









-
Domestication represents the earliest form of plant breeding and is a pivotal technological milestone in human history. During domestication, humans cultivated and improved wild plants, promoting reliable harvests that met their needs. Since the rise of agricultural civilizations around 10,000 years ago, numerous plant species have been domesticated from their wild ancestor, including legumes, rice, wheat, and corn—crops that now sustain billions of people globally. Traditionally, the process of domestication was extremely slow, often taking over a millennium, or even several millennia, to fix a single trait in a founder population[1]. This slow pace was largely due to the lack of effective tools (such as CRISPR-Cas9, high-throughput phenotyping, marker-assisted selection, and genomic selection), and the limited seed dispersal of available germplasm by ancient farmers[2]. However, current rapid advances in biotechnology have made modern breeding much more efficient, often requiring only a few decades to fix an undesirable trait in a population. To expedite the de novo domestication of wild plants and improve existing crops, it is essential for plant geneticists and breeders to understand the genetic principles underlying specific domestication traits, and to trace the crucial molecular changes that have occurred during the domestication process.
Seeds are crucial not only as a primary source of nutrition, but also for propagating food crops[3]. With the global population steadily increasing, the demand for food crops is expected to rise considerably. However, seed production faces multiple challenges due to environmental changes, including global temperature rise, soil salinization, desertification, and extreme weather events, all of which threaten food security. These changes, many of which are unpredictable, are likely to dynamically alter the breeding objectives of crop breeders, and impact the domestication of new seed crops.
Cultivated soybean (Glycine max L. Merr.) is one of the most economically significant leguminous seed crops, providing a rich source of protein, essential amino acids, oil, and metabolizable energy. The seeds of the cultivated soybean typically contain approximately 40% protein, 20% oil, 35% carbohydrates, and 5% ash, in addition to other important components such as amino acids, macronutrients, micronutrients, and sugars[4,5]. Dietary protein and vegetable oil provide the two main economic benefits of soybean, contributing up to 69% and 30% of the world's food and animal feed, respectively (
www.usda.gov ). Cultivated soybean was domesticated from its wild ancestor (G. soja Sieb. & Zucc.) in China approximately 6,000–9,000 years ago[6]. During soybean domestication, various traits were selected for, including the loss of seed shattering, reduced seed oil content, and increased seed size. Accordingly, G. soja has been used to study the nature of domesticated seed traits in G. max[7−10].Since the release of the genomes for the soybean cultivar Williams 82[11], and the wild soybean variety IT182932[12], research into the evolutionary history of domesticated soybeans has advanced rapidly. Numerous nucleotide sequence differences between wild and cultivated soybeans have been identified[12,13]. In recent years, advances in resequencing and next-generation sequencing have significantly enhanced soybean population genetics. More than 500 selective regions related to domestication have been identified[7,14−16]. These recent domestication-selective strategies now provide a valuable resource for characterizing new genes involved in soybean domestication.
To further identify and characterize domestication-related genes, a genome-wide association study (GWAS) analysis was employed to examine the genetic basis of seed traits underlying domestication in soybean, traits such as seed size/weight, and oil and protein contents[5,17−20]. Recently, multi-omics analysis has identified candidate domestication genes associated with desirable soybean seed traits. For instance, the integration of GWAS, expression QTL (eQTL) information, and transcriptome-wide association studies (TWAS), has identified 22 candidate causal genes responsible for preferred seed traits. These include GmRWOS1, which regulates seed weight and oil content[21]. This review highlights recent research progress in understanding the domestication of soybean seed traits such as size, oil content, and protein content, and it focuses on the evolution of these traits during the domestication process (Fig. 1,Table 1). Additionally, we discuss the applications of new technologies in basic domestication research, along with the challenges and prospects for future soybean studies and breeding.
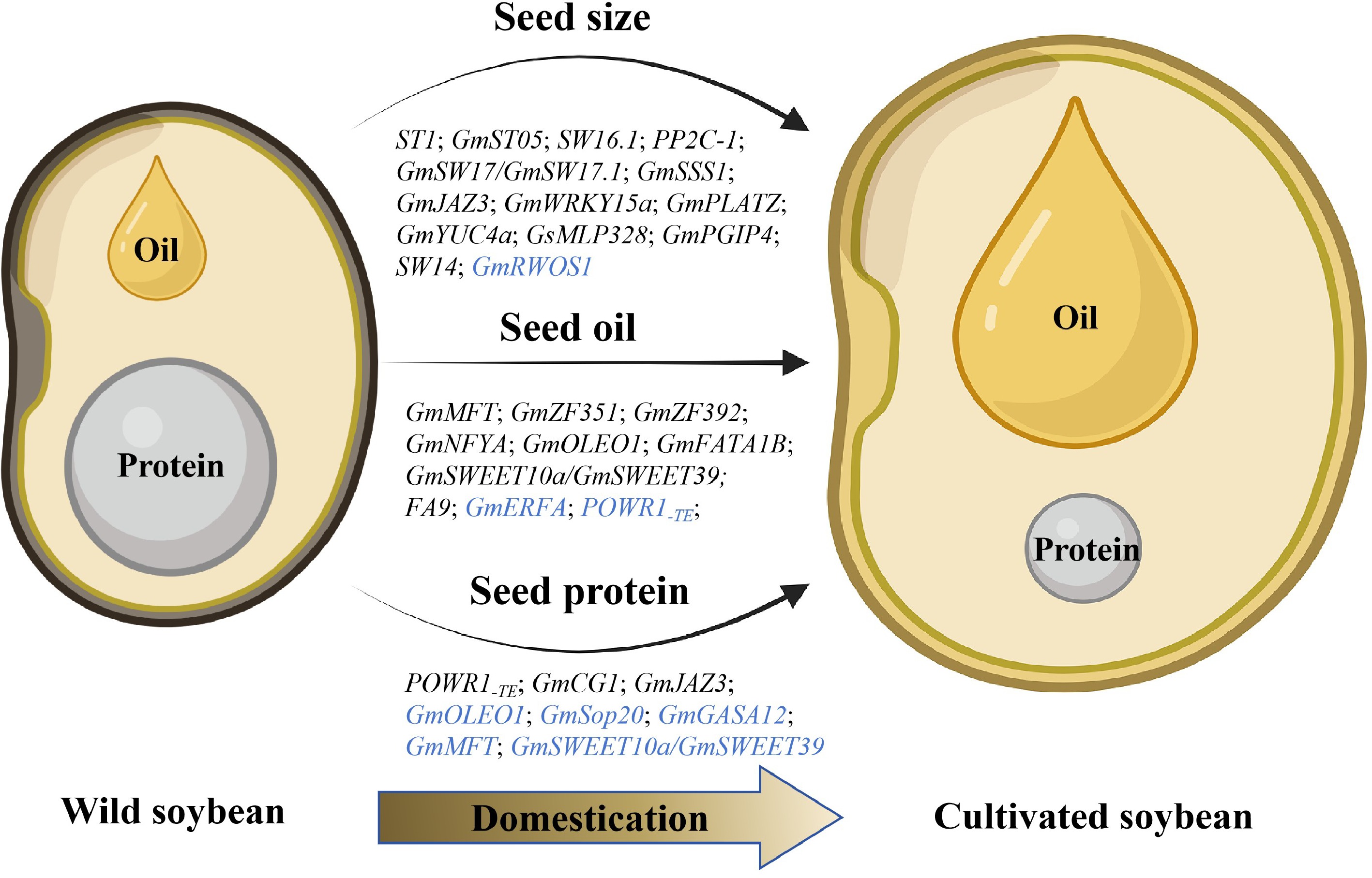
Figure 1.
Domestication genes related to critical seed traits in soybean. Black and blue letters represent positive and negative regulatory genes, respectively.
Table 1. Published domestication genes related to seed traits in soybean.
Trait Name Locus Conserved domain or function Mutant type Ref. Seed size ST1 Glyma.08G109100 UDP-D-glucuronate 4-epimerase SNP in CDS [29] GmST05/GmMFT SoyZH13_05G229200 Mother of TFL1 and FT (MFT) Indel and SNP in promoter
and SNP in CDS[17,50] Seed weight SW16.1 Glyma.16G198300 LIM domain-containing transcription factor Indel in CDS [35] SW14 Glyma.14G010000 Nuclear factor Y subunit SNP in CDS [33] PP2C-1 Glyma17g33690 Putative phosphatase 2C protein Indel and SNPin CDS [36] GmSW17/GmSW17.1 SoyZH13_17G105400/
Glyma.17G109100Ubiquitin-specific protease SNP in CDS [20,37] GmSSS1 Glyma.19G196000 Putative O-GlcNAc transferase SNP in CDS [40] GmJAZ3 Glyma.09G123600 JASMONATE-ZIM DOMAIN (JAZ) Indel and SNPin promoter [44] GmWRKY15a Glyma05g20710 WRKY transcription factor Indel in 5' UTR [45] GmPLATZ Glyma.13G147800 AT-rich sequence and zinc-binding (PLATZ) family protein Indel and SNP in promoter [46] GmRWOS1 Glyma.12G064800 K-stimulated pyrophosphate-energized sodium pump protein Indel and SNP in CDS [21] GsMLP328 Glysoja.20G052861 Soybean major latex protein (MLP) SNP in CDS [42] GmPGIP4 Glyma.08G079200 Polygalacturonase-inhibiting proteins (PGIP) SNP in CDS [42] GmYUC4a Glyma.14G124100 YUCCA (YUC) flavin monooxygenases Indel in 3' UTR and SNP in downstream region [10] Seed oil content and component GmST05/GmMFT SoyZH13_05G229200 Mother of TFL1 and FT (MFT) Indel and SNP in promoter
and SNP in CDS[17,50] GmZF351 Glyma06g44440 Tandem zinc-finger CCCH-domain protein Indel and SNP in promoter [51] GmZF392 Glyma.12G205700 Tandem zinc-finger CCCH-domain protein SNP in promoter [54] GmSWEET39/
GmSWEET10aGlyma.15G049200 Sugars will eventually be exported transporter Indel in CDS [18,19,56,57] GmNFYA Glyma02g47380 Nuclear factor Y subunit Indel in promoter [54] POWR1 Glyma.20G085100 CCT domain Indel in CDS [5] GmOLEO1 Glyma.20G196600 Putative oleosin protein Indel and SNP in promoter [59] GmFATA1B Glyma.08G349200 Acyl-ACP thioesterases (FAT) Indel in CDS [67] FA9 Glyma.09G250400 SEIPIN protein SNP in CDS [68] GmERFA Glyma.02G016100 AP2/ERF-type transcription factor Indel and SNP in promoter [55] Tocopherol content GmMQT/GmST05 Glyma.NDD2.05G269100 Mother of TFL1 and FT (MFT) SNP in promoter [79] GmZF351 Glyma06g44440 Tandem zinc-finger CCCH-domain protein SNP in promoter [76] Seed protein content POWR1 Glyma.20G085100 CCT domain Indel in CDS [5] GmSWEET10a/
GmSWEET39Glyma.15G049200 Sugars will eventually be exported transporter Indel in CDS [18,19,56,57] GmST05/ GmMFT SoyZH13_05G229200 Mother of TFL1 and FT (MFT) Indel and SNP in promoter
and SNP in CDS[17,50] GmOLEO1 Glyma.20G196600 Putative oleosin protein Indel in promoter [59] GmJAZ3 Glyma.09G123600 JASMONATE-ZIM DOMAIN (JAZ) SNP in promoter [44] GmSop20 Glyma.20G005900 C2H2-type zinc-finger
transcription factorIndel in CDS [84] GmCG-1 Glyma.10G246300 α' subunit of β-conglycinin SNP in CDS [88] GmGASA12 Glyma.08G291900 GA-regulated family protein SNP in CDS and promoter [90] -
Seed size is a crucial agronomic trait for domesticated crops. Increasing seed size is a key objective for yield improvement[22]. Therefore, enhancing yield by increasing seed size is a primary breeding objective for plant breeders. G. soja has small seeds compared to G. max[23], and consequently seed size is a key indicator of domestication and can be characterized by three main features: length, width, and thickness[24]. Compared to wild seeds, cultivated soybean seeds are longer, wider, and thicker, with a strong positive correlation between seed size and seed weight. This pleiotropic trait is controlled by numerous genes and environmental factors, making it a crucial selection factor during the domestication process[25]. In recent years, over 400 QTLs associated with soybean seed size have been mapped, and they are found across nearly all 20 of the soybean chromosomes[26−28]. Most of these QTLs are recorded in SoyBase (
www.soybase.org ).Overall, cultivated soybean seeds are rounder than wild soybean seeds. This morphological trait is primarily determined by thickness, assessed by the diameter perpendicular to the hilum. Through GWAS and map-based cloning, the Seed Thickness 1 (ST1) locus on chromosome 8 was identified; it encodes a UDP-D-glucuronate 4-epimerase that influences seed thickness by catalyzing pectin biosynthesis[29]. This locus is also located within a soybean oil content QTL, situated in a ~160-kb 'selective sweep' region shaped by soybean domestication[29−31]. Overexpression of ST1 enhances oil accumulation in seeds by indirectly affecting glycolytic levels, thereby influencing key metabolites in fatty acid (FA) metabolic pathways[29]. Analysis of sequence variation at the ST1 locus among 1,209 accessions revealed that ST1 has undergone selection during soybean domestication[29].
Through a GWAS of 1,853 soybean accessions, a natural allelic variation at GmST05 (Seed Thickness on Chromosome 5) was identified as predominantly controlling seed thickness and size in soybean[17]. GmST05 encodes a protein in the phosphatidylethanolamine-binding protein (PEBP) family that is homologous to MOTHER OF TFL1 AND FT (MFT) in Arabidopsis (Arabidopsis thaliana)[17,32]. Two haplotypes of GmST05 showed significant differences at the transcriptional level, with haplotype I exhibiting a significantly greater seed thickness than haplotype II[17]. The proportion of GmST05HapI to GmST05HapII increased progressively from wild soybean, to landraces, to cultivars, suggesting that GmST05 underwent artificial selection and subsequent local breeding during soybean domestication[17]. Transgenic experiments have demonstrated that GmST05 positively regulated seed size and oil content, while negatively regulating protein content, possibly by regulating the expression of GmSWEET10a (also known as GmSWEET39), which is involved in the transport of sucrose[17].
In addition to seed thickness, seed morphology is determined by seed length and width. Approximately 52 QTLs are related to seed length, and 32 QTLs are related to seed width (
http://soybase.ncgr.org ). Recently, a major QTL, GmSW17 (Seed Width 17), that determines soybean seed width/weight in the natural population, was reported on chromosome 17[20]. Transgenic experiments have demonstrated that GmSW17 positively regulated seed size. GmSW17 encodes a ubiquitin-specific protease orthologous to UBP22 of the ubiquitin-specific protease family. It forms a deubiquitinase module with GmSGF11 and GmENY2 that modulates H2B ubiquitination levels, thereby negatively regulating the expression of GmDP-E2F-1. Although population analysis showed that GmSW17 underwent artificial selection during soybean domestication, its functional alleles have not been fixed in modern breeding. As no other key genes controlling seed length or seed width have been identified to date, further investigation is needed to identify genes determining these two traits, and to pinpoint precisely how seed morphology is regulated in soybean.Regulation of seed weight
-
Seed weight is a crucial trait influencing yield, and it is typically measured by the weight of 100 seeds. Over 300 QTLs associated with soybean seed weight have been identified, scattered across its 20 chromosomes (
www.soybase.org ). On chromosome 14, Nuclear factor Y Subunit A (NFYA) (Glyma.14G010000), which encodes Seed Weight 14 (SW14), a positive regulator of seed weight, was identified through GWAS[33]. SW14 physically interacts with soybean Leafy Cotyledon 1 (GmLEC1), a key regulator of seed development, and subsequently inhibits the formation and transactivating activity of the GmLEC1/GmNF-YC2/GmbZIP67 trimeric complex. A haplotype analysis found that the natural allele, SW14H3, has undergone artificial selection during soybean domestication. A QTL for 100-seed weight, SW16.1, was identified on chromosome 16 using mapping populations derived from chromosome segment substitution lines of wild soybean[34,35]. SW16.1 encodes a transcription factor containing a LIM domain, which contains two zinc fingers, and is involved in protein interactions. It controls 100-seed weight by regulating the transcription of Metallothionein 4 (MT4), which encodes a positive regulator of seed weight[35]. Polymorphism analysis reveals that the frequencies of superior SW16.1 alleles have increased in cultivated soybean compared to wild soybean[35]. On chromosome 17, a QTL for 100-seed weight, phosphatase 2C-1 (PP2C-1), was mapped using a genetically stable population of RILs derived from a cross between wild soybean ZYD7, and cultivated soybean HN44[36]. PP2C-1 promotes cell size in association with GmBZR1, and the PP2C variant ZYD7 from wild soybean contributes to increased seed weight, suggesting a potential role in determining seed weight during soybean domestication[36].GmSW17.1 (also named GmSW17) is another locus on chromosome 17 that is associated with 100-seed weight. It was identified using GWAS and joint linkage mapping[37]. Knockout of GmSW17.1 significantly reduced 100-seed weight in 'Williams 82'. GmSW17.1 encodes a ubiquitin-specific protease that regulates 100-seed weight by interacting with a deubiquitinating module component, GmSGF11, in plant cell nuclei[37,38]. Two natural allelic variants of GmSW17.1 (GmSW17.1T and GmSW17.1C) result in significantly different 100-seed weights, with GmSW17.1T being more frequent in cultivated soybean, aligning with the trend of increasing 100-seed weight during domestication[37].
Association mapping showed that qHSW18-1 (Hundred-Seed Weight 18-1) on chromosome 18 was associated with 100-seed weight, with Glyma.18G242400 emerging as the most promising candidate gene for regulating 100-seed weight in soybean[39]. Haplotype analysis indicated that a superior haplotype of qHSW18-1 underwent intense selection during domestication and subsequent cultivar development[39]. On chromosome 19, Soybean Seed Size 1 (SSS1), which regulates seed weight, was cloned using MutMap analysis using an F2 segregating population, derived from crossing 'Zhong-pin 661' (Zp661) with an sss1 mutant (following EMS mutagenesis of Zp661)[40,41]. Haplotype evolution and molecular function analysis showed that an amino acid variant (Glu-to-Gln) at position 182 in one of its haplotypes, is characteristic of elite alleles selected during soybean domestication[40].
A recent GWAS study by Yang et al.[42] identified GsMLP328 (Major Latex Protein 328), a gene also associated with 100-seed weight, localized on chromosome 20, using a population of 236 wild soybean accessions[42]. GsMLP328 encodes an MLP that interacts with Polygalacturonase-Inhibiting Protein 4 (GmPGIP4), to coordinately regulate seed weight and associated traits, including seed dimensions, protein, and oil content. Two superior haplotypes, Hap_2 and Hap_3, exhibited significantly higher 100-seed weights and seed weights per plant compared to Hap_1. Hap_3 showed a predominant distribution in Asia, especially the Huang-Huai-Hai region of southeast China. However, this haplotype may be in the nascent stage of selection, as its proportion increased only slightly in landrace and cultivated accessions, relative to wild accessions. Moreover, the GmPGIP4 is located in a selection region, and the distribution ratio of its elite haplotype (Hap_2) has been continuously expanding from wild to landrace and cultivated accessions[42].
Transcriptome analysis of differentially expressed genes (DEGs) between wild and cultivated soybeans, combined with gene coexpression network analysis (WGCNA), offers a valuable perspective for exploring domestication genes. Through this approach, GA20OX and NFYA (Glyma02g47380), which are associated with seed weight and oil content, were found to be significantly more highly expressed in cultivated soybeans compared to their wild counterparts[43]. Utilizing gene coexpression network analysis, JASMONATE-ZIM DOMAIN 3 (GmJAZ3) was identified as a key regulator of soybean seed weight and size that promotes cell proliferation[44]. Haplotype analysis of the JAZ3 promoters revealed that Hap3 of JAZ3 underwent selection and fixation during domestication[44]. Using DEGs from RNA-seq data and then mapping these DEGs with QTLs associated with seed size, a candidate regulatory gene, GmWRKY15a, was identified. GmWRKY15a in G. max is significantly more highly expressed than GsWRKY15a in G. soja, and its expression level correlates with seed weight[45]. GmPLATZ, which encodes a Plant AT-rich sequence and zinc-binding (PLATZ) family protein, was identified through the analysis of seed transcriptomes from 45 cultivated soybean varieties and developing seeds at seven stages[46]. As a PLATZ-type transcription factor, GmPLATZ can directly bind to the promoters of cyclin genes and GmGA20OX to regulate their expression and promote cell proliferation[46]. Hap3 of GmPLATZ is an elite allele with elevated promoter activity that is associated with increased seed weight[46].
De novo genome assemblies of nine legume species and pangenome analysis of the genetic variations that impacted seed weight revealed convergent selection in hundreds of genes during legume evolution[10]. Among these, a YUCCA family gene, GmYUCCA4a, located on soybean chromosome 14, was identified as a potential target of convergent selection for increased seed weight in soybean, pigeon pea [Cajanus cajan (L.) Millspaugh], chickpea (Cicer arietinum L.), and pea (Pisum sativum L.). The Hap1 haplotype of GmYUCCA4a exhibited lower seed weights than Hap2 or Hap3, with a frequency of 94.5% in wild soybeans that declined to 4.3% in landraces, and 0.4% in cultivars. Through integrating GWAS, eQTL analysis, and TWAS, a candidate causal gene for regulating seed weight and oil content, Regulator of Weight and Oil of Seed 1 (GmRWOS1), was identified[21]. GmRWOS1 encodes a K+-stimulated pyrophosphate-energized sodium pump that regulates seed weight and oil content. Allelic variation analysis revealed that GmRWOS1 was strongly selected during soybean domestication[21].
-
Due to the high global demand for edible vegetable oil, increasing seed oil content is a crucial breeding goal for soybeans. Soybean breeding has more than doubled the oil content in soybean seeds, with cultivated soybean seeds typically containing about 18%–22% oil, whereas wild soybean seeds have only about 8%–10%[47]. The oil content in soybean seeds is a complex trait regulated by multiple genes. More than 300 QTLs related to oil content have been mapped across all 20 soybean chromosomes (
https://soybase.org )[48].qOil-5-1, a stable QTL associated with soybean oil content, was identified at the end of chromosome 5 using RILs from the cultivars Huachun 2 and Wayao[49]. A combination of fine mapping and GWAS confirmed that GmMFT (also known as GmST05), is the causal gene for this QTL[50]. GmMFT belongs to the PEBP family, and the loss of function of GmMFT causes downregulation of FA biosynthesis, and decreased expression of SWEET genes[50]. Haplotype analysis has revealed that the high seed oil content haplotype of GmMFT was selected during modern soybean breeding[50].
By comparing the transcriptomes of developing seeds from cultivated and wild soybeans, the domestication gene GmZF351, which is associated with seed oil accumulation, has been identified on chromosome 6. GmZF351 is located within the known QTL Seed Oil Plus Protein-2[43,51]. This gene encodes a zinc-finger protein that positively regulates lipid biosynthesis by enhancing the activity of WRINKLED1 (WRI1)[51]. WRI1 is a transcription factor in the APETALA2/ethylene-responsive element-binding protein family that is considered to be a master regulator of seed oil accumulation. It interacts with multiple oil-regulated proteins and enhances the expression of various genes involved in glycolysis and FA biosynthesis[52,53].
Additionally, another tandem Cys3-His (CCCH) zinc-finger protein GmZF392 functions as a positive regulator of lipid accumulation by physically interacting with GmZF351[54]. Domestication analysis of the promoter sequence of GmZF392 indicates that it has undergone selection from wild to cultivated soybeans[54]. Both GmZF392 and GmZF351 are upregulated by GmNFYA, a transcription factor associated with oil content accumulation that is expressed at significantly higher levels in cultivated soybeans compared to wild ones[43,54]. An AP2/ERF-type transcription factor (GmERFA) can inhibit the transcriptional activity of GmNFYA via a physical interaction[55]. There are three haplotypes of the GmERFA promoter (Hap1-3), of which Hap3 is the major haplotype in cultivated soybeans and is associated with a lower GmERFA expression level, higher oil content, and lower protein content. This indicates that GmERFA may have undergone selection during domestication[55].
Association analysis showed that the sugar transporter gene GmSWEET10a is closely linked to seed oil and protein QTLs[18,19,56,57]. GmSWEET10a is highly expressed in soybean seeds[18]. Variations in its promoter and coding regions results in differing oil contents in the seeds of RILs, and the superior alleles have been selectively transferred from G. soja to G. max by breeding[18,19,57]. Additionally, GmSWEET10b, which is functionally redundant with GmSWEET10a, is also undergoing selection in current breeding programs[57,58].
Important QTLs affecting oil content in soybean seeds have also been mapped on chromosome 20 in a region including POWR1 (protein, oil, weight, regulator 1), which encodes a CONSTANS, CO-like, and TOC1 (CCT)-domain protein, playing a major role in oil and protein production[5]. The favorable allele has a transposable element (TE) insertion that truncates the protein's CCT domain, significantly increasing seed oil content, weight, and yield[5]. POWR1 is considered to be a domestication gene, and the TE insertion in POWR1 underwent artificial selection during soybean domestication, resulting in higher seed oil content and weight in cultivated soybeans compared to wild-type soybeans[5]. Through GWAS analysis, an environmentally stable QTL, GqOil20, that is associated with oil content was also identified on chromosome 20. Molecular assays have demonstrated that this QTL contains GmOLEO1, which encodes an oleosin that contributes to oil accumulation in soybean seeds by stabilizing oil bodies within cells[59]. Further analysis revealed that the promoter region of GmOLEO1 is located within artificial selection sites, leading to its higher expression in cultivated soybeans compared to wild soybeans, thereby increasing oil accumulation in seeds of cultivated soybeans[59].
Improvement of oil composition
-
In recent years, an increasing number of studies have focused on the relationship between domestication and seed oil components. Soybean oil primarily consists of five types of fatty acids (FAs): approximately 10% palmitic acid (16:0), 4% stearic acid (18:0), 18% oleic acid (18:1, ω-9), 55% linoleic acid (18:2, ω-6), and 13% α-linolenic acid (18:3, ω-3), with 14% being saturated and 86% unsaturated FAs[48]. Unsaturated FAs are known to have beneficial effects on human health. One important unsaturated FA in determining soybean oil quality is ω-3. On one hand, higher levels can reduce oxidative stability[48]. On the other hand, the World Health Organization and numerous studies recommend that reducing the ω-6/ω-3 ratio to less than 4:1 in modern diets is beneficial to human health[60]. Notably, oil from cultivated soybeans generally has a ω-6 to ω-3 ratio of 6:1 to 7:1[61]. In contrast, wild soybeans have nearly double the ω-3 concentration, and a much lower ω-6/ω-3 ratio of 4:1 compared to cultivated soybeans[61]. Additionally, wild soybeans possess different QTLs controlling ω-3 levels than those found in cultivated soybeans[62]. This suggests that the higher ω-6 to ω-3 FA ratio in cultivated soybeans was selected during domestication. The high ω-3 concentration trait in wild soybeans is governed by a set of FA desaturase (FAD) alleles involved in ω-3 biosynthesis, including FAD3, FAD7, and FAD8[48,62−64]. However, none of these genes were preferentially selected during domestication, despite exhibiting a significantly lower missense mutation frequency than the genome-wide average in domesticated populations[65].
In the soybean genome, over 228 QTLs associated with FA metabolism have been identified across all 20 chromosomes (
www.soybase.org )[30,66]. Recently, a few causal genes within these QTL loci have also been identified. One such gene, GmFATA1B, which encodes an acyl-ACP thioesterase, was pinpointed as a causal gene in the seed oil-associated locus qQil-8-1[49,67]. Heterologous overexpression of GmFATA1B in Arabidopsis led to an increase in oleic acid, and its derivative, linoleic acid, of approximately 27% and 53%, respectively[67]. Polymorphism spectrum analysis revealed that GmFATA1B has undergone strong purifying selection, resulting in a very low frequency of deleterious alleles at this locus[67]. Another gene, GmSEIPIN1A, located within a meta-QTL region for soybean oil content on chromosome 9, known as Fatty Acid 9 (FA9), was identified as a significant contributor to seed FA content[68]. Loss of FA9 function decreases linoleic acid and oil contents, while significantly increasing oleic acid content, indicating that FA9 is a key regulator of the seed FA profile. Haplotype 2 of FA9 is absent in wild soybeans, but present in 13% of landraces and 26% of cultivars, suggesting it may have been selected during post-domestication improvement of soybeans[68].Improvement of tocopherol content
-
Tocopherols, commonly known as vitamin E, play a crucial role in human health due to their unique antioxidant properties. These compounds are composed of four types of fat-soluble molecules: α-, β-, γ-, and δ-tocopherol[69]. Among these, α-tocopherol is the primary contributor to the antioxidant activity of tocopherols, and it offers significant benefits to human cells[70]. Tocopherols reduce lipid peroxidation, thereby enhancing the storage stability of soybean oil[71]. As lipophilic antioxidants, tocopherols are present in all plant species, with soybean seed oil containing a higher tocopherol content than most other oilseed crops[72]. In soybean oil, γ-tocopherol is the most abundant (60%–66% of total tocopherol), followed by δ-tocopherol (24%–29%), α-tocopherol (4%–10%), and β-tocopherol (less than 3%)[73]. Due to their importance in oil quality, understanding the genetic basis of tocopherol content has become a significant focus of recent research.
QTLs related to tocopherol content have been identified on chromosomes 5, 9, 11, and 12 using methods such as mGWAS and BSA-Seq[74−77]. On chromosome 9, the promoter of γ-TMT3, which encodes a tocopherol methyltransferase from a high α-tocopherol cultivar (Keszthelyi Aproszemu Sarga), had higher activity than the promoter from a low α-tocopherol cultivar (Ichihime), likely due to single-nucleotide polymorphisms (SNPs) in cis-regulatory elements[77]. A molecular genetic study of wild soybean accessions with high α-tocopherol levels suggested that novel γ-TMT3 promoter haplotypes could enhance the genetic diversity of α-tocopherol biosynthesis in soybeans[78]. Analysis of a soybean population of over 800 accessions showed that the tocopherol content increased during domestication, with a strong positive correlation observed between tocopherol and oil content[76].
Further analysis revealed that the FA regulatory transcription factor GmZF351 activated the expression of several tocopherol pathway genes, thereby increasing both FA and tocopherol contents in soybean seeds[76]. On chromosome 5, SV (structural variation)-GWAS analysis identified GmMQT (Multiple Quality Traits, also known as GmST05) as affecting important seed traits, including tocopherol and oil content[79]. Twelve structural variations in GmMQT in the 547 accessions examined defined two haplotypes, Hap-Ref and Hap-Alt. Hap-Ref exhibited significantly higher oil content (19.05%) than did Hap-Alt (17.11%), with similar trends for γ-tocopherol[79]. These findings demonstrate that tocopherol content was domesticated along with the domestication of fatty acids.
-
In addition to being a vital oil source, soybeans are a significant source of high-quality protein for both human and animal consumption, contributing over 25% of the global protein supply[80]. Soybean protein is particularly nutritious as it contains all the essential amino acids required by humans. Seeds of cultivated soybean varieties contain 35%–40% protein, making them among the most abundant plant-based protein sources. In contrast, wild soybean seeds typically have about 50% protein[81]. During domestication, an increase in seed oil content was accompanied by a decrease in seed protein content due to a negative genetic correlation[48]. This suggests that the artificial selection of soybeans throughout domestication primarily targeted oil content and yield[82]. In modern soybean breeding, both seed oil and protein content are key objectives. Therefore, understanding the genetic mechanisms that determine soybean oil and protein content, and any potential tradeoffs, is crucial for breeding efforts.
To date, 241 QTLs related to protein content in soybean have been identified, with 16 confirmed (
www.soybase.org ). Two major QTLs for protein and oil content were discovered on chromosomes 15 and 20 using RFLP markers in an F2 population derived from a cross between G. max (A81-356022), and G. soja (PI 468916)[83]. POWR1, another key domestication-related gene associated with soybean protein content, is a pleiotropic regulator that influences both seed weight and oil content[5]. Besides POWR1, several domestication genes involved in soybean seed development have been identified. These genes have pleiotropic effects on seed size, oil, and protein content, often negatively regulating protein content. Notable among them are GmSWEET10a/GmSWEET39[18,68], GmMFT[50], GmOLEO1[59], and GmJAZ3[44]. A recent study integrating GWAS/TWAS has identified GmSop20 (Glyma.20G005900) on chromosome 20 as a key regulator of the oil-to-protein ratio[84]. The domestication-selected allele GmSop20C has undergone intense artificial selection, as revealed by genetic diversity analysis, and is prevalent in cultivars across northern China and the USA[84]. Functional analysis showed that GmSop20 directly activates the expression of GmSWEET10a. This synergizes two artificially selected loci within a unified regulatory network, enhancing sugar distribution from the seed coat to the embryo, and thereby increasing oil accumulation[84]. Although the precise metabolic connections between oil and storage protein synthesis remain unclear, the carbon flux in developing soybean seeds is mainly divided between protein and oil[69,84,85]. Understanding any shifts in the carbon flux favoring oil or protein will help improve soybean varieties.Improvement of protein quality
-
To enhance the quality of soybean seeds, it is crucial not only to increase protein content, but also to improve the efficiency of soybean protein utilization, and optimize seed protein quality. The primary storage proteins in soybeans are 7S and 11S globulins, which constitute 70%–80% of the total seed protein, and greatly influence soybean quality[86]. The 11S globulin is abundant in sulfur-containing amino acids essential for human nutrition, and the 7S globulin is a major allergen often responsible for allergies in humans and animals[87]. Consequently, strategies to enhance soybean seed quality focus on reducing 7S globulin levels and increasing 11S globulin levels. GmCG-1 encodes the α' subunit of β-conglycinin. Suppressing expression of GmCG-1 and its paralogues GmCG-2 and GmCG-3 decreases β-conglycinin content while also increasing the 11S/7S ratio, total protein content, and sulfur-containing amino acid content, thereby enhancing the nutritional profile of soybean seeds. Population evolution analysis indicates that GmCG-1, GmCG-2, and GmCG-3 were selected during soybean domestication, and the frequency of the superior Hap1 haplotype of GmCG-1 has gradually declined[88].
A recent study has demonstrated that High Seed Storage Protein (HSSP1), a B3 domain protein increases seed storage protein content by directly binding to the cis-acting element of GmCG1, upregulating its expression[89]. Selecting for larger seed size and higher oil content in soybeans may result in slightly less desirable seed protein traits. Another study demonstrated that Glycine max gibberellic acid-stimulated Arabidopsis 12 (GmGASA12), which encodes a gibberellin-regulated protein, cooperatively regulates the biosynthesis of β-conglycinin and glycinin by interacting with GmCG6[90]. Evolutionary analyses have shown that elite GmGASA12 haplotypes were strongly favored during domestication, with 94% of cultivars possessing beneficial alleles[90].
-
Because several domesticated traits of soybean seeds are influenced by multiple genes[16], comparative genomics analysis related to the seed's morphological index is a promising approach for identifying domestication genes[10]. With the advancement of sequencing technology and the growing availability of population data, numerous QTLs controlling seed traits in soybeans have been identified within the domestication region of the soybean genome[16]. However, only a few key genes have been isolated and functionally validated.
Integrating phenomics and multi-omics to uncover domestication genes in soybean seeds
-
A primary barrier to gene isolation and characterization is that most seed traits are significantly influenced by the growing environment. This makes it challenging to collect reproducible seed phenotypic data from various geographic areas and time periods, hindering the fine mapping of target traits. However, recent advances in seed phenomics, such as high-throughput phenotyping analysis systems, have greatly enhanced the accuracy and reliability of seed trait measurements. The high-throughput of these systems also facilitates the analysis of more crosses and replicates, in more diverse environments[91,92]. Various high-throughput image analysis and spectroscopic tools have been developed for seed phenology[93]. These systems also provide precise and detailed seed phenotypic information for individual lines and large mapping populations, potentially identifying correlations between genomic variation and phenotypic information in seed traits. Notably, the development of multispectral imaging, which offers a nondestructive physical technique, is a valuable tool for evaluating seed quality traits such as vigor, moisture, and germination status[94−96]. These advances are likely to reveal additional potential domestication genes and identify domesticated seed traits.
Genomic approaches have provided valuable insights into the domestication of seed traits in soybeans, and the advent of -omics technologies have transformed traditional methods. These technologies integrate information from various biological levels, including transcriptomics, proteomics, metabolomics, epigenomics, and even single-cell and tissue spatial transcriptomics[21,97]. The multidimensional and dynamic nature of seed -omics approaches has not only accelerated the identification of domesticated genes, but also opened up new opportunities for crop improvement. For instance, three-dimensional genome architecture (Hi-C) epigenomic technology, when combined with GWAS and long-read sequencing, has been used to construct a high-quality chromosome-scale reference genome for oilseed Camellia (Camellia oleifera)[98,99]. This research demonstrated that the artificial selection of elite alleles involved in oil biosynthesis significantly contributed to the domestication of oilseed Camellia[98].
Furthermore, a spatially resolved single-cell atlas of gene expression and chromatin accessibility in developing soybean seeds has led to the identification of several genes specifically expressed in the endosperm or embryo, including SWEET10a, a domesticated gene that regulates soybean seed size and oil content[100]. In the future, combining -omics technologies to deepen our understanding of seed trait domestication throughout seed development will become increasingly valuable. These innovative -omics technologies have significantly enhanced our comprehension of genotype–phenotype associations during seed domestication, and have provided information that is essential for plant breeding, particularly in modern breeding efforts.
Bottlenecks in exploring key domesticated genes of soybean
-
There are several bottlenecks in exploring key domesticated genes in soybeans during current research. First, most domesticated traits are controlled by multiple genes, which form different pathways, networks, or modules, that collectively regulate one or more traits. For example, GmZF392 physically interacts with GmZF351 to synergistically promote the expression of downstream genes. Furthermore, both GmZF392 and GmZF351 are upregulated by GmNFYA[54]. This complexity limits our ability to identify the central genes controlling specific domesticated traits. Second, many pleiotropic genes have been confirmed to regulate multiple traits during soybean domestication. For instance, GmST05 can regulate seed oil and protein content by affecting GmSWEET10a expression, in addition to influencing seed thickness[17,50]. This complexity complicates phenotypic screening and results in difficult trade-offs regarding which traits are beneficial for breeding objectives during the breeding process. Third, some traits exhibit negative coupling that cannot be resolved through traditional breeding approaches during domestication, such as the relationship between soybean protein and oil content. Breeders and early farmers likely shifted the balance of pleiotropic gene expression through selection pressures to mitigate this negative correlation.
Moreover, due to directional selection at specific genomic regions underlying agronomically important traits, the genome-wide genetic diversity of domesticated crops has been reduced[101]. Approximately half of the genetic diversity of soybean has been lost during domestication, resulting in a genetic bottleneck in current soybean breeding[7]. Meanwhile, deleterious mutations have accumulated during the domestication process, increasing the costs associated with soybean domestication[102]. Therefore, exploring genetic resources from wild soybean varieties, and utilizing modern biotechnologies such as genome editing and mutagenesis techniques to create more desirable allelic variations, could help overcome this genetic bottleneck and reduce the costs of future soybean breeding.
Progress and applications of gene editing technology in soybean improvement
-
In recent years, gene editing technology has rapidly advanced in the field of plant cultivation. The development of CRISPR/Cas9 technology has significantly expanded its range of applications. For instance, the introduction of the Flanking Nicks Prime Editor (FLICK-PE) system facilitates precise genome modification in soybeans, enhancing editing efficiency by over 21.1%[103]. Moreover, efficient genetic transformation methods have gradually been established. Recently, the Cut-Dip-Budding (CDB) gene delivery system, along with developmental regulators (DRs) such as WUSCHEL2 (WUS2) and the gene encoding isopentenyltransferase (IPT), has significantly accelerated soybean tissue culture, reducing the transformation process from months to weeks[104,105]. These advances not only have the potential to accelerate the functional analysis and breeding improvement of soybean domestication genes, but have also been demonstrated to be effective strategies for uncoupling the negative correlation between protein and oil content. For example, mutation in nodule inception gene Rhizobially Induced Cle1a/2a (RIC1a/2a), achieved through a multiplexed CRISPR–Cas9 mutagenesis system, lead to a significant increase in seed protein content without a concomitant decrease in oil content[106]. The soybean invertase inhibitor GmCIF1, which has been shown to reduce protein content without affecting oil content, may serve as a viable strategy for dissociating the negative correlation between protein and oil content when knocked out using CRISPR-Cas9 technology[107]. Furthermore, gene editing combined with AlphaFold prediction can create novel genotypes that do not exist in natural populations, overcoming the limitations of natural variation and addressing genetic bottlenecks[108].
In conclusion, with advancements in agricultural technologies and cropping systems, the focus of breeding efforts has evolved. Breeding objectives now must not only encompass improved yield and seed quality, but also align with current agricultural practices, such as selecting traits suitable for machine harvesting. Consequently, new technologies are swiftly transforming the domestication patterns of crop seeds. In the next stage of seed domestication, there is a pressing need to develop more diverse food systems through genomic editing-based breeding, or introgression breeding with multiple breeding populations. Therefore, we have summarized the key domestication genes regulating major seed traits in soybean (Fig. 1, Table 1). These genes can serve as elite targets for overcoming the bottleneck of low yield and quality in soybean by integrating them into modern breeding.
This work was supported by the National key R&D program of China (2024YFF1000502), the National Natural Science Foundation of China (32322062, 32441057), the Biological Breeding-National Science and Technology Major Project (2024ZD0407802), and the Natural Science Foundation of Shandong Province (ZR2023JQ009).
-
The authors confirm their contributions to the paper as follows: study conception and design: Chen B, Zhang D; data collection: Chen B, Wang C; draft manuscript preparation: Chen B, Wang C, Zhuang Y, Li X, Zhang J, Zhang D; funding: Chen B, Zhuang Y, Li X, Zhang D. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article, as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Baoyin Chen, Congcong Wang
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen B, Wang C, Zhuang Y, Li X, Zhang J, et al. 2026. Advances in understanding domestication-related genes for critical seed traits in soybean. Seed Biology 5: e008 doi: 10.48130/seedbio-0026-0004 

Advances in understanding domestication-related genes for critical seed traits in soybean
- Received: 05 November 2025
- Revised: 21 December 2025
- Accepted: 12 January 2026
- Published online: 27 March 2026
Abstract: Soybeans are a vital seed crop, supplying a substantial amount of protein and oil globally, for both human and animal consumption. Modern soybean (Glycine max L. Merr.) varieties have been developed through long-term domestication and genetic introgression of wild ancestors (Glycine soja Sieb. and Zucc.). This process involved significant alterations in seed traits, including seed morphology, size, and quality. In recent years, quantitative trait loci (QTLs), and key genes associated with seed domestication traits have been identified via analytical methods such as QTL mapping, genome-wide association studies, whole-genome resequencing, and -omics applications. This review summarizes recent research on domestication-related genes that regulate key seed traits, including size, oil content, and protein content. Additionally, we discuss the application of new technologies, such as -omics technologies, to advance research involving domestication-related genes. This work will help us better understand soybean domestication in terms of seed traits and should expedite the development of elite soybean varieties to meet the growing demands for soybean production, and ensure global food security.
-
Key words:
- Soybean /
- Domestication /
- Seed trait /
- Seed size /
- Seed protein /
- Seed oil