacteria are the most numerous and abundant soil microorganisms that regulate soil quality[44] and represent the main driving force of soil organic matter and nutrient cycling[45], illustrating that their diversity is an indicator for assessing the quality of soil. The diversity and abundance of bacteria in the soil also directly affect the soil's ecological cycle[46]. In this study, the soil comprising 100% waste slag had the lowest number of bacterial OTUs and the least endemic bacteria, but the OTUs increased to some extent with the addition of other soil types. The differences in the number of bacterial phyla between the different treatments were small, but the relative abundance of the top five dominant bacterial communities at the genus level was lowest in the 100% waste slag treatment. This suggests that the addition of other soil types enhances the relative abundance of dominant bacterial communities, and more profoundly affects the microbial community composition at lower taxonomic levels, but less at the higher levels[47].
The 100% waste slag treatment group had the lowest Chao1 and Shannon indices, suggesting that the addition of other soils increased the diversity of dominant bacterial communities in each treatment, including Proteobacteria, Actinobacteriota, Acidobacteriota, Chloroflexi, and Firmicutes in the soil, consistent with the findings of Lin et al. on the improvement of cinder[48]. These microorganisms have been reported as the dominant flora in other related studies[49]. The relative abundance of Actinomycetes, Chloroflexi, and Firmicutes was elevated in treatments including the addition of other soils compared to 100% waste slag treatments. Actinomycetes are the predominant component of metabolically active bacteria in rhizosphere soils[50], Chloroflexi are bacteria that produce energy using CO2 as a carbon source[51], while Firmicutes are fast-growing bacteria that vigorously in high-carbon environments[52], and prefer soil environments with readily degradable organic matter and a slightly higher pH[53]. Consequently, the addition of other soils enhances their growth and reproduction. Acidobacteria were higher in the L5, L6, L8, and L9 treatment groups than in the L15 treatment, mostly because they metabolize carbon and N[54] and promote denitrification[55], and the plants that were cultivated in soils corresponding to the L5, L6, L8, and L9 soil mixtures had high aboveground biomass to provide a rich source of carbon. The phylum Proteobacteria was the most abundant across all 15 treatments due to its diversity in species composition and metabolic forms, and it plays a major role in maintaining soil ecosystems[56]. Notably, Proteobacteria were most abundant in L15 and least in L6. Proteobacteria demonstrate an absolute competitive advantage in the complex environment of rare earth mining areas[57], which may support their abundance in L15 treatment with pure waste slag. This is supported by the fact that Sphingomonas, a soil-dominant bacterial genus in Proteobacteria, which promotes plant growth and development in infertile soils[58], was more abundant in the L7 treatment and least abundant in the L15 treatment, corroborating the poor performance of plants grown in the L15 treatment waste soils.
Soil fungi possess a greater capacity to degrade complex compounds than soil bacteria and serve as the primary agents for the decomposition of plant and animal residues in soil[59]. They can decompose organic matter, such as sugars, starch, cellulose, lignin, and tannins, participate in the formation and decomposition of humus, and carry out ammonification and nitrification[60]. Therefore, the diversity and abundance of soil fungi play an important role in the material cycle and energy flow of ecosystems[61]. In the present study, the 100% waste slag treatment group had the lowest number of fungal OTUs, with the number of OTUs increasing when other soil types were added as mixtures. The 100% residue treatment group also had the lowest Chao1 and Shannon indices, suggesting that the addition of the other soil types to the treatments increased the abundance and diversity of the dominant fungal community in the soil.
Ascomycota, Basidiomycota, and Mortierellomycota were the most dominant fungal phyla in the present study, similar to other studies[62]. Ascomycota is a group of fungi present in large numbers in the soil that decompose keratin, cellulose, and lignin[63], and also cycle the soil nutrients[64]. Among these fungi, most of the saprophytic microorganisms like Basidiomycota and Mortierellomycota decompose lignocellulose, promoting humus utilization in the soil[65]. In this study, Ascomycota was most abundant in the L15 treatment and exhibited the lowest abundance in the L6 treatment with added waste slag, indicating a negative correlation of Ascomycota with soil moisture content. Pure waste slag treatments also contained low levels of soil nutrients, which corroborates the Ascomycota's dominance. In contrast, the relative abundance of both Basidiomycota and Mortierellomycota was lower in the L5 treatment since Basidiomycota are dominant in nutrient-rich ecosystems[66], while most of the beneficial Mortierellomycota are present in healthy soils[67]. This phenomenon may have been ameliorated by the addition of other soil types, which improved the nutrient status of the soil and favored the growth of Basidiomycota and Mortierellomycota.
Effects of different residue ratios on plant-soil-microbe interactions
-
As an important component of terrestrial ecosystems, the interaction between plants, soil, and microorganisms drives the flow and circulation of nutrients in response to external environmental changes[68]. For example, plant nutrients originate from the soil, while soil nutrients are derived from the microbial population[69]. It has been demonstrated that the appropriate DCGS (coal-based solid waste) application is a potential approach to improve soil quality and facilitate the growth of Leymus chinensis in a soil-plant-microbe system[70]. In this study, both plant coverage and aboveground biomass were positively correlated with soil total N, available P and K, and organic matter, bacterial communities (Chloroflexi, Firmicutes, Actinobacteriota), and fungal communities (Mortierellomycota and Basidiomycota), on the one hand, and plant nutrient supply was mainly dependent on microbial decomposition functions. Soil microorganisms have the properties of decomposing organic carbon components such as cellulose and lignin in soil organic matter[71], while plant litter is a source of soil organic matter and nutrients for soil microorganisms. The lower carbon-to-nitrogen ratio of plant litter facilitates the development of microbial communities, enhancing decomposition and nutrient release, thereby improving soil nutrient content[72].
In this study, a positive correlation was identified between soil total N, available P, available K, and organic matter and plant growth indicators, bacterial communities, and fungal communities. This correlation can be attributed to the low degree of weathering of waste slag, the inter-root effect of plant root secretions, uptake of weathering products by plants, and the auxiliary influence of plant symbiotic mycorrhizae, which can facilitate the weathering of waste slag[73]. The survival and reproduction of microbial communities are also closely linked to soil nutrients, and changes in their function are positively correlated with changes in soil nutrients[74]. When plant and soil nutrients have an effect on microorganisms, microorganisms also react to both. In this study, the plant growth indexes and soil nutrient indexes of the treatments with higher microbial abundance, α diversity, and β diversity were higher than those of the treatments with lower microbial abundance, α diversity, and β diversity. Soils with high microbial diversity may accelerate the turnover rate of nutrients, which implies that microbial diversity facilitates more nutrients for plant growth[75]. This suggests that bacterial diversity and abundance in the soil directly influence the ecological cycle, including nutrient cycling and energy transformation within the soil[46]. Overall, soil microbial communities promote positive plant-microbe interactions[76].
-
Various residue mixing ratios had varying effects on soil microorganisms, soil nutrients, and plant growth. The addition of subgrade soil, guest, and reclaimed soils was more beneficial to plant growth than the treatments with individual residues. This was further manifested by the increase in the vegetation cover and aboveground biomass of Elymus nutans, the increase in the content of soil nutrients, including the total N, available P, available K, and organic matter, and the increase in the number of OTUs and diversity indexes of soil bacteria and fungi. Considering plant growth, soil nutrients, soil microorganisms, and the use of waste slag resources, the L9 treatment (25% waste slag + 75% subgrade soil) was the most optimal soil mixture that can facilitate the ecological restoration area of railway engineering in high-altitude areas.
-
The authors confirm contributions to the paper as follows: methodology, software: Chen B, Liu M; investigation: Liu M; supervision, resources, project administration: Chai Q; writing – original draft, data curation: Chen B; writing – review & editing: Chen B, Chai Q, Lu J, Yang C, Chen A. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
-
We thank the China Construction Railway Investment and Construction Group Co. for providing the experimental basic platform for this study. We also thank Shanghai Meiji Biomedical Technology Co. for determining the microbiological indicators. We would like to thank MogoEdit (www.mogoedit.com) for its English editing during the preparation of this manuscript.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Benju Chen, Minting Liu
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen B, Liu M, Chai Q, Lu J, Yang C, et al. 2025. Effects of waste slag soil reconstruction on the growth of Elymus nutans and microbial diversity in the western Sichuan Plateau. Grass Research 5: e021 doi: 10.48130/grares-0025-0018 

Effects of waste slag soil reconstruction on the growth of Elymus nutans and microbial diversity in the western Sichuan Plateau
- Received: 05 March 2025
- Revised: 14 June 2025
- Accepted: 02 July 2025
- Published online: 20 August 2025
Abstract: This study explored the effects of mixing different proportions of railway engineering waste slag with other types of soils to improve waste resource utilization of railway engineering waste slag in high-altitude areas. Waste slag, subgrade soil, guest, and reclaimed soils were mixed in different volume ratios for a total of 15 treatments and used to grow Elymus nutans spanning two growing seasons. Subsequently, the diversity of the soil microbiome was assessed using high-throughput sequencing analyses. The L9 treatment (25% waste slag + 75% subgrade soil) resulted in the highest plant cover and aboveground biomass, while the L15-treated (100% waste slag) plants were unsuccessfully established in the first year. The soil's total nitrogen, available phosphorus and potassium, and organic matter content were significantly increased (p < 0.05) in the L9 treatment when compared to the L15 treatment. The bacterial and fungal communities at the genus level were more abundant and diverse in all other types of soil treatments compared to the 100% waste slag treatment. The bacteria and fungi in the L4 (75% subgrade soil + 25% soil + 25% guest soil & reclamation soil), L5 (100% subgrade soil), L6 (25% subgrade soil + 75% guest soil and reclamation soil), and L9 treatments were clustered in one quadrant, according to β-diversity analysis, indicating the similarity in the microbial community structure in these treatments. Therefore, the addition of other types of soil can improve slag and positively affect plant growth, soil nutrient content, and the soil microbiome. The study recommends mixing 25% waste slag with 75% subgrade soil to improve railway engineering waste slag utilization in high-altitude ecological restoration areas.