






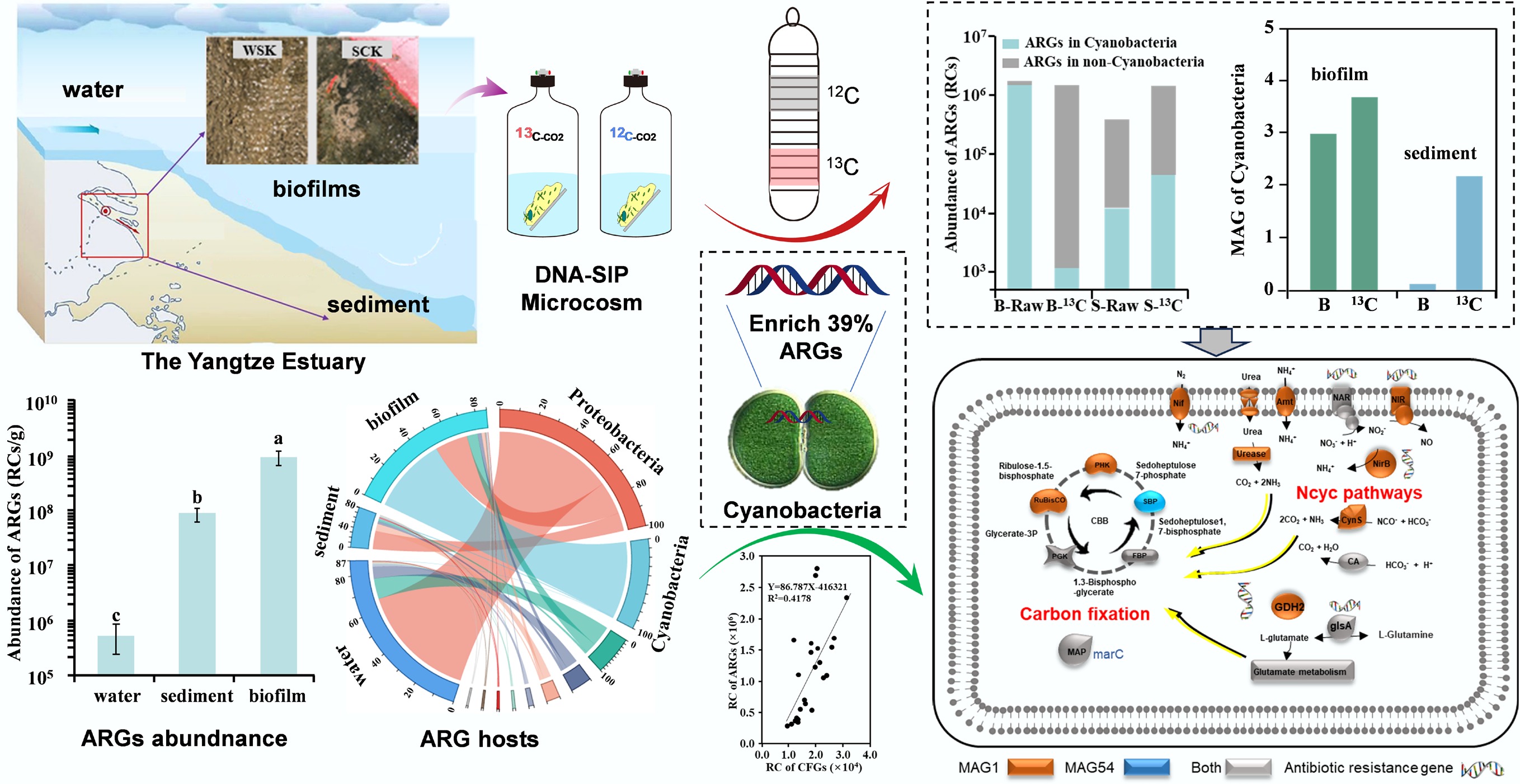



-
Antibiotics, which significantly contribute to human health and livestock production, also negatively affect the environment due to their abuse and indiscriminate discharge[1]. One of the major concerns regarding residual antibiotics is the spread of antibiotic resistance genes (ARGs), an emerging environmental pollutant that seriously affects global public health and ecological environmental security[2]. ARGs can help host bacteria withstand antibiotic treatment, thereby posing risks to humans and livestock[3]. In the environments, the host bacteria dominate the transcription, expression, proliferation, and dissemination of ARGs, and other hosts, such as viruses, can transfer ARGs through transduction[1]. These hosts affect the proliferation and dissemination of ARGs through vertical gene transfer, which involves inheriting genetic information, or horizontal gene transfer via conjugation, transduction, or transformation to acquire foreign ARGs[4]. Consequently, both biological and abiotic factors can regulate ARGs by influencing their hosts under complex environmental conditions[5,6]. Among abiotic factors, nutrient elements like C, N, P, and S are crucial for microbial growth and reproduction, and various forms of these nutrients, such as HCO3− NO3−, PO43−, SO42−, are associated with the presence and abundance of ARGs in environments[7,8]. However, the impact of the geochemical cycling processes of elements (such as carbon and nitrogen), particularly through metabolic pathways or functional genes linking microorganisms to target chemicals, on ARGs remains largely unknown.
Carbon-nitrogen coupled metabolism generally occurs within microbial cells, forming the foundation for maintaining the structure of living organisms and executing biochemical processes driven by various microorganisms[9,10]. Several functional nitrogen metabolism genes, such as norB, napA, and gdhA, are significantly correlated with ARGs[11], and dissimilatory nitrate reduction profoundly influenced the fate of ARGs during wastewater treatment processes[11]. Moreover, microbial communities involved in the nitrogen cycle, such as the ammonium-oxidizing Nitrosomonas and nitrite-oxidizing Nitrospira, may act as hosts for ARGs[12]. The multidrug-resistant plasmid RP4 increases NO and N2O yields via the electron transport system during bacterial ammonia oxidation[3]. Regarding carbon metabolism and ARGs, Cyanobacteria can affect carbon fixation types and ARGs, respectively. For example, CO2 fixation mediated by Cyanobacteria is important for the organism and global carbon balance[13], and Cyanobacterial blooms are a crucial driver of ARG diffusion and enrichment in aquatic environments[14]. Moreover, the formation of plastispheres, including Cyanobacterial communities, can alter the carbon cycle and ARGs in coastal areas[15]. A recent study has explored the relationship between carbon fixation and ARGs using the network analysis[16]. However, the specific metabolic pathways and their roles in promoting or inhibiting ARGs were not investigated or confirmed. Taken together, carbon-nitrogen metabolism might influence the fate of ARGs[17], and more attention should be paid to the effects of carbon-nitrogen cycling processes and to the microorganisms that execute them on ARGs.
Estuaries and coastal areas are crucial transitional zones where the ocean meets rivers or land, and serve as critical sites for carbon-nitrogen metabolism[18]. Salinity gradients or dissolved oxygen levels, mediated by tidal fluctuations, regulate the microbial communities involved in carbon fixation and nitrogen nitrification[19,20]. Additionally, significant amounts of anthropogenic pollutants, including nutrients, antibiotics, and heavy metals, are introduced into these areas through river runoff and sewage outfalls[7,13,20]. These pollutants can act as electron donors or acceptors in carbon-nitrogen metabolic processes or serve as nutrients for carbon-nitrogen sequestrating microorganisms. Consequently, the interplay between natural physicochemical conditions and human impacts shapes the complex carbon-nitrogen cycle. In addition, these areas are important for the occurrence and distribution of ARGs[7,13]. Metagenomic sequencing technology provides extensive information on ARGs and carbon-nitrogen metabolism, facilitating the exploration of their relationships[15,16]. Isotope tracing and DNA-based stable isotope probing (DNA-SIP) can localise elemental metabolic processes, functional genes, and associated microbes[20], helping investigate the connections between ARGs and carbon-nitrogen metabolism more precisely.
This study employed metagenomic sequencing and microcosm experiments utilizing DNA-SIP technology to comprehensively elucidate the relationships between carbon-nitrogen metabolic processes and functional genes, as well as antibiotic resistance genes (ARGs), in biofilm, sediment, and water samples from the Yangtze Estuary. The specific objectives of this study were: (1) to detect the ARGs profiles and their dominant hosts in these samples; (2) to elucidate the distribution characteristics of carbon-nitrogen metabolism functional genes and their relationship with ARGs; and (3) to identify and verify the key carbon-nitrogen metabolic processes and elucidate their potential impact on ARGs. This study offers new insights into how carbon-nitrogen metabolic processes and their executors regulate ARGs in estuarine and coastal marine environments.
-
The biofilm, sediment, and surface water samples were collected from eight sampling sites along the Yangtze Estuary in April 2023 (Fig. 1a). The detailed information is provided in the Supplementary File (section Sample collection). The physicochemical properties, including salinity, dissolved oxygen (DO), pH, dissolved organic carbon (DOC), total nitrogen (TN), total organic carbon (TOC), inorganic nitrogen ( NO3−, NO2−, and NH4+), and chlorophyll a, were described in our recent publication[19]. The properties of the samples are listed in Supplementary Table S1.
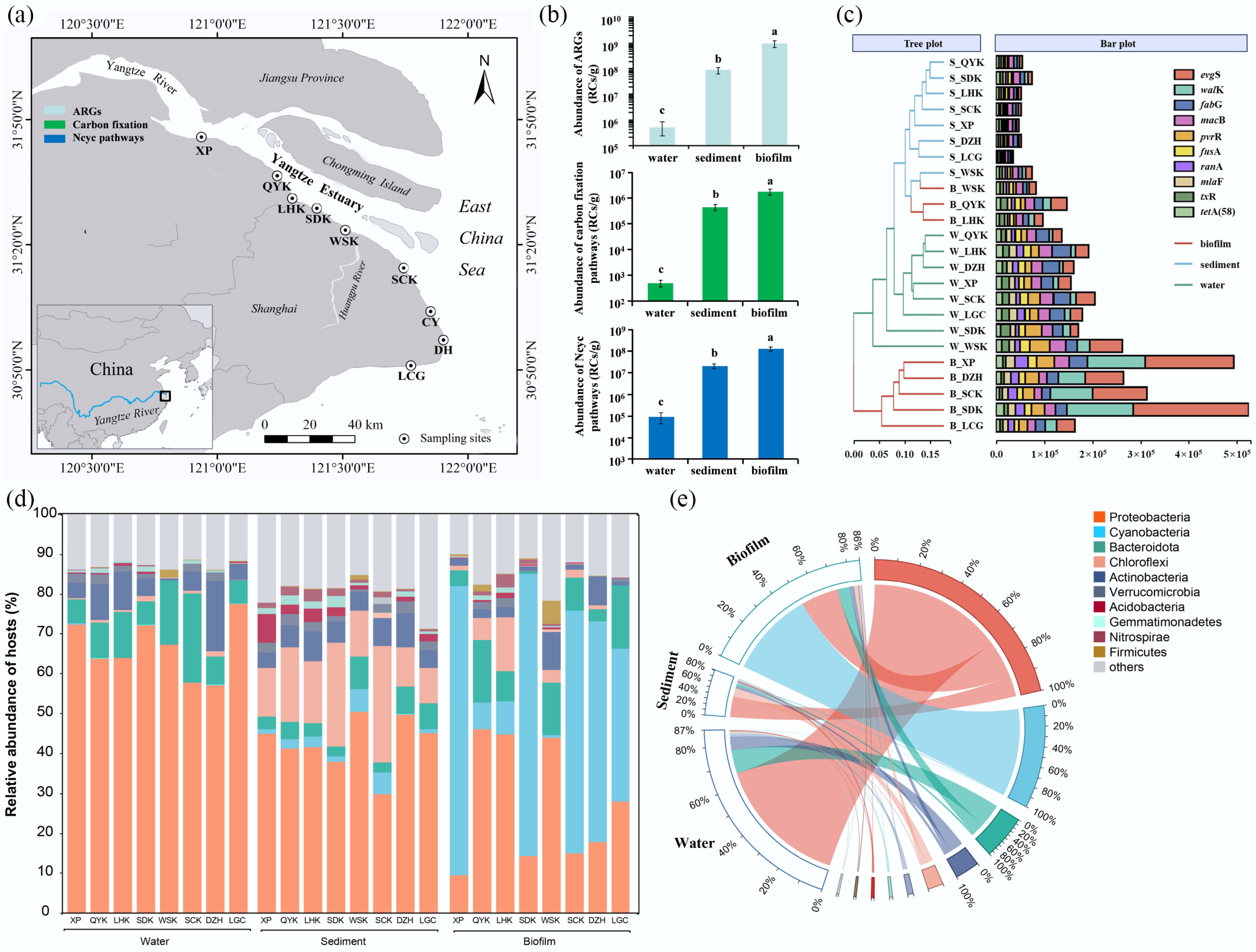
Figure 1.
(a) The eight sampling sites along the Yangtze Estuary, (b) the average abundances of ARGs, carbon fixation, and nitrogen cycling genes in the biofilms, sediments and water samples, (c) the abundance of individual ARGs in biofilm, sediment and water samples in the eight sampling sites, and their cluster analysis, (d) the relative abundance of ARGs hosts showed by sampling site, and (e) medium.
DNA-SIP microcosms
-
The DNA-SIP microcosms were set up as described in a previous study[20]. Briefly, after homogenization by mechanical stirring (100 rpm), 12.0 g of fresh biofilms or sediments and 20 mL of sterilized water sample were added to 120 mL serum bottles. The treatment groups were designed as follows: (1) blank, (2) 15NH4Cl + 5% (v/v) 13CO2. The concentration of 15NH4Cl was 20 μg/g for enriching and cultivating microorganisms related to carbon-nitrogen metabolism. All microcosms were incubated at 25 °C in the dark for 15 d. The samples were collected for DNA extraction and subsequent metagenomic sequencing. Each treatment was replicated three times.
DNA extraction and ultracentrifugation
-
Total DNA from the natural biofilm, sediment, and water samples, and from the cultured biofilms and sediments in the microcosms, was extracted using the Soil DNA Extraction Mini Kit (ONREW, Foshan, China) according to the manufacturer's protocols. The concentration and purity of DNA were determined by a UV-VIS spectrophotometer (SMA4000, Merinton, Beijing, China). Detailed information on the ultracentrifugation of DNA is provided in the Supplementary File (section Ultracentrifugation of DNA). All the DNA samples were stored at −20 °C prior to metagenomic sequencing.
Metagenomic sequencing and bioinformatic analyses
-
A total of 1 μg of extracted DNA from each water, sediment, and biofilm sample was used to construct a shotgun library using the TruSeq Nano DNA LT Sample Preparation Kit (Illumina, San Diego, CA, USA). The DNA fractions were sequenced using an Illumina Genome Analyzer (Novaseq PE150, Illumina Inc., San Diego, CA, USA) at Personal Biotechnology Co., Ltd, Shanghai, China. The analysis methods, including Fastp software[21], De Bruijn-graph-based software[22], De Bruijn graph[23], MMSeqs2 software, and several other databases, are listed in the Supplementary File (section Bioinformatic analyses). Gene abundances were derived by mapping all reads[24], and read counts (RCs) were normalized by per gram. Metagenomic data from estuarine sediments in China and the Nile River Estuary were collected according to previous studies[25,26], and the raw data were downloaded from the NCBI database. The metagenomic data for the sediment and water samples were obtained from the East China Sea. The detailed information on metagenome-assembled genomes (MAGs) and Nucleotide Identity (ANI) values is listed in the Supplementary File (section Bioinformatic analyses).
Statistical analysis
-
All statistical analyses were performed using the SPSS 19.0 software (SPSS, Chicago, IL, USA). Comparisons of gene abundance in water, sediment, and biofilm samples were evaluated using a one-way analysis of variance (ANOVA) followed by a least significant difference (LSD) test. A significant difference was considered at p < 0.05. The relationships between ARGs and environmental factors were examined using a Spearman's correlation analysis. The vegan package in R software was used to perform the cluster analysis and redundancy discriminant analysis (RDA) and partial redundancy analysis (pRDA). The association network analysis of ARGs and functional genes, and the chord diagram of the ARG hosts, were performed using the genes-cloud tools, a free online platform for data analysis (
www.genescloud.cn ). -
In total, 342 ARG subtypes were annotated, with average ARG counts of 271 ± 9, 275 ± 5, and 285 ± 4 in the biofilm, sediment, and water samples, respectively (Supplementary Fig. S1a). No significant difference in ARG subtype was observed among the sample types (p > 0.05; Supplementary Fig. S1a). However, the average abundance of ARGs in the biofilms was 9.0 × 108 RCs/g, which was significantly higher than that in sediment and water, with abundances of 8.4 × 107 and 5.1 × 105 RCs/g, respectively (p > 0.05; Fig. 1b). This finding reaffirms that biofilms serve as a critical sink for ARGs in estuarine environments, more than water or sediment, as observed in prior study[7]. Importantly, data further indicate that this biofilm-mediated enrichment is particularly pronounced in the hypertidal zone of this estuary, suggesting that hydrodynamic extremes may intensify ARG sequestration in biofilm matrices. In terms of the spatial distribution of ARGs, no evident spatial variation in ARGs with respect to salinity changes or land-based river discharge was observed among the biofilm, sediment, and water samples (Supplementary Fig. S1c). In Fig. 1c, ARGs in the biofilms at WSK, QYK, and LHK cluster with ARGs in the sediments, implying that the composition and abundance of ARGs in these three biofilms are similar to those in the sediments. A higher abundance of single ARGs was observed in the biofilms (Fig. 1c). In biofilms from XP, SDK, SCK, DZH, and LCG, these ARGs formed a cluster and subsequently grouped with ARGs in the eight water samples. This result indicated that ARGs in biofilms may mainly originate from water, due to increased exchange of substances or biological material between biofilms and water as tides rise and fall[27].
For individual ARGs, dominant ARGs, such as evgS, macB, fabG, and tetA(58) (Fig. 1c), aligned with those reported in earlier aquatic surveys[28]. However, the co-dominance of fabG, a gene less highlighted in prior estuarine studies, may highlight a distinctive selection pressure in this system, potentially linked to local contaminant profiles or microbial community structures. In biofilms, evgS dominated (9.8%), which was 2.5 and 3.5 times higher than that in water and sediment samples, respectively (Fig. 1c). As a multi-antibiotic resistance gene, evgS is a sensor protein of a two-component signal transduction system (TCSs) that phosphorylates the regulatory protein evgA, which confers multi-antibiotic resistance to fluoroquinolones, macrolides, penam, and tetracycline antibiotics[29]. Another notable feature in biofilms was the abundance of walK, and the distribution was consistent with evgS (Fig. 1c). Specifically, walK was significantly more abundant in biofilms (6.3%) than those in sediments (1.9%) and water samples (1.7%), especially for those at XP (8.8%), SDK (10.9%), SCK (10.8%), DZH (6.9%), and LCG (4.5%) (Fig. 1c). Moreover, walK is a histidine kinase sensor in TCSs that controls peptidoglycan metabolism, and confers resistance to daptomycin[30]. The synonymous genes of walK are vicK and yycG, which are sensitive to temperature changes and are beneficial for biofilm formation[31]. Additionally, pvrR is a response regulator of a TCS that regulates multi-antibiotic resistance and biofilm formation through porin deletion/gene absence[32]. The abundance of pvrR was relatively high in biofilms from XP (2.7%), SDK (2.1%), SCK (2.9%), DZH (3.5%), and LCG (3.3%), which closely resembled the distributions of evgS and walK. In general, TCSs are responsible for modulating genes involved in a multitude of bacterial functions, including biofilm formation, antibiotic resistance, and cell motility[29]. Therefore, the three most abundant genes, including evgS, walK, and pvrR, may represent common hosts with TCS-mediated resistance mechanisms in biofilms.
Furthermore, Chl-a concentration, nitrogen nutrient content, and TOC/DOC, rather than pH, DO, and salinity, were the major physiochemical factors regulating ARGs, with Pr values < 0.05 (Supplementary Fig. S2). The concentration of Chl-a was significantly correlated with the abundance of ARGs (p < 0.05). Chl-a may be primarily provided by Cyanobacteria or other microalgae that are visible in the biofilms in Supplementary Fig. S3, and previous studies have found that Cyanobacteria may be potential hosts for ARGs or may regulate ARGs in the environment[14,33]. Therefore, Cyanobacteria or other microalgae providing Chl-a may be crucial for regulating ARGs in estuarine biofilms. A previous study has reported that NH4+ and NO3− were positively correlated with the abundance of ARGs in the sediments of the East China Sea[34]. In this study, the significant correlations between ARGs and nitrogen nutrients, including TN, NO3−, NO2−, and NH4+, indicate that nitrogen cycling may be involved in the occurrence and regulation of ARGs. TOC is usually significantly associated with ARGs, suggesting that TOC or DOC can adsorb ARGs and protect them from nuclease degradation or physical separation[7]. Additionally, organic carbon can serve as a carbon source for microbial metabolism, which may play a role in regulating ARGs[35]. Thus, it can be hypothesized that metabolic processes or nutrient cycle processes, such as the nitrogen and carbon cycles, and Cyanobacteria, are likely involved in the regulation of ARGs.
In the biofilms, Cyanobacteria were highly enriched in XP, SDK, SCK, DZH, and LCG, with an average relative abundance reaching 60% (Fig. 1d, e). In addition, the clustering of ARG hosts was consistent with that of the ARGs (Supplementary Fig. S4). In sediment and water, the most abundant ARG hosts were Proteobacteria, with relative abundances of 66.4% and 42.5%, respectively (Fig. 1d). Consistent with recent reports[4,6], Proteobacteria dominated the ARG-host phyla in the samples. Beyond confirming this pattern, network analysis revealed that Proteobacteria in biofilms formed more robust co-occurrence associations with diverse ARGs compared to those in sediment, underscoring the biofilm environment as a potential hotspot for horizontal gene transfer within this phylum. As mentioned above, ARGs in biofilms at WSK, QYK, and LHK clustered with the ARGs in sediments (Fig. 1c), and the dominant ARG host was Proteobacteria in biofilms at WSK, QYK, and LHK, with an average relative abundance of 44.8% (Fig. 1d, e), which was comparable to that in sediment.
Cyanobacteria are known to serve as both the reservoirs and sources of ARGs[14,36]. In biofilms, several ARGs are positively correlated with Cyanobacteria, as shown by correlation and network analyses[37,38]. However, owing to technological limitations, the entire ARG profiles of the Cyanobacteria community have not been fully presented[36]. In this study, Cyanobacteria were not only the primary hosts of total ARGs, but also the primary hosts of the individual ARGs, including walK and evgS, with a relative abundance of nearly 80% in biofilms (Supplementary Fig. S5). In addition, as the third most abundant ARGs in biofilms, the primary host of macB is also Cyanobacteria (Supplementary Fig. S5). Therefore, in estuarine biofilms, Cyanobacteria may serve as a reservoir for ARGs, and the hosted ARGs, including walK and evgS, may serve as indicators of ARG pollution by Cyanobacteria. Among the Cyanobacteria, at the genus level, 141 host genera were predicted, and more than 50%, 44%, and 43% of the ARGs were hosted in these genera in water, sediment, and biofilm, respectively, whereas others were hosted in an unclassified Cyanobacterium (Supplementary Table S2). Harmful cyanobacterial algal blooms are an ecological and global public health concern, as they can produce cyanotoxins and disrupt ecosystems[36]. In this study, only one species, Microseira wollei, belongs to Microseira sp., which is capable of carbon-nitrogen fixation, produces several congeners of saxitoxin[39], and adapts to alternating wet and dry environments[40]. Thus, due to the presence of ARGs, Cyanobacteria and Cyanobacterial harmful algal blooms should be given greater attention, as Cyanobacteria such as Microcystis aeruginosa, which contain ARGs, may spread into the surrounding environments from biofilms.
Carbon-nitrogen metabolic processes and functional genes and their effects on ARGs
-
The average abundance of carbon fixation genes in biofilms was 1.5 × 106 RCs/g, which was significantly higher than those in sediment and water, with average abundances of 4.8 × 105 and 1.4 × 102 RCs/g, respectively (p < 0.05; Fig. 1b). The average abundance of nitrogen metabolism genes in biofilms was also significantly higher than that in sediment and water (p < 0.05), with values of 8.8 × 107, 2.5 × 107, and 7.5 × 104 RCs/g, respectively (Fig. 1a). These results suggested that biofilms might have a more substantial potential for carbon-nitrogen metabolic ability than those in sediment and water samples in the Yangtze Estuary. Although biofilms exist as thin layers in the environment, they can affect the migration and transformation of nutrients or pollutants at the sediment-water interface, and further regulate their ecological functions, such as carbon cycling[41,42]. This may be related to the unique three-dimensional structure, extracellular polymeric substances, and microbial community composition of biofilms[41,42].
The relationships between ARGs and the metabolic processes of nutrients, including carbon, nitrogen, phosphorus, and sulphur, were preliminarily analyzed in Fig. 2a, b and Supplementary Fig. S6. The results showed that most of them were significantly and positively correlated with ARGs, except for the nitrogen metabolism pathways, which showed both positive and negative correlations with ARGs (Supplementary Fig. S6). Furthermore, carbon fixation genes and nitrogen metabolic genes were significantly and positively correlated with ARGs (p < 0.05; Fig. 2a, b), suggesting that the microbial carbon-nitrogen metabolic processes may regulate the occurrence of ARGs in estuarine and coastal environments[16]. Among the carbon fixation progresses, the Calvin cycle was significantly and positively correlated with ARGs (p < 0.05; Fig. 2a). Based on pRDA, the Calvin cycle was found as the second largest regulator of variation in ARGs (13.26%) (Fig. 2a). The network analysis revealed that rbcL was significantly and positively correlated with several ARGs (Fig. 2c). These results imply that the Calvin cycle may be the dominant carbon fixation process for ARGs in estuarine environments. For the nitrogen metabolic processes, nitrogen fixation was significantly and positively correlated with ARGs (p < 0.05; Fig. 2b). Furthermore, nitrogen fixation was found to be the dominant regulator of the variation in ARGs (54.07%). Networks revealed that the functional genes of nitrogen fixation, including nifH, nifD, and nifK, were significantly correlated with most selected ARGs (Fig. 2d). Taken together, nitrogen fixation was the dominant nitrogen cycling process regulating the ARGs in this study.
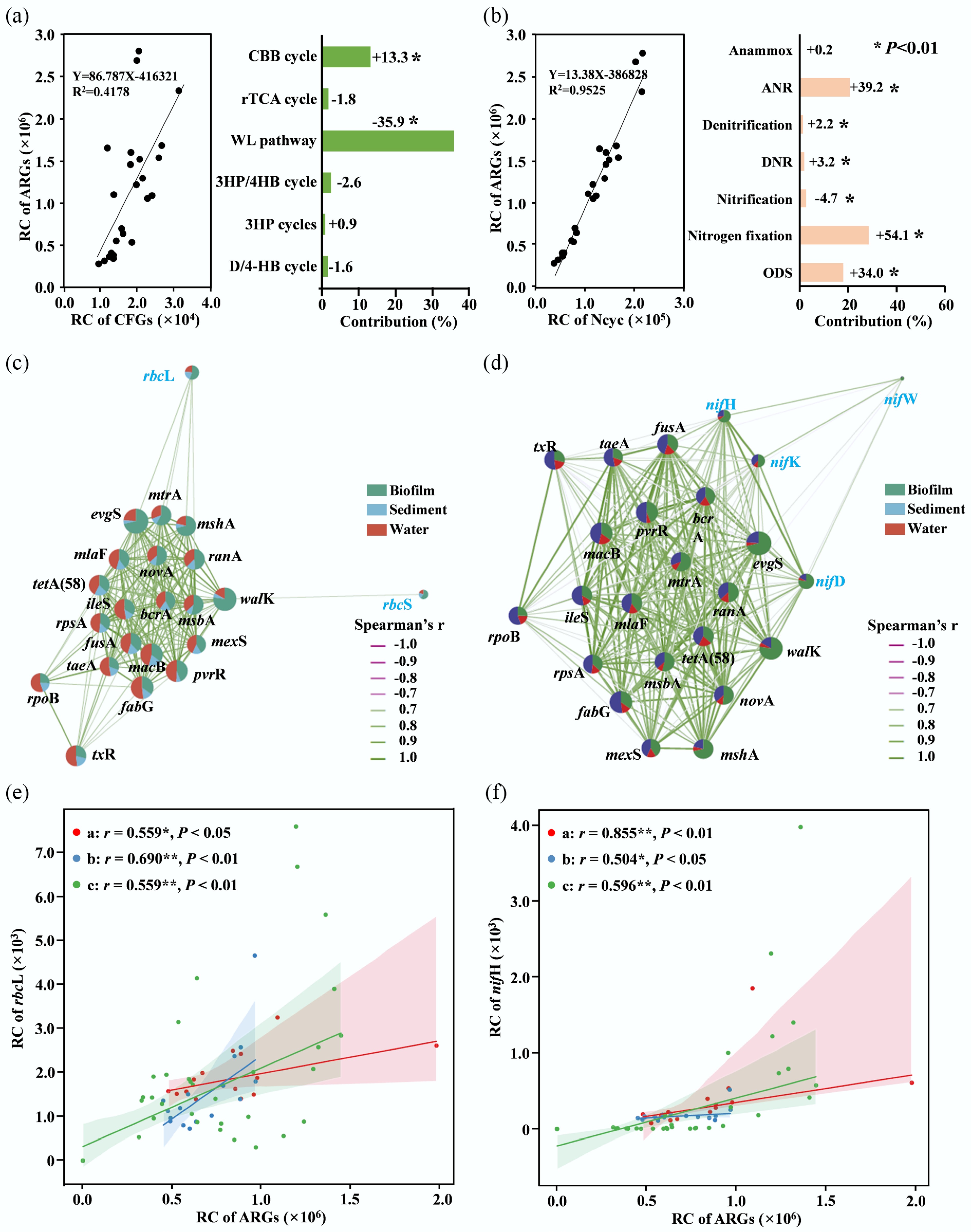
Figure 2.
Corelationship between ARGs and carbon-nitrogen fixing genes or pathways. (a) Relationship between the total abundances of ARGs and carbon-fixing genes, with contributions of different pathways to ARG variation by pRDA. (b) The relationship between the total abundances of ARGs and nitrogen metabolic genes, with contributions of different pathways to ARG variation by pRDA. (c) Network of functional genes of the Calvin cycle and the top 20 abundant individual ARGs. (d) Network of functional genes in nitrogen fixation and the top 20 abundant individual ARGs. (e) Relationship between rbcL and ARGs. (f) Relationship between nifH and ARGs. Red indicates the Nile River Estuary, blue indicates Chinese estuaries, and green indicates the East China Sea. Symbols: +, positive correlation; −, negative correlation; *, significant correlations between ARGs and functional genes (p < 0.05).
Microbial communities are key hosts for antibiotic ARGs and serve as one of the primary regulatory factors[6,43]. As described earlier, Cyanobacteria in biofilms play a central role in the Calvin cycle and nitrogen fixation, and they are also the dominant microorganisms harboring ARGs (Fig. 1d, e; Supplementary Fig. S7). Similar patterns were observed in the sediment and water samples, in which Proteobacteria were the dominant hosts for the Calvin cycle, nitrogen fixation, and ARGs (Supplementary Fig. S7). Therefore, the positive correlation among the Calvin cycle, nitrogen fixation, and ARGs may be due to their shared hosts, suggesting that the presence of the Calvin cycle and nitrogen fixation may promote the enrichment of ARGs. For instance, nitrogen-fixing microorganisms are generally categorized into heterotrophic and autotrophic groups, with Proteobacteria and Cyanobacteria, respectively, representing the dominant representatives of these groups[44]. In estuarine sediment, the contribution of heterotrophic nitrogen-fixing bacteria is actually greater[45]. However, this study demonstrated that autotrophic nitrogen fixation occurs predominantly in estuarine biofilms, potentially contributing to the spread of ARGs and posing a significant environmental risk.
To investigate the relationships between ARGs and carbon-nitrogen fixation in estuarine environments with larger geographic scales or varying degrees of human influences, the metagenomic data from 74 samples (water or sediment, Supplementary Table S3) from three regions, including the Chinese estuaries (17 sediment samples, Supplementary Fig. S8a), the East China Sea (15 water samples and 16 sediment samples, Supplementary Fig. S8b), and the Nile River Estuary (26 sediment samples, Supplementary Fig. S8c, S8d), were further analyzed. The results revealed a significant positive correlation between the abundance of rbcL, a marker of the Calvin cycle, and the total abundance of ARGs across all three regions (p < 0.05; Fig. 2e). Meanwhile, the abundance of nifH, representing nitrogen fixation, was positively correlated with the total abundance of ARGs in all three regions (p < 0.05; Fig. 2e). These findings suggest that a positive relationship between the Calvin cycle, nitrogen fixation, and ARGs may indeed exist in estuarine and coastal environments. However, as sufficient metagenomic data from biofilms are not available, it remains unclear whether these relationships hold true for biofilms. Although these correlations have been determined from the perspectives of gene abundance and microbial hosts, how changes in the complex physicochemical properties of estuaries affect their relationships remains to be studied. Moreover, despite varying levels of antibiotic contamination, positive correlations between ARGs and the Calvin cycle and nitrogen fixation have been observed in regions with severe antibiotic pollution, such as the Yangtze Estuary[7], and other Chinese estuaries[26], and in areas with less antibiotic pollution, such as the East China Sea far from the nearshore[34]. Therefore, this trend may be a natural phenomenon in estuarine environments. Nevertheless, the emission of key nutrient elements by high-intensity human activities may result in adverse environmental issues, such as eutrophication[46], in which large amounts of ARG hosts, such as Cyanobacteria, can prevail in aquatic environments[14,36]. Although the microbial Calvin cycle can fix carbon dioxide and mitigate greenhouse gas emissions, it may also facilitate the generation and transmission of ARGs.
DNA-SIP microcosms reveal the impact of carbon-nitrogen metabolism on ARGs
-
To further explore the effects of carbon and nitrogen fixation on ARGs, the CO2 labelled with the 13C isotope and NH4Cl labelled with the 15N isotope were added to the microcosms with the biofilm and sediment (Fig. 3a). Compared with those in initial samples, the total abundance of ARGs decreased by 13.6% in the biofilm and increased by 271% in the sediment (Fig. 3b). Although few changes in total ARGs in biofilms were found, the abundance of ARGs in Cyanobacteria significantly decreased by 129,600%, whereas those hosted in non-Cyanobacteria increased by 640% (Fig. 3b). In the sediments, the abundance of ARGs hosted by Cyanobacteria and non-Cyanobacteria increased by 280% and 270%, respectively (Fig. 3b). To explain these results, the relative abundance and composition of ARGs hosts were analyzed, which changed significantly in the biofilms and sediments (Fig. 3c). For example, Cyanobacteria decreased by 76,000% in biofilms (Fig. 3c), which was the primary reason for the decrease in ARGs in the biofilm microcosms, such as the above-mentioned dominant and indicative ARGs, including walK and evgS (Supplementary Fig. S9). Although the dominant host of macB was Cyanobacteria, the other major host was Proteobacteria, which had a comparable relative abundance to Cyanobacteria (Supplementary Fig. S4c), which developed into the first dominant phylum, with the relative abundance increasing by 1,100% in the13C-group compared to that in the control group (Fig. 3c). This is the reason for the increase in macB levels in the biofilms. In addition, all dominant ARGs, except for walK, all increased (Supplementary Fig. S9), which may be due to a decrease in the number of walK-positive host Cyanobacteria of walK. The DNA-SIP microcosm conditions are more conducive to nitrification research, leading to the extinction of Cyanobacteria[19]. This inversely supports the idea that there is a coupled relationship between carbon-nitrogen fixation and ARGs mediated by Cyanobacteria in biofilms.
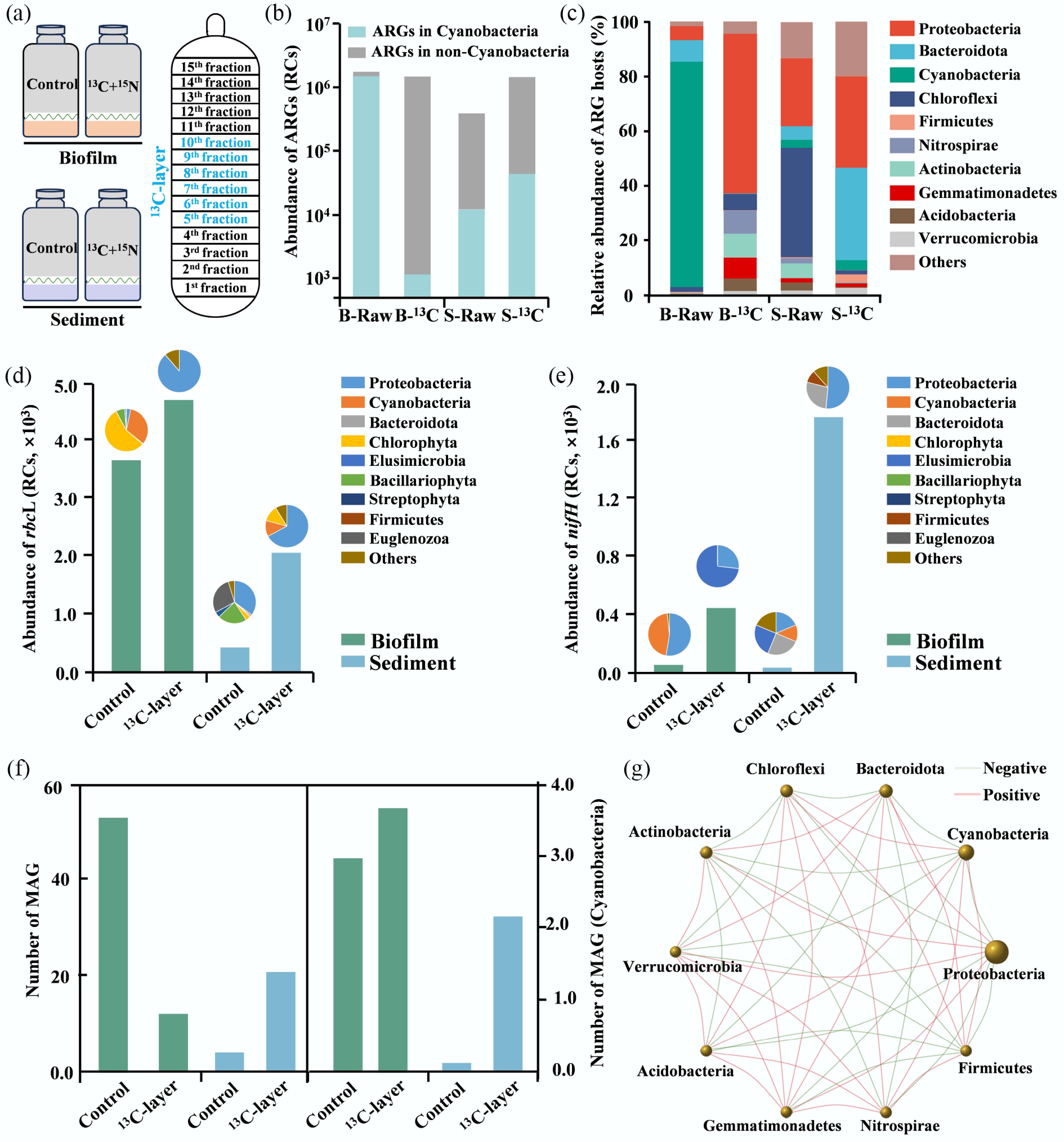
Figure 3.
The DNA-SIP microcosms of biofilm and sediment taken along the Yangtze Estuary. (a) The DNA-SIP microcosm and DNA stratification. (b) The abundance of ARGs in control and 13C-layer groups. (c) The relative abundance of ARGs hosts in control and 13C-layer groups. (d) The abundance of rbcL and its hosts in control and 13C-layer groups. (e) The abundance of nifH and its hosts in control and 13C-layer groups. (f) The abundance of MAGs and the MAGs of Cyanobacteria in control and 13C-layer groups. (g) The network of ARGs hosts in the biofilm, sediment, and water samples.
In terms of carbon-nitrogen fixation, the abundance of rbcL increased by 30% and 250% in the 13C-labeled layers of biofilms and sediments, respectively (Fig. 3d), while the abundance of nifH increased by 700% and 5,430% in 13C-layer of biofilms and sediments, respectively (Fig. 3e). Interestingly, the changes in functional genes and ARGs in biofilms were inconsistent, which differed from the above result that functional genes showed a significant correlation with ARGs (Fig. 2e, f). Further analysis revealed that the abundance of rbcL and nifH genes hosted by Cyanobacteria decreased, indicating that carbon-nitrogen fixation mediated by Cyanobacteria decreased, whereas that mediated by non-Cyanobacteria increased. For example, in the 13C-labeled rbcL hosts, Betaproteobacteria and Gammaproteobacteria accounted for 84.5% of the total identified microbial communities in the biofilms (Supplementary Fig. S10), and these microbes were also dominant in the sediments with a relative abundance of 65.7% (Supplementary Fig. S10). These findings support a positive correlation between ARGs, the Calvin cycle, and nitrogen fixation, specifically mediated by Cyanobacteria.
To investigate the relationship between ARGs and carbon-nitrogen metabolism in the microbial species, MAGs from the initial and 13C-labeled groups were further analyzed, and the results showed that 65 MAGs were predicted in Fig. 3f and Supplementary Table S4. Among them, several MAGs belonged to Cyanobacteria, which were labelled with 13C in the heavy layer and had complete genomes > 99% (Fig. 3f; Supplementary Table S4), confirming their involvement in carbon-nitrogen metabolism. Network analysis showed that, as dominant hosts of ARGs, Cyanobacteria exhibited negative correlations with Proteobacteria and Bacteroidota (Fig. 3g), indicating that competition between microbial communities may affect the occurrence of ARGs in estuarine biofilms[14]. Phylogenetic analysis and ANI revealed that the predominantly active Cyanobacteria, marked by 13C in the biofilm and sediment, belonged to Microcoleaceae sp. and Geitlerinemaceae sp. (Fig. 4a, b). Based on these findings, the metabolic mechanisms linking ARGs to nitrogen-carbon metabolism were further explored using these two MAGs in subsequent analyses.
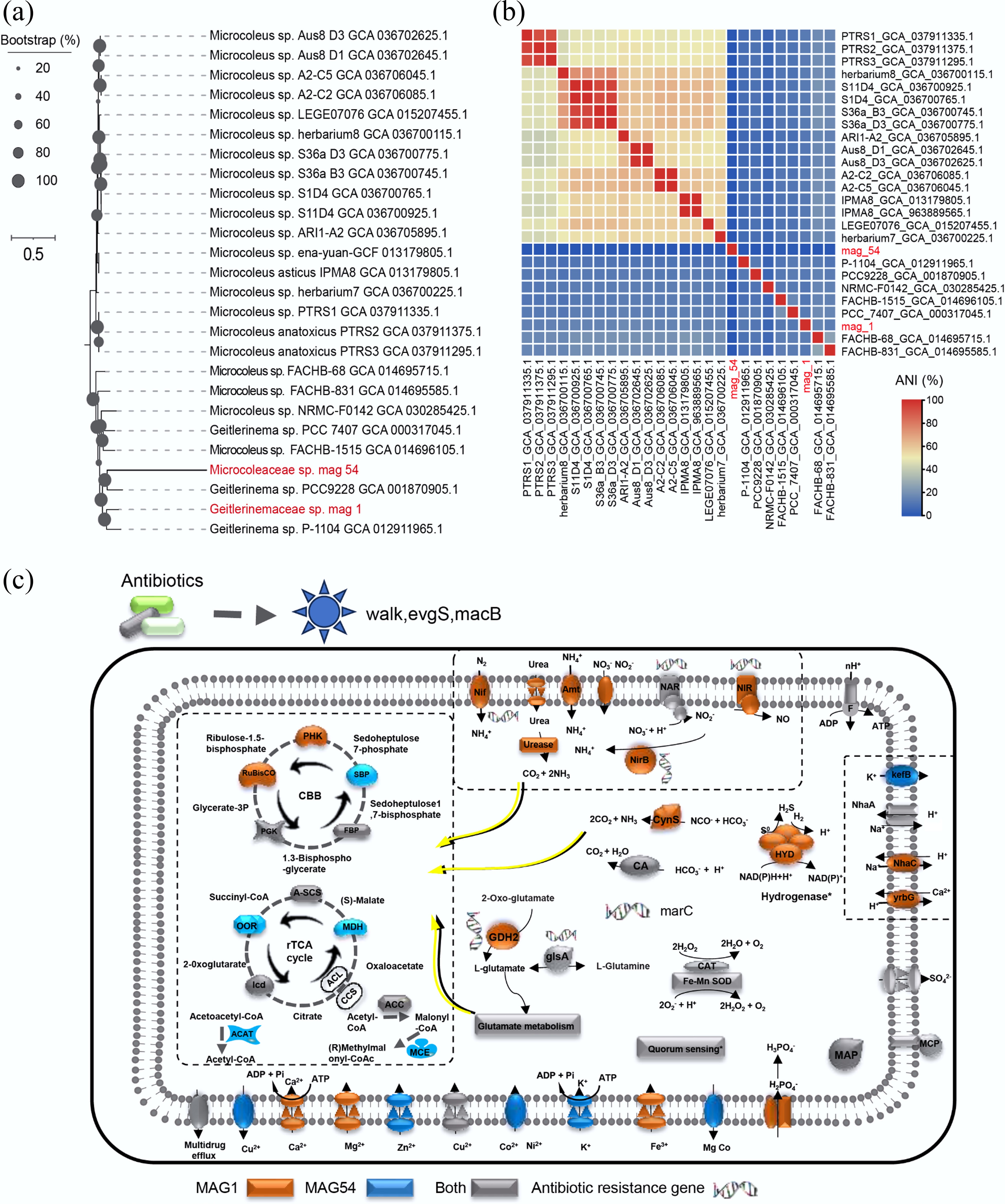
Figure 4.
(a) A phylogenomic tree was constructed to investigate the evolutionary relationships among Cyanobacterial genomes: (b) Average Nucleotide Identity (ANI) analysis was performed to assess further assess the genomic similarities and differentiation among the Cyanobacterial strains. (c) Metabolic analyses of cyanobacteria metagenome-assembled genomes (MAGs), providing insights into their survival strategies in estuarine coastal biofilms. Colors denote the presence of specific genes in MAG1 (orange), MAG54 (blue), or both (grey). Asterisks represent incomplete features. Key enzymes and pathways are abbreviated as follows: Nif, nitrogen fixation protein, amt, ammonium transporter; NAR, ferredoxin-nitrate reductase; NIR, ferredoxin-nitrite reductase; NirB, nitrite reductase (NADH) large subunit; CynS, cyanate; CA, carbonic anhydrase; HYD, 3b [NiFe] hydrogenase; CAT, catalase; kefB, glutathione-regulated potassium-efflux system protein; NhaA, Na+ : H+ antiporter; yrbG, Na+ : H+ antiporter; MAP, methionyl aminopeptidase; GDH2, glutamate dehydrogenase; glsA, glutaminase; MarC, nitrite reductase (NADH) large subunit; MCP, methyl-accepting protein; MSP, methionine salvage pathway; SOD, superoxide dismutase. PGK, phosphoglycerate kinase; RuBisCO, ribulose-bisphosphate carboxylase large chain; FBP, fructose-bisphosphate aldolase; SBP, fructose-1,6-bisphosphatase II/sedoheptulose-1,7-bisphosphatase; PRK, phosphoribulokinase; A-SCS, succinyl-CoA synthetase alpha subunit; MDH, malate dehydrogenase; ICD, isocitrate dehydrogenase; OOR, 2-oxoglutarate/2-oxoacid ferredoxin oxidoreductase subunit alpha; MCE, methylmalonyl-CoA/ethylmalonyl-CoA epimerase; ACAT, methylmalonyl-CoA/ethylmalonyl-CoA epimerase; ACC, acetyl-CoA C-acetyltransferase. Enzyme complexes of the respiratory chains are labeled using Roman numerals.
Possible metabolic mechanisms between ARGs and nitrogen-carbon metabolism
-
Several ARGs, including marC, were annotated to nitrogen transformation genes (nifH, NAR, NIR, and nirB) and carbon-related metabolic pathways, suggesting potential mechanisms through which ARGs influence Cyanobacterial metabolism (Fig. 4c; Supplementary Table S5). Nitrogen fixation is an energetically demanding process, reliant on nitrogenase activity encoded by nifH and sensitive to intracellular redox balance[47]. ARGs, such as those co-localized with nifH (Fig. 4c), may influence nitrogenase stability by mitigating oxidative stress or modulating energy availability[48]. Similar mechanisms have been observed in antibiotic-stressed bacterial communities, where ARGs enhance stress resilience[49]. NAR and NIR, which reduce nitrate and nitrite to ammonium, respectively, play crucial roles in the nitrogen cycle. ARG-associated modulation of these genes could ensure sustained ammonium availability for nitrogenase under antibiotic pressure[50]. nirB, which is involved in ammonium assimilation, further integrates nitrogen metabolism into broader metabolic networks, a process that is potentially enhanced by ARG-driven regulatory mechanisms[51]. The co-expression of ARGs with these genes in Cyanobacteria aligns with the observations in antibiotic-contaminated soils, where nitrogen metabolism is reshaped by ARG activity[52].
Carbon metabolism in Cyanobacteria involves pathways, such as formate oxidation, carboxylic acid metabolism, and urease activity, all of which provide CO2 for the Calvin cycle[53]. The detection of ARGs (such as the dominant gene walK) in GDH2 and glsA suggests that ARGs may co-occur within the same microbes possessing GDH2/glsA, thereby enhancing microbial metabolic plasticity under stressful conditions[54]. GDH2 integrates carbon and nitrogen metabolism, while glsA supports amino acid biosynthesis, which is essential for growth and stress adaptation[55]. Furthermore, ARGs may influence urease activity, which releases CO2 and ammonium, thereby supporting both carbon fixation and nitrogen assimilation. Similar roles for ARGs in reshaping metabolic pathways have been documented in microbial communities exposed to antibiotics[56].
The marC gene, associated with multidrug resistance, plays a critical role in membrane transport and stress adaptation[57]. In Cyanobacteria, marC may stabilize the CO2 transport systems, thereby supporting RuBisCO activity in the Calvin cycle. Additionally, marC likely contributes to the oxidative stress defense by protecting the nitrogenase and carbon-fixation machinery under environmental pressures[58]. The involvement of marC in metabolic regulation parallels findings in other microbial systems, where ARGs enhance resilience and metabolic efficiency[59]. The integration of ARGs into metabolic networks suggests their broader ecological roles beyond conferring antibiotic resistance. ARGs may act as hubs that coordinate stress responses to nitrogen and carbon metabolism, as seen in other stress-adapted microbes[60]. Furthermore, biofilm formation and extracellular polymeric substance production in Cyanobacteria, influenced by ARGs, optimize the micro-environment for nitrogen fixation and carbon sequestration[61].
These findings highlight the dual roles of ARGs in both antibiotic resistance and metabolic regulation. Cyanobacteria harboring ARGs could be pivotal in ecological nitrogen and carbon cycling and have potential for applications in sustainable agriculture and carbon capture technologies[62]. However, the dissemination of ARGs in natural ecosystems warrants caution, as it could exacerbate the spread of resistance traits[63]. Future research incorporating multi-omics approaches, isotope tracing, and mutant analyses is crucial for elucidating the molecular mechanisms underlying these processes.
-
This study underscores the pivotal role of Cyanobacteria in hosting ARGs within Yangtze estuarine biofilms and establishes a genetic co-occurrence association between ARGs and microbial carbon-nitrogen metabolism. Metagenomic analysis identified Cyanobacteria as the dominant hosts of ARGs and key carbon-nitrogen fixation genes, such as rbcL and nifH, suggesting an intrinsic connection between these processes. These findings revealed a positive correlation among ARGs, the Calvin cycle, and nitrogen fixation, suggesting a widespread and natural phenomenon in estuarine and coastal environments. Stable isotope labelling with 13C and 15N further confirmed that microorganisms executing the Calvin cycle and nitrogen fixation were primarily Cyanobacteria, which were also major reservoirs of ARGs. The detection of ARGs, including marC, co-occurring with nitrogen transformation genes (nifH, NAR, NIR, and nirB) and carbon metabolism genes (GDH2 and glsA), highlights the potential mechanisms by which ARGs influence Cyanobacterial metabolism. Overall, this study provided novel insights into the dual roles of ARGs in both antibiotic resistance and metabolic regulation in aquatic ecosystems. Cyanobacteria harboring ARGs may contribute to nitrogen and carbon cycling, with potential applications in sustainable agriculture and carbon capture technologies. However, their role in ARG dissemination also raises concerns, particularly regarding the eutrophication and expansion of Cyanobacterial blooms. These findings emphasize the need for continued monitoring and mitigation strategies to balance the ecological benefits of Cyanobacteria with the risks associated with ARG proliferation in estuarine and coastal systems.
-
It accompanies this paper at: https://doi.org/10.48130/ebp-0025-0021.
-
The authors confirm contributions to the paper as follows: Xing-Pan Guo: conceptualization, methodology, resources, validation, formal analysis, investigation, writing – original draft, writing – review & editing, visualization, project administration, funding acquisition; Xiu-Feng Tang: conceptualization, methodology, resources, validation, formal analysis, investigation, writing – original draft, writing – review & editing, visualization; Nazupar Sidikjan: investigation; Xiang-Yang Zhao: investigation; Long-Ling Wang: investigation; Zhi Guo: writing – review & editing; Ping Han: writing – review & editing; Ye Huang: writing – review & editing; Li-Jun Hou: project administration; Yi Yang: writing – review & editing, project administration. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available upon request from the corresponding author.
-
This study was funded by the National Natural Science Foundation of China (Grant Nos 42125102, 42576166, and 42507400), the National Key Research and Development Program of China (Grant No. 2022YFC3105800), and The China Postdoctoral Science Foundation (Grant No. GZB20250575).
-
The authors declare that they have no conflict of interest.
-
Cyanobacteria were the dominant hosts of ARGs, accounting for 39% of ARGs in biofilms.
Calvin cycle and nitrogen fixation genes were significantly correlated with ARGs.
The effects of carbon-nitrogen fixation on ARGs were universal in estuaries.
Several ARGs were associated with carbon/nitrogen-related metabolic genes and pathways.
-
# Authors contributed equally: Xing-Pan Guo, Xiu-Feng Tang
Full list of author information is available at the end of the article. - The supplementary files can be downloaded from here.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Guo XP, Tang XF, Sidikjan N, Zhao XY, Wang LL, et al. 2026. Cyanobacteria-mediated carbon-nitrogen coupling promotes the enrichment of antibiotic resistance genes in the Yangtze estuarine biofilms. Environmental and Biogeochemical Processes 2: e004 doi: 10.48130/ebp-0025-0021 

Cyanobacteria-mediated carbon-nitrogen coupling promotes the enrichment of antibiotic resistance genes in the Yangtze estuarine biofilms
- Received: 31 October 2025
- Revised: 13 December 2025
- Accepted: 26 December 2025
- Published online: 21 January 2026
Abstract: The metabolism of biogenic carbon-nitrogen elements mediated by Cyanobacteria is a fundamental process for microbial physiological and metabolic activities, playing vital roles in the proliferation and dissemination of antibiotic resistance genes (ARGs). However, the relationships between carbon-nitrogen metabolism and ARG patterns among Cyanobacteria remain understudied. Here, the study investigated the relationships between ARGs and carbon-nitrogen metabolism in naturally occurring biofilms, sediments, and water bodies in the Yangtze Estuary, via metagenomic analysis combined with DNA-based stable isotope probing (DNA-SIP). The results showed that Cyanobacteria were the dominant ARG hosts in biofilms, contributing 39% of ARGs, compared with only 0.2% and 2.5% in water and sediment samples, respectively. The Calvin cycle and nitrogen fixation genes were significantly positively correlated with ARGs, explaining 13.3% and 54.1% of the variation in the ARGs, respectively. DNA-SIP further confirmed that organisms involved in the Calvin cycle and nitrogen fixation, such as Cyanobacteria, were dominant hosts of ARGs. Several metagenome-assembled genomes, belonging to Cyanobacteria, were labelled with 13C and identified as Microcoleaceae sp. and Geitlerinemaceae sp. Certain ARGs were annotated in nitrogen transformation genes (nifH, NAR, NIR, and nirB) and carbon-related metabolic pathways, further confirming the genetic co-occurrence of ARGs and microbial carbon-nitrogen genes. This study provides new insights into the dual roles of ARGs in antibiotic resistance and metabolic regulation of Cyanobacteria in estuarine biofilms.
-
Key words:
- Carbon fixation /
- Nitrogen fixation /
- Antibiotic resistance genes /
- DNA-SIP /
- The Yangtze estuary