










-
Tea, a beverage with a long and storied history, is favored not only for its distinctive flavor but also for the variety of bioactive compounds its contains, which have garnered significant attention in scientific research[1,2]. These active components have been scientifically proven to promote sleep, accelerate ethanol metabolism, and alleviate anxiety[3−5]. Taihu Cuizhu tea, a signature specialty of Wuxi City, Jiangsu Province, enjoys a prestigious reputation in the international tea market for its exceptional quality and unique appearance. The tea is characterized by its flat and elegant shape, vibrant green and oily color, clear and bright infusion, lofty and enduring aroma, fresh and mellow taste, and tender and uniform leaf base. In evaluations of a tea's quality, appearance, aroma, taste, color infusion, and leaf base are all important considerations. Among these, aroma, as one of the key factors in distinguishing tea quality, varies significantly among different types of tea[6,7].
Fixation, the first step in tea processing, plays a crucial role in tea quality[8]. Catechins, as secondary metabolites of tea plants, are one of the unique chemical components in tea leaves and are also one of the main constituent substances, possessing the ability to scavenge free radicals[9,10]. Research indicates that when the fixation temperature is too high, catechins are prone to thermal degradation, affecting tea quality[11]. Microwave fixation involves energy conversion through dipole rotation and ionic conduction[12], converting microwave energy into heat energy and evaporating some of the moisture in the tea leaves. Concurrently, at high temperatures, proteins in the tea leaves decompose, releasing free amino acids[13].
The harvesting season is crucial for tea quality. Research by Ye et al. indicates that the 'Huangyu' mutant of the 'Yinghong 9' variety exhibits the best physicochemical quality when harvested in spring[14]. Theaflavins and thearubigins, as important phenolic substances that determine the quality of black tea, account for more than half of the phenolic compounds in black tea[15]. Tea quality is closely related to the seasonal variation in total polyphenol content[16]. Further research by Dai et al. deepened this understanding; they found that the bitter components in tea leaves harvested in summer and autumn significantly increased, a fac that is related to the seasonal changes in several flavonoid compounds and amino acids, the main factors affecting the seasonal variation in the taste quality of tea[17].
The microbial community is also an important factor affecting tea quality[18], as microorganisms can participate in the metabolism of theaflavins, caffeine, organic acids, and other products[19,20]. Research by Xiao et al. found that fermentation with Eurotium cristatum can increase the floral scent of autumn tea and reduce astringency[21]. Zhang et al. discovered that fermentation with Cordyceps militaris can improve the flavor of summer Keemun black tea[22].
This study focuses on fresh leaves of Taihu Cuizhu tea from specific plots in Doushan area of Wuxi, including special-grade and first-grade fresh leaves. The study employed various fixation processes for fresh leaves harvested in different seasons. By integrating sensory quality evaluations, biochemical indicators, and microbiomic and metabolomic analyses, the aim was to explore the optimal fixation temperature, time, and harvesting season for producing high-quality Taihu Cuizhu tea. The results of the study aim to provide scientific evidence and practical guidance to produce Taihu Cuizhu tea in the Wuxi region.
-
Fresh leaves of Taihu Cuizhu tea were collected from the same plot in the Doushan area of Wuxi, Jiangsu Province. Tea leaves were categorized into special-grade and first-grade tea leaves, adhering to the 'single bud' and 'one bud with one leaf' plucking standards, respectively. Among them, special-grade fresh leaves are those composed of single buds with a uniform length of about 1.5–2 cm, without single or broken leaves, and the fresh leaves are uniform and clean. First-grade fresh leaves refer to fresh leaf buds that are 2–2.5 cm long, composed of one bud and one newly unfolded leaf, with the bud being longer than the leaf, and no single or fragmented leaves. This standard is based on the production standard samples of Taihu Cuizhu tea in the Doushan area of Wuxi (the original habitat of Taihu Cuizhu tea)[23]. Target tea samples were collected from Wuxi Doushan Bai Cha Yuan Tea Culture Co., Ltd. in the autumn of 2023 and the spring of 2024. These samples were collected in accordance with the standard processing techniques of leaf picking, spreading, fixation, and drying. The spreading time was uniformly set at 8 h, and the blanching steps were completed relative to the control group set in the experiment. To examine the effects of thermal parameters on tea quality, three experimental fixation temperatures (160, 180, and 200 °C) were established, each with corresponding treatment durations of 6, 8, and 10 min. The fixation process was conducted using automated fixation equipment (Sichuan Dengyao Machinery Equipment Co., Ltd., 6CL-1280), which is widely used in the Doushan production area, ensuring consistent product quality through standardized operational protocols. Following the fixation process, the treated samples were dried in a tea roasting machine (Fujian Jiayou Tea Machinery Intelligent Technology Co., Ltd., 6CHZ-01). The drying temperature was uniformly set at 100 °C for 10 min. The dried samples were subjected to comprehensive biochemical analyses. The remaining tea samples were ground and passed through a 100-mesh Tyler sieve, then stored at –20 °C until further analysis.
Sensory evaluation
-
The evaluated tea products and chemical standards were both deemed to be safe. A panel consisting of eight trained evaluators was selected to form the sensory evaluation group. The participants gave their consent to take part in the sensory study and for us to use their information. In accordance with the methodology for the sensory evaluation of tea (GB/T23776-2018), the evaluation group conducted a sensory assessment of five factors: Appearance, infusion color, aroma, taste, and leaf base. Prior to the formal evaluation, additional calibration training was conducted, specifically addressing the characteristics of the tea samples used in this experiment to ensure a consistent understanding of the aroma and taste descriptors. To ensure the reliability of the evaluation results, Cronbach's alpha coefficient was used to assess inter-evaluator consistency. A value of 0.85 was obtained, indicating good agreement among the panelists and reliable data. In the appearance evaluation, 200 g of tea leaves was weighed, and then 3 g of each tea sample was added to 150 mL of boiling water and steeped for 4 min. The tea infusion was filtered out at a consistent rate, leaving the tea leaves in the teapot. The tea infusions were labeled with three-digit random codes and distributed to the evaluators for tasting, with the results being recorded. The average of the results from the eight evaluators was taken as the outcome for the sample.
Calculation of pH
-
A tea sample weighing 5 g was measured and combined with distilled water at a ratio of 1:5 (w/v) in a centrifuge tube. The mixture was allowed to undergo complete hydration through thorough soaking. Subsequently, the hydrated mixture was subjected to centrifugation at 3,500 r/min for a duration of 10 min. Following centrifugation, the supernatant was carefully decanted into a volumetric flask. The pH measurement was then conducted by immersing a calibrated pH electrode into the supernatant, and the corresponding pH value was recorded.
Analysis of chemical composition
Determination of moisture content
-
In accordance with the national food safety standard (GB 5009.3-2016), the direct drying method was employed to determine the moisture content of tea leaves. A clean aluminum pan was selected and placed in a drying oven at a temperature of 101~105 °C for 1.0 h, then cooled in a decorator for 0.5 h and weighed. The drying process was repeated until the mass difference between two consecutive weighings did not exceed 2 mg, which was considered to be the constant weight of the aluminum pan. The aluminum pan containing the sample was weighed on an analytical balance with an accuracy of 0.01 g. The sample was placed in a preheated oven at a temperature of 65–75 °C for drying. After drying, the pan was removed, cooled to room temperature in a desiccator (which takes approximately 30 min), and weighed immediately. Then the sample was placed back in the oven at 65–75 °C for about 1 h of further drying. The sample was removed, cooled in a decorator for 0.5 h, and weighed again. This procedure was repeated until the mass difference between two consecutive weighings did not exceed 2 mg, which was considered to be the constant weight.
Determination of total polyphenol content
-
According to the National Standard of the People's Republic of China (GB/T8313—2018), the determination of tea polyphenols was conducted as follows. First, we weighed 0.2 g (accurate to 0.0001 g) of a uniformly ground tea sample into a 10 mL centrifuge tube, then added 5 mL of a 70% methanol–water solution, preheated the mixture to 70 °C, and mixed it well with a glass rod until thoroughly moistened. The sample was immediately transferred to a 70 °C water bath and infused for 10 min (stirring every 5 min). After infusion, the sample was cooled to room temperature and then centrifuged at 3,500 r/min for 10 min. The supernatant was transferred to a 10-mL volumetric flask. The residue was extracted once more with 5 mL of a 70% methanol–water solution, repeating the previous steps. The extracts were combined and made up with the solvent to a volume of 10 mL, shaken well, and filtered through a 0.45-µm membrane. Next, 1.0 mL of the mother liquid was transferred to a 100-mL volumetric flask, diluted to the mark with water, and shaken well. Absorbance (E) was measured at 765 nm using a spectrophotometer (Model 752), and tea polyphenols were quantified using gallic acid as the calibration standard.
Determination of total free amino acid content
-
According to the national standard (GB/T8314-2013), the total amount of free amino acids was determined using the ninhydrin method. For this, 1 g of the sample was taken and addedto a centrifuge tube with 50 mL of distilled water, then infused in a boiling water bath at 100 °C for 45 min. The mixture was removed, cooled, and filtered, and the filtrate was reserved. The next step involved accurately pipetting 1 mL of the test solution into a 25-mL colorimetric tube, adding 0.5 mL of phosphate buffer (pH 8.0) and 0.5 mL of a 2% ninhydrin solution, and heating the mixture in a boiling water bath for 15 min. After cooling, the sample was diluted to 25 mL with water. After the mixture was allowed to stand for 10 min, the absorbance (A) was measured at 570 nm using a 5-mm cuvette, with the reagent blank solution as the reference. This operation was repeated with 1 mL of a standard theanine working solution to determine the absorbance.
Determination of total flavonoid content
-
The determination of plant flavonoid content was performed using the Plant Flavonoid Content Test Kit (Aluminum Hydride Method) from Beijing Solarbio Technology Co., Ltd., following the manufacturer's instructions. Briefly, 0.1 g of a ground tea sample was placed in a centrifuge tube, to which 1 mL of an extraction solution was added, and the sample was extracted using an ultrasonic extraction method. The mixture was then centrifuged at 12,000 rpm for 10 min, and the supernatant was collected and made up to a final volume of 1 mL. Reagents were added according to the kit's protocol and mixed thoroughly. The absorbance was measured at 470 nm. Using the standard curve of rutin (0.1–2 mg/mL), the total flavonoid content was calculated and expressed as micrograms of rutin equivalents (RE) per milliliter of tea infusion (μg RE/mL)[24].
Determination of soluble polysaccharide content
-
The determination of soluble polysaccharide content in the leaf samples was conducted using the Plant Soluble Sugar Content Test Kit (Aluminum Hydride Method) from Beijing Solarbio Technology Co., Ltd., following the manufacturer's instructions. Briefly, glucose working solutions (200, 150, 100, 50, and 25 μg/mL) were prepared as standard tubes, distilled water was used as the blank tube, and 1 mL of a tea infusion was used as the test tube. Each solution was placed in a 25 mL volumetric flask, and, after cooling, 8 mL of anthrone reagent was added. The mixture was then heated at 100 °C for 7 min and subsequently cooled to room temperature. The absorbance was recorded at 620 nm, and the soluble polysaccharide content (μg/mL) was calculated using the standard glucose curve.
16S rRNA gene sequence amplification and sequencing
-
Refer to Edwards' method for the surface sterilization steps for tea leaves and validation of the efficacy of sterilization[25]. Subsequently, the samples were treated by soaking in 75% alcohol for 1 minute, rinsed three times with sterile water, disinfected with 5% sodium hypochlorite for 5 min, and rinsed three times with sterile water for sterilization and set aside for use. They were then ground using a grinding device. We employed the CTAB (hexadecyltrimethylammonium bromide) technique to extract genomic DNA from the tea leaves and detected the quality of DNA extraction using 1% agarose gel electrophoresis. We used the primers 799 F (5′-AACMGGATTAGATACCCKG-3′) and 1,193 R (5′-ACGTCATCCCCACCTTCC-3′) to construct bacterial genomic libraries. Additionally, to achieve specific amplification of the V5–V7 region of the 16S rRNA gene for each sample, we incorporated a unique 6-nt barcode at the 5′ end of the forward primer. To ensure high fidelity of the reaction, all polymerase chain reaction (PCR) reactions were conducted using QuasisonTM High-Fidelity PCR Master Mix (New England BioLabs, Ipswich, MA, USA). The detection of PCR products was performed using a one-step loading buffer containing SYBR green dye, which was mixed with the PCR products at a volume ratio of 1:1, and then the samples were analyzed by electrophoresis on a 2% agarose gel. Purification was carried out using the Qiagen Gel Extraction Kit (Qiagen, Hilden, Germany).
Following the manufacturer's instructions, we constructed a library from the pooled PCR products using the TruSeq DNA PCR-Free Sample Preparation Kit (Illumina, San Diego, CA, USA). We assessed the quality of the constructed library using the Qubit@2.0 fluorometer (Thermo Scientific, Waltham, MA, USA) and the Agilent Bioanalyzer 2100 system. Ultimately, we performed high-throughput sequencing of the library using the Illumina MiSeq PE 300 platform, obtaining paired short-read sequences. The raw sequencing data were processed for quality control using QIIME (Version 1.9.1,
http://qiime.org ), and paired reads were processed using FLASH (Version 1.2.7,http://ccb.jhu.edu/software/FLASH )[26,27]. The sequencing depth of the samples is shown in Supplementary Table S1, and rarefaction curves are shown in Supplementary Fig. S1. The raw 16S data were deposited in the NCBI database under accession code PRJNA1321724.Determination of metabolites by liquid chromatography–mass spectrometry
-
On the basis of our analysis of the physicochemical properties, we selected four samples with the best overall performance (AS160_6, AF180_6, SF180_8, and SS160_10) for untargeted metabolomic analysis. For this, 0.4 g of freeze-dried tea powder was placed into a 10-mL centrifuge tube, and 8 mL of a 70% methanol solution containing 4-chloro-DL-phenylalanine (200 mg/L) was added as an internal standard. After ultrasonic treatment for 30 min, the mixture was left to stand for 4 h. Next, 1 mL of the supernatant was centrifuged in a 2-mL centrifuge tube (4 °C, 12,000 rpm, 10 min). The centrifuged supernatant (0.1 mL) was diluted 40-fold and placed in a vial for analysis. The quality control (QC) samples were prepared by pooling 10 μL of each sample and analyzed together with the other samples. The QC samples were inserted regularly and analyzed in all five samples.
Untargeted metabolomic analysis was performed using an ultrahigh-performance liquid chromatograph (UltiMate 3000, Dionex, Sunnyvale, CA, USA) and a mass spectrometer (Q-Exactive Focus, Thermo Fisher Scientific, Waltham, MA, USA). Sample separation was achieved using a Waters ACQUITY UPLC HSS T3 C18 1.8 μm × 2.1 mm × 100 mm column (Waters, Milford, Massachusetts, USA). The elution program was consistent with that described by Zhang et al.[28]. The mobile phases for this method were (A) a 0.05% aqueous formic acid solution and (B) 0.05% formic acid in acetonitrile, with a column temperature of 40 °C, a flow rate of 0.2 mL/min, and an injection volume of 2 µL. The mass spectroscopy (MS) conditions were electrospray ionization (ESI) ± mode MS conditions; the ion source voltage was 2,500/1,500 V; the auxiliary gas flow rate was 8 L/min; the fragmentation voltage was 135 V; the ion source temperature was 325 °C; the sheath temperature was 325 °C; the sheath flow rate was 11 L/min; and the spray voltage was 40 V. The MS–MS scan mode was set to a data-dependent ms2 scan mode with a resolution of 17,500, and collision-induced dissociation was set to a stepped mode of 10, 20, and 60 eV. Each sample was subjected to three biological replicates. Compound identification of metabolites was performed by comparing the accuracy of the m/z value [<10 parts per million (ppm)] and MS–MS spectra with an in-house database established with available authentic standards. The detailed identification of the metabolites is shown in Supplementary Table S2.
Statistical analysis
-
For the analysis of alpha diversity, we employed three distinct diversity indices to assess the species diversity within the samples: Pielou evenness, richness, and Shannon's index. The calculations of these indices and beta diversity indices based on Bray–Curtis distances were performed using QIIME software (version 1.9.1). To more intuitively present the results of these diversity indices, we utilized the ggplot2 software package (version 2.15.3) within the R programming language for data visualization. In the analysis of alpha diversity indices, we conducted a one-way analysis of variance (ANOVA) followed by Duncan's test using SPSS 18.0 statistical software to determine if there were significant differences among the samples, with p < 0.05 considered to be statistically significant. Beta diversity analysis is another method used for assessing the differences in species composition between samples. Principal component analysis (PCA) was conducted and heatmaps were drawn using Origin 9.0 software (Hampton, Massachusetts, USA). Partial least squares discriminant analysis (PLS-DA) was performed using MetaboAnalyst 5.0 (
www.metaboanalyst.ca ). Orthogonal partial least squares discriminant analysis (OPLS-DA) of the metabolomic data was carried out using SIMCA-P 14.0 software (Umetrics, Umeå, Sweden). Variables with a variable importance in projection (VIP) > 1.0, p < 0.05, and a fold change (FC) > 1.50 or < 0.67 were identified as differential metabolites. Metabolic pathways were enriched and topologically analyzed by searching the Kyoto Encyclopedia of Genes and Genomes (KEGG) database[29−31], with Camellia sinensis as the corresponding species. All samples were analyzed in triplicate, and standard deviations were calculated. -
The quality of Taihu Cuizhu tea is profoundly influenced by the interplay between harvest season, microbial communities, and metabolite profiles, which collectively determine its sensory and nutritional attributes (Figs 1−3, Table 1). Sensory evaluations revealed that special-grade autumn teas consistently excelled in appearance, aroma, and leaf integrity, exhibiting a balanced flavor profile with reduced bitterness. In contrast, first-grade spring teas stood out in overall sensory quality, likely caused by elevated levels of soluble polysaccharides, polyphenols, and free amino acids (Fig. 1a–d, Table 1). These seasonal variations align with findings from Longjing tea, where spring harvests benefit from favorable climatic conditions—such as increased sunlight and moderate temperatures—that enhance biosynthetic activity[32]. However, this study reveals, through an in-depth analysis of Taihu Cuizhu tea, that its quality formation mechanism may exhibit cultivar-specific characteristics. Microbiomic analysis further elucidated this relationship: Spring teas displayed greater microbial diversity, suggesting that warm, humid conditions promote microbial colonization (Fig. 2a−d), amplifying the synthesis of flavor compounds like L-theanine, a key contributor to umami flavors[33]. Proteobacteria dominated all samples, with Sphingomonas being the predominant genus, which was particularly abundant in spring teas, followed by Methylobacterium and Rhizobium in autumn and spring, respectively (Fig. 3a, e). Sphingomonas, known for its polyphenol oxidase activity in Sichuan black tea, may enhance amino acid accumulation[34]. We speculate that it may play a similarly yet more distinctive role in the formation of Taihu Cuizhu's unique flavor profile (such as its refreshing taste). Strong correlations between Proteobacteria and eight flavonoid compounds support this notion. These correlations with flavor-related metabolites indicating microbial metabolism amplify seasonal biochemical differences (Fig. 4). This suggests that microbial metabolism not only amplifies general seasonal differences but may also directly participate in shaping the unique flavor characteristics that distinguish Taihu Cuizhu from other tea categories like Longjing. Conversely, autumn teas accumulated higher levels of alkaloids like cephalotaxine, cinchonidine (Fig. 5), possibly as a stress response to cooler temperatures and reduced photoperiod, mediated by Proteobacteria's metabolic flexibility[35,36]. Beta diversity analysis confirmed distinct microbial communities between the seasons (Fig. 2d), suggesting that seasonal microbial succession drives the metabolite profiles: Spring teas favor flavonoids, whereas autumn teas accumulate alkaloids (Fig. 5). Unlike fermented Pu-erh tea, where microbial activity drives theaflavin's formation, Taihu Cuizhu's unfermented nature highlights the unique role of endophytic microbiota in green tea quality[37]. This linkage redefines tea quality as a co-metabolic outcome of plant physiology and microbial ecology, where environmental factors shape the microbial communities that modulate the flavor and nutritional compounds. These findings suggest that microbial communities are not merely indicators of seasonal variation but active regulators in the formation of tea flavor. By directly participating in the biochemical transformation of tea leaves, they translate and amplify seasonal environmental signals into distinct metabolite profiles. For instance, spring conditions may enhance microbial enzyme activity, boosting compounds like L-theanine, whereas autumn's cooler climate triggers alkaloid production. This opens avenues for manipulating environmental factors, such as the abundance of Sphingomonas, to tailor flavor profiles. Future research could explore how pre-harvest factors such as soil microbiota and rainfall influence this dynamic, potentially using metagenomics to map microbial enzyme pathways or field studies to assess climate-driven microbial shifts.
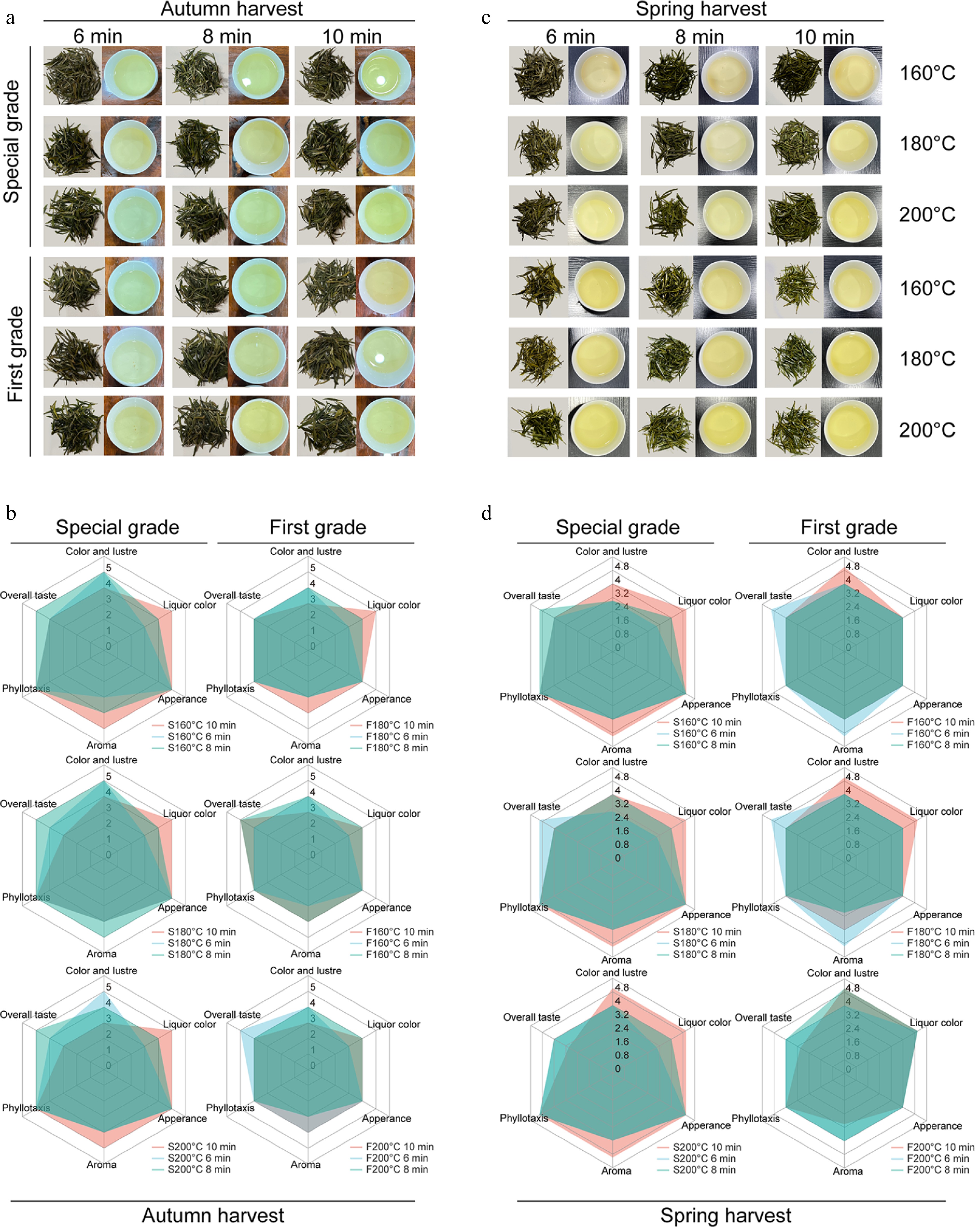
Figure 1.
The sensory evaluation of all the treatments of Taihu Cuizhu tea. (a), (b) Images of dry tea and tea infusions from different harvesting seasons of Taihu Cuizhu tea. (c), (d) The quantitative descriptions of different treatments of Taihu Cuizhu tea.
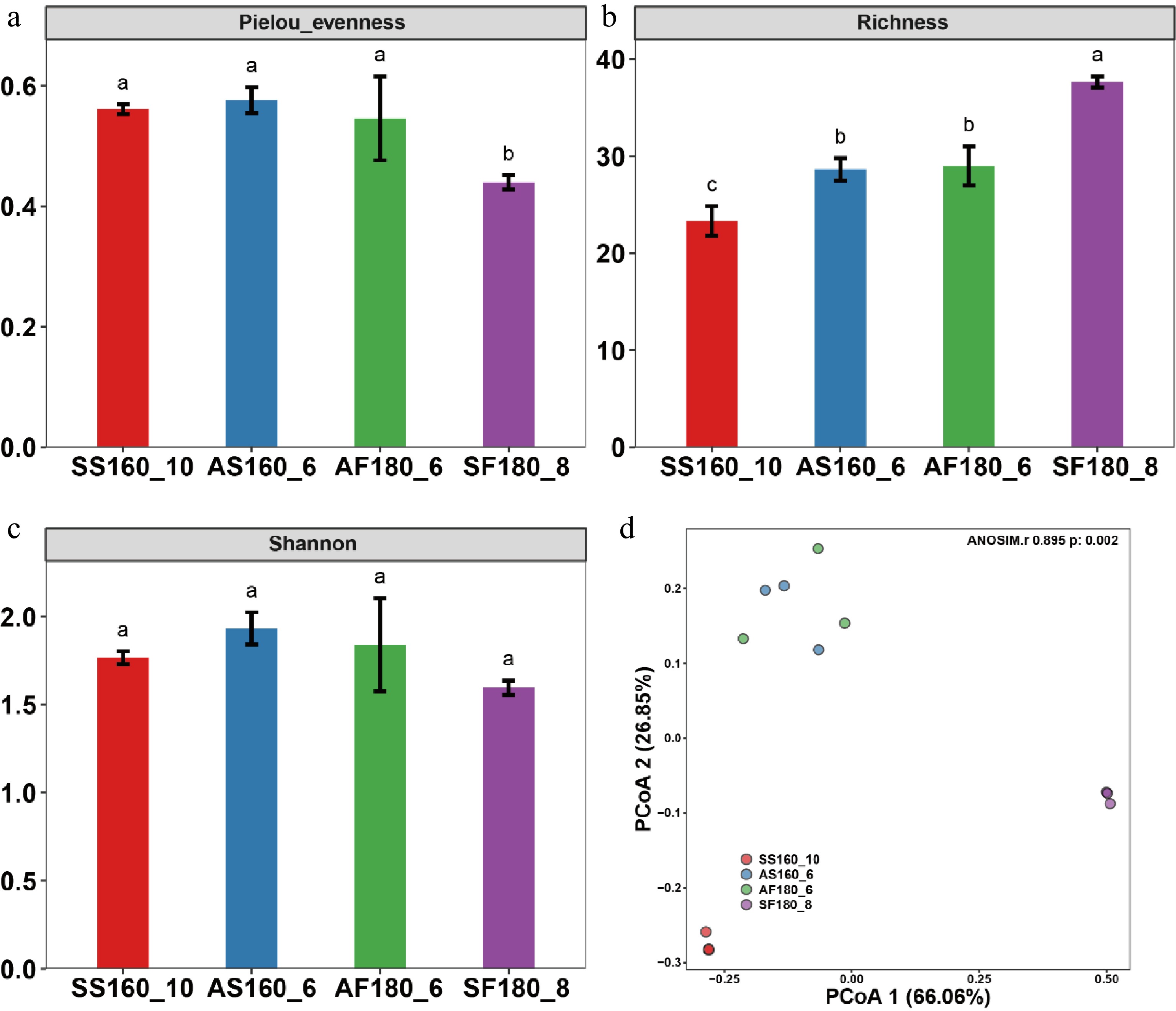
Figure 2.
Alpha and beta diversity analysis of tea bacterial communities. (a) Pielou eveness; (b) richness; (c) Shannon diversity index; and (d) principal coordinate analysis. SS160_10: Special-grade tea fixed at 160 °C for 10 min harvested in spring; AS160_6: Special-grade tea fixed at 160 °C for 6 min harvested in autumn; AF180_6: First-grade tea fixed at 180 °C for 6 min harvested in autumn; SF180_8: First-grade tea fixed at 180 °C for 8 min harvested in spring.
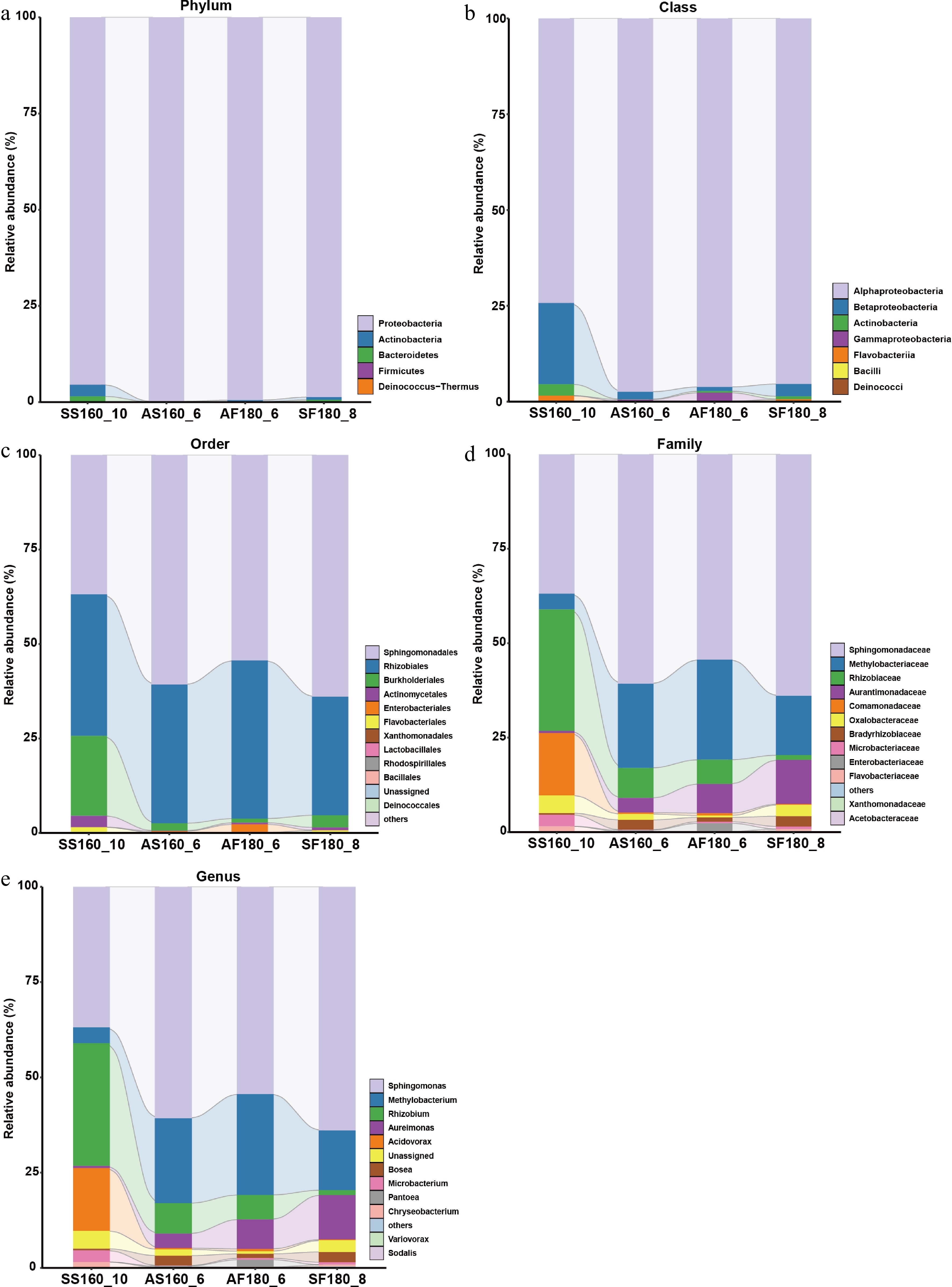
Figure 3.
The bacterial community composition of tea leaves after different optimized methods of fixation: (a) Phylum, (b) class, (c) order, (d) family, and (e) genus.
Table 1. Content of quality components of Taihu Cuizhu tea under different treatments.
Temperature
(°C)Time
(min)pH value Water content Soluble polysaccharides Total flavonoids Tea polyphenols Free amino acids Spring Autumn Spring Autumn Spring Autumn Spring Autumn Spring Autumn Spring Autumn Special-grade 160 6 5.60 ± 0.03c 5.99 ± 0.05a 0.17 ± 0.00a 0.04 ± 0.00a 12.91 ± 1.03b 3.07 ± 0.03a 45.57 ± 0.39b 108.06 ± 39.58a 0.11 ± 0.00b 0.13 ± 0.00a 38.05 ± 0.13b 29.93 ± 0.45a 8 6.15 ± 0.01a 5.98 ± 0.06a 0.07 ± 0.02b 0.03 ± 0.00b 12.14 ± 0.79b 2.49 ± 0.04a 52.02 ± 1.08a 47.36 ± 9.54b 0.10 ± 0.00c 0.10 ± 0.01b 39.53 ± 0.42a 25.53 ± 0.39c 10 6.10 ± 0.00b 5.98 ± 0.02a 0.12 ± 0.04ab 0.03 ± 0.00c 15.71 ± 1.50a 3.62 ± 0.07b 54.95 ± 2.91a 39.07 ± 0.41b 0.14 ± 0.00a 0.10 ± 0.00b 37.73 ± 0.26b 26.66 ± 0.56b 180 6 6.07 ± 0.02b 6.18 ± 0.03a 0.15 ± 0.00a 0.03 ± 0.00a 15.47 ± 1.10a 3.87 ± 0.15a 47.67 ± 1.53b 53.93 ± 22.66a 0.13 ± 0.00b 0.11 ± 0.00a 32.21 ± 0.32c 19.05 ± 0.29c 8 6.14 ± 0.01a 6.19 ± 0.02a 0.05 ± 0.05b 0.03 ± 0.00b 15.83 ± 0.42a 3.16 ± 0.04a 51.39 ± 2.63ab 53.20 ± 9.71a 0.14 ± 0.00a 0.10 ± 0.00b 35.87 ± 0.08b 25.34 ± 0.24a 10 6.16 ± 0.03a 6.22 ± 0.03a 0.04 ± 0.03b 0.02 ± 0.00c 10.82 ± 0.81b 3.37 ± 0.15b 54.86 ± 3.09a 74.23 ± 41.24a 0.14 ± 0.00a 0.06 ± 0.00c 40.58 ± 0.50a 24.45 ± 0.54b 200 6 6.19 ± 0.01a 6.22 ± 0.02a 0.08 ± 0.04a 0.03 ± 0.01a 13.04 ± 0.76b 2.46 ± 0.15a 47.63 ± 3.03a 44.84 ± 5.91b 0.15 ± 0.00a 0.08 ± 0.00b 34.10 ± 0.30c 13.40 ± 0.07c 8 6.16 ± 0.01b 6.15 ± 0.04b 0.01 ± 0.01b 0.02 ± 0.00b 14.12 ± 0.93ab 2.93 ± 0.06b 46.87 ± 1.10a 80.93 ± 13.41a 0.13 ± 0.00c 0.08 ± 0.00b 37.09 ± 0.29b 31.21 ± 0.47a 10 6.16 ± 0.01b 6.09 ± 0.03c 0.00 ± 0.00b 0.02 ± 0.00b 15.36 ± 0.86a 2.77 ± 0.15b 46.37 ± 2.76a 51.81 ± 6.68b 0.14 ± 0.00b 0.11 ± 0.00a 40.77 ± 0.52a 26.99 ± 0.55b First-grade 160 6 6.00 ± 0.01b 6.13 ± 0.02b 0.13 ± 0.00a 0.04 ± 0.00a 17.22 ± 0.61a 2.83 ± 0.08a 53.81 ± 1.84b 50.02 ± 2.76a 0.11 ± 0.00b 0.09 ± 0.00a 34.56 ± 0.30a 23.80 ± 1.05b 8 6.10 ± 0.01a 6.26 ± 0.05a 0.06 ± 0.00b 0.03 ± 0.00ab 18.21 ± 0.28a 3.14 ± 0.05b 51.42 ± 1.65b 42.45 ± 5.27a 0.11 ± 0.00a 0.07 ± 0.00c 32.97 ± 0.29b 23.96 ± 0.78b 10 6.11 ± 0.03a 6.21 ± 0.05a 0.02 ± 0.00c 0.03 ± 0.00b 15.49 ± 2.37a 3.34 ± 0.29b 69.00 ± 5.91a 52.27 ± 7.96a 0.11 ± 0.00a 0.08 ± 0.00b 30.80 ± 0.17c 30.68 ± 0.87a 180 6 6.15 ± 0.02a 6.24 ± 0.03a 0.09 ± 0.00a 0.04 ± 0.00a 12.91 ± 1.21c 3.30 ± 0.05a 48.20 ± 1.32b 67.13 ± 21.00a 0.11 ± 0.00c 0.10 ± 0.00b 31.43 ± 0.35c 32.27 ± 0.88a 8 6.09 ± 0.01c 6.32 ± 0.03a 0.04 ± 0.01b 0.03 ± 0.00b 16.20 ± 1.33b 3.06 ± 0.05b 58.41 ± 2.46a 83.85 ± 47.69a 0.15 ± 0.00a 0.07 ± 0.00c 32.52 ± 0.40b 26.60 ± 0.55b 10 6.12 ± 0.01b 6.29 ± 0.05a 0.01 ± 0.01c 0.03 ± 0.00b 19.19 ± 1.42a 3.29 ± 0.08a 51.19 ± 1.74b 58.64 ± 9.08a 0.13 ± 0.00b 0.18 ± 0.00a 34.38 ± 0.40a 20.50 ± 0.37c 200 6 6.23 ± 0.01a 6.29 ± 0.05a 0.04 ± 0.00a 0.03 ± 0.00a 17.45 ± 2.84a 2.29 ± 0.02a 51.53 ± 1.90a 64.74 ± 12.12a 0.13 ± 0.00b 0.12 ± 0.01a 36.63 ± 0.48a 30.37 ± 0.64c 8 6.12 ± 0.01b 6.16 ± 0.04b 0.07 ± 0.04a 0.03 ± 0.00b 15.56 ± 0.52a 2.45 ± 0.11a 52.65 ± 0.61a 40.66 ± 8.08b 0.11 ± 0.00c 0.10 ± 0.01b 28.93 ± 0.10c 33.09 ± 0.57a 10 6.10 ± 0.01c 6.16 ± 0.03b 0.17 ± 0.13a 0.03 ± 0.00b 15.04 ± 0.75a 2.33 ± 0.08a 54.54 ± 3.94a 50.08 ± 2.99ab 0.14 ± 0.00a 0.13 ± 0.01a 34.58 ± 0.91b 31.69 ± 0.55b Different letters indicate significant differences among different drying times within the same temperature. 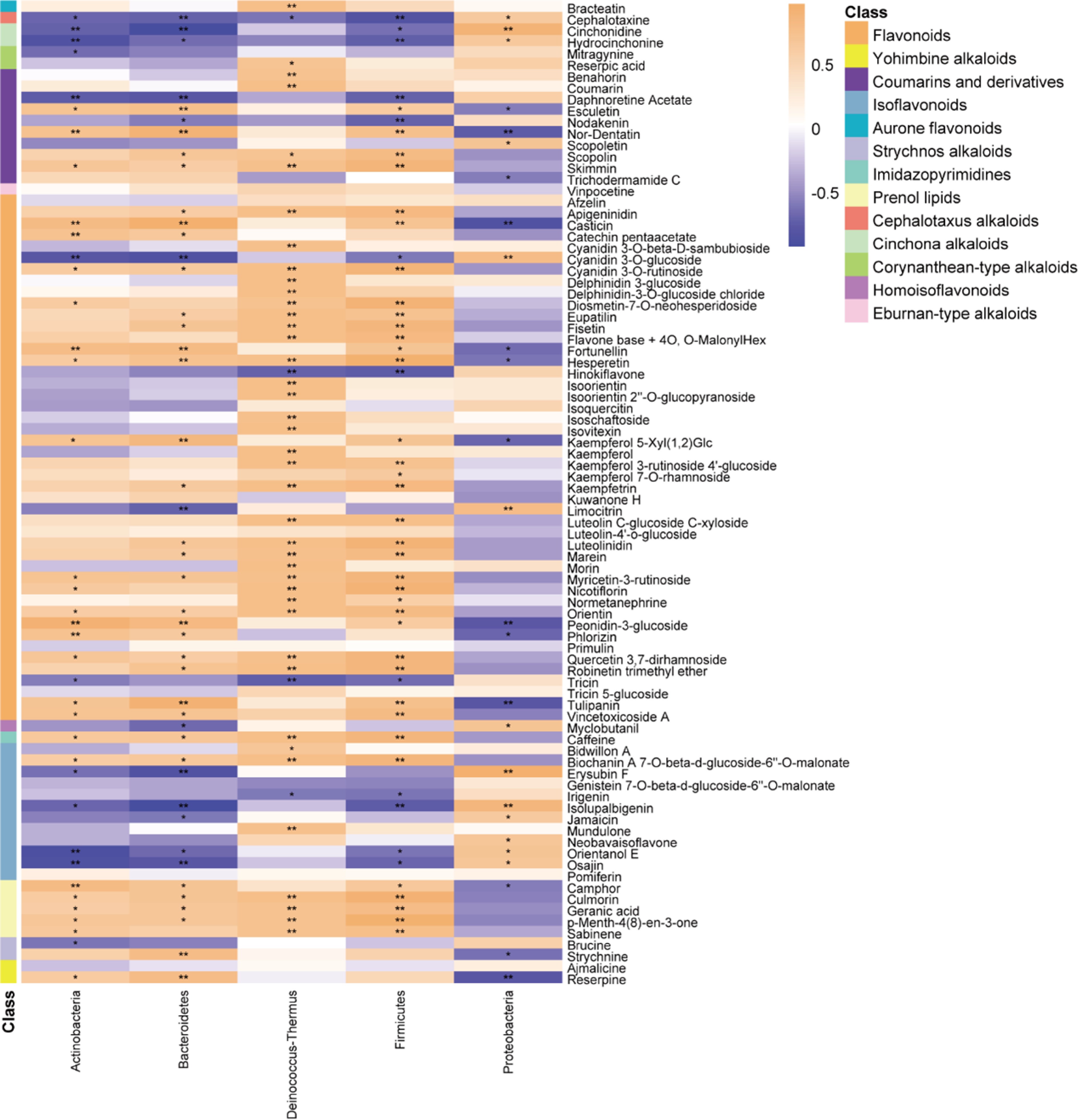
Figure 4.
Correlation between tea metabolites and bacteria in Taihu Cuizhu tea (* p < 0.05, ** p < 0.01; *** p < 0.001).
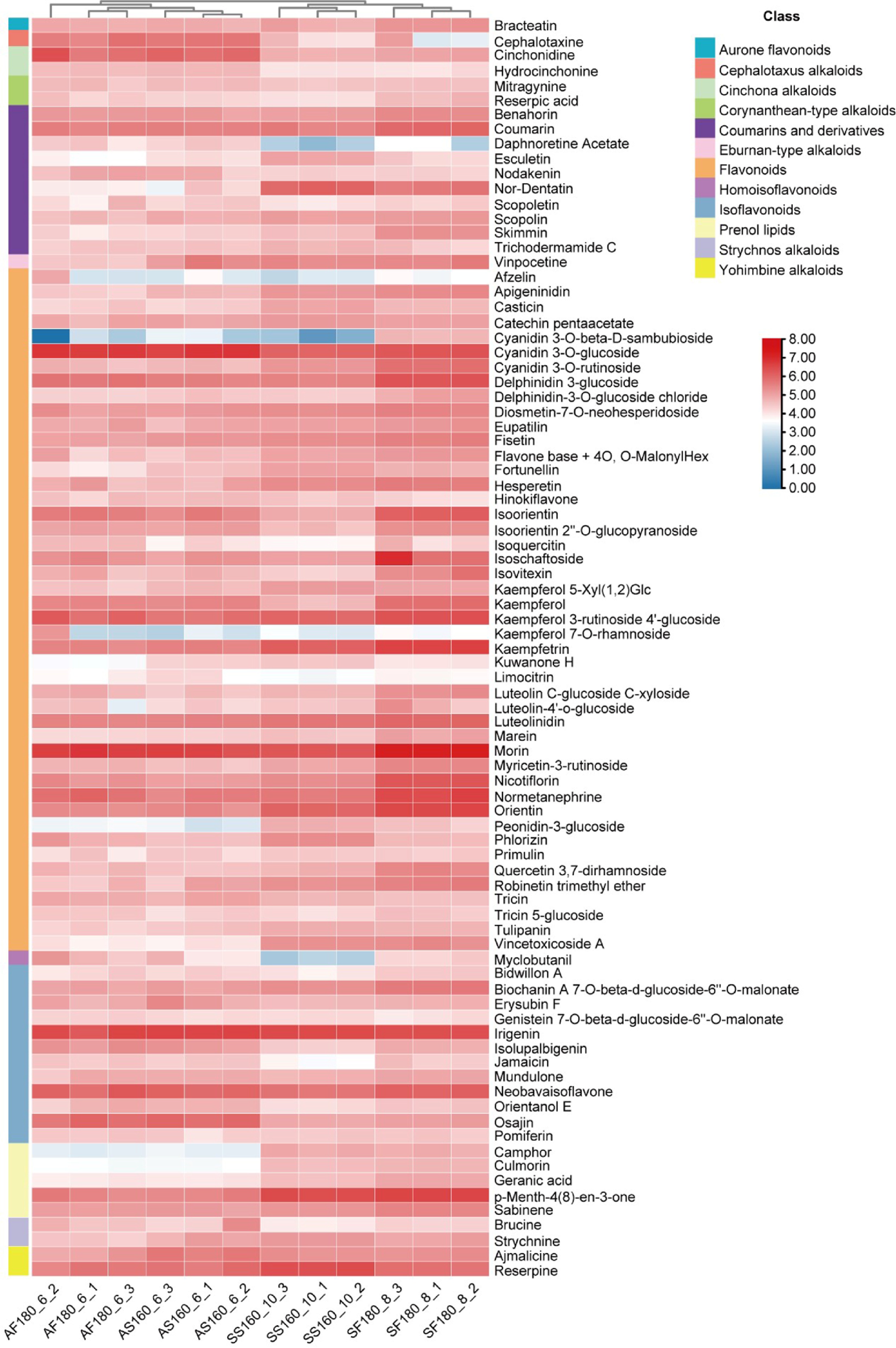
Figure 5.
Heatmap of the expression of tea metabolites in different samples. The color scale represents values normalized by log10(N+1), where N denotes the original measurement for each metabolite.
Fixation as a catalyst for optimizing bioactive compounds
-
The fixation conditions act as a metabolic switch, dynamically regulating Taihu Cuizhu's biochemical and metabolomic profiles to optimize bioactive compounds (Table 1, Figs 6 and 7). We compared the contents of the four bioactive components under different fixation conditions (Table 1) and found that the optimal fixation time for special-grade spring tea was 10 min, with the corresponding optimal fixation temperature being 160 °C. Moreover, special-grade autumn tea, first-grade spring tea, and first-grade autumn tea each had two relatively favorable fixation conditions, namely AS160_6, AS200_8, SF180_8, SF200_6, AF180_6, and AF200_8. Further comparisons revealed significant differences in the bioactive components under different fixation conditions. The contents of soluble polysaccharides, total flavonoids, and tea polyphenols in AS160_6 were higher than those in AS200_8, especially the content of tea polyphenols, which showed a significant difference. The contents of tea polyphenols and total flavonoids in SF180_8 were significantly higher than those in SF200_6, and the content of polysaccharides in AF180_6 was significantly higher than that in AF200_8. In summary, the optimal fixation conditions for the four types of tea are SS160_10, AS160_6, SF180_8, and AF180_6. The optimal fixation temperature for special-grade tea is 160 °C; for first-grade tea, it is 180 °C. Additionally, spring tea benefits from a longer fixation time (8 min), whereas autumn tea requires a shorter fixation time (6 min). These conditions maximized catechin, flavonoid, and free amino acid levels (Table 1). Lower temperatures preserved catechins in special-grade teas by minimizing thermal degradation, consistent with green tea processing[38], whereas a shorter fixation time reduced teh oxidation of polyphenols in autumn teas, retaining a desirable liquor color (Fig. 1c, d). Tea infusions remained mildly acidic, adhering to green tea standards, and higher fixation temperatures reduced the moisture content, reflecting enhanced water evaporation (Table 1). Metabolomic analysis revealed that spring teas, especially SF180_8, exhibited greater diversity in differential metabolites, including flavonoids and catechin derivatives, driven by fixation-induced changes (Figs 5, 7c and d). SS160_10 was enriched in catechin pentaacetate (Fig. 5), likely because of reduced enzyme inactivation at lower temperatures[39]. The bacterial co-occurrence networks in SF180_8 showed higher connectivity (Figs 4 and 6), suggesting that elevated fixation temperatures strengthen microbial interactions, potentially upregulating flavonoid biosynthesis via the polyphenol metabolism of Sphingomonas[40]. This microbial–metabolic interplay is novel for green tea, as prior research has focused on fermentation-driven effects in black and dark teas[37,41]. Taihu Cuizhu's fixation-driven changes offer a unique mechanism for quality customization. For example, low-temperature fixation in autumn teas may preserve alkaloids, enhancing astringency, whereas high-temperature fixation in spring teas could amplify flavonoid-driven antioxidant properties. This metabolic switch concept positions fixation as a precision tool for targeting specific bioactive profiles, such as antioxidant-rich flavonoids for health-focused teas or autumn teas with a distinct balance between alkaloids and other compounds for favorable traditional flavors. Microbial contributions suggest that the fixation parameters could be tuned to harness specific enzymes, such as the polyphenol oxidase of Sphingomonas, to elevate desired metabolites[42]. Liu found that supplementing the green tea fermentation process with Eurotium cristatum altered the activity of polyphenol oxidase and α-amylase during fermentation, leading to changes in the content of bioactive compounds in tea, as well as significantly higher in vitro inhibition of angiotensin-converting enzymes[43]. Such approaches align with precision processing trends in food science, like controlled coffee roasting for flavor modulation[44]. Future studies might explore real-time microbial activity during fixation using proteomics or enzyme assays or investigate fixation's impact on microbial gene expression to optimize bioactive synthesis.
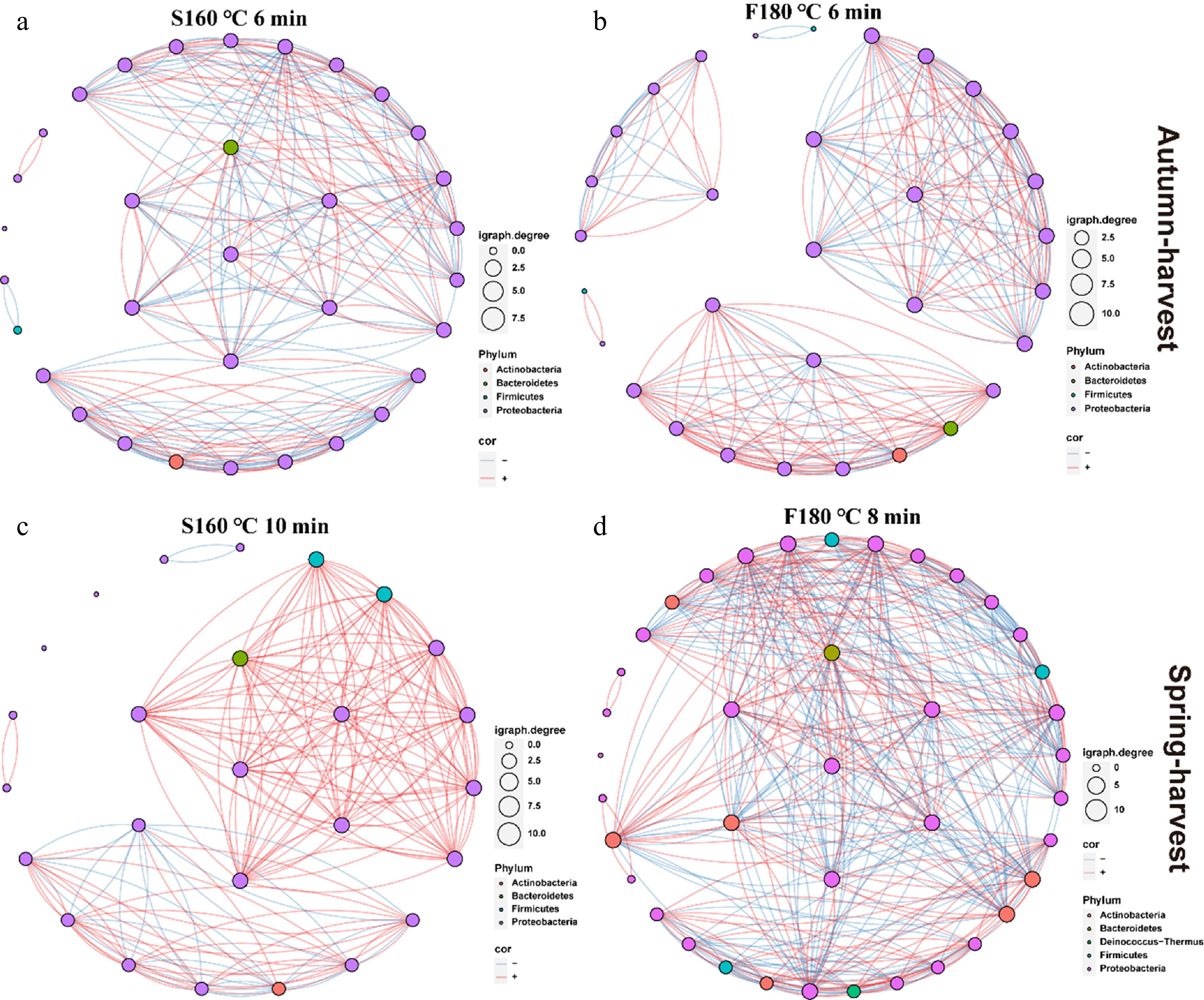
Figure 6.
Bacterial co-occurrence network diagrams of tea leaves treated with four different optimized fixation methods. The size of each node is proportional to the number of connections. Red and blue edges represent positive and negative correlations, respectively. (a) S160 °C 6 min, (b) F180 °C 6 min, (c) S160 °C 10 min, (d) F180 °C 8 min.
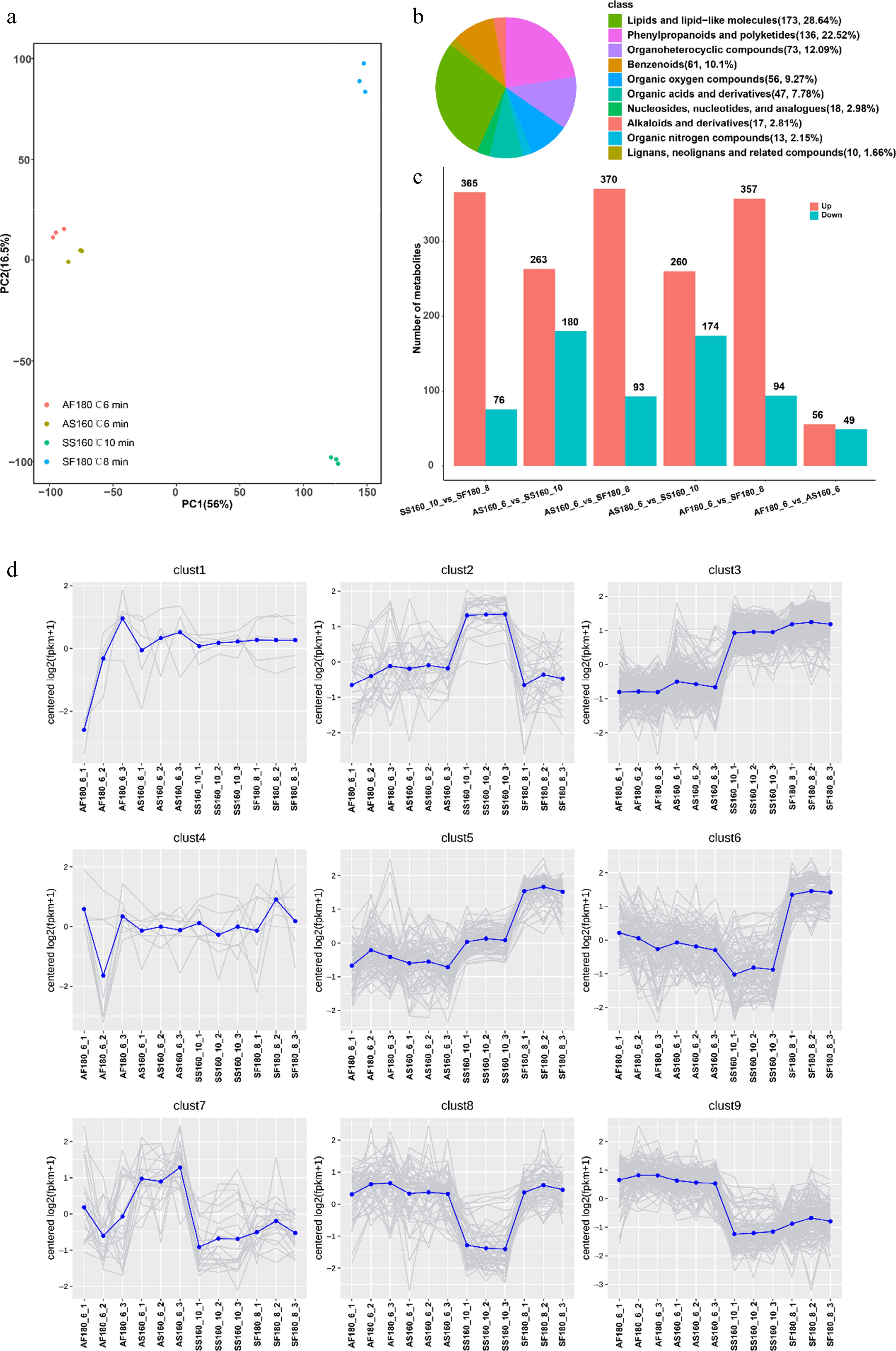
Figure 7.
Metabolites diversity analysis among four treatments. (a) PCA score plots for all samples; (b) The classification of metabolites; (c) The number of DAMs in different comparison groups; (d) K-means plot of differential metabolite.
Microbial community stability as a quality regulator
-
Microbial community stability, particularly in spring teas, emerged as a key regulator of Taihu Cuizhu's quality, maintaining consistent sensory and bioactive attributes through beneficial microbial interactions (Figs 2−4, and 6). Proteobacteria, with Sphingomonas as the dominant genus, prevailed across all samples, but spring teas exhibited greater microbial diversity and stability, evidenced by uniform α-diversity patterns and robust co-occurrence networks (Figs 2a–c and 6). This stability likely stems from spring's favorable climate, supporting microbial colonization and sustained enzyme activity that enhances flavor compounds like L-theanine and polysaccharides (Fig. 4)[45]. In contrast, autumn teas displayed less stable communities, with a higher abundance of Methylobacterium, possibly as an adaptation to cooler, drier conditions (Fig. 3e)[46]. Spring teas' stronger bacterial network connectivity, especially in SF180_8 (Fig. 6), implies that stable microbial interactions amplify bioactive compounds like flavonoids[47]. This stability ensures reliable production of flavor and nutritional compounds, particularly in spring teas, where higher microbial diversity correlates with superior sensory traits (Fig. 1b, d). The interaction between fixation and microbial stability suggests that processing conditions (such as SF180_8's higher temperature) reinforce beneficial microbial interactions, as evidenced by stronger network connectivity (Fig. 6). This insight offers a new perspective for tea production: Stabilizing the microbial communities through environmental or processing control could enhance the consistency of quality.
Seasonal biosynthetic trade-offs in metabolite profiles
-
Seasonal biosynthetic trade-offs between flavor-enhancing and stress-related metabolites, driven by environmental differences, shape Taihu Cuizhu's sensory and nutritional characteristics (Figs 5, 7 and 8). Metabolomic analysis revealed spring teas' greater diversity in differential metabolites, particularly flavonoids and glycerophospholipids linked to anthocyanin and caffeine metabolism, supporting flavor synthesis and antioxidant properties (Figs 5, 7c, and 8b–e)[48−50]. Autumn teas, however, accumulated alkaloids such as cephalotaxine, cinchonidine, and stilbenoids, reflecting stress responses to cooler conditions (Figs 5, 7c, and 8f)[51]. Clustering analysis identified distinct metabolic expression profiles: Spring teas were rich in catechins (SS160_10), whereas autumn teas favored alkaloids, highlighting a trade-off between flavor-enhancing compounds (flavonoids, amino acids) and stress-related compounds (alkaloids, stilbenes) (Figs 5 and 7d). Heatmap analysis confirmed this (Fig. 5), with spring teas' flavonoid abundance contributing to sweetness and antioxidants[52], and autumn teas' alkaloids enhancing astringency[53]. This seasonal biosynthetic trade-off is primarily driven by key environmental stress factors such as temperature and photoperiod. The relatively warm temperatures and gradually increasing daylight in spring promote the accumulation of photosynthetic products and energy metabolism (the tricarboxylic acid cycle), providing ample precursors for the substantial synthesis of flavor compounds (catechins, amino acids). Simultaneously, higher light intensity may directly upregulate the phenylpropanoid metabolism pathway [phenylalanine ammonia-lyase (PAL) and chalcone synthase (CHS) enzyme activity], thereby specifically enhancing the biosynthesis of flavonoids[54]. In contrast, the low-temperature stress and shortened daylight hours in autumn trigger plants' stress signaling pathways. This may lead to a shift in the metabolic flow of carbon and nitrogen. On the one hand, low temperatures may inhibit some kinds of growth-related primary metabolism; on the other hand, as a defense mechanism, stress signals activate the synthesis pathways of alkaloids and stilbenoids.[55]. KEGG pathway analysis supported this, showing that spring teas were enriched in pathways for active bud growth, and autumn teas were enriched in stress responses (Fig. 8). This trade-off framework enables producers to target specific metabolite profiles by adjusting the time of harvest, such as early spring for flavonoid-rich health teas or late autumn for a distinct balance between alkaloids and other compounds for favorable traditional flavors. Similar to cocoa, where environmental stress influences the alkaloid content[56], Taihu Cuizhu's model offers a parallel for tea.

Figure 8.
Enrichment analysis of KEGG pathways in different comparison groups. (a) SS160_10 vs. SF180_8; (b) AS160_6 vs. SS160_10; (c) AS160_6 vs. SF180_8; (d) AF180_6 vs. SS160_10; and (e) AF180_6vs. SF180_8; (f) AF180_6 vs. AS160_6.
Multi-omics framework for precision tea production
-
Integrated multi-omics, combining sensory, biochemical, microbial, and metabolomic data, establishes Taihu Cuizhu as a model for precision tea production, where harvest and fixation protocols can be tailored to seasonal and microbial contexts to maximize quality (Figs 5, 7 and 8). This blueprint enables data-driven optimization. For instance, producers could leverage seasonal climate differences to predict microbial and metabolite distributions to optimize the time of harvest. Early spring harvests may maximize flavonoids, whereas late autumn harvests enhance alkaloids. Spring teas processed at higher temperatures could target health-conscious markets with flavonoid-rich antioxidants[57,58], whereas autumn teas fixed at lower temperatures may appeal to traditional markets favoring alkaloid-driven flavors[59]. This precision approach contrasts with empirical methods, aligning with precision agriculture trends[60]. Taihu Cuizhu's multi-omics framework links terroir to flavor, integrating environmental and microbial factors with quality. Future research could refine this model using real-time environmental monitoring, machine learning, and metabolomics, potentially extending it to other teas such as Longjing, matcha, or crops like coffee.
-
Integrated multi-omics analyses combining sensory evaluations, biochemical profiling, microbial characterization, and metabolomic data, establishes a comprehensive framework for optimizing Taihu Cuizhu tea quality. This approach synthesizes seasonal–microbial–metabolite relationships, positions fixation as a metabolic switch, explains the microbial community's stability, and reveals biosynthetic trade-offs. Sensory assessments demonstrate the superior flavor profile of special-grade autumn teas and the robust sensory characteristics of first-grade spring teas, driven by biochemical variations in polysaccharides, polyphenols, and free amino acids. Microbiomic analysis revealed Proteobacteria's dominance, with Sphingomonas enhancing flavor compounds in spring teas. The optimal fixation temperature for special-grade tea is 160 °C; for first-grade tea, it is 180 °C. Additionally, spring tea benefits from a longer fixation time (8 min), whereas autumn tea requires a shorter fixation time (6 min). Fixation conditions modulated these characteristics: Lower temperatures preserved catechins, whereas shorter fixation times reduced polyphenol oxidation in autumn teas. The stable microbial communities in spring teas ensured consistent quality, with fixation processes amplifying the microbial–metabolic interactions. Metabolomic profiling showed that spring teas were flavonoid-rich, whereas autumn teas accumulated alkaloids, reflecting distinct seasonal biosynthetic strategies. This integrative framework enables producers to customize harvesting and processing for targeted quality outcomes, such as antioxidant-rich spring teas or astringent autumn variants. The Taihu Cuizhu model exemplifies how terroir influences flavor through dynamic interactions among environmental factors, microbial ecology, and processing parameters, showcasing the multi-omics framework's significant potential for precision tea production. Future work could focus on the validation and manipulation of functional microorganisms, the precise optimization of the entire processing chain, and the extension of this framework to other specialty tea varieties.
-
The authors confirm their contributions to the paper as follows: conception and design of the research: Li C, Yuan Y, Shen R; acquisition of data: Li C; analysis and interpretation of data: Zhou R, Zhu Y; statistical analysis: Zhou R; drafting the manuscript: Li C, Yuan Y, Zhou R; revision of manuscript for important intellectual content: Li C, Zhu Y, Yuan Y. All authors read and approved the final manuscript.
-
The datasets generated during and/or analyzed in the current study are available from the corresponding author on reasonable request.
-
Not applicable.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Chutian Li, Ru Shen
- Supplementary Table S1 Sequencing depth of samples.
- Supplementary Table S2 Detailed metabolite information
- Supplementary Fig. S1 Rarefaction curves of the 16S rRNA gene sequencing.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li C, Shen R, Zhou R, Zhu Y, Yuan Y. 2026. Decoding Taihu Cuizhu tea quality: Multi-omics reveals seasonal and fixation effects. Beverage Plant Research 6: e008 doi: 10.48130/bpr-0025-0038 

Decoding Taihu Cuizhu tea quality: Multi-omics reveals seasonal and fixation effects
- Received: 03 July 2025
- Revised: 13 September 2025
- Accepted: 05 October 2025
- Published online: 11 February 2026
Abstract: Through an integrated multi-omics approach, this study deciphers how seasonal, microbial, and metabolic factors collectively shape the quality of Taihu Cuizhu green tea. Special-grade autumn teas exhibit superior appearance, aroma, and balanced flavor with reduced bitterness, whereas first-grade spring teas contain higher levels of soluble polysaccharides, polyphenols, and amino acids, contributing to their enhanced sensory quality. Spring teas showed greater microbial diversity, with Sphingomonas (Proteobacteria) promoting the synthesis of flavor compounds such as L-theanine. In contrast, autumn teas accumulated stress-induced alkaloids like cephalotaxine under cooler conditions. Fixation acts as a metabolic switch: optimal results were achieved at 160 °C for special-grade tea and 180 °C for first-grade tea, with spring tea benefiting from longer fixation (8 min) and autumn tea requiring a shorter duration (6 min). These parameters maximized the preservation of catechin and flavonoids while maintaining sensory traits. Spring teas demonstrated more stable microbial communities, strengthened by fixation, with high-temperature fixation (SF180_8) enhancing bacterial interactions and flavonoid biosynthesis. Metabolomic profiling confirmed a seasonal biosynthetic trade-off: Flavonoids dominated in spring, supporting antioxidant capacity, whereas alkaloids accumulated in autumn, enhancing astringency. This multi-omics framework provides a foundation for precision tea production, enabling targeted harvesting and processing strategies to achieve specific quality profiles. Future studies may integrate real-time monitoring and machine learning to optimize and extend this model to other tea varieties and agricultural products.
-
Key words:
- Taihu Cuizhu Tea /
- Sensory evaluation /
- Fixation conditions /
- Microbial communities /
- Multi-omics