









-
Tea is one of the most widely consumed beverages globally, with significant economic, cultural, and health benefits. As it is a perennial woody crop cultivated across diverse geographical regions, tea production has expanded significantly over the past decades to meet increasing global demand, with current annual production exceeding 6 million tons (International Tea Committee,
https://inttea.com ). The quality and yield of tea are intrinsically linked to the growing environment, particularly the soil conditions, making soil health in tea plantations a critical factor in sustainable tea production. Global tea plantations are primarily distributed in tropical and subtropical regions. China is a key area for tea plant cultivation. The soil in this region has characteristics that differ from those of other typical acidic soils[1]. The soil in tea plantations has developed unique properties as a result of long-term cultivation, including uneven fertilization and the decomposition of pruning residues. These factors create a unique environment in tea plantation ecosystems, characterized by natural acidification and a high Al content[2]. In fact, in most regions of the world, particularly in China, the pH value of tea plantation soils ranges from approximately 3.96 to 5.48[3].Healthy soil is essential for the vigorous growth and the production of high-quality tea. The soil's nutrient composition, particularly the nitrogen (N) and potassium (K) levels, significantly influences the accumulation of amino acids and tea polyphenols. Additionally, other nutrient elements, including magnesium (Mg), zinc (Zn), and manganese (Mn), are crucial for developing flavor compounds[4,5]. The physical structure of tea plantation soil affects root growth and water availability, both of which are vital for bud and leaf germination. Furthermore, the soil conditions affect the synthesis of secondary metabolites in tea plants, influence the mineral composition of tea leaves, and are critical to the development of distinctive aroma compounds, ultimately determining the tea's quality and yield[5].
Soil microorganisms are vital to the ecosystem. They contribute significantly to soil health, enhance plant growth, and support the circulation of materials and energy within the natural ecosystem. Previous research has indicated that the structure of soil microbial communities serves as a crucial indicator of soil health and productivity in a complete agricultural ecosystem[6,7]. Consequently, the condition of tea plantation soil hinges on the rational structure of its microbial community and the efficient operation of its functions. Numerous studies have shown that specific microbial communities play crucial roles in several key physiological processes in tea plants' growth, including nutrient absorption, plant hormone balance, and responses to stress[8−11]. At the same time, an increasing number of researchers have recognized and appreciated the roles of soil microorganisms in the decomposition and formation of humus; the cycling and energy flow of substances like carbon (C), N, and phosphorus (P); and the fixation and storage of organic C within the global ecosystem[12,13]. Understanding the diverse composition and functions of microbial communities in tea plantation soil is crucial for grasping their role in production and ecology[14]. Despite this, comprehensive research on how soil microorganisms in tea plantations affect and modulate the complex physiological activities of tea plants, along with their role in sustaining the ecosystem's stability, remains insufficient. In addition, continuous tea monocropping has led to numerous problems, characterized by decreased soil fertility, elevated disease incidence, and declining tea quality, all of which threaten the sustainable development of the tea industry[2,3,10]. Key contributing factors, including the excessive application of chemical fertilizers and pesticides and soil acidification, undermine the diversity and functional stability of soil microorganisms[3,15].
In this review, we will systematically review the relationships among tea plantation soils, microorganisms, and tea plants; assess how various agricultural practices influence the microbial community; and discuss emerging research techniques and methodologies. Against this backdrop, this review highlights the paucity of research on microbial functions as a core challenge, especially regarding the practical barriers to establishing causal and predictive relationships between the microbial community's structure and key ecosystem indicators, including sustained yield, the development of quality, and stress adaptation. This critical gap has hindered in-depth analyses of soil–microbe–tea plant interactions and the development of science-based microbial regulation strategies for managing tea plantations. Additionally, we propose improving the sustainability of tea plantations by leveraging the roles of soil microorganisms and highlighting future research directions.
-
The tea plantation soil microbiome contributes to multiple ecosystem functions and services, such as organic matter decomposition, nutrient cycling, and support for tea quality and health regulations, through its interactions with a complex environment and other organisms. Therefore, revealing the composition of tea plantations' soil microbial communities can help us better understand the functional profiles that the soil microbiome provides.
Bacteria
-
Bacteria are the primary microbial group in tea plantation soils, significantly influencing both tea plants' growth and ecosystem health. The main bacterial phyla in these environments include Proteobacteria, Actinobacteria, Acidobacteria, Chloroflexi, and Firmicutes[15]. In particular, nitrifying bacteria from the Proteobacteria phylum, including certain β-proteobacteria and γ-proteobacteria, can convert ammonium N into nitrate N, which plants can absorb and use, thereby enhancing N use efficiency in tea plants. N-fixing bacteria, such as Rhizobia and Helicobacter, convert atmospheric N2 into ammonia, boosting soil N levels. This process accelerates plants' growth and enhances their ability to absorb CO2 during photosynthesis, thereby improving the soil's C sink capacity[16]. Actinomycetes constitute the majority of soil bacteria, particularly exemplified by Streptomyces. Studies suggest that this microorganism can produce various antibiotics, such as streptomycin and tetracycline, which effectively prevent and manage infections caused by pathogenic bacteria in tea plants[17]. Kielak et al. found that Acidobacterium contributes to the synthesis of the plant growth hormone indole-3-acetic acid (IAA) and facilitates iron uptake, thus functioning as a plant growth-promoting bacterium that stimulates plant–bacteria interactions[18]. Acidobacteria, a pivotal functional microbial group in tea plantation soils, synthesize and secrete low-molecular-weight organic acids via genome-encoded C metabolism enzymes, thereby acidifying the rhizosphere environment[19,20]. The acidified microenvironment and organic acid anions act synergistically to solubilize minerals, release nutrients (P, K, Fe), and chelate free Al3+, which aligns with the Al-tolerant and acidophilic traits of tea plants. By activating the nutrient supply, mediating carbon and N cycling, and regulating signals through secondary metabolites, these acidophilic microbes promote the root development of tea plants to exert a dual growth-promoting effect, thus serving as the core microbial community that sustains nutrient cycling in the plant–soil ecosystem and safeguards tea plants' health. For instance, Bacillus subtilis is a predominant species found in the rhizosphere of tea plants. It can break down complex organic materials such as cellulose and lignin while facilitating the cycling of essential nutrients, including P, K, and Fe, in the soil[21]. Acidobacteria can produce organic acids, reducing soil pH and promoting the dissolution of minerals and nutrient release[22]. Although existing research has clarified the functional roles of tea plants in growth and soil health, the specific molecular mechanisms underlying these plant–microbe interactions remain underexplored in tea plantations. For example, a recent study demonstrated that a synthetic microbial community (isolated from tea roots) can promote NH4+ homeostasis and theanine synthesis in the tea plant[23].
Fungi
-
Alongside bacteria, fungi are essential components of tea plantation ecosystems, with Ascomycota, Basidiomycota, and symbiotic fungi as the primary functional groups[24]. Ascomycetes are widely distributed in tea plantation soils and are also dominant in soils worldwide. The reason for this might be that this type of microbial community has evolved survival mechanisms to adapt to complex, variable soil environments[25]. Penicillium and Trichoderma are the main members of Ascomycetes. Research has demonstrated that this group can produce secondary metabolites (antibacterial substances) that inhibit other microorganisms, thereby protecting plants from infection by pathogens[26]. Moreover, tea plantations produce a large amount of pruned leaves and litter every year. These biological residues provide favorable living conditions for Basidiomycetes. For instance, Saitozyma, Russula, and Hygrocybe mostly feed on the surface of lignin-rich residual substances. They act as decomposers, degrading pruned leaves and litter, providing C sources and other nutrients for the growth of tea plants[24,27]. There are also many mutualistic symbionts, such as arbuscular mycorrhizal fungi (AMF), distributed in the rhizosphere soil of tea plants. Plant roots and symbiotic fungi establish a close relationship through a series of complex signaling and mycelial interactions. Among them, Glomus and Acaulospora play a vital role in helping plants to synergistically absorb and transport nutrients and metal ions such as Zn and Fe[28]. Furthermore, previous research using an inoculation experiment with the Longjing 43 cultivar found that AMF not only promoted the growth of tea plants but also significantly increased the levels of flavonoids, anthocyanins, and free amino acids in tea, revealing the prominent role of fungi in improving tea quality[29].
Other microbes
-
Additionally, protozoa and viruses also play vital roles in the microecology of tea plantation soils. The abundance and distribution of protozoa are often linked to the water conditions, aeration, and nutrient levels. These organisms, including amoebas and ciliates, typically feed on bacteria and fungi. Moderate predation helps regulate the excessive growth of certain microorganisms, thereby maintaining the structure and quantity of the soil microbial community in tea plantations. This contributes to the diversity and stability of the microbial community within the tea plantation. Soils are the largest viral reservoirs on Earth, containing 107 to 1010 viruses per gram of soil[30]. They are now considered critical to the soil's element cycling, though they have been neglected for a long time. Though current research on tea plantations has focused on using viruses to develop biological agents that target hosts to control harmful insects such as Ectropis obliqua during tea production[31], research on tea plantation viruses has rarely been reported.
Factors influencing microbial communities in tea plantation soils
-
Many factors can directly or indirectly influence the soil microbiome in tea plantations, given the unique characteristics of tea plantations and tea plants. Previous research has indicated that variations in plant varieties and genotypes can affect the composition of soil microbial communities, including fungi and bacteria. Root exudates, aboveground litter, and secondary metabolites vary among different tea tree cultivars, resulting in distinct microbial communities in the rhizosphere. Studies on the rhizosphere microbiota of four typical tea tree cultivars in southwestern China revealed significant differences in fungal communities across distinct tea garden soils. It is hypothesized that these variations may stem from host-specific root exudates and litter[14]. Concurrently, an investigation of bacterial communities in the rhizosphere across 44 tea cultivars demonstrated substantial variation in bacterial diversity and abundance among cultivars[32]. Beyond varietal differences, the shrubs' age represents another significant factor. Gui et al. conducted a co-occurrence network analysis of the relationship between the soil bacterial community's composition and shrub age across three tea gardens in eastern China, revealing that older tea gardens exhibit tighter and more complex bacterial community associations. Ultimately, these shifts in the microbial community stem from the progressive accumulation of soil carbon and nitrogen over extended cultivation periods as tea plants mature[7].
In terms of tea plantation types, conventional, organic and ecological tea plantations exhibit significant differences in their soil microbial communities, influenced by the soil's physicochemical properties and human management practices. Wu et al. found that the abundance of Gram-positive bacteria (G+) and actinomycetes in organic tea plantations, along with the ratio of Gram-negative to Gram-positive bacteria (G−/G+), were all higher than in conventional tea plantations. This indicates that the distribution of soil microbial communities in organic tea plantations is superior to that in conventional tea plantations[33]. Monoculture tea plantations often exhibit reduced microbial diversity caused by uniform litter inputs and chemical fertilizer applications. In contrast, increased plant diversity in mixed tea gardens promotes the proliferation of functional microorganisms such as AMF and N-fixing bacteria, through litter heterogeneity, root interactions, and regulation of the microclimate. Notably, the Shannon index for fungi increased significantly in organic tea plantations, whereas bacterial abundance remained largely unchanged. At the same time, the most significant change was in the abundance of the Proteobacteria phylum, which plays a key role in converting soil-available N[34]. This phenomenon may provide insights into and inspiration for practices aimed at improving soil nutrient levels in tea plantations by modifying the microbial functional activity groups. Previous studies have shown that bacteria are the dominant microbes in terraced and ancient tea plantations, whereas fungal communities show notable differences. This echoes earlier research which found that both bacterial and fungal communities in the rhizosphere soils of terraced tea plantations showed higher alpha diversity than those in ancient tea gardens. This difference may arise because ancient tea gardens create nutrient-rich ecological niches that also put intense selective pressures on microorganisms[35]. Therefore, the type of tea garden has diverse and complex effects on the soil microbial community's structure.
In addition, natural factors like climate and geography greatly limit tea plantations' production. High temperatures can directly affect the soil's microbial biomass, composition, and abundance[36], and drought limits substrate diffusion and induces physiological stress in microorganisms, reducing microbial biomass and abundance in the soil[37]. When subjected to various temperature and humidity conditions, the microbial communities in tea garden soil experienced notable changes. At 30% and 50% saturated pore space conditions, high temperatures (35 °C) significantly suppressed the abundance of bacteria and archaea, and the expression of ammonia-oxidizing archaea (AOA), ammonia-oxidizing bacteria (AOB), and nitrite reductase genes (nirK) while increasing the copy number of fungal internal transcribed spacer (ITS) genes[38]. Altitude also influences soil microorganisms, with temperature variations resulting from differing elevations significantly impacting the abundance of fungal communities within tea plantations[39]. The effect of elevation on soil microorganisms mainly results from changes in the environmental factors, including shifts in the soil's physicochemical properties such as pH, which indirectly affect the distribution of microbial communities[40]. Although studies have reported on the effects of climate and altitude on soil microorganisms in tea plantations, the underlying mechanisms governing their interactions with microbial communities and functions warrant further investigation.
The enrichment of Al in tea plantation soils, coupled with the extensive use of N fertilizers in production, increasingly displaces Al from the deeper soil layers to the root zone and back to the soil surface. This process exacerbates soil acidification in most tea plantations[2,41]. Currently, over 50% of tea garden soils in China have a pH below 4.5[3]. Soil acidification significantly affects the abundance of acidophilic microorganisms, such as Acidobacteria[42]. Relevant literature shows that soil bacterial diversity peaks at a neutral pH and decreases with deviations from this. This hump-shaped pattern highlights the crucial role of pH in bacteria's abundance, distribution, and diversity[43]. Moreover, favorable soil structures provide diverse habitats for microorganisms. For example, aggregates are relatively stable internally, which protects sensitive microorganisms from environmental fluctuations. However, their surfaces interact with the external environment, enabling microbial exchange via nutrient and moisture exchange in the soil. Fungal hyphae can entangle soil particles to form aggregates that provide attachment sites and habitats for fungi and other microorganisms, influencing their community composition and distribution.
Effect of agronomic practices on the soil microbiome in tea plantations
-
Over recent decades, many studies have revealed the complex relationship between management practices and microbial responses, providing valuable insights for optimizing tea cultivation systems. To achieve substantial tea yields and quality, traditional tea gardens inevitably rely on chemical fertilizers and pesticides in routine management, which adversely affect the soil microorganisms[44]. Gu et al. found that organic fertilization, compared with traditional methods, increased microbial diversity associated with fermentation, nitrate oxidation, N fixation, and chemotrophic activities[45]. This diversity better supports soil fertility and boosts plants' nutrient absorption capacity. The application of organic fertilizers can promote microbial activity and enhance synergistic effects within the soil microbiome, increasing the availability of soil organic matter and nutrients, ultimately boosting plant biomass[46].
Monoculture of the tea plant, combined with the ongoing use of chemical fertilizers and pesticides, often leads to imbalanced soil nutrient levels, decreased soil pH, deterioration of the soil structure, and increased heavy metal contamination[3,47,48]. These issues seriously undermine the stability of soil microbial communities. They may even increase the abundance of certain pathogenic microorganisms, thereby threatening soil health and the sustainable growth of tea plantations. A study examining changes in the soil microbial community over 20 years of continuous tea cultivation found that beneficial microbes like Pseudomonas and Rhodanobacter decreased, whereas pathogenic fungi such as Fusarium became more prevalent[49,50]. In contrast, intercropping systems can effectively reduce the negative effects of continuous cropping on soil microbial communities. If we take the Alnus–tea intercropping system in southwest China as a typical example, this planting approach not only significantly improves N fixation efficiency but also boosts the soil fungal and bacterial biomass by 41% and 10%, respectively, compared with monocropped tea plantations. These changes include the elevated biomass of functional microorganisms, including ectomycorrhizal fungi, G− and G+, and actinomycetes[51]. Furthermore, intercropping leguminous crops in tea plantations can enrich beneficial rhizosphere microbes such as Mycobacterium, Saccharimonadales, and Bacillus, while inhibiting the proliferation of certain denitrifying bacteria[15,52], thereby providing a feasible strategy for addressing N fertilizer dependence and continuous cropping obstacles.
Proper pruning is essential in tea garden management, promoting plant growth, improving quality and yield, and increasing stress resistance. The pruning debris decomposes into soil humus via microbes, adding nutrients for tea plants. These plant-derived C sources (prunings and litter) act as the main substrates for soil microorganisms. Additionally, numerous studies have shown that their presence can affect the microbial community's composition and enhance the soil environment[53,54]. Interestingly, different pruning methods may also be a factor influencing variation in the microbial communities in tea plantations. Jiang et al. found that microbial diversity indices in both the rhizosphere and phyllosphere were higher under umbrella pruning than under flat-top pruning treatment, while the proportion of beneficial fungi increased by 0.9-fold, indicating that different pruning treatments can also affect the microbial community's distribution[55]. Furthermore, improving the soil's texture and physical structure by applying biochar can increase the abundance of the AOB gene and the nirK gene of denitrifying bacteria while decreasing the abundance of the AOA gene. This promotes microbial functions in mitigating the N2O and NO emissions generated by denitrification in acidic soils[56].
-
The vast soil microbial community supports the coordination and functional performance of tea garden ecosystems. This review clarifies specific roles that soil microorganisms play in maintaining the stability of tea garden ecosystems. Soil microorganisms facilitate nutrient cycling within tea gardens, which is essential for plant growth and ecosystem sustainability. They mediate processes such as N fixation, P solubilization, Fe acquisition, Al tolerance, and the decomposition of prunings and litter, thereby promoting the cycling of elements like C, N, and P (Fig. 1). By breaking down organic matter, microorganisms release nutrients that tea plants can absorb, while also engaging in biogeochemical processes that sustain soil fertility during continuous cropping. For instance, N-fixing microorganisms help reduce N limitations in tea plantation soils, which are often acidic and tend to immobilize nutrients, whereas P-solubilizing microorganisms convert insoluble inorganic and organic P into forms that are accessible to tea plants' roots (Fig. 1). Soil microbial activity encourages the formation of soil aggregates, thereby enhancing soil porosity, water retention, and aeration. This stable soil structure, in turn, provides a beneficial habitat for microorganisms and tea plants, establishing a positive feedback loop in the ecosystem (Fig. 1). Soil microorganisms are crucial in improving tea's quality and helping plants withstand stress. They regulate the nutrient supply and soil chemistry, affecting the production of important secondary metabolites in tea leaves such as catechins, amino acids, and volatile compounds, which shape tea's flavor, aroma, and health benefits (Fig. 1). Moreover, they bolster tea plants' resilience against environmental challenges and pests like drought, extreme temperatures, and pathogen infections by triggering systemic immunity and releasing antimicrobial substances (Fig. 1). This stress tolerance is vital for maintaining tea production in the face of increasing climate change and biological threats.
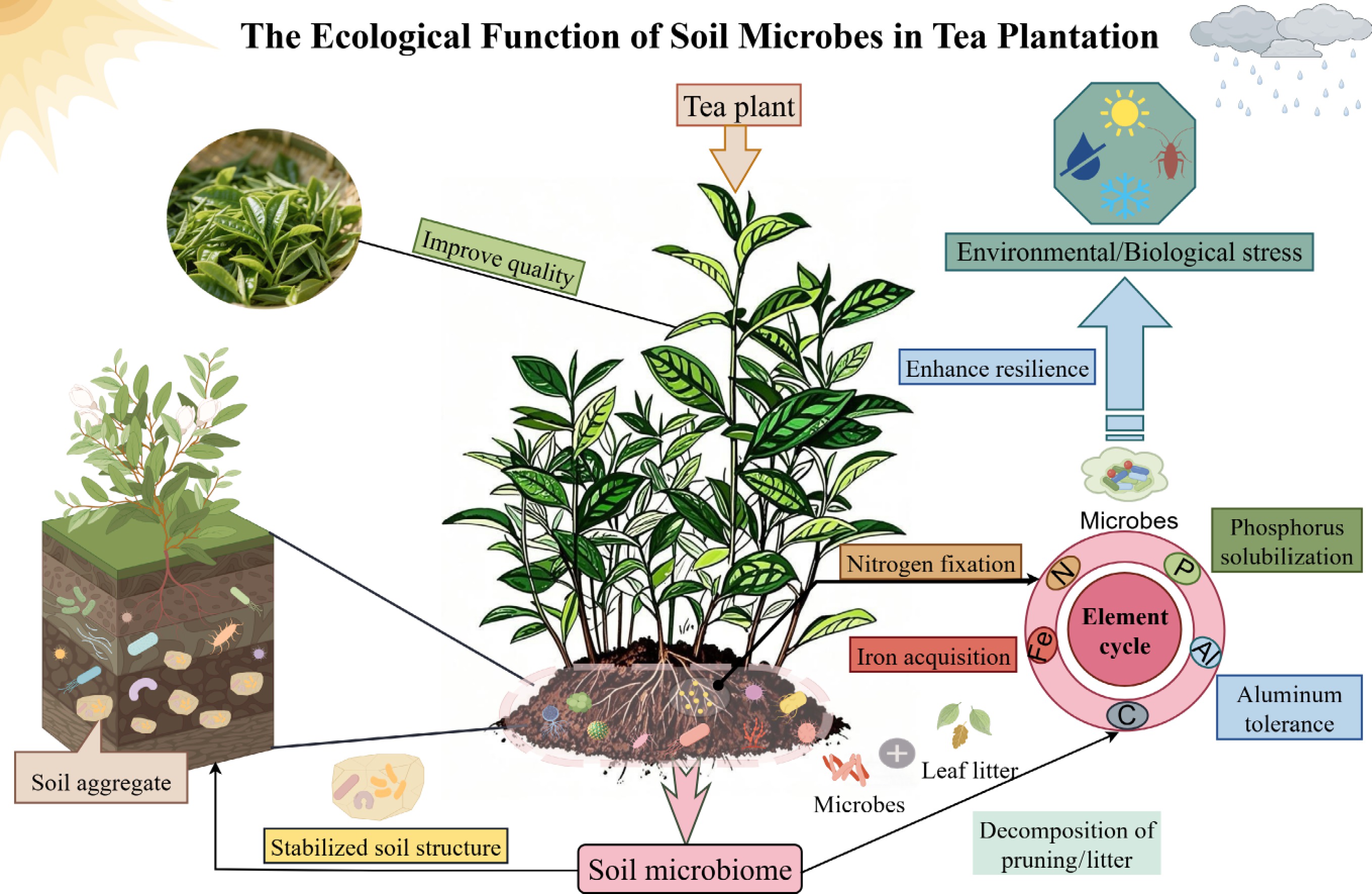
Figure 1.
Ecological function model of soil Microorganisms in tea plantations. Soil microorganisms in tea gardens can enhance tea's quality; they also strengthen tea plants' resistance to environmental and biotic stresses; participate in nutrient cycles such as C, N, P, Fe, and Al; and decompose litter. They also stabilize the soil's structure and form soil aggregates. This figure was drawn with Figdraw (
www.figdraw.com/# ).Effect of the soil microbiome on the qualities of tea
-
It is widely recognized that microorganisms play a crucial role in the development of plants' quality within the agricultural sector, a principle that has also been extensively demonstrated in tea production. The functional microbial communities colonizing plant roots are closely linked to the plants' quality. These microbial populations primarily influence tea plants' growth and quality through four mechanisms. First, soil microorganisms function as nutrient transformers: They fix, dissolve, and convert soil-borne N, P, and K into bioavailable forms, thereby establishing a basis for the the fundamental nutrient supply for tea plants' growth and the development of quality[57]. Second, these microorganisms regulate tea plants' physiological metabolism by promoting the biosynthesis of endogenous growth hormones (e.g., IAA), thereby accelerating the root system's development and enhancing nutrient uptake efficiency. Certain root-associated soil microorganisms isolated from Indian tea plantations were found to effectively secrete IAA, significantly promoting the growth of tea plants[58]. Third, endophytic bacteria and fungi improve the stability of the rhizosphere microenvironment by enhancing the secretion of extracellular polysaccharides (EPS). This process not only stabilizes soil aggregates' structure but also elevates tolerance to abiotic stresses (drought, salinity) and resistance to biotic stresses (pests and diseases)[11]. Last, under stress conditions, certain beneficial microorganisms in the rhizosphere help alleviate stress by producing aminocyclopropane-1-carboxylic acid (ACC) synthase. This enzyme inhibits excessive ethylene production in tea plants, reducing the toxic effects of high ethylene levels on plant cells. ACC deaminase is a key enzyme in environmental stress resistance, inhibiting ethylene synthesis and thereby mitigating the damage caused by external stressors. For instance, microorganisms capable of producing ACC dehydrogenase are present in the soil surrounding tea plants' roots. These microorganisms can cleave the ACC secreted by pathogens, thereby enhancing the stress resistance of tea plants[59].
Amino acids, catechins, polyphenols, flavonoids, caffeine, and volatile compounds often determine the flavor, aroma, and nutritional value of tea. The involvement of microorganisms has been shown to significantly influence the content and metabolism of these compounds, which are crucial to tea's quality[60]. Bag et al. found that inoculating tea plants with AMF can increase the expression of genes encoding amino acid synthases, leading to significantly higher total amino acid content and ultimately increasing the levels of sucrose, soluble glycoproteins, flavonoids, and tea polyphenols compared with untreated controls[8]. Shao et al. also found that Claroideoglomus etunicatum, Diversispora spurca, and Diversispora versiformis significantly increased the levels of metabolites such as amino acids, polyphenols, catechins, and flavonoids in the leaves of Fuding white tea seedlings[61]. The total phenol/amino acid ratio (TP/AA) serves as a crucial indicator for evaluating the quality of summer tea, particularly green tea. A sustained increase in TP/AA leads to a decline in green tea's quality. Interestingly, foliar application of Bacillus amyloliquefaciens significantly reduces TP/AA levels. Moreover, caffeine and catechin levels are lowered considerably, significantly improving the quality of summer tea[62].
Effect of soil microbiome on element cycling
-
Soil microorganisms play a central role in promoting nutrient transformation and the cycling of key elements within tea garden ecosystems. They have evolved intricate networks of interactions and mechanisms that ensure the sustained flow of energy and efficient utilization of elements across all trophic levels.
Previous studies have consistently indicated that the decomposition of organic matter is primarily controlled by microorganisms, influencing terrestrial ecosystems' C, N, and P balances, as well as the nutrients' availability to plants[63]. The sequestration and stabilization of soil organic carbon (SOC) represent crucial processes in the global C cycle. Soil organic matter serves as the largest terrestrial reservoir for C and N and also also acts as a vital source of both macronutrients and micronutrients[64]. Soil microorganisms in tea plantations play a pivotal role in the formation and transformation of SOC. On one hand, photosynthetic autotrophic and chemoautotrophic microorganisms use photosynthesis and chemoautotrophic substances to fix CO2 and synthesize organic matter, thereby incorporating C sources into the soil. On the other hand, substantial plant residues such as litter and pruning debris in tea gardens are decomposed by extracellular enzymes secreted by bacteria and fungi into simple sugars, amino acids, and other small molecules. These are then absorbed and utilized by numerous microorganisms, ultimately releasing C back into the atmosphere as CO2 through respiration[65].
In tea cultivation, large amounts of N fertilizer are often used to enhance leaf quality by increasing the levels of free amino acids. Microorganisms are crucial in N cycling, with different groups performing specific roles. Yang et al. identified Proteobacteria, Actinobacteria, Firmicutes, and Nitrospirae as the dominant phyla associated with nitrogen through high-throughput sequencing, noting variations across soils with different pH levels. Bifidobacterium and Luteimonas were the primary nitrogen-associated bacteria in alkaline soils, whereas Nitrospira and Bdellovibrio dominated in acidic soils[66]. The effect of N-fixing microorganisms in the soil on plant growth has been widely examined. N-fixing bacteria like Rhizobia transform atmospheric N into forms that are accessible to plants, meeting the plants' N needs and supporting the soil N balance[67]. Furthermore, ammonification of nitrate N is vital for maintaining the stability of soil N. In addition to N-fixers, many ammonifying bacteria and Actinomycetes are present in the soil. These microbes decompose N-rich organic materials to produce ammonia under anoxic or anaerobic conditions. They occupy different ecological niches and operate under specific conditions, collectively facilitating the mineralization of soil organic N and stabilizing soil N levels[68]. The pathways by which nitrosating and nitrifying bacteria produce nitrite and nitrate, respectively, and denitrifying bacteria convert nitrate back into N2 constitute key routes of N loss in the soil[69].
Phosphorus is one of the essential nutrients for plants' growth and development. However, its uneven distribution in the soil, diverse forms, and low mobility limit its availability for uptake and utilization by plants[70]. Soil microorganisms play a pivotal role in driving soil P cycling through three primary processes. First, certain soil bacteria and fungi secrete phosphatases to hydrolyze insoluble organic P in the soil, converting it into inorganic forms of P that plants can absorb, a process known as the mineralization of organic P. Research into the improvement of acidic soils has revealed that the addition of exogenous inorganic P stimulates the growth of microorganisms that degrade soluble phosphates, thereby accelerating the mineralization of organic P[71]. The dissolution of inorganic P is achieved through the secretion of organic acids by P-solubilizing bacteria. Microorganisms release the inorganic P chelated by oxides in the soil minerals, whereby the phosphate ions within insoluble phosphates are dissociated and dissolved into the soil solution for uptake by plant roots[72]. When soil contains relatively high levels of available P, microorganisms actively absorb phosphate ions and store them within their cells to meet their metabolic needs. Upon the death of these microorganisms or when they experience nutrient deficiency, the P-containing substances stored within them are either decomposed and utilized by other microorganisms or released back into the soil as inorganic P. This process involves the fixation and release of P. Soil microorganisms can alleviate P limitations by regulating the P-cycle-related community's composition. More than 20 P-solubilizing microbial genera have been identified, with Actinomycetes and certain fungi significantly enhancing the soil-soluble P content[73].
Aluminum is detrimental to most plants, yet tea plants exhibit strong tolerance to Al. It has been reported that low concentrations (0.2–1 mM) of Al can significantly promote root growth, thus making it beneficial and even indispensable for tea plants' development[74,75]. The role of soil microorganisms in Al metabolism is often recognized for enhancing Al tolerance and mitigating Al toxicity in tea plants. Researchers have isolated Al-tolerant microorganisms from tea garden soils. Culturing these microorganisms on Al-containing media effectively reduces Al concentrations, with distinct Al tolerance mechanisms observed across different media types[76]. Iron is crucial for chlorophyll production; a lack of Fe causes yellowing of leaf veins and leaf wilting, disrupting normal plant growth. This is especially important for crops like tea plants, where the leaves are the main harvestable part[77]. Microorganisms primarily absorb and utilize Fe through two mechanisms, secreting organic acids (such as citric acid and oxalic acid) and producing Fe-chelating carriers (siderophores). Microbes secrete low-molecular-weight organic acids such as citric, oxalic, and succinic acids into the rhizosphere, where they acidify the surrounding microenvironment. This process helps desorb adsorbed iron from soil colloids and also reduces insoluble ferric iron (Fe3+) to soluble ferrous iron (Fe2+), which most microbes can use. When Fe is severely limited in the environment, microbes produce siderophores. As low-molecular-weight, high-affinity Fe-chelating secondary metabolites, siderophores are secreted extracellularly and rapidly sequester free Fe3+ in the environment via their high affinity, forming stable water-soluble Fe3+–siderophore complexes. The complexes are recognized by membrane-bound receptors on the microbial cell surface and are transported into the cell via transporters or the proton motive force. Inside the cell, Fe3+ is reduced to Fe2+, dissociated from the complexes, and then utilized by the microbes, and the siderophores can be recycled, secreted again, and reused[78,79].
Effect of the soil microbiome on soil structure
-
The soil structure in natural ecosystems refers to the arrangement and combination of soil particles. It is the product of interactions among biological activity, climate, and soil minerals, exerting profound effects on the soil's fertility, aeration, and water-holding capacity, as well as the growth of plant roots. An important concept in soil structure is soil aggregates. Heterogeneous bonding between organic and mineral particles forms soil aggregates, which serve as the fundamental units of soil structure. On the basis of their particle size, they can be classified into macroaggregates (> 0.25 mm) and microaggregates (< 0.25 mm). Macroaggregates provide space and nutrients for root growth, whereas the formation of microaggregates relies on the mediation of persistent binding microbial substances[80]. EPS acts as a natural binder that binds soil particles of varying sizes, including clay, silt, and sand, forming the foundation for soil with a stable physical structure. EPS directly drives microaggregate formation and serves as a "filler" and "bridge" to fill the voids between aggregates, further reinforcing the soil structure and maintaining its stability, which, in turn, shelters the SOC entrapped within it from rapid decomposition[81]. The mycelia of fungi such as AMF are widely present in the soil. Beyond facilitating nutrient uptake for plants, these fungal networks act as a biological scaffold to form dense interwoven networks that entangle soil particles and aggregates, particularly in the rhizosphere of tea plants. This microbial network structure effectively buffers the soil displacement and fragmentation caused by external forces, stabilizing the spatial arrangement of soil aggregates. This microbial network structure effectively buffers the soil displacement and fragmentation induced by external forces, stabilizing the spatial arrangement of soil aggregates and thus physically protecting SOC from degradation to enhance long-term C sequestration[82]. Zhang et al. emphasized the relationship between the microbial community's composition in aggregates of varying sizes and soil function in their research, advocating for the use of aggregate microbial assembly functions as factors and indicators for investigating the interactions between agricultural soils and microorganisms[81,83]. Some studies have also indicated that the close relationships among biological activity, particularly the presence of microorganisms; the sequestration of SOC; the increase in biopores; and the formation of stable food webs provides researchers with a perspective for understanding the mechanisms by which the soil's natural structure performs its ecological functions[82].
-
The development of microscopy and isolation culture techniques laid the foundation for soil microbiology in the 20th century. Woese et al. (1990) found that ribosomal RNA sequences could serve as molecular clocks for tracing evolution, leading to the three-domain model of life and transforming biodiversity studies[84]. Microbiologists then adopted molecular biology methods, such as 16S rRNA sequencing, to analyze the soil's microbial diversity. Since the early 21st century, new methods like various 16S rRNA-based techniques, stable isotope probing, and environmental metagenomics have emerged, revolutionizing explorations of the soil's microbial diversity and functions. This progress has made soil microbiology a vibrant interdisciplinary field combining soil science, ecology, microbiology, and environmental science. Today, research offers valuable insights and tools to understand biogeochemical cycles, promote sustainable soil use, assess the impacts of global change, and develop bioremediation strategies.
Over recent years, three major emerging technologies have driven advances in soil microbiology research, namely soil microbiome omics technologies, in situ characterization technologies for soil microbiomes, and soil microbiome resource regulation technologies[85]. Soil microbiome omics includes microbial single-molecule and single-cell techniques, molecular markers, genomics, transcriptomics, proteomics, metabolomics, lipidomics, and their coupling mechanisms. It also covers functional genomics, applications in diversity research, and integrated platforms combining physical, chemical, and microbiological data based on the soil's attributes. In situ characterization uses single-cell screening for soil microbes, including isotope-traced markers, modern mass spectrometry, and imaging for continuous monitoring of microbial functions and ecological processes across scales and field network platforms for research under natural and controlled conditions. Soil microbiome resource regulation technologies include comparative studies of modern molecular and traditional methods, high-throughput screening, and functional analyses of soil microorganisms and their bioactive compounds. They also involve preserving, utilizing, and evaluating microbial resources; developing microbial databases and standards; and creating synthetic microbiomes for healthcare, industry, and agriculture.
However, the current research on tea plantation soil microorganisms remains confined mainly to using second-generation sequencing technologies for 16S or ITS sequencing to characterize the bacterial and fungal community's composition, with only a limited number of studies using metagenomic sequencing to capture the microorganisms driving elemental cycling in tea plantation soils. Compared with other ecosystems such as paddy fields, grasslands, and forests, advanced approaches and techniques, including high-throughput microbial screening, single-cell selection, and multiomics integration, have not yet been applied to tea plantation soils. The absence of these cutting-edge technologies in tea plantation research has substantially constrained our understanding of the soil's ecological functions in tea ecosystems and, more critically, has limited our capacity to harness microbial-based strategies to promote the green, efficient, and sustainable development of the tea industry. This knowledge gap can be primarily attributed to the fact that a significant proportion of researchers in the tea science community have concentrated their efforts on cultivar breeding and tea processing, inadvertently overlooking the crucial roles that soil microorganisms play in determining tea's quality, enhancing plants' resistance, and supporting healthy tea plant growth. Therefore, we hereby appeal for greater research attention to the tea plantation soil microbiome. After all, healthy tea plantation soils represent the fundamental cornerstone for the green and sustainable development of the tea industry. Only by comprehensively understanding and effectively managing the soil microbiome can we unlock the full potential of sustainable tea production systems that balance productivity, quality, and environmental stewardship.
-
Despite progress in understanding the microbiomes of tea plantation soils, critical limitations remain. Focusing on individual microbial groups or isolated functions oversimplifies ecosystem processes. Many studies have examined bacteria or fungi separately or have focused on single biogeochemical processes, missing the complex interactions between functional groups and their collective impact on tea plants' health and productivity[21,23,24,66,74]. Microbial interaction networks, including competition, cooperation, and syntrophy, are still poorly understood in tea plantations. Moreover, the geographic coverage remains notably skewed, with most research concentrated in the major tea-producing regions of China, Japan, and India. Tea-growing regions in Africa, Southeast Asia, and South America remain largely understudied, despite accounting for a substantial proportion of global tea production. This limited geographic scope constrains our ability to develop generalizable principles for managing the tea soil microbiome and hinders the identification of region-specific microbial resources. Additionally, the temporal dynamics of microbial communities are poorly understood because of limited long-term data and snapshot studies. Most research captures only single time points or seasonal samples, missing responses to tea plants' phenology, management, and climate change. The mechanistic links between microbial structure and ecosystem functions are also unclear, with few studies establishing causality despite the observed associations. Thus, this review strongly calls for the establishment of standardized, long-term monitoring networks across these understudied tea-growing regions. These networks should incorporate unified sampling protocols, standardized molecular analysis methods, and synchronized data recording systems to ensure cross-regional comparability of the results. Critically, such standardized long-term monitoring efforts are indispensable for addressing the existing research gaps: They will not only lay the foundational groundwork for developing generalizable principles for managing the tea soil microbiome that can be tailored to diverse climatic and edaphic conditions but will also facilitate the systematic identification and characterization of region-specific microbial resources.
Furthermore, as we mentioned above, technical limitations pose significant challenges for the study of the tea plantation soil microbiome. Current methods, predominantly relying on 16S rRNA and ITS amplicon sequencing, often fail to capture the full spectrum of microbial diversity. Rare species, which may serve as keystone taxa or harbor critical functional genes, are frequently overlooked because of sequencing depth limitations. Most studies have focused on prokaryotes and fungi while neglecting protists, viruses, and phage-bacterial networks that regulate the microbial population's dynamics. The inability to culture the vast majority (> 90%) of soil microorganisms further limits functional characterization and biotechnological applications.
Future directions
-
The future of tea plantation soil microbiology should focus on integrated multiomics approaches like metagenomics, metatranscriptomics, metaproteomics, and metabolomics to link community structure with ecosystem function. Metagenomics shows microbial potential, whereas metatranscriptomics and metaproteomics reveal active genes and proteins. Metabolomics studies the small molecules influencing plant–microbe interactions and tea's quality. Promising areas include single-cell genomics and cultivation-independent screening, using techniques like Raman-activated cell sorting and fluorescence-activated cell sorting combined with microfluidics, and mini-metagenomics to isolate uncultured microbes. Moreover, co-occurrence analysis should be experimentally validated using synthetic microbial communities to study specific interactions, such as cross-feeding and competition, before being confirmed in soil systems.
Quality-related research presents one of the most crucial frontiers. Preliminary evidence suggests that tea flavor compounds may be influenced, directly or indirectly, by soil microbial activity, yet the specific mechanisms remain largely unknown[86]. Future research linking soil microbiomes to the quality determinants of tea must emphasize a paradigm shift beyond mere correlational analyses. Specifically, it should systematically elucidate the regulatory mechanisms through which soil microorganisms modulate the biosynthesis of key tea quality-related compounds including L-theanine (a primary contributor to umami taste), polyphenols, and catechins (which confer antioxidant properties and astringency), volatile organic compounds (VOCs, shaping tea's aroma profiles), and the TP/AA. To this end, the mandatory integration of advanced analytical chemistry techniques (e.g., gas chromatography–mass spectrometry, liquid chromatography–mass spectrometry, and nuclear magnetic resonance spectroscopy) with microbiome profiling is imperative. When complemented with stable isotope labeling experiments using 13C and 15N, this combined approach should enable precise tracing of C and N fluxes from soil microbes through the plant to the tea quality compounds. Collectively, these strategies will facilitate the establishment of robust causal relationships between soil microbial communities and tea quality traits, thereby providing direct scientific evidence for microbial contributions and actionable insights to guide the targeted enhancement of tea's quality. In addition, synthetic biology and microbiome engineering offer transformative potential for developing more effective biofertilizers and biocontrol agents, ultimately improving tea's quality[87].
In addition, the development of sustainable, microbiome-informed management strategies is the most pressing practical challenge. This requires moving beyond descriptive studies toward intervention-based research that tests specific management practices for enhancing beneficial microbial functions while suppressing pathogens and nutrient losses. Priority areas include optimizing organic amendment applications to maximize beneficial microbial activity and soil C sequestration; developing region-specific microbial inoculants tailored to specific tea varieties, soil types, and climatic conditions; designing intercropping and agroforestry systems that enhance ecosystem services; and integrating soil microbiome monitoring into decision-support systems for precision agriculture.
The core functions of the soil microbiome are highly vulnerable, as their stability depends heavily on climatic and environmental homeostasis and is easily disrupted by climate perturbations, a phenomenon particularly pronounced in tea plantations. Climate adaptation is pivotal to sustaining the functional integrity of the microbiome and ensuring the sustainability of tea cultivation. Climate models project substantial shifts in temperature and precipitation across major tea-producing regions, with potential alterations of 20%–40% in the composition of microbial communities in subtropical areas[88], which directly impairs their core functionalities. Beneficial microbial taxa are stress-sensitive, and a decline in their abundance can exacerbate soil degradation and compromise the stress resistance of tea plants.
Given this vulnerability, enhancing climate adaptation mechanisms is imperative. Research priorities include clarifying the impacts of climate change on microbial functions, identifying vulnerable taxa/functions, and validating microbial engineering of resilience in tea plants. Preserving climate-resilient microbes in germplasm banks through targeted management (e.g., adaptive fertilization) would aids their colonization of tea plantations. Integrating insights into the microbiome's vulnerability with adaptation mechanisms is key to mitigating climate risks and safeguarding tea ecosystems' resilience.
-
The future of tea plantation soil microbiology lies in transforming from a descriptive to a predictive and ultimately a prescriptive science. This transition requires a mechanistic understanding to establish the causal relationships between microbial characteristics and ecosystem outcomes, predictive modeling that accurately forecasts the microbial community's responses to enable proactive management, prescriptive applications translating fundamental knowledge into actionable recommendations, and a sustainability focus ensuring that microbiome-based strategies enhance not only productivity and quality but also the soil's long-term, biodiversity conservation, and climate change mitigation.
By addressing the current limitations and pursuing the research priorities mentioned in this review, the field can unlock the full potential of soil microbiomes to support a thriving, sustainable tea industry that balances productivity, quality, and environmental stewardship. The challenges are substantial, but so are the opportunities to revolutionize our understanding and management of these critical belowground ecosystems that ultimately determine the character and sustainability of one of the most cherished beverages.
This work was supported by the National Natural Science Foundation of China (32402632), the Jiangsu Province Young Scientific and Technological Talents Promotion Plan (Grant No. JSTJ-2025-481), the Jiangsu Key-technology Integration and Extension Project for Modern Agriculture (Grant No. JCTG[2025]11), Jiangsu Academy of Agricultural Sciences Scientific Research Fund Project (Grant No. KYJJ(25)0715), and the Jiangsu Modern Seed Industry Revitalization Project (Grant No. JBGS[2021]085).
-
The authors confirm their contributions to the paper as follows: draft manuscript writing and revision: Fang X, Ji L; manuscript review: Zhu C, Chen S, Ni K, Wang Y, Ren H, Yang Y. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing does not apply to this article, as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xiwen Fang, Lingfei Ji
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Fang X, Ji L, Zhu C, Chen S, Ni K, et al. 2026. Understanding and managing soil microorganisms for sustainable tea production: a review of current knowledge and future perspectives. Beverage Plant Research 6: e010 doi: 10.48130/bpr-0026-0005 

Understanding and managing soil microorganisms for sustainable tea production: a review of current knowledge and future perspectives
- Received: 06 December 2025
- Revised: 12 January 2026
- Accepted: 23 January 2026
- Published online: 13 March 2026
Abstract: Tea is one of the most widely consumed beverages globally, and its quality and yield are intrinsically linked to soil health, particularly the soil microbiome. As a perennial woody crop, tea plantations develop unique soil characteristics, including natural acidification and high aluminum content, resulting from long-term cultivation and fertilization practices. Soil microorganisms are vital components of tea plantation ecosystems, contributing significantly to soil health, plant growth, and nutrient cycling. However, comprehensive research on how microorganisms affect the plants' physiological activities and the ecosystem's stability remains insufficient. This review examines the current understanding of soil microbial communities in tea plantations, focusing on diversity patterns of bacteria, fungi, and other microbes, and factors influencing these communities, including tea varieties, plantation types, climate, geography, soil properties, and agricultural management practices. We systematically review the ecological functions of soil microorganisms, emphasizing their roles in improving tea quality through transforming nutrients, regulating secondary metabolites, facilitating element cycling, and stabilizing the soil structure. The review discusses research methods and technological advances, highlighting the transition from traditional sequencing to multiomics technologies. We identify current challenges, including our limited understanding of microbial interaction networks, geographic coverage gaps, and technical limitations. Future directions focus on integrated multiomics approaches, quality-related research linking microbiomes to tea flavor compounds, sustainable microbiome-informed management strategies, and adaptation to climate change. This comprehensive review aims to promote sustainable tea production by leveraging soil microorganisms and provides valuable insights for formulating management strategies that balance productivity, quality, and environmental stewardship.