









-
Plant diversity is a core attribute of ecosystems, commonly described through measures such as species richness and patterns of community composition[1,2]. It underpins ecosystem functioning and is often linked to the stability of ecological communities[3,4]. As a key component of these ecosystems, plant diversity reflects the overall health and viability of forest communities. Previous research has highlighted the characteristics and influencing factors of diversity in forestry contexts[5,6]. Plantation forests, characterized by extensive distribution, rapid growth, and high yields, constitute a significant segment of terrestrial ecosystems. They are vital for stabilizing plant diversity, recovering and restoring forest ecosystems, providing forest products, and improving ecological conditions[7,8]. However, despite the recognized importance of understory vegetation for nutrient cycling and ecosystem stability, the specific dynamics of understory species diversity and community structure in Quercus acutissima plantations, particularly in relation to stand age development, remain poorly understood. A critical knowledge gap exists regarding how understory vegetation responds to the aging process of oak monocultures.
In structurally layered forests, the understory constitutes a key element of vertical organization and can contribute to the persistence of plantation ecosystems[9]. Understory diversity is closely associated with forest functioning through its contributions to nutrient turnover, carbon (C) inputs, and soil biotic processes, with implications for resilience and productivity[10−12]. Differences in stand attributes, such as canopy cover and litter quantity/quality, can modify near-ground conditions and thereby shift understory species composition[6,13,14]. For example, a study in the Canadian boreal forests found that increased light availability reduces understory plant diversity[15]. In addition, variation in soil physicochemical properties is frequently linked to patterns of understory structure and diversity[9,14,16].
As plantation forests expand, land degradation intensifies, prompting focus on the interplay between understory plants and soil factors[17−19]. In established planted forest ecosystems, changes in stand age alter understory vegetation diversity, soil nutrients, temperature, and light, triggering changes in soil physicochemical properties, such as porosity, organic matter, and soil bulk density[20,21]. For instance, in different-aged Castanopsis hystrix plantation forests, plant diversity and soil physicochemical properties are shown to improve with increasing forest age, particularly rising soil organic C and total phosphorus (P) levels[18,22]. Studies have also indicated strong correlations between understory vegetative diversity and soil pH, hydrolyzed nitrogen (N), and quick-acting potassium (K) content[23]. Research on lemon mallow in the hilly wind-sand area of Northwest China revealed that forest age positively correlates with herbaceous biomass and soil nutrient levels[24]. In uneven-aged Eucalyptus plantations, soil moisture was identified as a primary factor influencing understory plant diversity, alongside soil bulk density and organic matter content[25]. These findings underscore how increasing forest age alters soil physicochemical properties, ultimately impacting understory plant diversity[18,26]. In ecological restoration, variations in soil physicochemical properties are often influenced by geographic location, soil depth, and plant species[27,28]. Therefore, understanding these properties is essential for studying understory plant diversity.
Q. acutissima is a deep-rooted, light-loving tree species known for its drought resistance and ability to improve soil and water conservation, making it a pioneer species in ecological restoration[29,30]. Its seeds, bark, and wood serve as valuable industrial materials, and its extensive canopy provides dense shade, making it suitable for use as a street tree, and for creating windbreaks and fire-prevention forests[31]. The long-term single monoculture forest management of Q. acutissima has led to a continuous decline in soil fertility, which has become the main factor limiting the growth of plantation forests in China, thereby greatly restricting their long-term development.
Despite the ecological importance of Q. acutissima plantation forests in subtropical regions, research mainly focuses on single stand ages for plantation design and silvicultural operations[32], with insufficient evaluation across different stand ages (Fig. 1). From an ecosystem restoration perspective, few investigations have examined the characteristics of understory plant communities and their influential factors across stand ages. Understory plants play crucial roles in nutrient cycling, maintaining soil fertility, and carbon sequestration in subtropical forests[33]. However, how their composition and diversity change across stand ages, and which factors primarily control these patterns, remain poorly understood for Q. acutissima plantations (Fig. 1). Filling this knowledge gap is essential for elucidating the relationships among soil nutrient status, soil chemical properties, and understory diversity, and thereby optimizing sustainable management strategies for Q. acutissima plantations (Fig. 1). To address this gap, we examined Q. acutissima plantations spanning five stand ages (4, 10, 25, 45, and 50 years) to: (1) quantify how understory community attributes vary across development; (2) evaluate the environmental correlates most strongly associated with these patterns; and (3) explore plausible pathways underlying observed diversity shifts.
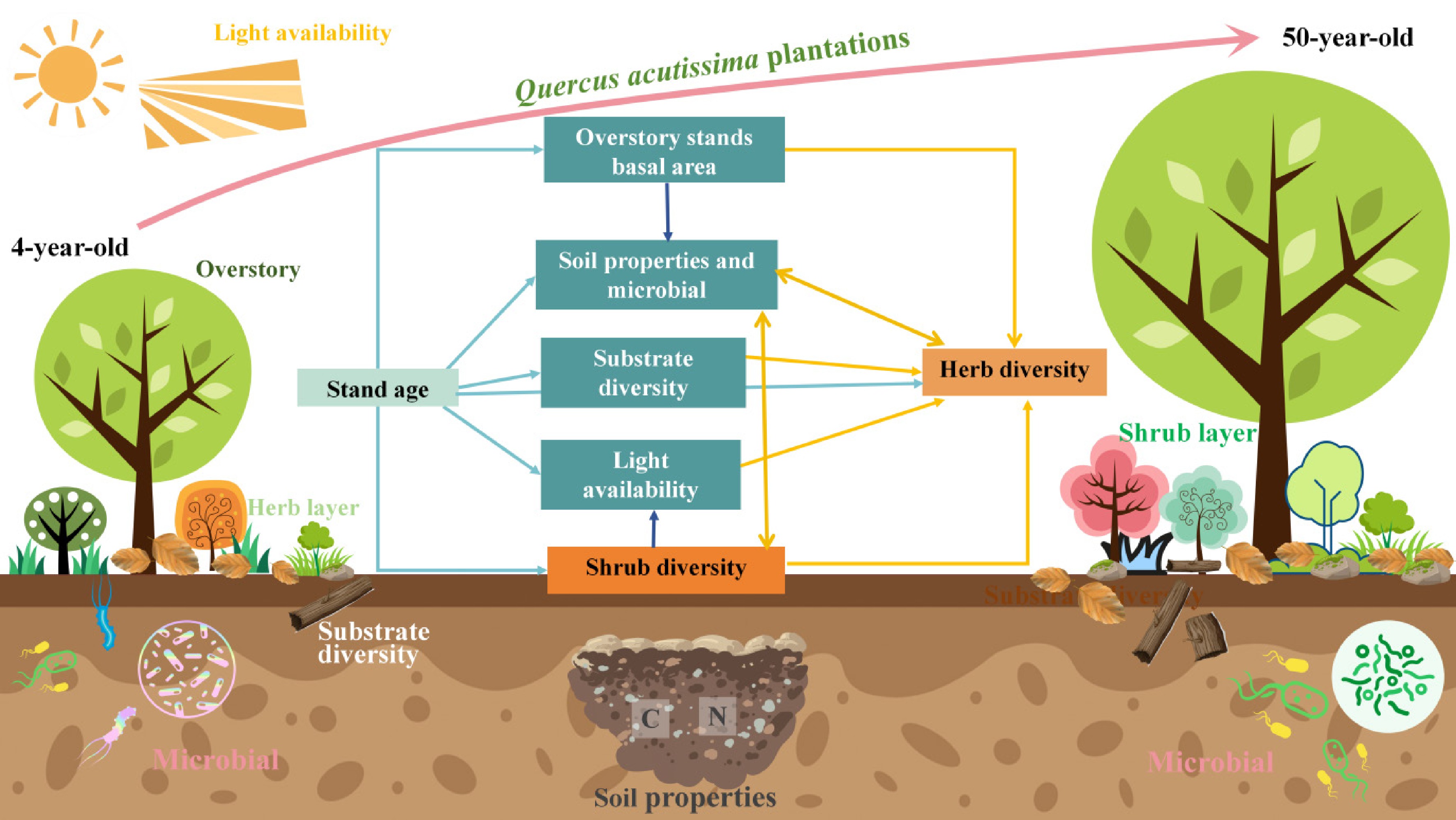
Figure 1.
Conceptual model of hypothesized relationships between the light availability, substrate diversity, and soil conditions on understory vegetation diversity.
Specifically, we hypothesize that: (1) Understory plant community attributes (richness, diversity, and cover) will vary significantly with stand age, exhibiting a successional shift where light-demanding herbs dominate in young forests, and shade-tolerant shrub diversity peaks in mid-successional stages. (2) Patterns of understory diversity and composition will be most strongly associated with environmental gradients, showing positive correlations with soil nutrient availability (specifically soil C, N, and organic matter) and negative correlations with understory light availability (Fig. 1). (3) Shifts in understory diversity will be driven by pathways mediated by stand development (Fig. 1), where maturation enhances soil chemical properties while reducing light availability, thereby filtering the community composition.
-
The study was conducted in the state-owned Huangfu Mountain forest region, Chuzhou, Anhui Province, China (117°58'–118°03' E, 32°17'–32°25' N), situated within a north-south climatic transition zone[34]. The forested landscape extends over 35,515 hm2, including a 1,751 hm2 core protected area, and supports approximately 72% forest cover. The average annual precipitation is about 1,060 mm, and the yearly average temperature ranges from 14 to 15.7 °C, including an extreme minimum of −19 °C, and a maximum of 41.2 °C. The planting density of the artificial stand of Q. acutissima was 2,000 trees·ha−1. Dominant canopy species in the study area include Quercus acutissima, Acer buergerianum, and Dalbergia hupeana. The shrub species Lindera glauca and Rhus chinensis cover the middle forest story, and the major herb species include Persicaria filiformis, Erigeron canadensis, and others. Zonal soils are of the yellow-brown loam type.
Historically, there have been large-scale fires in the area, and forests have been re-established or restored through artificial planting of Q. acutissima seedlings on fire trails and other trails. In the Huangfu Mountain region, the sampled Q. acutissima stands originated from planting events in 1973, 1978, 1998, 2013, and 2019 (Fig. 2). Management practices were broadly comparable among stands: early post-planting tending (e.g., nutrient inputs and site care) was applied and then discontinued after approximately 3–5 years. Before establishment, the sites shared similar land-use and geologic backgrounds.
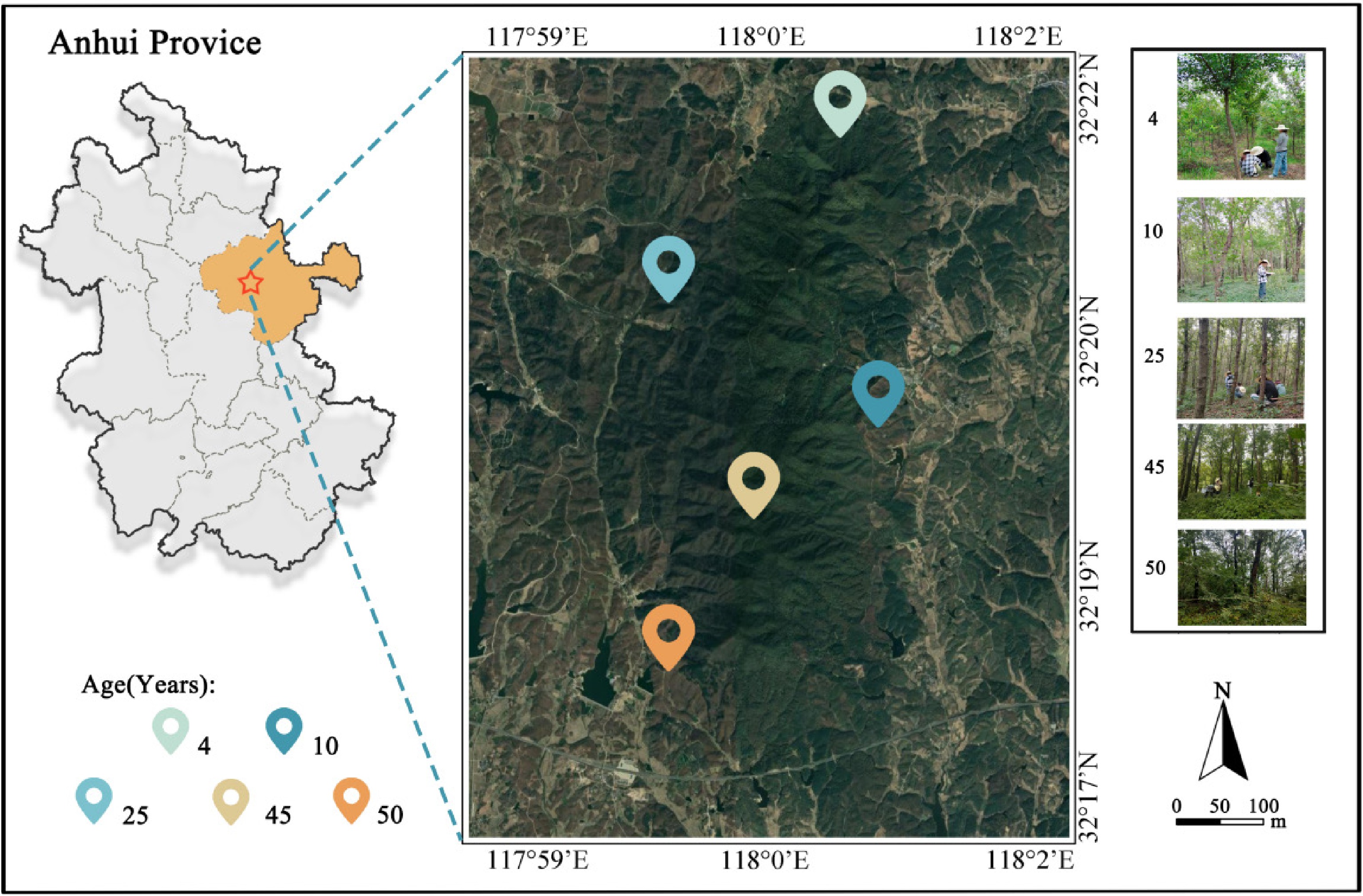
Figure 2.
Geographic context and distribution of sampling plots in Q. acutissima plantations at Huangfu Mountain, Chuzhou, Anhui Province, China. Geographical location of sampling sites in Anhui Province. Base map: Google Earth satellite imagery. Sampling sites were labeled by the authors using Photoshop. Map approval number: GS (2024) 0650.
Sampling plot selection and field survey
-
Sampling plots were selected at sites with similar elevation, slope, and soil texture, and were managed under the same silviculture system across the plantation age groups. We used the details of the plantation of different-aged forests at the Huangfu Mountain Forestry Farm, determined by the field investigation department. Based on the sample plots' data, we used the time-series method to select 4-, 10-, 25-, 45-, and 50-year-old Q. acutissima plantation forests. Since this is a chronosequence study, we inferred successional dynamics by comparing static community composition metrics among different age classes, rather than monitoring changes within individual stands over time. We set three replicates for the five forest ages, resulting in 15 survey areas, illustrated in the map (Fig. 2). For each stand-age class, we established three 20 m × 20 m plots located well within the stand interior, away from edge influences (Table 1). To minimize spatial dependence and selection bias, plots were chosen using a stratified random design. Briefly, we compiled a sampling frame of all candidate stands per age class from historical inventory records (stand ID codes). We then used computer-generated random numbers (Microsoft Excel 2023, and R, v4.2.3) to select stand IDs, and the selected stands were surveyed as sampling sites.
Table 1. Stand and site attributes of the sampledplots in eastern China (values are mean ± SD; n = 3 plots per stand-age class).
Stand age (years) Longitude Latitude Altitude (m) Direction Slope Canopy density Shrubs layer cover (%) Herbs layer cover (%) 4 118°00'00" 32°24'00" 130 South 10 0.38 ± 0.03a 18.56 ± 2.43b 27.23 ± 8.08a 10 118°01'12" 32°19'48" 139 South 12 0.10 ± 0.01b 17.43 ± 4.57b 10.05 ± 1.44b 25 117°59'24" 32°21'36" 166 South 13 0.11 ± 0.01b 13.35 ± 3.58b 13.37 ± 1.90b 45 118°00'36" 32°19'12" 153 South 12 0.11 ± 0.01b 33.17 ± 6.50a 13.46 ± 2.77b 50 117°59'24" 32°18'00" 169 South 11 0.10 ± 0.02b 15.33 ± 5.62b 11.99 ± 3.89b Different letters among stand age means different levels of significance, p < 0.05. Within each stand, we established a 20 m × 20 m plot using a random selection procedure[33,34]. For each plot, we recorded geographic position (latitude/longitude), elevation, slope, and aspect. All trees (height ≥ 4 m) were identified and measured for abundance, height, and diameter at breast height (DBH). Understory vegetation was surveyed within nested subplots: a 5 m × 5 m quadrat for shrubs (1.3 m ≤ height < 4 m), and ten 1 m × 1 m quadrats for herbs (height < 1.3 m). For trees, we recorded species identity, DBH, and height; for shrubs and herbs, we recorded species identity and percent cover[12,13,19]. This data was used to compute diversity metrics.
To characterize the understory light environment, canopy openness was measured at 1.3 m above ground level in each herb quadrat as the fraction of the visible sky not obscured by foliage. Readings were taken in the four cardinal directions (N, S, E, and W) with a spherical densitometer[35,36]. For each plot, mean openness was used as an index of average light availability (LM), and the among-quadrat standard deviation was used to describe within-plot spatial variability in light (LSD). Forest-floor heterogeneity was quantified by visually estimating the proportional cover of major substrate types within each 20 m × 20 m plot (bare rock, litter, exposed soil, uprooted root-soil mounds, and coarse woody debris). We calculated a substrate diversity index (SubD) as the Shannon diversity of substrate cover proportions:
$ {\rm{SubD}} = {-}\sum {(}{{p}}_{{i}}\;\log {{p}}_{{i}}{)} $ where, pi is the relative cover of substrate category (i) within the plot.
Soil sample collection and determination
-
Within each 20 m × 20 m plot, we collected soils from 3–5 locations selected at random after removing the surface litter layer. At each location, a 0–20 cm core was obtained using a ring-core sampler. The material from the sampling points was homogenized and split into two subsamples: one was immediately cooled on dry ice for subsequent microbial analyses, and the second was retained for measurements of soil moisture, pH, bulk density, organic matter (SOM), and total C and N.
Soil water content (SWC) was determined gravimetrically by drying fresh soil at 105 °C to constant mass. For physicochemical analyses, soils were air-dried and passed through a 100-mesh sieve. Soil pH was measured with a pH meter (Mettler Toledo, Shanghai, China) in a 1:2.5 (w/v) soil-water suspension. Total C and N concentrations (mg·g−1) were quantified by flash dry combustion with gas chromatographic separation and thermal conductivity detection (Vario EL Cube, Elementar, Germany)[21,34]. Microbial richness was expressed as the number of observed fungal OTUs, and microbial diversity was characterized following Huang et al.[34].
Data analysis
-
We evaluated stand-age differences in environmental variables and understory diversity using one-way analysis of variance (ANOVA). Diversity was summarized with Shannon's index:
$ {{H}'}={-}\sum \nolimits_{{i=1}}^{{S}}{{p}}_{{i}}\ln {{p}}_{{i}} $ where, pi denotes the relative abundance of species (i) and (S) is species richness. The same index was calculated for the tree layer using overstory species abundances within each plot. When ANOVA indicated significant age effects, pairwise comparisons were conducted with appropriate post hoc tests. Assumptions were checked using Shapiro-Wilk tests for normality, and Bartlett tests for homogeneity of variances.
To test whether community composition varied among stand ages, we used permutational multivariate analysis of variance (PerMANOVA). Differences in composition were visualized with non-metric multidimensional scaling (NMDS). We identified age-associated taxa with indicator species analysis (indicspecies package) using the multipatt function and 999 permutations (α = 0.05); indicator values were partitioned into specificity and fidelity to quantify species-group associations[37]. Community stability was assessed using the Godron approach by ranking species by relative importance, computing cumulative relative importance ([x]) and cumulative species proportion ([y]), and fitting a quadratic model ([y = ax2 + bx + c]). The coordinate ([x, y]) derived from the fitted curve was used as a summary descriptor of dominance structure.
Associations between community composition and environmental variables (overstory attributes, soil properties, and microbial metrics) were evaluated using envfit, in the vegan package. To explore predictor importance, we applied random forest models (randomForest package) that included soil nutrient indices, light availability (LM and LSD), and stand-structure variables; importance scores ranked predictors. Variables consistently identified as influential were carried forward into a structural equation model (SEM, lavaan package) to examine hypothesized pathways linking stand development, environmental conditions, and understory diversity (Fig. 1). All analyses were performed in R (v4.2.3).
-
Using PerMANOVA, the analysis of the distance matrix-based showed significant differences in the species composition of the shrub layer (p = 0.001) and the herb layer (p = 0.001) across the different stand groups (Fig. 3). NMDS spatial ordination showed that species composition and interspecific number ratios differed among age groups. In the shrub layer, species composition and interspecific number varied significantly. The species composition of the shrub layer in the age groups (4, 10, 25, and 45 years old) of the Q. acutissima plantation shows no dependency, whereas the 25- and 50-year-old groups overlap. On the other hand, in the herb-layer, species composition and interspecific numbers diverged considerably across age groups, and no overlap was observed, indicating that the ratio of species composition to species number in the herb-layer varied more than in the shrub layer with increasing plantation age.
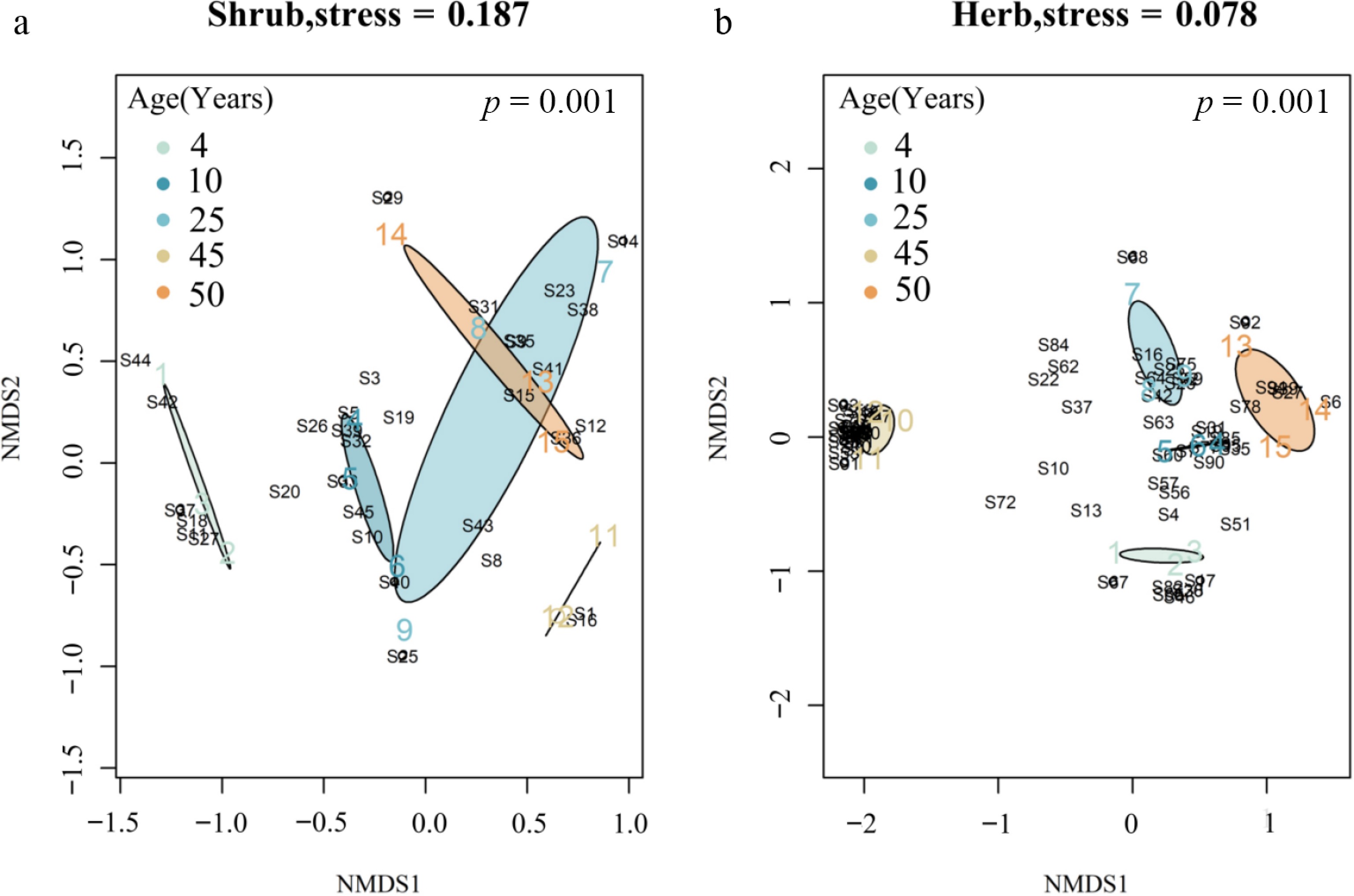
Figure 3.
Non-metric multidimensional scaling (NMDS) ordinations of shrub- and herb-layer species composition across stand-age classes. Each point represents a plot, distances among points reflect differences in community composition (closer points indicate more similar assemblages). Ellipses show the standard error around stand-age centroids, based on weighted-average scores.
Differences were found in the indicator species of Q. acutissima plantations by plantation age. In the shrub layer (4 years), Triadica sebifera (IndVals = 1.000, p < 0.05) and Cymbopogon citratus (IndVals = 1.000, p < 0.05) were warm and humid-loving plant species, while in the herb-layer, indicator species primarily belonged to light demanders named Erigeron annuus (IndVals = 1.000, p < 0.05), Rubus hirsutus (IndVals = 1.000, p < 0.05), Oxalis corniculata (IndVals = 0.904, p < 0.05), and annual herbs named Euphorbia maculata (IndVals = 1.000, p < 0.05), Elsholtzia cyprianii (IndVals = 1.000, p < 0.05), Kummerowia striata (IndVals = 1.000, p < 0.05), the aster family Xanthium strumarium (IndVals = 1.000, p < 0.05), Erigeron sumatrensis (IndVals = 1.000, p < 0.05), and the warm and humid-loving Justicia procumbens (IndVals = 1.000, p < 0.05), Gelsemium elegans (IndVals = 1.000, p < 0.05), and Cuscuta japonica (IndVals = 1.000, p < 0.05). In the 10-year-old plantation, the shrub-layer indicator species mainly belonged to the family Rubiaceae, Paederia foetida (IndVals = 0.943, p < 0.05), and family Anacardiaceae, Rhus chinensis (IndVals = 0.942, p < 0.05), and comprised herb-layer indicator species from the family Aristolochiaceae, Aristolochia debilis (IndVals = 1.000, p < 0.05) (Table 2).
Table 2. Indicator taxa associated with stand-age classes in the shrub and herb layers of Q. acutissima plantations.
Stand age (years) Indicator species Life forms Specificity Sensitivity Indicator value p Shrub layer 4 Triadica sebifera Tree (seedling) 1.0000 1 1.000 0.013 Cymbopogon citratus Herb 1.0000 1 1.000 0.013 10 Paederia foetida Herb 0.8896 1 0.943 0.012 Rhus chinensis Tree (seedling) 0.8879 1 0.942 0.022 50 Maclura tricuspidata Shrub 0.8261 1 0.909 0.010 Ligustrum sinense Shrub 0.8333 1 0.913 0.010 Herb layer 4 Erigeron annuus Herb 1.0000 1 1.000 0.011 Euphorbia maculata Herb 1.0000 1 1.000 0.011 Justicia procumbens Herb 1.0000 1 1.000 0.011 Elsholtzia cyprianii Herb 1.0000 1 1.000 0.011 Gelsemium elegans Herb 1.0000 1 1.000 0.011 Xanthium strumarium Herb 1.0000 1 1.000 0.011 Erigeron sumatrensis Herb 1.0000 1 1.000 0.011 Rubus hirsutus Shrub (seedling) 1.0000 1 1.000 0.011 Cuscuta japonica Herb 1.0000 1 1.000 0.011 Kummerowia striata Herb 1.0000 1 1.000 0.011 Oxalis corniculata Herb 0.8166 1 0.904 0.011 10 Aristolochia debilis Herb 1.0000 1 1.000 0.016 25 Isotrema mollissimum Vine (seedling) 0.9055 1 0.797 0.020 Ophiopogon japonicus Herb 0.6359 1 0.797 0.033 45 Quercus acutissima Tree (seedling) 0.7894 1 0.888 0.014 50 Juncus effusus Herb 1.0000 1 1.000 0.009 Maclura tricuspidata Tree (seedling) 0.8980 1 0.948 0.009 Paederia foetida Herb 0.3746 1 0.612 0.016 Similarly, indicator herbaceous species in 25-year old forests included Isotrema mollissimum (IndVals = 0.797, p < 0.05) and Ophiopogon japonicus (IndVals = 0.797, p < 0.05), both found on loose, fertile soils, while the shrub layer lacked indicator species. In 50-year Q. acutissima plantations, indicator species in the shrub layer were mostly shade-tolerant like Maclura tricuspidata (IndVals = 0.909, p < 0.05), and Ligustrum sinense (IndVals = 0.913, p < 0.05), while the herb-layer comprised of the perennial herbs, Juncus effusus (IndVals = 1.000, p < 0.05) and shade-tolerant Maclura tricuspidata (IndVals = 0.948, p < 0.05), as well as Paederia foetida (IndVals = 0.612, p < 0.05).
Differences in understory cover, species richness, evenness, and Shannon's index in Q. acutissima plantations among stand age groups
-
Our results indicated that plant diversity in the shrub and herb layers of Q. acutissima plantations varied significantly with stand age. The 45-year-old plantation showed robust shrub cover compared to other forest stand ages. Herb cover was significantly higher in the 4-year group than in the other stand groups (Fig. 4). Also, shrub layer richness (p < 0.01), herb layer richness (p < 0.001), shrub layer Shannon's index (p < 0.01), and herb layer Shannon's index (p < 0.01) differed significantly across stand ages. However, the shrub-layer Shannon index and the shrub-layer richness index showed a trend of initially increasing, and then decreasing with forest age, reaching their highest values at 45 years. On the other hand, the highest herb layer Shannon index and herb layer richness index were observed in the 4-year-old plantations, with a decreasing trend with increasing plantation age (Fig. 4).
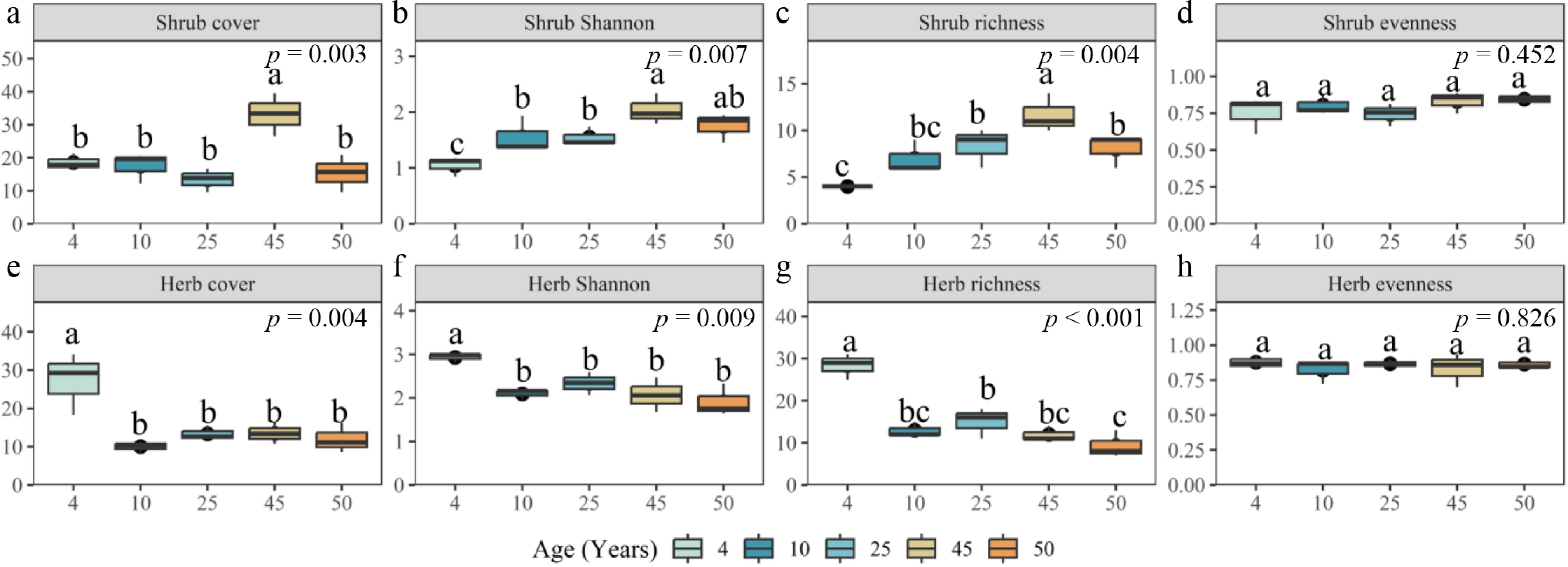
Figure 4.
Effects of stand age on understory vegetation cover and diversity in the shrub and herb layers of Q. acutissima plantations. For each age class, the figure summarizes plot-level estimates (n = 3) of (a), (e) percent cover; (b), (f) Shannon diversity (H′); (c), (g) species richness, and (d), (h) evenness. Data are presented as mean ± standard error. Where group comparisons were significant, different lowercase letters indicate statistically distinguishable age classes based on post hoc tests following one-way ANOVA (α = 0.05).
Community stability in Q. acutissima plantations
-
The Euclidean distance (d) between the modeled intersection (x, y) and the ideal stabilization point (20, 80) varied across stand ages (Fig. 5). Compared to 4 years (d = 10.07) and 45 years (d = 14.76), the vegetation communities of 10 years (d = 19.72) and 20 years (d = 20.00) were found unbalanced, suggesting that early conservation measures destabilized the community. Remarkably, the vegetation community in the 50-year was the most unstable (d = 23.07) in Q. acutissima plantations.
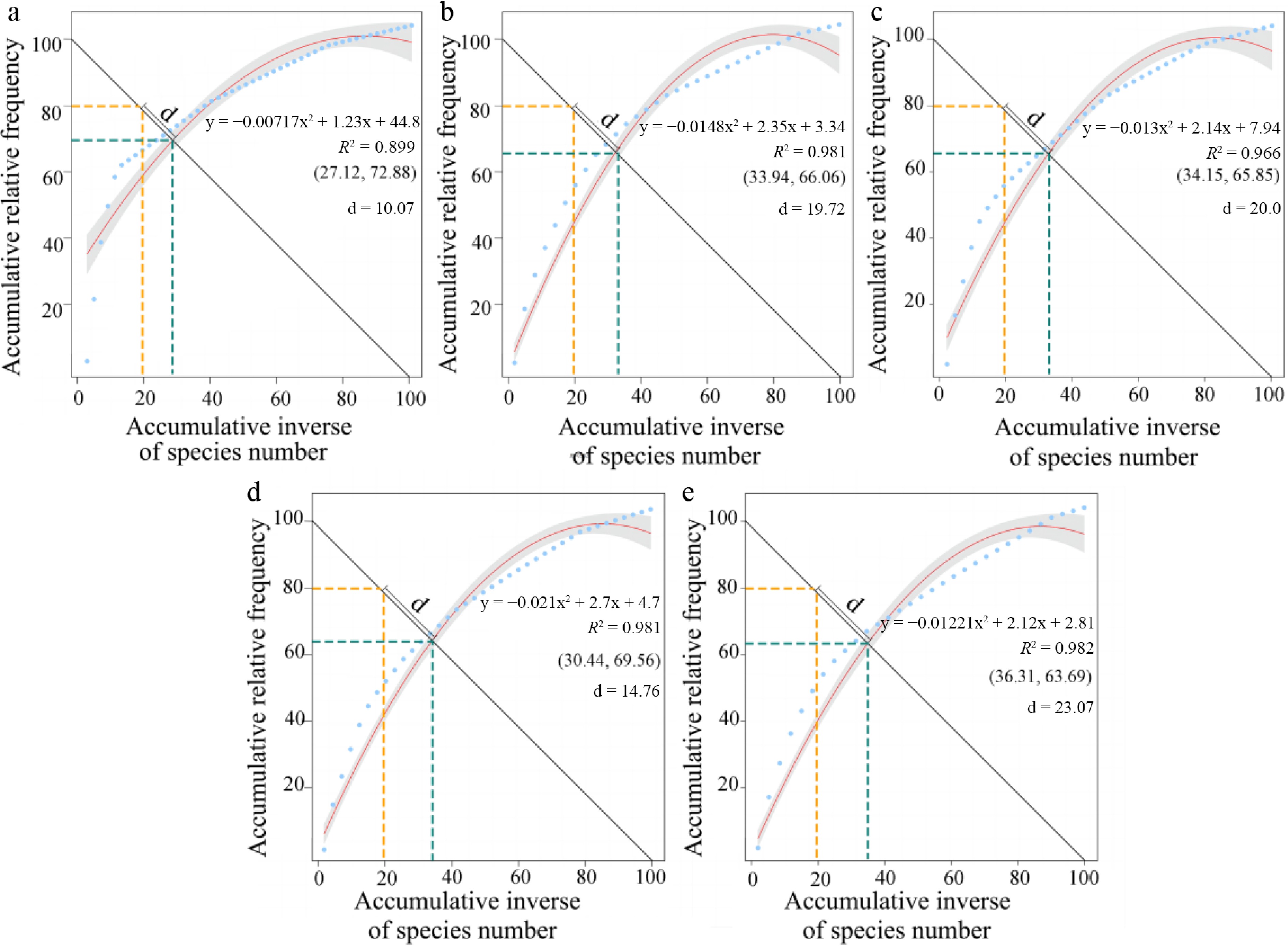
Figure 5.
Stand-age variation in dominance structure and inferred community stability in Quercus acutissima plantations based on the Godron approach. Panels (a)–(e) correspond to stands aged 4, 10, 25, 45, and 50 years, respectively. In each panel, species are ranked by decreasing relative importance and plotted as cumulative relative importance (x), against cumulative species proportion (y). A quadratic function is fitted to the (y)–(x) relationship to describe community dominance structure. The stability index (d) is calculated as the Euclidean distance between the fitted intersection point ([x, y]), and the theoretical reference point (20, 80), where smaller (d) values indicate a dominance structure closer to the reference configuration. Line types indicate the reference point (yellow), the fitted intersection (green), and the fitted curve (red).
Effects of different forest ages on environmental factors in Quercus acutissima plantations
-
In the analysis, the Shannon index overstorey evenness, overstorey stand basal area, light availability, light variability, as well as substrate diversity, are significantly different among various stand ages (p < 0.001) (Fig. 6). Similarly, soil C, N, C/N, soil organic matter, soil pH, soil water content, soil bacterial Shannon index, soil fungal Observed OTUs, and soil fungal Simpson index showed significant influence by uneven age plantation groups (p < 0.05).
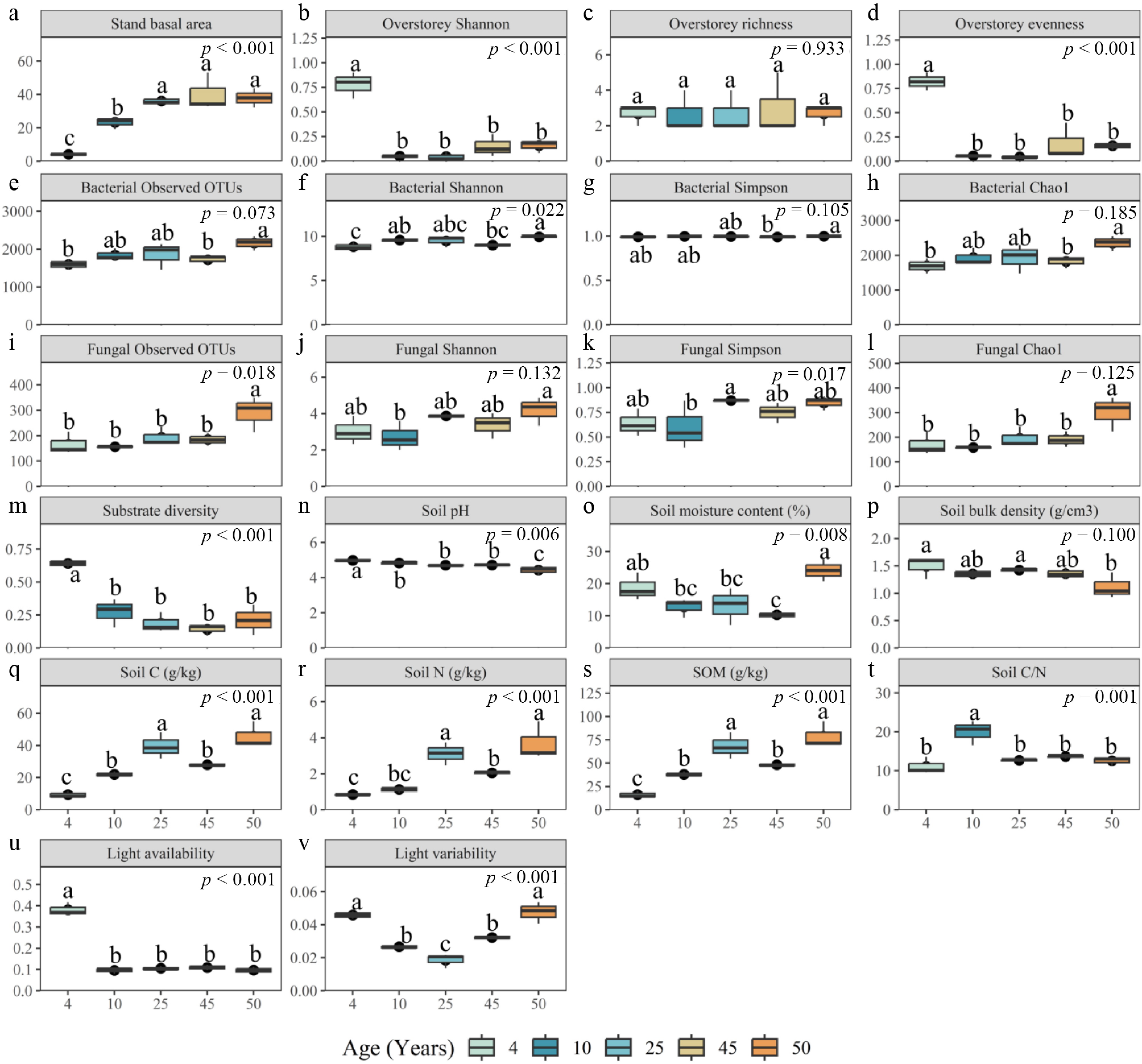
Figure 6.
Stand-age differences in overstory structure, soil properties, light environment, substrate heterogeneity, and microbial diversity in Quercus acutissima plantations. Panels show (a) stand basal area; (b)–(d) overstory diversity metrics (Shannon, richness, evenness); (e)–(h) bacterial diversity indices (observed OTUs, Shannon, Simpson, Chao1); (i)–(l) fungal diversity indices (observed OTUs, Shannon, Simpson, Chao1); (m) substrate diversity; (n)–(t) soil physicochemical variables (pH, moisture content, bulk density, soil C, soil N, organic matter, and C/N), and (u)–(v) canopy light metrics (mean light availability and light variability). Data is summarized across plots within each age class (4, 10, 25, 45, and 50 years; n = 3 per class). Boxes indicate the interquartile range with the median; whiskers show the 5th–95th percentiles, and points represent the plot values. Reported p values are from one-way ANOVA. Different letters denote significant pairwise differences among age classes based on post hoc comparisons (α = 0.05).
Overstory stand basal area, soil fungal observed OTUs, soil C, soil N, and soil organic matter content showed an overall increasing trend with forest age groups. The tree crown Shannon index, overstory evenness, substrate diversity, soil pH, and light availability decreased. With increasing stand age, light variability first decreased, reached a minimum at 25 years, and then gradually increased. Inversely, the soil C/N ratio increased and then decreased with forest stand age, reaching its highest value at 10 years plantation age. The soil bacterial Shannon index showed a trend of increasing, decreasing, and then increasing with forest age, reaching a maximum at 50 years (Fig. 6).
Factors affecting the species composition of the understory
-
NMDS scaling ordination of understory communities related to resource availability and heterogeneity, indicates that substrate diversity, soil C, N, organic matter content, pH, and light availability were the key factors driving changes in plant community composition in the shrub and herb layer (Fig. 7). Environmental gradients related to these factors contributed significantly to the high differences of species groups. Soil properties, particularly organic matter content, showed strong correlations with plant community positioning along NMDS axes, highlighting their importance in shaping community structure, and underscoring the roles of resource availability and heterogeneity as drivers of understory community dynamics.
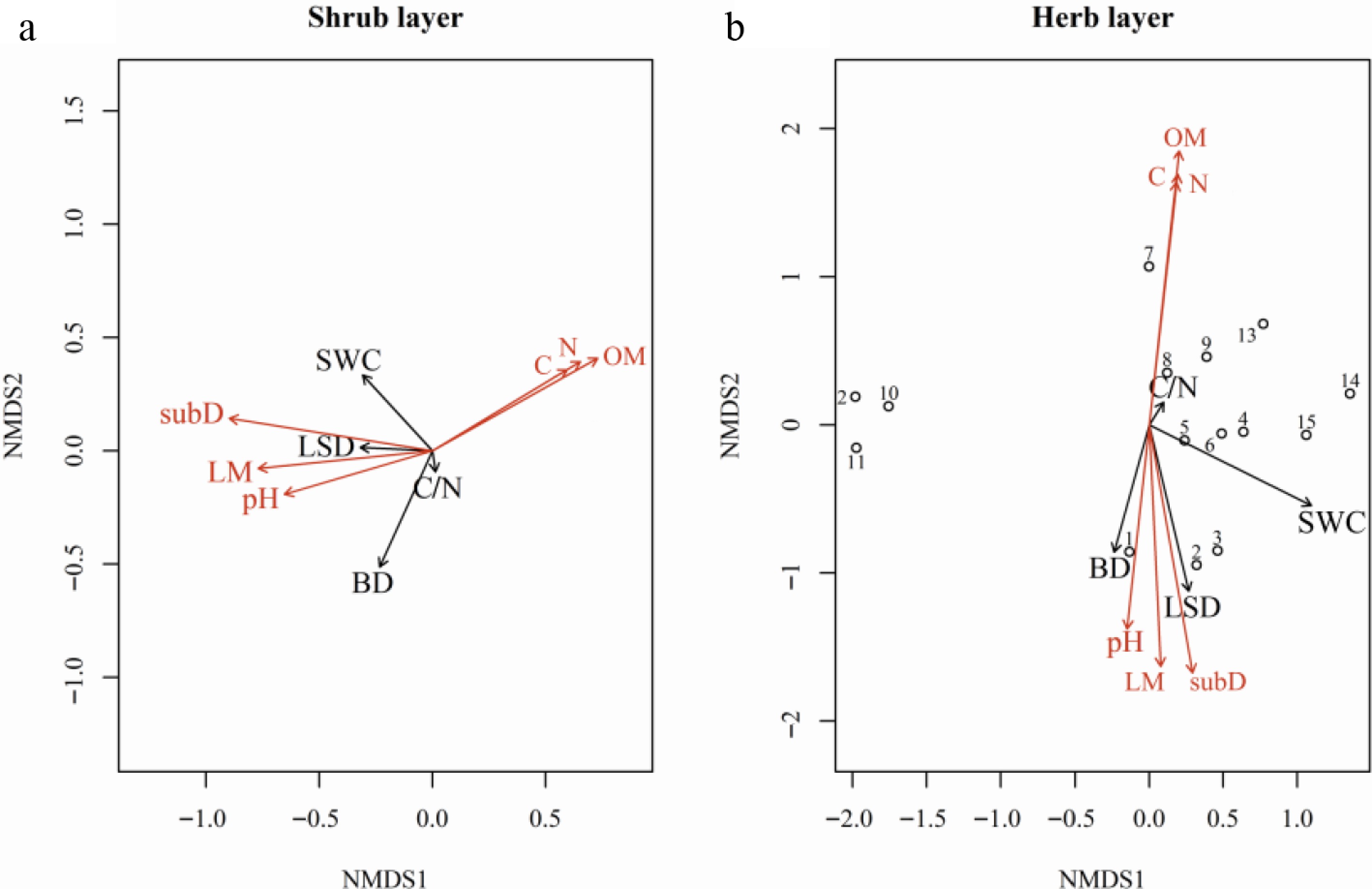
Figure 7.
NMDS ordinations of understory community composition with fitted environmental vectors for the (a) shrub, and (b) herb layers. Points represent plots, and arrows show the direction and strength of correlations between community structure and measured variables: mean light availability (LM), light variability (LSD), substrate diversity (subD), soil carbon (C), soil nitrogen (N), soil organic matter (SOM), soil C/N ratio, soil pH, soil water content (SWC), and soil bulk density (BD). Arrow length is proportional to the correlation magnitude, and arrow direction indicates increasing values of the corresponding variable. Vectors are colored by significance of the fitted relationship (red: p < 0.05; black: p ≥ 0.05).
Linking environmental factors to understory plant diversity
-
In the SEM analysis, shrub richness and herb Shannon's index were significantly fitted with plant diversity (Fig. 8). We found that overstory stands basal area had a positive direct effect on shrub richness (r = 0.951) and exhibited an indirect adverse impact through soil N and soil bacterial Shannon's index. Overstory evenness had a positive effect (r = 0.496) on herb Shannon's index and a negative correlation with shrub Shannon's index (r = −0.601) and soil C (r = −0.549). These relationships revealed the complex interactions among plants, soil nutrients, and soil microorganisms in the understory of a Q. acutissima plantations. SEM showed that 62.6% of the factors affecting shrub richness were due to overstory stand basal area, soil N, and soil bacterial Shannon, while 62.4% of the factors influencing Shannon were due to overstory evenness, shrub Shannon index, and soil C.
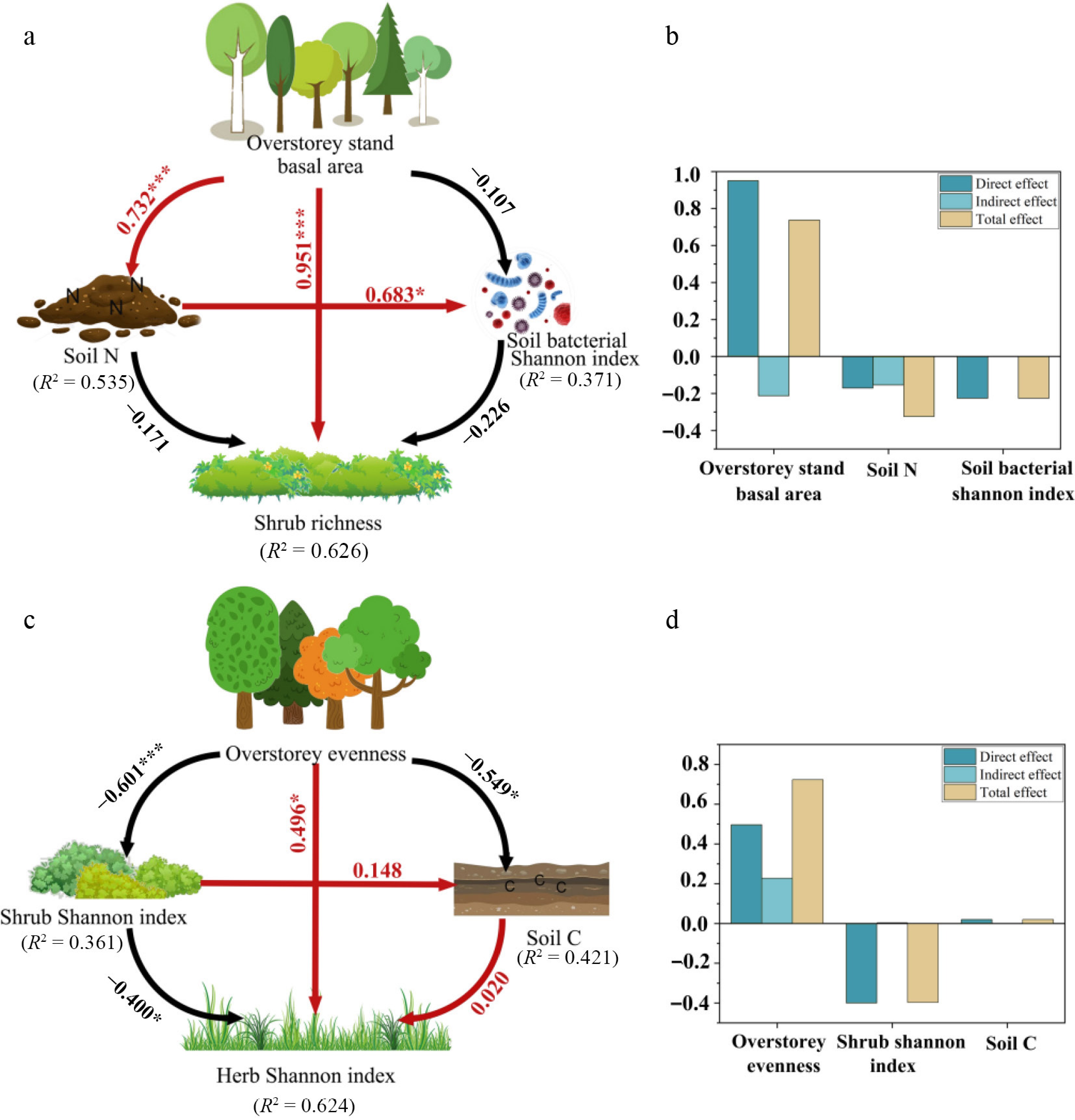
Figure 8.
Structural equation models summarizing hypothesized pathways linking overstory attributes, soil conditions, soil bacterial diversity, and understory diversity in Quercus acutissima plantations. Panels presenting the models for (a), (b) shrub richness, and (c), (d) for the herb-layer Shannon diversity. Solid arrows indicate directional relationships; arrow colors denote the sign of the effect (red = positive, black = negative), and only statistically supported paths are shown (p < 0.05). Numbers on arrows are standardized path coefficients. Values adjacent to response variables report the proportion of variance explained (R2). Bar plots on the right decompose the total effects for the predictors retained in each model into direct and indirect components. ***, * denote p < 0.001, and p < 0.05, respectively.
-
Soil nutrient content is an essential indicator of forest soil quality, which is significant for the establishment and management of plantation forests[18,38]. As forests mature, vegetation cover and biomass increase, while inputs of litter and root exudates rise, providing a continuous source of organic matter and nutrients to the soil[39,40]. These inputs not only improve soil structure, but also enhance microbial activity and C, N cycling, thereby increasing soil nutrient availability and long-term storage capacity[41]. Our findings demonstrate that forest stand age significantly affects soil C and N as well as soil C/N ratio. Specifically, we observed an overall increase in soil C, N, and organic matter content with advancing forest age. This aligned with the findings of a previous study[42], which reported the characteristics of vegetation and soil changes in uneven-age Robinia pseudoacacia plantations. The observed forest litter layer and forest age likely contributes to enhanced soil nutrient availability through increased decomposition[18,33].
Plant diversity, which is primarily influenced by soil nutrient levels, is a vital component of terrestrial ecosystem health. In the context of forest management, conserving biodiversity can enhance economic efficiency and community stability[21,43−45]. Usually, the composition and diversity of plants in the understory of plantation forests changes with forest age, becoming more variable[21,43−45]. The outcomes of this study demonstrated significant differences in understory species composition among stand age groups[21,43−47]. Indicator species analysis of the shrub and herb layers revealed a shift in understory communities toward shade-tolerant species with stand development, consistent with previous findings[21,46−48]. This shift is likely driven by reduced light availability and intensified competition for space and resources as canopy closure increases.
In Q. acutissima plantation forests, the species richness index of the understory shrub layer showed that mature forests (45 years) exhibited the highest species richness[49,50], consistent with a previous study, which is consistent with the 'mid-species richness hypothesis'. This hypothesis posits that species richness follows a unimodal pattern throughout succession, peaking during the mid-succession stage[50]. In the context of Q. acutissima plantations, the forest-age gradient can be regarded as a chronosequence, reflecting secondary succession of understory communities following afforestation. The understory plant richness of Q. acutissima plantation forests is correlated with the stand structure (Supplementary Fig. S1). The observed increases in species evenness, Shannon index, and coverage of the herb layer with higher light availability and substrate diversity suggests that stand structure, light, and substrate diversity are key drivers of understory plant diversity. As the Q. acutissima plantation matures, the understory vegetation diversity gradually transitions from light- and moisture-loving herbaceous plants, to shade-tolerant, nutrient-poor-tolerant shrubs and herbaceous plants (Fig. 2, Table 2). This phenomenon may be attributed to the immediate growth advantages that annual herbaceous plants exhibit following disturbance, thereby impeding the establishment of late-successional species that are characterised by their capacity to tolerate shade and nutrient-poor conditions[6,9]. Conversely, higher shrub cover and diversity in mature Q. acutissima plantations has been shown to reduce available resources for the herbaceous layer through shading and competition for soil resources[9,33]. These findings highlight the importance of managing stand structural and substrate conditions to promote biodiversity and maintain ecosystem functioning in Q. acutissima plantations[50].
The physical and chemical properties of soil are fundamental to plant growth and development, influencing both their physical structure and nutrient availability[21,43−45]. Correlation analysis demonstrated that understory plant diversity was strongly and positively associated with multiple soil environmental parameters, suggesting that soil nutrient availability plays a crucial role in shaping understory diversity patterns (Supplementary Fig. S1). Results from redundancy analysis (RDA) further identified soil C, N, organic matter content, pH, the C/N ratio, and light availability as significant determinants of understory plant diversity. Among these variables, soil C content showed a general upward trend with forest maturation, consistent with Liu et al.[51] but contrasting with Zhang et al.[52]. Such variation may stem from differences in litter production and decomposition dynamics among stand types, thereby influencing soil carbon turnover and accumulation. Soil N was significantly correlated with soil C content (p < 0.05), which refers to the fact that soil N can be used as an indicator of changes in soil organic C pools[52], and soil N content generally increased with forest age group, with a slight decrease in the middle-aged forest stands, apparently because Q. acutissima was in a period of rapid growth, which resulted in a strengthened consumption of soil N. The soil N content of plantations was also significantly correlated with soil C content, with a slight decrease in the middle-aged forest stands.
As Q. acutissima has high quality habitat requirements, it favors warm, humid climates, and grows best on fertile, well-drained soil[29,30]. In this study, an investigation was conducted into the soil physical properties present in Q. acutissima plantations of varying ages. It was determined that the soil bulk density in the young (4-year) stand exceeded those in other stands. This finding indicated that Q. acutissima could affect soil compactness, thereby reducing soil aeration. This reduction in soil aeration was identified as the primary cause of the decline in species diversity observed in Q. acutissima plantations. Given the observed decline in community stability during the intermediate stages (10 and 25 years), we recommend implementing targeted management interventions to accelerate the transition toward a stable, mature community structure. Specifically, moderate thinning should be prioritized in these mid-successional stands to increase understory light availability and heterogeneity, thereby mitigating the adverse effects of canopy closure on light-demanding herbaceous species. Furthermore, to facilitate succession and address the structural gaps identified in the middle-aged stands, the introduction of functional shade-tolerant shrub species is advisable to establish a multi-layered vertical structure and optimize resource utilization. These interventions, combined with the monitoring of soil physical conditions, are essential for mitigating the 'stability trough' during rapid stand development, thereby enhancing the long-term ecological resilience and biodiversity maintenance of Q. acutissima plantations. Consequently, in accordance with the principle of suitable land and suitable plants, it is not appropriate to select land with strong clayey soil for future afforestation; rather, it is preferable to choose sandy or sandy loam land for planting Q. acutissima. The findings indicate that stand development and soil conditions are central correlates of understory diversity and community structure, providing evidence to inform biodiversity-conscious management of plantation forests.
-
This study investigated how understory vegetation communities respond to stand age in Q. acutissima plantations in the Huangfu Mountain forest. By integrating vegetation surveys with analyses of soil properties, light conditions, and overstory plants, we revealed that understory vegetation communities in Q. acutissima plantations varied across age groups and environmental factors. As plantation forests matured, species composition shifted from light-demanding to shade-tolerant species, revealing distinct successional patterns with varying stand ages. Significant variations in understory vegetation cover, richness, and diversity were observed among age groups, underscoring the importance of accounting for forest age in conservation planning. Further, plantations at 10 and 25 years showed instability, highlighting the need for adaptive management during these stages. The highest plant diversity observed in the mature forests (45 years) reaffirms the functional importance of mature forest ecosystems. Factors such as light availability, substrate diversity, and the interactions between overstory and soil properties play a vital role in shaping understory plant communities. However, given the geographical limitation of this research to a single region, further studies across different geographic areas are necessary to confirm the broader applicability of these findings. In the future, cross-regional studies will be conducted to further validate these patterns.
This work was supported by the Anhui Provincial Forestry Research and Innovation program ('Research on Forest Plant Diversity Restoration Techniques'), the Anhui Province University Natural Science Research Foundation (KJ2024AH051402), Anhui Provincial Natural Science Foundation (2308085Y27), the Science Research Project in Universities of Anhui Province for Distinguished Young Scholars (2022AH020070), and the National Natural Science Foundation of China (42307055). We are grateful to Fengyu Xu, Tao Huang, Xiaoxiang Hou, Qiying Sun, Rui Hua, Yangyang Chen, Weijie Tong, and Shuai Ma for assistance with field sampling and laboratory work.
-
The authors confirm their contributions to the paper as follows: conceptualization: Ma Y, Zang L, Li X; investigation and formal analysis: Ma Y, Xu D, Xu H, Liu K, Wang R; statistical analyses, visualization, writing – original draft, project administration, funding acquisition: Ma Y; writing – review & editing: Wu Z, Xiang M, Haider FU, Zhang L, Li X; supervision: Zhang L, Li X. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
-
accompanies this paper online at: https://doi.org/10.48130/fra-0026-0003.
- Supplementary Fig. S1 Effects of different environmental factors on understory diversity in different forest ages.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ma Y, Xu D, Xu H, Liu K, Wu Z, et al. 2026. Understory vegetation diversity dynamics across a chronosequence of Quercus acutissima plantations: roles of stand development and environmental factors. Forestry Research Advances 1: e005 doi: 10.48130/fra-0026-0003 

Understory vegetation diversity dynamics across a chronosequence of Quercus acutissima plantations: roles of stand development and environmental factors
- Received: 29 November 2025
- Revised: 12 February 2026
- Accepted: 02 March 2026
- Published online: 14 April 2026
Abstract: Investigating the dynamics of species composition and their influencing factors is critical for evaluating plant communities' current state and future trajectory. However, how their composition and diversity vary with forest development, and which environmental factors control these patterns, remain poorly understood in Quercus acutissima plantations. This study addresses key knowledge gaps by examining understory vegetation composition dynamics in Q. acutissima plantations of different stand ages. Here, we measured understory vegetation composition dynamics and the related biotic and environmental factors in Q. acutissima plantations of several ages (4, 10, 25, 45, and 50 years). Results show that soil carbon (C), nitrogen (N), and organic matter content (SOM) improved significantly with forest age, whereas soil bulk density and water content remained stable. Understory species composition and relative abundance differed markedly among forest age classes, showing a clear shift from heliophilous (light-demanding) to shade-tolerant species as stands matured. Herb and shrub layer richness was highest at 45 years of forest. The highest Shannon diversity index and herb richness, observed in young forests (4 years old), were strongly influenced by sunlight availability. Key factors influencing understory plant diversity included C, N, SOM, pH, and C/N ratio. Moreover, both soil chemical properties and light availability exerted strong effects on plant community composition across forest stands. These results show that stand development and soil conditions are central correlates of understory diversity and community structure, offering evidence to guide biodiversity-conscious management of plantation forests.
-
Key words:
- Quercus acutissima plantations /
- Plant diversity /
- Stand ages /
- Understory