









-
Mango (Mangifera indica) is a commercially significant crop of the Anacardiaceae family, which is also known as the king of tropical fruits. It is the fifth most grown fruit globally and is known for its nutritional richness, including vitamins, bioactive compounds, carbohydrates, organic acids, lipids, and proteins[1]. They are not only eaten fresh but are also generally processed into numerous products, such as powders, jams, juices, nectars, and jellies[2]. China ranks as the 2nd largest mango producer in the world, with major growing regions in Hainan, Guangxi, Yunnan, and Sichuan provinces[3]. Mango domestication is over 4,000 years old, and it was first domesticated in Southeast Asian and Indo-Burma, with its global dissemination occurring by the 14th century[4]. Although mango production is economically significant, it has a number of environmental issues that influence the crop yield[5]. Some of these challenges are UV radiation, hot temperatures, bright lights, salinity, and drought that may affect the growth of plants[6]. The UV-B radiation (280–315 nm), which is mostly caused by depletion of the ozone layer, is now a significant plant abiotic stressor. These radiations may have detrimental effects on the growth, development, and productivity of the plants[7]. These adverse conditions induce the overproduction of ROS, such as hydrogen peroxide (H2O2), superoxide anion (O2−), and hydroxyl radicals (OH−), which damage the cellular membranes, organelles, and other macromolecules, and ultimately result in cell death[8]. Plants have developed complex antioxidant systems with enzymatic and non-enzymatic compounds as a mechanism of alleviating the damage caused by ROS.
One of the most important ROS-scavenging enzymes is Ascorbate peroxidase (EC: 1.11.1.11), an important heme-containing enzyme that keeps the redox homeostasis in plants. It is involved in the reduction of H2O2 to water with ascorbate as the particular electron donor[9−11]. This activity underscores its critical role in plant defence mechanisms, particularly in stress tolerance[12,13]. APX originated as a soluble enzyme in chlorophytes, and over evolutionary time, it diversified in land plants through gene duplications, resulting in the formation of cytosolic, peroxisomal, and chloroplastic isoforms[14]. In land plants, chloroplastic APX also underwent alternative splicing, producing both soluble and thylakoid-bound enzymes. Furthermore, in certain angiosperms, independent duplications gave rise to distinct soluble and membrane-bound APX isoforms[15]. This diversification highlights the functional specialization of APX proteins across different plant species. In Arabidopsis thaliana, eight APX genes have been identified, which include three cytosolic (APX1, APX2, APX6), three peroxisomal (APX3, APX4, APX5), and two chloroplastic isoforms (sAPX and tAPX)[16−18]. A similar distribution of APX genes is maintained in rice (Oryza sativa), which contains eight genes, with two each for cytosolic, peroxisomal, mitochondrial, and chloroplastic isoforms[19,20]. Similar APX gene families are found in other species, tomato (Solanum lycopersicum), upland cotton (Gossypium hirsutum), sorghum (Sorghum bicolor), and kiwifruit (Actinidia chinensis)[21−24]. The APX gene family is highly diversified in terms of size in various species, which is a sign of the evolutionary diversification and functional specialization of these enzymes. For example, sorghum contains nine APX genes[23], while peanut has an exceptionally high count of 166 APX genes[25]. The cytosolic APX enzymes are significant in stress conditions, including high temperatures, as they scavenge H2O2 and reduce photo-oxidative stress, which in turn enhances plant tolerance to abiotic stress[26,27]. These enzymes are crucial for preserving redox equilibrium within cells and defending plants from oxidative stress triggered by ROS, which is a by-product of stress. Other than its role in antioxidant metabolism, recent reports have revealed an additional role of APX in lignin biosynthesis, whereas Arabidopsis and Brachypodium distachyon have been shown to exhibit coumarate 3-hydroxylase, which is involved in lignin biosynthesis[28]. Nevertheless, in earlier experiments, it was shown that improved UV-B stress led to APX gene upregulation, leading to an increase in the amount of ascorbate in fruits, thus enhancing their capacity to counteract ROS[29].
Although APX plays a significant role in plant development and stress tolerance, comprehensive research on this gene family in mango is limited. The objective of the study is to characterize the occurrence and localisation of MiAPX genes in mango chromosomes as well as to examine the functional diversity of the genes in terms of exon-intron structure, gene-regulatory frameworks, physicochemical characteristics of the encoded proteins, and the phylogenetic relations. Further, to profile the gene expression under different stress conditions, transcriptomic and quantitative real-time polymerase chain reaction (qRT-PCR) analyses were done. These findings provide insight into the potential roles of MiAPX genes in stress tolerance and hormone-mediated regulation, which will be the basis for further studies on the response of mango to biotic and abiotic stresses.
-
The APX gene family in mango was determined by conducting the BLASTP searches using the reference genome of Alphonso present on the NCBI database (
www.ncbi.nlm.nih.gov/datasets/genome/GCF_0110750555.1 ). Ultimately, ten APX gene family members of the mango were found. In order to obtain some functional information, sequences of the candidate genes were subjected to domain identification and functional analysis through the InterPro database (www.ebi.ac.uk/interpro ) to identify their possible functions[30]. To examine evolutionary relationships, the mango APX protein sequences were used to identify homologs in A. thaliana and rice through the NCBI platform, resulting in the identification of eight A. thaliana, and eight rice family members. The ProtParam tool (https://web.expasy.org/protparam ) was used to identify physicochemical parameters such as molecular weight, isoelectric point (pI), amino acid content, and GRAVY score. Subcellular localization predictions were done on Cell-PLoc 2.0 (www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2 ).Three-dimensional (3D) and secondary structure of APX genes
-
The 3D model of the APX proteins was generated with the help of the web tool of the platform Swiss-Model (
https://swissmodel.expasy.org )[31]. To analyze the secondary structure, the SOPMA software (http://npsa.lyon.inserm.fr/cgi-bin/secpred_sopma.pl ) was applied[31]. Additionally, selected sequences were further subjected to detailed 3D structural modeling using the SWISS-MODEL platform.Gene structures, domains, and conserved motifs analysis
-
Motif analysis of the APX gene family was performed using the MEME suite (
https://meme-suite.org/meme/tools/meme ), which identified up to ten conserved motifs[32]. To analyze domain architecture, the NCBI's Batch CD-Search tool (Conserved Domain Database) was used[33]. Gene structure and domain visualization were carried out using TBtools-II v2.096[34].Chromosomal mapping and Ka/Ks ratio evaluation
-
Ka/Ks ratios were calculated using MEGA 11 to test the selection pressure, whereby a value above 1 indicated positive selection, = 1 neutral evolution, and a value below 1 purifying selection[35]. Moreover, the calculation of gene divergence times was done through the following formula given by Tahir ul Qamar et al.[36]. The homology analysis of APX genes of M. indica, rice, and A. thaliana was done with the help of TBtools-II v2.096[37].
Phylogenetic relationship of the APX gene family in mango
-
A multiple sequence alignment was used to determine the evolutionary relationship among mango APX genes and their homologs in A. thaliana and rice with the help of the MUSCLE algorithm. The construction of a phylogenetic tree was done using the neighbour-joining (NJ) method of MEGA 11 software[38]. Bootstrap tests were done (1,000 replications) to ensure that the tree is stable. The resulting tree was then plotted in a graphical way via (iTOL) platform (
https://itol.embl.de/ )[38] to be visualized and compared with.Identification of cis-elements in the promoter region of APX genes
-
The 2,000 bp upstream promoter sequences of mango APX genes were purified to examine regulatory sequences. The presence of Cis-acting regulatory elements in these regions was tested in the plantCARE database (
https://bioinformatics.psb.ugent.be/webtools/plantcare/html )[39]. Besides, the identified regulatory motifs were graphically displayed with the help of TBtools-II software (2.096).Plant materials and sample collection of mango
-
The experiment was carried out on a 15-year-old Tainong 1 mango orchard in Sanya, Hainan, China (18°25′ N, 109°46′ E), at an average temperature of 25 °C and annual rainfall of 1,700 mm. The UV-B treatment (313 nm, 96 kJ·m−2·d−1) was done by placing four 40 W lamps 30 cm above the canopy, which simulated a 15% increase in UV-B. Mango trees were randomly assigned to control and UV-B treatment groups, with five fruits from each tree sampled at 30, 40, and 91 d after flowering. All the samples were then kept at −80 °C.
Validation by qRT-PCR
-
Ten APX genes with differential expression rates were used to validate the transcriptome data by using qRT-PCR. Primer Premier 6 was used to design the primers which were synthesized by Shanghai Bioengineering Co., Ltd (Shanghai, China, Supplementary Table S1). The total RNA was isolated, reverse transcribed using the HiScript II kit, and qRT-PCR was performed on the qTOWER3 system with the SYBR Master Mix. Actin was used as the reference gene, and the relative expression was determined using the 2−ΔΔCᴛ technique[40].
Statistical analysis
-
A one-way ANOVA was conducted to determine significant differences among the experimental groups. Data visualization and graphical analyses were carried out using GraphPad Prism 8.0.1. Additionally, heatmap expression patterns of DEGs were created using TBtools-II (v2.096)[34].
-
Ten members of the APX gene proteins were identified in the mango genome and were named MiAPX1 to MiAPX10. Physicochemical parameters of the respective APX proteins, such as molecular weight and isoelectric point (pI), were analysed (Table 1). The MiAPX genes varied in length from 240 to 621 amino acids. MiAPX8 and MiAPX9, respectively, coded the smallest and the largest protein. The molecular weights were found to be about 26.4 to 69.3 kDa, and this was due to structural diversity in the family. Among all the members, six were acidic, with four being basic. Whereas eight genes were stable with a measure of instability below 40, and two not stable. Additionally, the negative GRAVY scores for all MiAPX genes suggest that they are predominantly hydrophilic. This study offers an in-depth characterization of the MiAPX gene family, highlighting its structural diversity and physicochemical properties. The findings contribute valuable information for further functional studies on MiAPX genes and their roles in mango physiology and stress responses.
Table 1. Biological features of the APX gene family.
Gene IDs Given name A.A MW (kDa) pI Instability index Aliphatic index Gravy Subcellular localization LOC123198607.1 MiAPX1 246 27,152.47 5.38 39.61 81.34 −0.433 Peroxisome LOC123212144.1 MiAPX2 289 31,770.3 7.69 37.02 86.85 −0.272 Peroxisome LOC123214773.1 MiAPX3 286 31,561.02 6.67 36.65 84.93 −0.325 Peroxisome LOC123228152.1 MiAPX4 286 31,574.88 6.72 36.32 83.92 −0.366 Peroxisome LOC123201325.1 MiAPX5 250 27,500.05 5.38 32.41 80.04 −0.388 Peroxisome LOC123209556.1 MiAPX6 245 26,945.6 5.04 32.41 79.71 −0.255 Peroxisome LOC123224991.1 MiAPX7 249 27,394.17 5.52 28.7 78.84 −0.33 Peroxisome LOC123200457.1 MiAPX8 240 26,441.18 5.39 33.99 91.12 −0.135 Peroxisome LOC123203234.1 MiAPX9 621 69,336.45 5.9 48.18 75.3 −0.526 Chloroplast, mitochondrion Analysis of the secondary structure
-
The ten APX proteins were selected and analyzed for secondary structure using the SOPMA online server. The analysis of the selected individual proteins showed that they had different ratios of alpha-helices, extended strands, bends, and random coils (Supplemental Fig. S1). These variations in secondary structural components affect the overall conformation, rigidity, flexibility, and interactions with other molecules. The α-helices ranged from 32.72% in MiAPX10 to 43.71% in MiAPX4, extended strands from 9.09% in MiAPX4 to 12.60% in MiAPX1, β-turns varied from 2.77% in MiAPX2 to 5.78% in MiAPX1, and random coils from 41.87% in MiAPX1 to 54.95% in MiAPX10 (Supplementary Table S2).
Investigating the 3D structures
-
The ability to predict 3D protein models links the gap between protein structure and sequence, making it easier to analyze activities and interactions with other proteins. The 3D structures of ten MiAPX genes were generated using SWISS-MODEL (Fig. 1). The results revealed that all genes were identified as monomers. The GMQE value of these proteins varied from 0.79 to 0.97, whereas the sequence identities revealed a range of variability, with high values observed in MiAPX3 (86.715%), MiAPX7 (85.83%), MiAPX4 (85.31%), MiAPX5 (85.08%), MiAPX9 (80.885%), and MiAPX6 (80.82%), suggesting these proteins have more reliable structural models. In contrast, some moderate identities were also found, like MiAPX1 (78.37), MiAPX8 (77.08), MiAPX2 (76.33), and MiAPX10 (75.08) (Fig. 1, Supplementary Table S3).
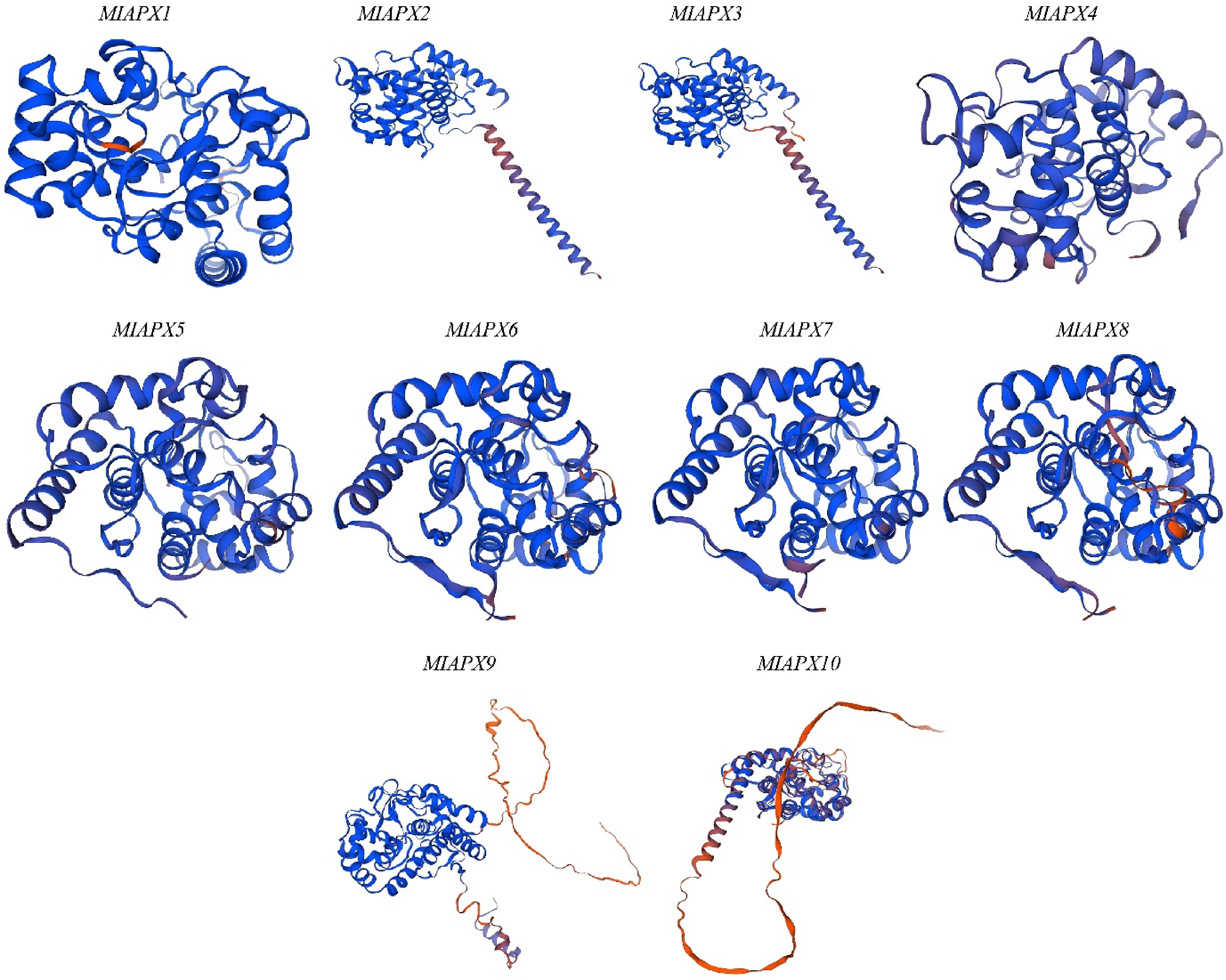
Figure 1.
Represents the 3D structures of ten MiAPX genes which were generated using SWISS-MODEL.
Conserved motifs, conserved domains, and gene structure analysis
-
Conserved motifs of the mango MiAPX gene sequences were predicted and identified using the MEME online analysis tool. The statistical findings showed that there were ten conserved motifs (named motif 1–10) in ten members of the MiAPX gene family. Distinct differences in the conserved motifs were found in each cluster. For example, MiAPX10 has seven conserved motifs, whereas the other genes have eight to ten motifs (Fig. 2a). Among these, motif three is widely distributed, appearing in 100% of the MiAPX family members, indicating its prominence and potential importance. These results indicate that some motifs, such as motif 3, are highly conserved motifs that might have critical functions in some vital biological processes. The combined outcomes of phylogenetic tree and motif analysis further identified unique motif distribution patterns between different branches, which emphasized the evolutionary and functional diversity in the MiAPX gene family.
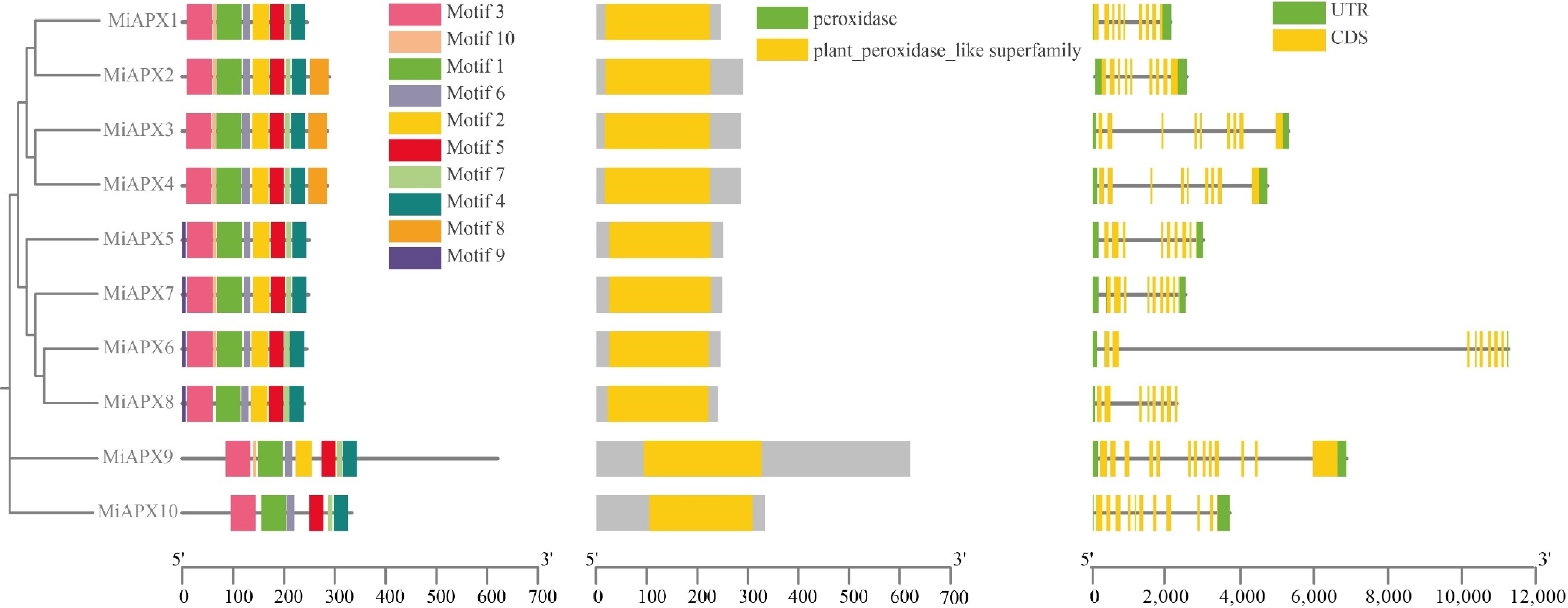
Figure 2.
Comparative analysis of APX gene motifs, domains, and structures. (a) Boxes of different colors represent the ten conserved motifs. (b) APX family protein domains. (c) The UTR–exon organization of APX genes, with UTRs shown in green, and exons (CDS) shown in yellow. Introns are represented by black lines connecting the exons.
The analysis of the ten identified domains of MiAPX genes was done in Fig. 2b, showing that all the genes belong to the plant peroxidase-like superfamily. This consistent presence of the plant peroxidase-like domain across all MiAPX genes highlights their functional conservation and suggests a shared role in peroxidase-related biological processes within the MiAPX gene family.
To explore the structural diversity of duplicated genes, it is crucial to examine their gene structures. The intron and exon can give useful information about the diversity and evolutionary connections between the APX gene family in mango. Genomic DNA sequences of the MiAPX genes were aligned with their full-length cDNA sequences to visualize gene structures. This study compared the structural composition of ten MiAPX genes with the help of the mango genome, focusing on exon/UTR positions. The UTR/exon structure, along with the type and number of introns, served as a hallmark of gene family evolution. The analysis revealed that nine MiAPX genes contain two introns, while MiAPX8 has eight exons, and all other genes possess nine to 11 exons (Fig. 2c). To further assess the structural properties of these genes, a phylogenetic tree and exon/ intron structure map were drawn with the highlighting of the UTRs.
Chromosome distribution, collinearity, and Ka/Ks analysis
-
Chromosomal mapping was conducted to determine the genomic positions and gene duplication of the MiAPX gene family members in the mango genome. The outcome of the mapping indicated the uneven distribution of the ten MiAPX genes in nine out of the 20 chromosomes. Chromosome 2 had the greatest number of genes with two MiAPX genes, while all other chromosomes had one MiAPX gene (Fig. 3a).
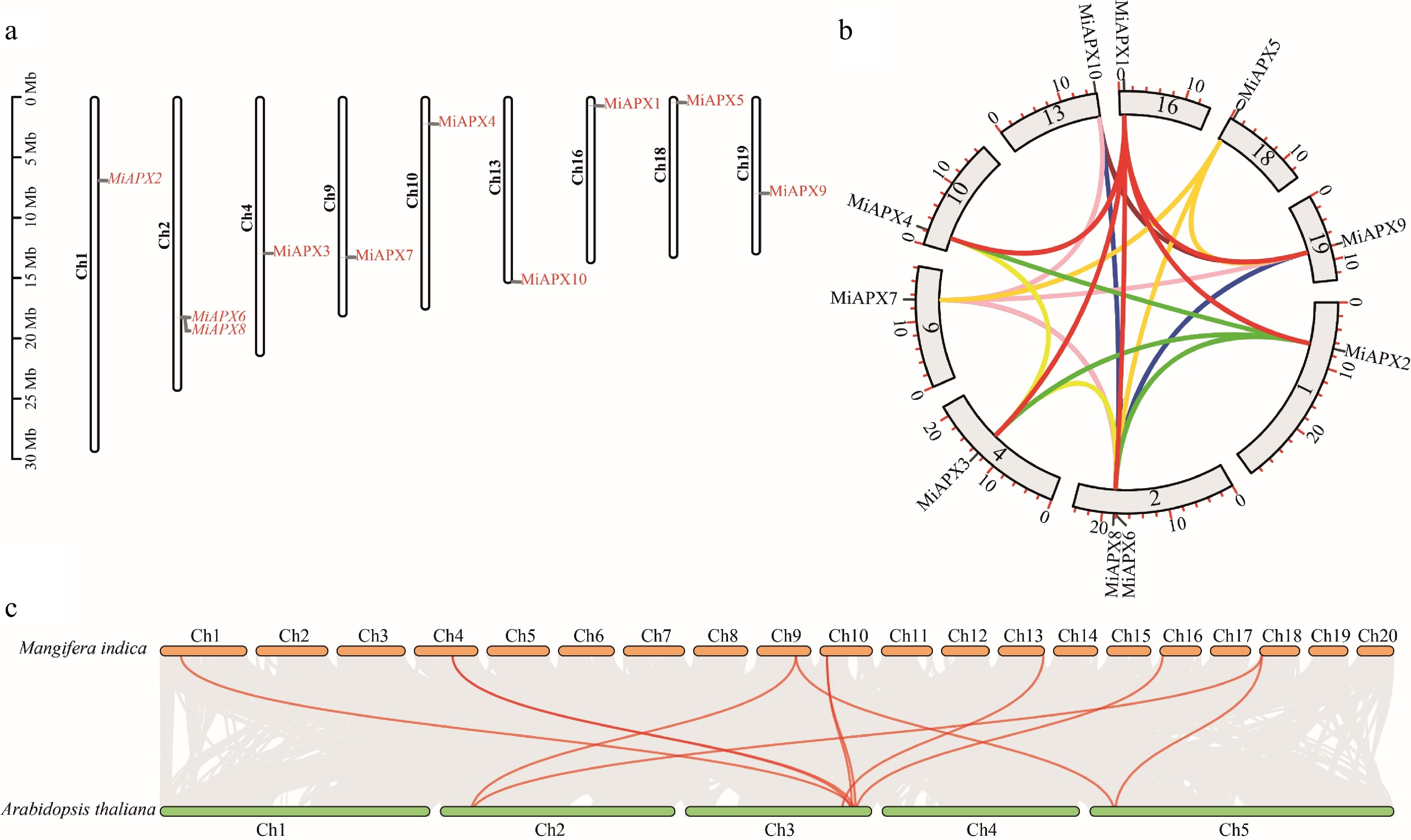
Figure 3.
(a) The distribution of ten MiAPX genes across ten mango chromosomes is shown, with genes marked in red and chromosomes in black. (b) A circos plot illustrates the duplicated MiAPX gene pairs within the mango genome. (c) Syntemic relationships between APX genes in mango and A. thaliana were shown, with orange and green boxes representing the respective genomes. Red lines highlight conserved orthologous gene pairs.
To explore the evolutionary dynamics of the MiAPX gene family, the Ka/Ks substitution rates were estimated for each gene pair. (Supplementary Table S4). Twenty-six homologous pairs of genes within the MiAPX gene family were identified. The values of Ka/Ks showed that all pairs had values lower than one, which implied they had experienced purifying selection. The divergence of these 26 pairs of duplicated genome-wide gene pairs was 8.10–106.01 million years ago (Mya). Analysis of Ka/Ks ratios and divergence time indicates that the MiAPX genes have been under a strong purifying selection in the course of their evolution.
Gene duplication is a key feature of plant genomics and has a role in the functional diversification of gene families. It may occur in various mechanisms, such as segmental and tandem duplications. In order to determine whether gene duplication contributes to MiAPX gene family evolution in mango, duplication events of the MiAPX genes were compared. Out of 26 homologous pairs, only one was found to be the tandem duplication, while the rest were categorized as segmental duplications (Fig. 3b). In order to further explore the syntenic relationships and evolutionary processes, collinearity analysis of the A. thaliana and mango genomes was performed (Fig. 3c). This comparison has shown that there are 11 MiAPX genes that are highly collinear with the APX genes in A. thaliana, as described in Supplementary Table S2.
Phylogenetic study of APX family members
-
Protein sequences were used to get phylogenetic analysis of mango, A. thaliana, and rice. The APX genes were divided into four major clusters (Groups 1–4), according to domain composition and phylogenetic clustering as indicated in Fig. 4. The largest cluster was Group 1, which included seven APX genes, including one of mango, two of Arabidopsis, and four of rice. Group 2 consisted of eight members, comprising four each of mango, Arabidopsis, and rice. Group 3 was also comprised of eight APX genes with four of mango, two of Arabidopsis, and two of rice. Group 4 consisted of three members, one of mango and two of Arabidopsis. Notably, certain MiAPX genes displayed close evolutionary relationships with specific Arabidopsis genes. For instance, in Group 1, MiAPX9 was closely related to AtAPX8; in Group 2, MiAPX5 clustered with AtAPX2; and in Group 4, MiAPX10 showed a close relationship with AtAPX6.
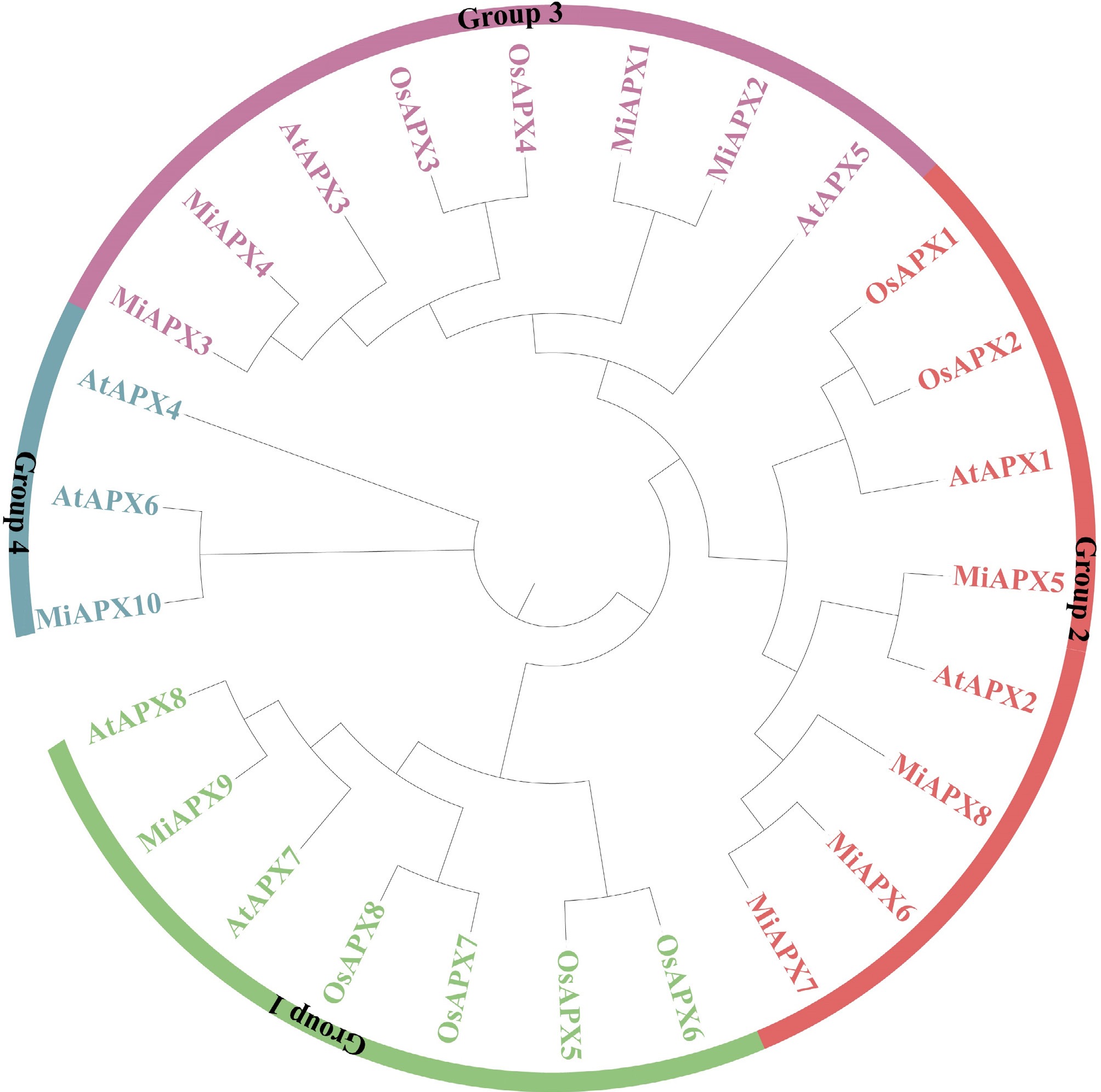
Figure 4.
Phylogenetic tree of MiAPX genes from A. thaliana, rice, and mango. The tree was constructed using the maximum likelihood method with 1,000 bootstrap replications in MEGA 11. Different colors represent distinct groups.
Cis-acting component analysis of the APX family promoter
-
Cis-acting elements are essential sequences in DNA found in the promoter regions of the genes that are involved in the regulation of transcription through interactions with transcription factors. This analysis involved the analysis of the 2,000 bp upstream areas of ten MiAPX genes to determine the possible regulatory motifs. After removing uncharacterized sequences, the identified elements were classified into three broad categories, i.e., elements related to abiotic stress responses, including plant hormone signaling, and growth or development (Fig. 5). In the stress-related group, four elements were discovered, which belonged to the light responsiveness, anaerobic conditions, drought, and low-temperature responses. These included: ACE, AE-box, Box 4, GATA, G-Box, I-box, and TCT, containing 79.31% of light-responsive elements, while ARE, LTR, and MBS accounted for 14.48%, 3.45%, and 2.75%, respectively. The abundance of light responsiveness might be an important regulator of photomorphogenesis. Plant hormones have six types, which were abscisic acid (ABRE, 27.18%), auxin (TGA, 5.82%), gibberellin (GARE and P-Box, 2.91%), methyl jasmonate (TGACG, 36.90%), salicylic acid (TCA, 6.79%), and ethylene (ERE, 20.39%). It is important to note that the most common ones were TGACG (methyl jasmonate) and ERE (ethylene), which comprised 35.2% and 29.5% of the elements, highlighting the involvement of the MiAPX family in hormone regulation. Additionally, six elements that are associated with growth and development were also identified, including meristem expression (CAT-box, 11.11%), circadian control (circadian, 11.11%), seed specificity (RY-element, 11.11%), zein metabolism (O2-site, 16.67%), MYBHv1 binding sites (CCAAT-box, 44.44%), and endosperm expression (GCN4, 5.55%). The dominance of CCAAT-box elements (44.44%) highlights the possibility of involvement of MiAPX genes in developmental processes. The Cis-acting element analysis demonstrates the widespread participation of the MiAPX gene family in photomorphogenesis, hormonal signaling, and other growth and developmental mechanisms. The abundance and diversity of these elements suggest that MiAPX genes are critical for mango's adaptive responses to environmental stimuli and developmental regulation.
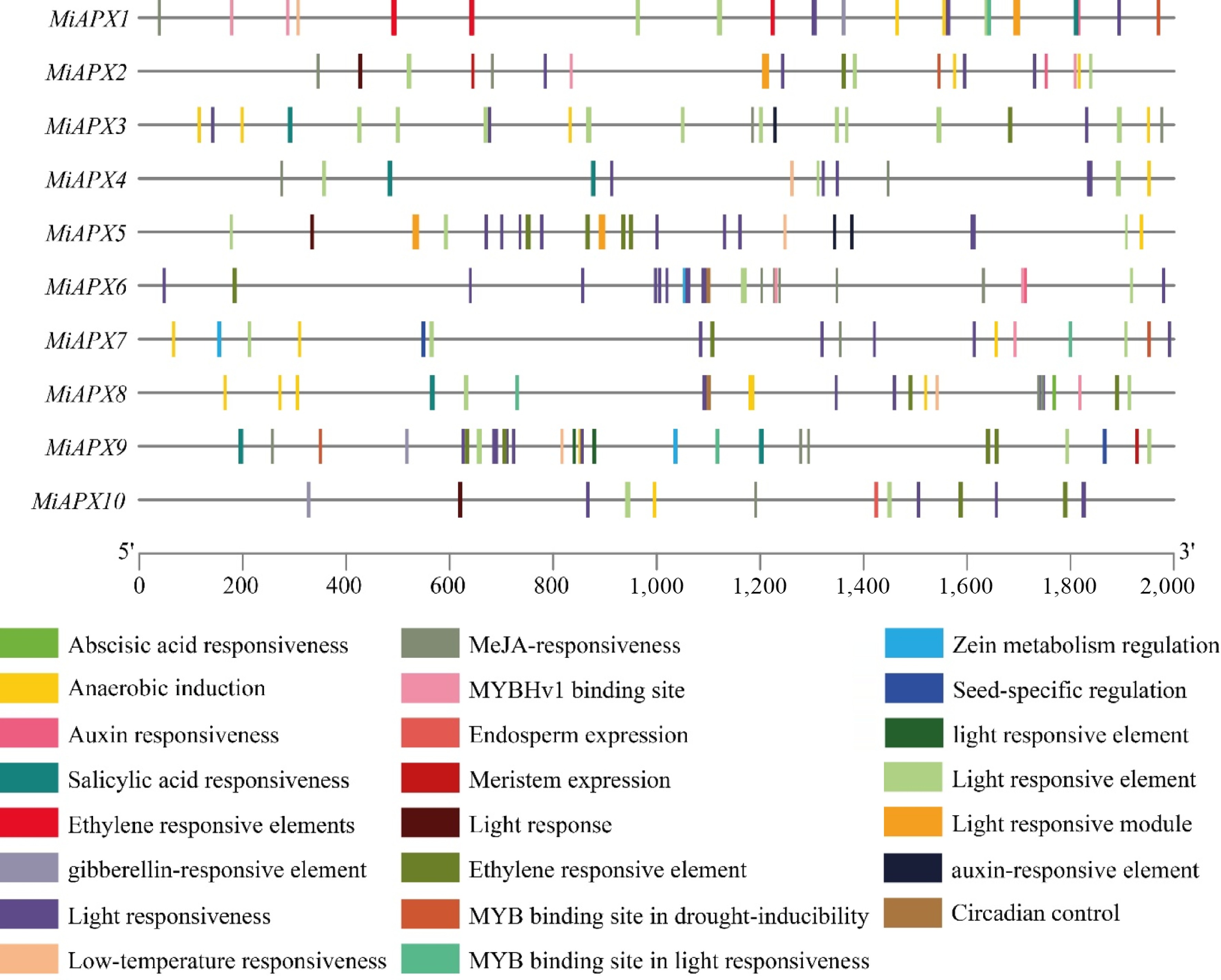
Figure 5.
The Cis-acting elements in the mango promoter, with altered colours representing each element and their respective proportions.
Expression analysis and qRT-PCR validation of APX gene family under UV-B stress in mango
-
To investigate the effects of UV-B treatment on mango pulp, the expression levels of APX genes associated with the AsA metabolism pathway were analyzed and compared with a control group. Transcriptome analysis indicated that there were seven APX genes whose expression was significantly upregulated by the 40th day of the UV-B treatment (Fig. 6a). On the 91st day, six APX genes exhibited increased expression levels.
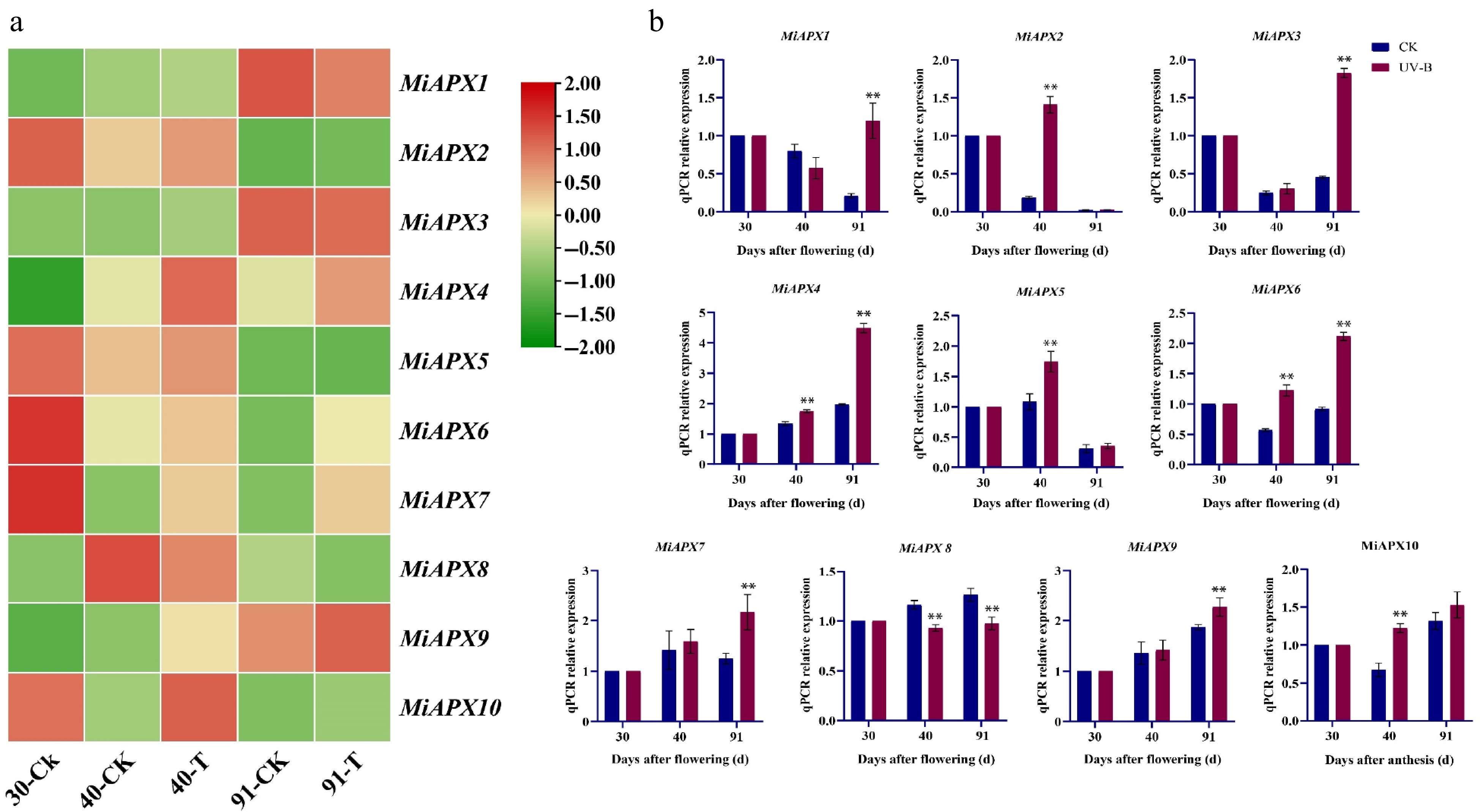
Figure 6.
(a) Heatmap showing the expression patterns of MiAPX genes in response to UV-B stress. (b) qRT-PCR validation of ten MiAPX genes in samples collected at 30, 40, and 91 d after flowering under UV-B treatment. The x-axis represents days after flowering (d), and the y-axis indicates the relative expression levels of APX transcripts.
Nonetheless, ten APX gene members were identified and studied to get a better understanding of their physiological functions in mango fruit pulp subjected to UV-B stress (Fig. 6b). These genes were analyzed by the expression levels with the help of qRT-PCR at various developmental stages (30, 40, and 91 d). By the 40th day, MiAPX2, MiAPX4, MiAPX5, MiAPX6, and MiAPX10 were significantly upregulated, whereas MiAPX8 was downregulated upon UV-B treatment. The expression patterns changed by the 91st day, with MiAPX1, MiAPX3, MiAPX4, MiAPX6, MiAPX7, and MiAPX9 found to be upregulated in response to UV-B stress whereas MiAPX8 continued to exhibit downregulation.
-
Mango production faces several environmental challenges that include tolerance to high temperatures, salt, drought, high light intensity, and UV-radiations that impact the productivity of the plant[41]. Plants coping with these issues have an effective antioxidant system, which consists of enzymatic and non-enzymatic elements that maintain redox and osmotic homeostasis, enhancing tolerance to environmental stresses[42]. Exposure to stress can lead to the formation of ROS, including singlet oxygen, superoxide, H2O2, and hydroxyl radical, which are detoxified mainly through the AsA–GSH cycle, whereas APX plays a key role[43,44]. APX is a class I heme-peroxidase that can be found mostly in the higher plants, chlorophytes, and red algae, with gene families varying across species[42]. It is critical in ROS scavenging and plant development[22]. Despite extensive characterization of APX genes in other species, their roles in mango remain unexplored. Advancements in mango genome sequencing offer critical insights into the APX gene family, enabling deeper exploration of their roles and regulatory mechanisms. Ten APX genes have been identified in mango, exceeding the number in rice and A. thaliana but fewer than in cotton[22,24,45].
The MiAPX gene family in mango comprises ten members (MiAPX1-MiAPX10), varied in length from 240 to 621 amino acids and 26.44–69.34 kDa molecular weights (Table 1). Six proteins are acidic, four are basic, and eight are stable, with two unstable proteins. All proteins are hydrophilic. These properties reflect the diverse structural characteristics of the MiAPX gene family in mango[24]. APX proteins are categorized into cytoplasmic, peroxisomal, chloroplastic, and mitochondrial types, each serving distinct functions in maintaining redox balance. Cytosolic APX regulates H2O2 levels during stress and development, coordinating inter-organelle signaling. Chloroplastic APXs protect the photosynthetic apparatus from photooxidative stress[46]. Peroxisomal APX works with catalase to control H2O2 leakage and redox communication, while mitochondrial APX maintains redox balance during stress and senescence[47]. In this study, phylogenetic analyses and subcellular localization predictions were consistent, with peroxisomal subfamily members (MiAPX1, MiAPX2, MiAPX3, MiAPX4, MiAPX5, MiAPX6, MiAPX7, MiAPX8), while MiAPX9 was found in mitochondria and MiAPX10 was found in cytoplasm (Table 1). These findings align with prior observations in rice, kiwifruit, and Arabidopsis[24,47,48]. All these isoforms are involved in the effective elimination of the ROS and signaling integration for plant adaptations. The protein's secondary structure primarily consisted of α-helices and random coils, aligning with the results of Wang[49]. It is assumed that the random coil can primarily be involved in forming the enzyme's active site and specific functional sites for interactions with other proteins[50]. The presence of such characteristics of the genes as secondary structure is a vital aspect of the functionality and control of nucleic acids[51]. The 3D arrangement of amino acids within the random coils contributes to the structural and functional versatility of the proteins[52,53]. In this study, secondary structure analysis (Supplementary Fig. S1 and Supplemental Table S2) and DNA modelling (Fig. 1; Supplementary Table S3) predicted most MiAPX genes as monomers. These findings align with previous studies suggesting that APX typically functions as a monomer[54]. The observed variability emphasizes the importance of validating structural predictions to ensure their accuracy[55]. The MiAPX gene family exhibits structural conservation and potential functional divergence, with all members classified in the plant peroxidase-like superfamily, indicating a role in ROS scavenging. This study aligns with Pitaya, which has the same conserved plant_peroxidase_like superfamily domain[56]. Gene structure analysis revealed overall conservation in the MiAPX family, with most genes containing two introns, while MiAPX8 shows a distinct exon–intron arrangement, indicating possible evolutionary adaptation and roles in organelle-specific targeting[22]. These results give information on the functional and evolutionary dynamics of the MiAPX gene family, which can be used in future studies on stress tolerance and plant development.
Duplication of genes is a usual phenomenon in plant genomes and has played a great role in plant evolution[57]. This evolution leads to the formation of gene clusters that carry similar proteins with similar sequences and structures. Such duplications of genes may be done by whole-genome duplication (WGD), segmental duplications, or tandem duplications[58]. Intraspecific collinearity analysis (Fig. 3b) identified 26 collinear pairs of genes in mango, indicating that events of gene duplication, including WGD, segmental, and tandem duplication, have played a role in the expansion of gene families in the mango genome. These duplications likely promoted functional diversification and genetic redundancy, which are essential for evolutionary processes. They enable genes to acquire new or specialized functions, enhancing the plant's adaptability and stress tolerance. The conservation in these collinear pairs of genes demonstrates that they are important biological processes and can be utilized to gain crucial information about the evolution of the mango genome.
To further examine the evolutionary connections of APX gene family members among the species, a comparative collinearity was done between Arabidopsis and mango. The analysis found 11 Arabidopsis collinear pairs of genes (Fig. 3c), which aligns with their genetic distance[59,60]. However, MiAPX6, MiAPX8, and MiAPX9 did not show collinearity with Arabidopsis, suggesting that these genes may be unique to mango. While the other members of the MiAPX showed collinearity with Arabidopsis genes. These findings revealed that most MiAPX genes originated through segmental duplication (Supplementary Table S4), consistent with reports for APX genes in peanut[25]. They also suggest that the duplicated MiAPX genes may contribute to the evolutionary diversification of the gene family. Phylogenetic analysis revealed 26 APX genes from mango rice, and A. thaliana revealed four groups (Fig. 4). Close evolutionary relationships, such as MiAPX5-AtAPX3 and MiAPX10-AtAPX10, suggest functional similarities and provide a foundation for further studies. Transcriptional regulation of complex gene networks requires the involvement of cis-acting regulatory factors that act as molecular switches. These factors are linked to control of various biological functions, such as environmental stress response, hormonal regulation, and developmental control[61]. Cis-regulatory elements observed in the promoter of MiAPX genes were found to be involved in various biological processes (abiotic stress tolerance (light, low temperature, drought, defences and anaerobic responses), phytohormone responses (ABA, auxin, SA, JA and GA), and developmental response (e.g., circadian regulation and zein metabolism) (Fig. 6). Abiotic stressors, including changes in temperature, toxic and drought, have a serious negative effect on the productivity and growth of plants. Nevertheless, phytohormones are the regulators that aid in alleviating the negative effects of these stresses[62].
APX genes have been known to have different expression patterns during stress conditions, and this has a great role to play in plant tolerance mechanisms. qRT-PCR analysis was employed in this study to determine the expression pattern of MiAPX genes when exposed to UV-B (Fig. 6b). The results revealed that MiAPX1, MiAPX3, MiAPX4, MiAPX6, MiAPX7, and MiAPX9 were significantly up-regulated under UV-B treatment after 91 d in comparison to the control group. These findings suggest that MiAPX genes play a critical role in ROS scavenging and fruit development during mango maturation under stress conditions. Previous studies support this observation, demonstrating that transgenic Arabidopsis, sweet potato, and cotton overexpressing APX genes exhibit enhanced ROS detoxification under abiotic stress[63−65]. Similarly, increased APX expression in kiwifruit leaves and roots regulates AsA content[66]. UV-B exposure has also elevated APX gene expression in mango, lettuce, and cucumber, improving antioxidant capacity[65−67]. These observations collectively support the hypothesis that APX genes are critical in maintaining oxidative homeostasis under various abiotic stresses. The upregulation of these genes after stress exposure likely enhances the plant's ability to scavenge ROS, boosting its tolerance to adverse environmental conditions. In mango, the process of fruit development and maturation is characterised by a rapid physiological and biochemical process and a high level of oxidative metabolism. These stages of APX gene differentiation probably help to sustain the homeostasis of ROS and regulate major signaling pathways associated with ethylene production, cell wall remodeling, and pigment buildup. Moreover, latex synthesis, as a specific activity of the Anacardiaceae family, is associated with oxidation processes in the resin canals that can also rely on a body of APX activity to regulate the defensive responses involving ROS. Collectively, these observations suggest strong evidence of the retained and multifunctional nature of APX genes in the various plant species, especially in integrating stress responses and development in order to maintain plant growth during adverse environmental conditions. Future research could focus on validating the functional roles of MiAPX genes using overexpression techniques or the CRISPR/Cas system. Such studies would provide a solid foundation for further exploring stress tolerance mechanisms and could contribute to breeding stress-resilient crops like mango and other economically significant plants.
-
This study provides a comprehensive analysis of the MiAPX gene family in mango, revealing their structural diversity, evolutionary conservation, and functional roles in stress responses. Phylogenetic analysis clustered MiAPX genes with their counterparts in A. thaliana, highlighting evolutionary conservation and close functional relationships. Cis-regulatory element analysis further supports their role in abiotic stress responses and phytohormone signalling. The expression profiling of MiAPX genes under UV-B stress demonstrated differential regulation, with MiAPX1, MiAPX3, MiAPX4, MiAPX7, and MiAPX9 showing significant upregulation, suggesting their involvement in scavenging ROS and enhancing stress tolerance. In contrast, the downregulation of MiAPX8 suggests potential functional specialization. These findings align with prior studies in other plant species, confirming the conserved role of APX genes in mitigating oxidative stress and maintaining redox homeostasis. These results provide a valuable framework for future functional validation of MiAPX genes, which may lead to the development of stress-tolerant mango cultivars and other crop species.
This work was financially supported by the National Natural Science Foundation of China (Grant No. 32160677) and the Hainan University Mango Research System.
-
The authors confirm contributions to the paper as follows: study conception and design: Tahir H, Zhou K; data collection: Tahir H, Gao Y; analysis and interpretation of results: Tahir H, Gao Y, Sajjad M, Wei L, Qian M, Zeeshan Ul Haq, Tahir A, Farooq MA, Zhou K; draft manuscript preparation: Tahir H; supervision: Zhou K; project administration: Zhou K; funding acquisition: Zhou K. All authors reviewed the results and approved the final version of the manuscript.
-
The sequencing data used in this study are publicly accessible through the NCBI Sequence Read Archive under the Accession No. PRJNA1245353.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0025-0032)
-
Received 29 July 2025; Accepted 31 October 2025; Published online 22 December 2025
-
Ten MiAPX genes identified in mango with diverse structures and phylogenetic relationships.
MiAPX1, MiAPX3, MiAPX4, MiAPX7, and MiAPX9 significantly upregulated under UVB stress.
Cis-regulatory element analysis revealed MiAPX gene involvement in stress responses.
-
# Authors contributed equally: Hassam Tahir, Yijia Gao
- Supplementary Table S1 Lists the primers used for quantitative qRT-PCR.
- Supplementary Table S2 Secondary structure modeling of 10 MiAPX proteins using SOPMA server.
- Supplementary Table S3 3D structure modeling of 10 MiAPX proteins.
- Supplementary Table S4 Ka, Ks and Ka/Ks substitution rates of MiAPXs gene pairs.
- Supplementary Fig. S1 The SOPMA web server was used to study the secondary structure of 10 MiAPX genes.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Tahir H, Gao Y, Sajjad M, Qian M, Wei L, et al. 2025. Genome-wide identification and expression analysis of the ascorbate peroxidase gene family in mango under enhanced UV-B radiation. Tropical Plants 4: e039 doi: 10.48130/tp-0025-0032 

Genome-wide identification and expression analysis of the ascorbate peroxidase gene family in mango under enhanced UV-B radiation
- Received: 29 July 2025
- Revised: 23 October 2025
- Accepted: 31 October 2025
- Published online: 22 December 2025
Abstract: Reactive oxygen species (ROS) are signal transducers that lead to oxidative damage to plants during abiotic and biotic stress. Ascorbate peroxidase (APX) is an important enzyme in the ascorbate-glutathione cycle that regulates redox and improves tolerance to stress. We reported by this research group that APX gene activity, which can induce the activity of ascorbate levels in mango fruit, was increased in enhanced UV-B radiation stress. The APX gene family in mango (Mangifera indica) was comprehensively analyzed, demonstrating the presence of ten APX family members (MiAPX1-MiAPX10) with varying structural features, conserved motifs, and phylogenetic features. Gene duplication and collinearity analyses revealed evolutionary events, highlighting segmental duplications as a driver of MiAPX gene expansion. Cis-regulatory element analysis further indicated their involvement in responses to abiotic stresses, phytohormones, and developmental processes. Expression analysis under UV-B stress showed significant upregulation of MiAPX1, MiAPX3, MiAPX4, MiAPX7, and MiAPX9, implicating their role in ROS scavenging and stress adaptation. These results are in alignment with the previous work in other plant species, indicating the evolutionary conservation of APX genes in oxidative stress defense. This work can be helpful in understanding the structural diversity of the MiAPX gene family, which is a prerequisite to further research on stress-tolerance and the development of stress-resistant mango cultivars.
-
Key words:
- Mangifera indica /
- APX gene family /
- Stress resilience /
- Hormone signalling /
- Antioxidant defense