








-
Distant hybridization, the crossing of genetically distinct species or genera, represents a powerful yet under-utilized tool in forest tree breeding and conservation. By introducing novel alleles from wild or distantly related taxa, it enhances genetic diversity, disease resistance and adaptive capacity. This is especially critical under climate change, habitat fragmentation and emerging pathogens, where a limited genetic base of many tree species may be insufficient for long-term survival[1]. Distant hybridization can broaden this base by conferring traits such as drought tolerance, accelerated growth and enhanced resilience[2].
Significant progress in distant hybridization has also been made in major crop species, where wide crosses in cereals, rice, and Brassica have enabled the introgression of disease resistance, stress tolerance, and fertility-related traits through techniques such as embryo rescue, chromosome doubling, and somatic hybridization[3−5]. Similar advances in horticultural crops, including citrus and sugarcane demonstrate how cytogenetics and tissue culture can overcome strong reproductive barriers[6]. These developments provide useful conceptual and technical parallels for distant hybridization efforts in long-lived forest trees.
This review evaluates the reproductive barriers that limit distant hybridization in forest trees and the strategies developed to overcome them. Barriers are organized into pre-zygotic and post-zygotic mechanisms, with emphasis on their operation in trees. Conventional techniques, including mentor pollen and cut-style pollination, are examined alongside biotechnological advances such as in vitro culture, genome editing and molecular breeding. Case studies from both model systems and tree genera (e.g., Populus, Chitalpa, Salix) are included to illustrate practical applications and challenges.
Despite recent advances, distant hybridization in forest trees faces persistent obstacles. Extended generation times slow breeding cycles, while large and complex genomes hinder molecular integration[7−9]. In vitro methods, though useful, remain difficult to scale and transfer to field conditions[10]. Moreover, hybrids that thrive under controlled conditions often fail in natural environments, reflecting unresolved questions of ecological viability[11,12]. The limited integration of genomic, physiological, and ecological data further constrains predictive capacity[13]. By synthesizing current progress and identifying these knowledge gaps, this review aims to outline more resilient and productive forest hybrid strategies.
-
Understanding reproductive barriers is essential for improving the success of distant hybridization in forest trees, as these barriers often limit interspecific and intergeneric breeding. Reproductive barriers may occur at pre-zygotic stages, such as differences in habitat preference, flowering time, reproductive structures, or at post-zygotic stages, including reduced hybrid viability or sterility[14,15]. These barriers may be shaped by environmental factors or operate independently of the ecological context.
Pre-zygotic barriers and overcoming strategies
-
Pre-zygotic barriers prevent fertilization before the formation of a zygote[16]. In distant forest tree hybridization, such barriers are common and may include differences in flowering time (a pre-pollination barrier), and pollen-pistil incompatibility (a post-pollination barrier)[17−20] (Fig. 1).
Other pre-zygotic barriers reported in herbaceous plants, such as pollinator specialization or floral morphological divergence, are less relevant in forest trees, which are predominantly wind-pollinated or artificially crossed[21]. Changes in the identity or behavior of pollinators can influence natural hybridization by altering pollen flow between species. In many angiosperms, transitions in pollination modes have been shown to modify or break down reproductive barriers[22]. Phylogenetic studies similarly demonstrate that pollinator-driven transitions can promote gene exchange and diversification across plant lineages[23]. Although such processes are less relevant in forest trees, which are mostly wind-pollinated or manually crossed, they illustrate how ecological factors can shape pre-zygotic compatibility in other systems (Fig. 1). Given its pivotal role in preventing successful fertilization and its complexity in tree hybridization, we will delve deeper into the mechanisms of pollen-pistil incompatibility and temporal isolation in subsequent sections.
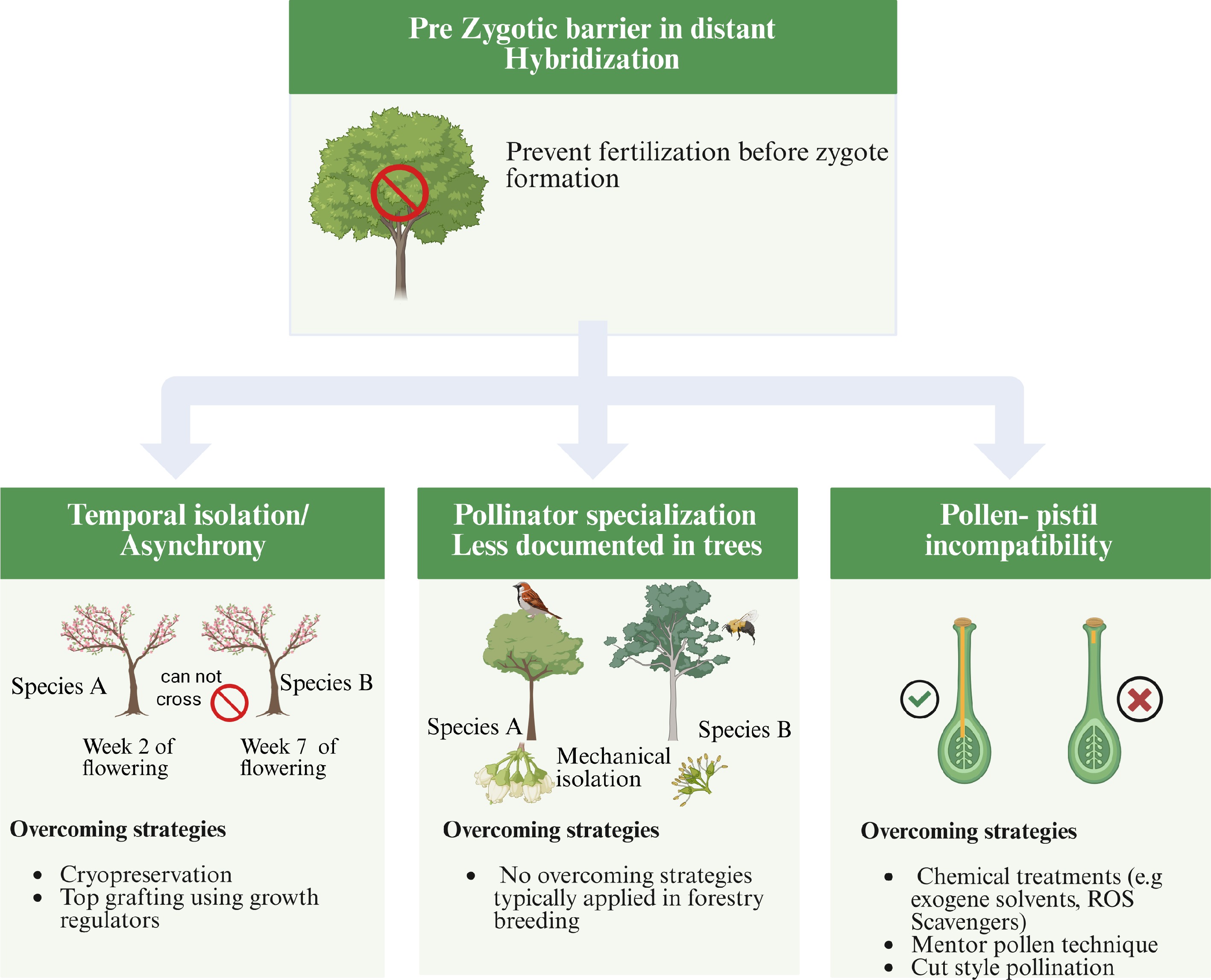
Figure 1.
Schematic illustration of major pre-zygotic barriers encountered in distant hybridization of forest trees, including temporal isolation/asynchrony, pollen–stigma incompatibility, and mechanical isolation. Each barrier prevents fertilization before zygote formation and limits hybrid success. Strategies such as controlled pollination, hormonal treatment, and style manipulation are used to overcome these reproductive obstacles and facilitate successful hybrid formation.
Temporal isolation or reproductive asynchrony
-
Temporal isolation, caused by mismatched flowering times, is a recognized pre-zygotic barrier in plant hybridization[24]. In crop wild relative systems, flowering asynchrony was shown to be a highly effective mechanism for preventing hybridization, emphasizing the role of phenological mismatches in limiting gene flow[24]. However, its role in forest trees is less documented. Variation in flowering time due to genotype or environment has been observed in trees[25]. For instance, delayed flowering was observed in Eucalyptus globulus clones compared with seedlings under different climates[26]. Further evidence of temporal isolation as a hybridization barrier comes from Fraxinus excelsior and Fraxinus angustifolia, where assortative mating based on flowering time contributed to isolation by time within a hybrid zone[27].
To address this challenge, researchers have explored methods to manipulate flowering time and extend pollen viability. One approach involves top grafting combined with the application of paclobutrazol, which has been shown to induce early flowering in Eucalyptus genotypes, thereby aligning reproductive timing and accelerating breeding efforts[28]. Another promising strategy is pollen cryopreservation, which enables the long-term storage of viable pollen for use in hybridization when female trees flower out of sync. Cryopreservation enables long-term storage of viable pollen, allowing breeders to overcome mismatched flowering times in distant crosses. This technique has been widely applied in crop and woody species, and is increasingly used in forestry to facilitate hybridization when flowering periods do not overlap. Rajasekharan & Rohini highlight cryobanking as a practical tool for synchronizing reproductive events, and early forestry work also demonstrated the feasibility of storing viable Pinus taeda pollen for breeding applications[29,30]. These techniques offer practical solutions to synchronize reproductive events and overcome temporal barriers in forest tree breeding programs[29].
Pollen–pistil incompatibility
-
Pollen-pistil incompatibility occurs when pollen fails to germinate on the stigma, or when pollen cannot reach the ovules[19]. This incompatibility arises from complex interactions between pollen and pistil tissues, which can actively reject foreign pollen or fail to support its growth due to incongruity[31]. Specific receptor-like kinases on the stigma surface, and peptides present on pollen surface play a crucial role in the pollen-pistil interaction[32]. These molecules form a 'lock-and-key' system, where the receptor-like kinases on the stigma interact with complementary peptides from the pollen tube. In species such as Arabidopsis thaliana and Brassica oleracea this interaction is critical for pollen compatibility. When the pollen is incompatible this system prevents the pollen tube from penetrating the stigma[33].
Recent work in Arabidopsis and other Brassicaceae has clarified key pollen–pistil signaling pathways underlying compatibility. Studies from Lijia Qu's group have shown that PCP-B peptides and stigma receptor kinases regulate pollen hydration and penetration, and their recent work highlights how male-female communication governs compatibility decisions during early reproduction[32,34]. In parallel, Qiaohong Duan's laboratory has demonstrated that RALF peptides, FERONIA signaling, and stigma-expressed receptors mediate both intra- and inter-specific pollen rejection[33,35]. Although derived from herbaceous plants, key elements like CrRLK1L receptors, cysteine-rich peptides, and ROS-mediated cell wall responses are conserved in angiosperms. Evidence from woody perennials such as ROS roles in Fraxinus pollen tube growth, and Betula female gametophyte development supports this[36,37]. These parallels suggest that analogous signaling systems may operate in forest trees and could contribute to pre-zygotic barriers observed in genera such as Populus and Salix.
In Brassicaceae, self-incompatibility (SI), and unilateral incompatibility (UI), are mediated by the SRK–FER–ROS signaling pathway (S-locus Receptor Kinase–FFERONIA–Reactive Oxygen Species), which triggers reactive oxygen species (ROS) production to prevent fertilization by incompatible pollen[35]. Additionally, the stigma-specific protein SPRI1, independent of ROS signaling, is essential for species-specific rejection of distantly related pollen, as shown in mutants of Arabidopsis thaliana[38].
Overcoming pollen-pistil incompatibility
-
Overcoming pollen-pistil incompatibility can be achieved through various techniques, including the use of mentor pollen, which provides specific recognition or growth-promoting substances that help overcome incompatibility barriers[39]. The mentor pollen technique uses irradiated or chemically treated pollen that is genetically compatible but incapable of fertilization. When mixed with viable but incompatible pollen, it stimulates the pistil's response and enables the incompatible pollen to germinate and grow[33,40]. Studies have shown that mentor pollen can provide specific recognition substances or growth-promoting substances that help overcome incompatibility barriers[33]. For instance, in the genus Populus, this approach has been used to facilitate both intra- and inter-specific hybridization by promoting pollen tube growth[39,40]. Applying synthetic pRALFs (Rapid Alkalinization Factor peptides), or mutating gate components allowed pollen from distant species to penetrate the stigma and form hybrid embryos[33]. The technique has been successfully applied in various plant species, including black cottonwood and Brassica, where it helped overcome stigma-level incompatibility barriers[41,42].
Chemical treatments are another way of overcoming pollen-pistil incompatibility. It involves the application of exogenous solvents like Ether, kinetin (KDA), α-indoleacetic acid (AIA), and α-naphthaleneacetic acid (ANA) on the stigma to stimulate pollen germination and enhance pollen tube growth[43]. Hormonal treatments involve the application of plant hormones such as gibberellic acid (GA3), and 2,4-Dichlorophenoxyacetic acid (2,4-D), to stimulate pollen germination and enhance pollen tube growth[44]. These hormones modulate auxin and gibberellin pathways, which are crucial for fertilization and embryo development[44]. Although specific studies on hormonal treatments in the provided contexts are limited, the general principle is well-documented in plant breeding literature[45,46].
Cut-style pollination is a technique where a portion of the style is removed to allow direct access of the pollen tube to the ovary, bypassing the incompatible upper style[47]. This method has been effectively used in loquat (Eriobotrya japonica) to overcome pre-fertilization barriers in hybridization[48]. The removal of the style's upper third, where pollen tube growth is typically impaired, allows successful fertilization between incompatible species[49,50].
While various techniques, such as mentor pollen and chemical treatments, help overcome pollen-pistil incompatibility, genetic and biochemical approaches targeting specific molecular pathways offer a more targeted solution to overcoming SI and UI barriers. To overcome SI- and UI-mediated pre-zygotic barriers in Brassica rapa, the SRK–FER–ROS signaling pathway was disrupted using biochemical and genetic approaches. Sodium salicylate (Na-SA) acted as a ROS scavenger, while S-nitrosoglutathione (GSNO), a nitric oxide donor, nitrosated FER to suppress ROS production. Additionally, antisense oligodeoxynucleotides (AS-ODNs) were used to silence SRK, FER, and RBOHs, reducing the stigma's rejection response. These interventions successfully enabled inter-specific and inter-generic hybrid embryo formation, offering a practical strategy for distant breeding in Brassicaceae crops[35].
For SPRI1 mediated barriers, although no direct solution was applied in the SPRI1 study, mutation of the SPRI1 gene (e.g., in spri1-1 knockout lines) allowed foreign pollen to germinate and grow into the pistil, bypassing inter-specific rejection[38]. This suggests that genetic modification or targeted gene editing could be used to disable SPRI1-mediated barriers. Such an approach holds potential for facilitating hybridization across species, independent of self-incompatibility mechanisms.
Recent studies highlight the critical role of ROS balance and cell wall dynamics in pollen tube growth and successful fertilization. In Fraxinus mandshurica, ROS were shown to be essential for pollen germination and tube elongation[36], while in Betula platyphylla, proper ROS homeostasis regulates female gametophyte maturation, linking oxidative signaling with ovule development[37]. Similarly, in Arabidopsis thaliana, defects in pollen cell wall assembly lead to sterility, acting as strong pre-zygotic barriers[50]. Imbalances in ROS signaling or defects in pollen cell wall formation can prevent successful fertilization, constituting important pre-zygotic barriers. Targeted manipulation of ROS pathways and pollen tube growth genes could support distant hybridization efforts in forest trees[51].
While each method has limitations, they collectively mark significant progress in overcoming pollen–pistil incompatibility, especially for distant hybridization in forest trees and crops. Classical approaches like mentor pollen, hormonal treatments, and cut-style pollination have shown success in bypassing stigma or style barriers[41,44,47]. Molecular strategies, such as disrupting the SRK–FER–ROS pathway or SPRI1 knockout, offer gene-targeted solutions in Brassicaceae[35,38]. These findings highlight how combining classical and molecular approaches can effectively break pre-zygotic barriers and enhance hybridization potential across species.
Post-zygotic barriers and overcoming strategies
-
Post-zygotic barriers arise after fertilization and prevent the normal development or reproductive ability of hybrid progeny[16]. These barriers are generally more difficult to overcome due to their complex genetic and developmental basis[52]. Key forms include hybrid inviability, hybrid sterility, and ecological inviability (Fig. 2).
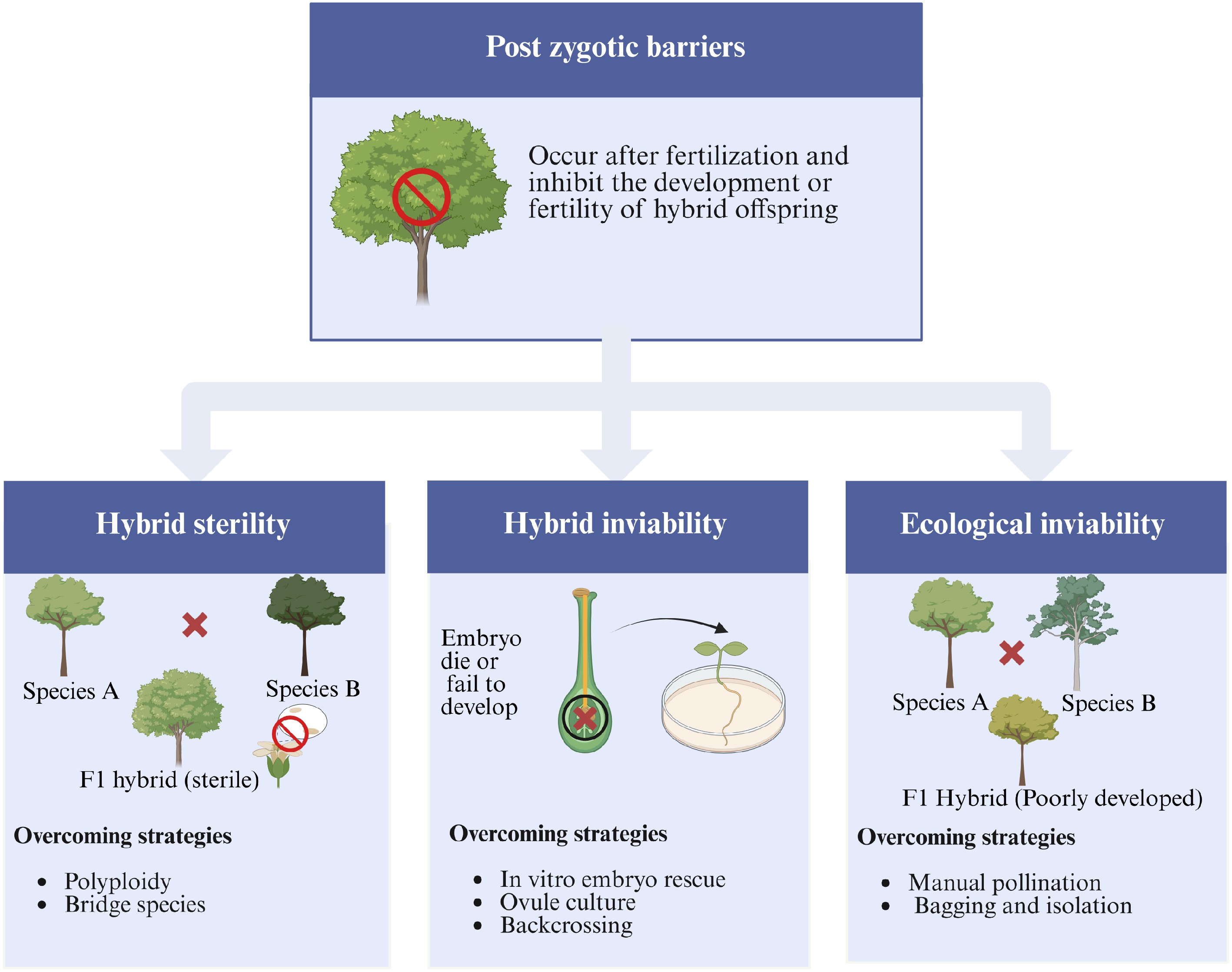
Figure 2.
Conceptual framework illustrating major post-zygotic barriers encountered in distant hybridization of forest tree species. The figure highlights three primary forms: (1) Hybrid sterility, where F1 hybrids are unable to produce viable gametes; (2) Hybrid inviability, characterized by embryo abortion or failure to develop; and (3) Ecological inviability, where hybrids are maladapted to environmental conditions. Associated strategies to overcome these barriers include polyploidization, embryo rescue, and ecological modeling approaches aimed at improving hybrid establishment and performance.
Hybrid inviability and sterility
-
Hybrid inviability occurs when hybrid embryos fail to develop or die shortly after germination, often due to developmental incompatibility or endosperm failure[53]. Hybrid sterility, by contrast, results when hybrids grow into mature individuals but are unable to produce functional gametes, often due to irregular chromosome pairing during meiosis[54].
These phenomena are common in distant crosses, such as between divergent Eucalyptus species, where hybrid survival is markedly lower than intraspecific crosses[11]. Strategies to overcome these barriers are explained below.
Embryo rescue is an in vitro technique that involves excising immature embryos from developing seeds before endosperm degeneration leads to abortion, and culturing them on nutrient media, effectively rescuing embryos that would otherwise fail in distant hybrids[55]. It has been applied in Populus breeding to overcome post-fertilization ovule loss and in conifers like Pinus ayacahuite, where cotyledonary embryos are cultured in vitro[56]. However, success depends critically on precise excision timing, optimized culture media, and strictly sterile lab practices[57]. Limitations include labor-intensive protocols, relatively low survival rates for some species, and the necessity for controlled sterile conditions[58].
Ovule culture is another method to overcome inviability. In some cases, culturing the entire ovule (instead of just the embryo) provides a more protective environment and improves recovery rates, particularly when embryo development is extremely early or delicate[59]. Ovule culture was used in the hybridization of Populus alba and Populus euphratica, resulting in over 90% success in plantlet development, compared to 70% with ovary culture. This technique facilitated the acclimatization of 75 interspecific hybrid plants in a greenhouse, demonstrating its effectiveness in Populus hybridization[60].
Backcrossing is used to overcome hybrid sterility[61]. This involves crossing a hybrid with one of its original parents (typically the recurrent parent) across successive generations to restore fertility and stabilize desirable traits[62]. Research on rice hybrids, particularly between Oryza sativa and Oryza glaberrima, has demonstrated that backcrossing can enhance pollen and spikelet fertility[61]. Similarly, in indica-japonica rice hybrids, pyramiding wide-compatibility alleles through backcrossing significantly increased seed-setting rates and overcame hybrid sterility barriers[63].
Ecological inviability
-
Ecological inviability arises when hybrid offspring, though viable and fertile, are poorly adapted to environmental conditions in either parent's habitat[64]. These hybrids may display maladaptive traits, such as intermediate phenotypes that perform poorly under field stressors (e.g., drought, temperature extremes, or soil type)[65]. The following are strategies to overcome ecological inviability.
Parental selection based on adaptive complementarity, i.e., choosing parents with non-overlapping but synergistic ecological traits (e.g., one drought-tolerant and one cold-tolerant), can help generate hybrids with broader environmental tolerance[66]. For instance, in the case of Eucalyptus, intense selection and testing of large populations have been used to develop elite hybrid clones that exhibit high environmental resilience, although their susceptibility to pests and ecological stressors highlights the challenges hybrids may face in certain conditions[67].
Predictive ecological modeling could be employed to overcome ecological inviability by incorporating genomic and phenotypic data to predict where hybrids will perform well. For instance, models based on hybrid zones in Eucalyptus have been used to guide planting locations under shifting climate regimes[66].
Ultimately, overcoming ecological inviability requires a combination of ecological insight, field trials, and long-term monitoring to ensure hybrids are not only viable, but sustainable in practical applications[68].
Advanced biotechnological approaches
-
In addition to conventional breeding methods, advanced biotechnological approaches have revolutionized the field of forest tree breeding by providing tools to overcome reproductive barriers inherent in distant hybridization[11]. These technologies enable precise manipulation of genetic, cytological, and developmental processes, facilitating the creation of novel hybrid combinations that are often unattainable through traditional breeding methods[69]. Genomic tools have further advanced the understanding of reproductive barriers by identifying loci linked to traits like hybrid sterility, growth, and stress tolerance, as shown in species like Populus and Eucalyptus, guiding early hybrid selection[70].
Genomic tools
-
Genomic technologies have significantly improved the understanding of reproductive barriers at the molecular level[71].Techniques such as whole-genome sequencing, quantitative trait locus (QTL) mapping, and genome-wide association studies (GWAS), have identified loci responsible for traits like hybrid sterility, growth performance, and stress tolerance in forest trees[72]. For instance, genomic tools have been successfully applied in Populus and Eucalyptus to uncover incompatibility genes and guide early selection of suitable hybrid combinations[7,72].
Marker-assisted selection (MAS) facilitates the early detection of desired traits in hybrid progeny, reducing the breeding cycle time. However, large and repetitive genomes, high heterozygosity, and long generation times continue to pose challenges in assembling high-quality reference genomes and applying these tools to understudied genera[72].
Chromosome doubling (polyploidization)
-
Polyploidization is a powerful approach to overcome hybrid sterility, particularly in interspecific and intergeneric hybrids, where meiotic pairing irregularities cause gamete dysfunction[73]. By artificially doubling the chromosome number, homologous pairing can be restored, allowing normal meiosis and fertility. This is typically induced using antimitotic agents such as colchicine or oryzalin[10].
In Populus and Cupressus, chromosome doubling has been used to restore partial fertility and enhance traits like stress tolerance and wood density[74]. While polyploidization offers advantages in plant breeding, it can also lead to genomic instability and reduced growth in some hybrids, underscoring the need for careful cytological evaluation, and tailored breeding strategies based on species-specific goals[75].
In vitro fertilization
-
In vitro fertilization (IVF) is a promising technique to overcome hybridization barriers in distant forest trees, which are often impeded by pre- and post-fertilization barriers. IVF allows for the manipulation of gametes outside the plant's reproductive structures, facilitating the fusion of gametes from different species, and potentially overcoming these barriers[76]. This approach is particularly useful in forest tree breeding, where traditional methods are limited by long generation cycles and inbreeding depression[77]. The following sections detail how IVF is applied to address these challenges.
Isolated gametes can be fused to bypass crossing incompatibility[76]. IVF involves the isolation of male and female gametes, which can then be fused using techniques such as electric pulses or chemical treatments like Ca2+ or PEG. This bypasses the natural barriers that prevent gamete fusion in vivo, allowing for the creation of hybrids that would otherwise be impossible due to crossing incompatibility[76].
Somatic hybridization directly combines genomes of different species. In vitro techniques also enable somatic hybridization, where protoplasts from different species are fused to create somatic hybrids. This method can circumvent pre-fertilization barriers by directly combining the genetic material of two species[78].
Embryo rescue prevents abortion and supports hybrid seedling development. Post-fertilization barriers, such as embryo abortion, can be addressed through embryo rescue techniques. This involves culturing embryos in vitro to prevent abortion and allow them to develop into viable plants[79].
Chromosome doubling restores fertility in sterile hybrid lines. IVF can also facilitate chromosome doubling in haploids, which helps restore fertility in hybrids that might otherwise be sterile. This is particularly useful in creating homozygous lines for breeding programs[80].
While IVF offers significant advantages in overcoming hybridization barriers, it is not without challenges. The regeneration of hybrids from in vitro cultures can be difficult, and the techniques require further refinement to be widely applicable in forest tree breeding programs[81]. Additionally, the complexity of forest tree genomes and the long lifecycle of these species pose unique challenges that are not present in herbaceous plants[82]. Despite these hurdles, the potential of IVF in forest tree breeding and conservation remains substantial, offering a pathway to enhance genetic diversity and improve forest tree species.
-
The following case studies on pre- (Table 1) and post-zygotic barriers (Table 2) exemplify how reproductive barriers in distant hybridization have been addressed. Although numerous artificial hybrids in forest trees have been successfully produced (e.g., Populus simonii × Populus nigra, Catalpa bignonioides × Chilopsis linearis, Franklinia alatamaha × Gordonia lasianthus), there is little published information on the specific pre-zygotic barriers encountered during their creation. Most studies focus on post-fertilization success, molecular confirmation, or phenotypic traits, rather than documenting pollen-stigma incompatibility, asynchronous flowering, or other pre-zygotic mechanisms. The lack of direct studies on pre-zygotic barriers in forest tree hybrids highlights a critical knowledge gap and represents an area for future research. The following are examples, including interspecific and intergeneric crossing, each highlighting different barriers and solutions.
Table 1. Pre-zygotic barriers documented in distant hybridization of forest trees and related genera, together with the approaches developed to overcome them.
Hybrid example Pre-zygotic barrier Solution/approach Ref. Salix viminalis × Populus spp. Stylar blockage: pollen tubes germinate but fail to penetrate the style due to incompatibility in transmitting tissue. Placental (cut-style) pollination to bypass the style and allow direct ovule access, enabling micropyle entry and embryo initiation. [83] Salix × Populus Extremely short stigma receptivity (1–2 d) and reduced pollen germination on Salix stigmas, resulting in low fertilization efficiency. Pollination on day 1 of anthesis using high pollen loads to maximize successful germination and improve crossability. [84] Catalpa × Catalpopsis Pollen–stigma incompatibility leading to failed pollen germination. preventing normal pollen tube growth and fertilization; cross failures occur at the initial stigma interface. Grafting onto dwarf Catalpopsis rootstocks; low-temperature storage of floral branches; mixed pollen application under high temperature; stamen removal. [85] Populus
intersectional crosses (e.g., Leuce × Aigeiros, Leuce × Tacamahaca)Species-specific stigma-level incompatibility; In Populus tremuloides, pollen from Aigeiros and Tacamahaca sections fails to germinate or induces callose deposition, indicating rejection. In P. trichocarpa, Leuce pollen germinates, but pollen tubes are arrested within the stigma before entering transmitting tissue. Use of mentor pollen (heat-treated compatible pollen mixed with incompatible pollen) to soften recognition barriers; stigma treatments with organic solvents that disrupt lipid-based incompatibility signals and temporarily enhance receptivity. [86−88] Table 2. Representative examples of distant hybrids in forest trees and related genera, the post-zygotic barriers encountered, and the strategies applied to circumvent them.
Hybrid example Post-zygotic barrier Solution/approach Ref. × Cupressocyparis leylandii Near-complete sterility caused by meiotic irregularities and unbalanced gamete formation; hybrids fail to set viable seeds. Vegetative propagation via cuttings; rooting improved using wounding + auxin (IBA) treatments to enable large-scale clonal production. [90] × Chitalpa tashkentensis Hybrid sterility and early embryo abortion; cytological studies confirm absence or irregular seed development. Polyploid induction (oryzalin) restoring partial fertility through tetraploid and cytochimeric plants; embryo rescue to recover hybrid embryos; routine propagation via stem cuttings. [91,92] Salix viminalis × Populus sp. Embryo abortion due to defective endosperm development, preventing normal seed maturation. In vitro pollination and embryo rescue at globular–cotyledonary stages to recover viable hybrid plantlets. [83] × Gordlinia grandiflora Poor hybrid viability and reduced fertility; hybrids exhibit malformed and non-functional pollen. Controlled pollination (emasculation + stored pollen), seed stratification and culture to obtain viable seedlings; vegetative propagation used to maintain hybrid lines. [93] Case studies illustrating pre-zygotic barriers
-
Distant hybridization between Catalpa and Catalpopsis is frequently repressed by pre-zygotic barriers, particularly pollen–pistil incompatibility that stops normal fertilization and seed formation. A Chinese patent described a strategy to overcome pre-zygotic barriers in distant hybridization by grafting parental branches onto dwarf Catalpopsis rootstocks, storing flower branches at low temperatures to preserve viability, applying mixed pollen from multiple paternal clones under high temperatures to enhance fertilization, and removing stamens from maternal flowers to prevent self-pollination. These techniques successfully bypassed stigma-pollen incompatibility, leading to viable hybrid seedlings and demonstrating effective management of pre-zygotic barriers through controlled pollination and floral manipulation[85].
Stigma receptivity in willow was investigated to optimize Salix × Populus crosses, revealing that the stigma itself constitutes a major pre-zygotic barrier[83]. Receptivity was extremely short, lasting only 1–2 d after anthesis, and Populus pollen showed markedly reduced germination on willow stigmas compared with conspecific pollinations. To overcome this barrier, it is recommended to pollinate on day 1 of anthesis. Using a high frequency of pollen also improves hybridization efficiency[84].
A study on pre-zygotic barriers in Populus identified species-specific and layered barriers in interspecific crosses. In Populus tremuloides (aspen), pollen from Aigeiros and Tacamahaca sections failed to germinate or penetrate the stigma, often forming callose deposits as a rejection marker. A similar barrier was observed in Populus trichocarpa, where pollen from the Leuce section germinated, but was arrested in the stigma before reaching the ovules[87]. Although these barriers initially appeared decisive, subsequent research demonstrated that they could be circumvented. Techniques such as the use of mentor pollen have provided ways to bypass stigma-level rejection[87], while treating stigmas with organic solvents has been shown to disrupt surface lipids and improve receptivity. Together, these approaches offer practical means to overcome incompatibility mechanisms, enabling successful hybridization between aspens and members of the balsam poplar and cottonwood groups[86,88].
Crosses between Salix viminalis and various Populus species represents one of the best-studied examples of intergeneric hybridization within Salicaceae[89]. In an intergeneric cross between Salix viminalis and Populus species, a strong pre-zygotic barrier was observed at the stylar level, where Populus pollen tubes germinated on the stigma but rarely penetrated through the style to reach the ovules. This stylar block could be partially bypassed by placental pollination, i.e., when styles were cut off, pollen germinated on the placentae, which allowed limited pollen tube entry into the micropyle and sporadic initiation of hybrid embryos[83].
These case studies highlight pre-zygotic barriers in distant hybridization, such as stigma-pollen incompatibility, limited stigma receptivity, and stylar blockage hindering crossability in forest trees. However, strategies like precise pollination timing, use of mentor pollen, stigma surface modification, and alternative techniques (e.g., placental pollination), can mitigate these barriers, expanding the genetic base for breeding. Despite progress, systematic studies on pre-zygotic mechanisms are limited, emphasizing the need for further cytological and molecular research to improve hybridization predictability.
Case studies illustrating post-zygotic barriers
-
The Leyland cypress (× Cupressocyparis leylandii) is a notable example of a spontaneous intergeneric hybrid between Cupressus macrocarpa and Chamaecyparis nootkatensis, first observed in the UK. This hybrid combines the rapid growth of C. macrocarpa with the cold tolerance of C. nootkatensis, making it ideal for use in hedges, windbreaks, and landscape planting[94]. Molecular studies have confirmed the hybrid origin of Leyland cypress, with genetic evidence showing contributions from both parent species, C. macrocarpa and C. nootkatensi[94].
Leyland cypress exhibits near-complete sterility, making seed production ineffective, a major post-zygotic barrier. This limitation was overcome through vegetative propagation, particularly cutting propagation, supported by wounding techniques and auxin treatments (e.g., IBA), enabling large-scale cultivation. As a result, Leyland cypress has become widely valued for its rapid growth, adaptability, and use in windbreaks, hedges, and timber production[90]. The sterility of Leyland cypress, while advantageous for maintaining uniformity, limits its genetic diversity and adaptability. This underscores the importance of vegetative propagation in its cultivation. Additionally, the hybrid's susceptibility to diseases like canker presents challenges that require careful management[95].
Chitalpa tashkentensis is an intergeneric hybrid between Catalpa bignonioides and Chilopsis linearis. This hybrid is primarily valued for its ornamental features, drought tolerance, large flowers, and extended blooming period[91]. Sterility is the post-zygotic barrier of hybrids, which is confirmed by DNA evidence, making it advantageous for ornamental planting since it does not produce unwanted seeds[91]. DNA sequencing has verified that it originated from a cross between Chilopsis linearis and a Catalpa species, although not definitively Catalpa bignonioides as once believed[96]. Propagation is typically achieved through cuttings, and recent research has explored methods to restore fertility through polyploid induction, showing experimental promise[92]. Polyploidy can restore fertility in Chitalpa tashkentensis. Oryzalin, a polyploidization agent, has been used to induce polyploidy in Chitalpa tashkentensis, which can restore fertility[91]. This process involves submerging meristems in an oryzalin solution, resulting in tetraploid and cytochimera plants. Tetraploid Chitalpa plants have shown sparse flowering, while cytochimeras flower profusely and have viable pollen, allowing for successful self-pollination and cross-pollination with Catalpa bignonioides to produce triploids[91,92]. Both propagation and embryo rescue support breeding. Propagation of Chitalpa tashkentensis is typically achieved through cuttings due to its sterility. However, embryo rescue techniques have been employed to germinate triploid and tetraploid embryos, enhancing the breeding program's success[91].
Despite the success in inducing polyploidy, challenges remain in achieving consistent fertility restoration across different hybrid lines. Reciprocal crosses with Chilopsis taxa have been unsuccessful, indicating a need for further research into compatible genotypes[91,92]. Future research may focus on developing new molecular markers and understanding the genetic pathways responsible for reproductive isolation, which could facilitate the creation of compatible hybrid lines[97]. The challenges in achieving consistent fertility restoration underscores the complexity of hybridization processes, but advances in molecular techniques and polyploidy research offer promising avenues for future development.
Another example is × Gordlinia grandiflora, an intergeneric hybrid between Franklinia alatamaha and Gordonia lasianthus in the family Theaceae. Earlier attempts at creating this hybrid produced seedlings that subsequently died, reflecting post-zygotic incompatibility that impaired hybrid viability[93]. Researchers successfully established viable hybrids through controlled pollination, where Franklinia flowers were emasculated and hand-pollinated with stored Gordonia pollen, followed by seed stratification and culture. The resulting plants displayed desirable traits, including semi-evergreen foliage, large white flowers, and vigorous growth[93]. However, cytological analysis revealed malformed pollen, indicating reduced fertility. To maintain the hybrid, vegetative propagation was used, demonstrating how post-zygotic barriers can be managed through careful pollination and clonal propagation for successful hybrid introduction[93].
One more successful case of overcoming post-zygotic barriers is the Salix × Populus hybrid. In hybrid crosses between Salix viminalis and Populus species, embryo abortion caused by defective endosperm development has been reported. This barrier was successfully circumvented through embryo rescue at the globular to cotyledonary stage, leading to the regeneration of viable hybrid plantlets[83].
These case studies reflect the spectrum of strategies and outcomes in distant hybridization. While some species, like Leyland cypress, have seen widespread application, others serve as important models for pushing the boundaries of reproductive compatibility in forest trees.
To provide a synthesized overview of key contributions to distant hybridization research, Table 3 summarizes representative studies across major tree genera, highlighting their principal findings and methodological advances. These examples complement the case studies above and illustrate how approaches such as polyploidy induction, embryo rescue, vegetative propagation, and controlled pollination have collectively broadened hybridization possibilities in forestry.
Table 3. Summary of selected studies illustrating major advances and methodological approaches in distant hybridization of forest trees.
Tree species/hybrid Key findings Main contribution to distant hybridization research Ref. × Cupressocyparis leylandii Spontaneous intergeneric hybrid with strong post-zygotic sterility; hybrids exhibit vigorous growth and adaptability. Demonstrated how complete sterility can be overcome through vegetative propagation (cuttings + auxin), enabling large-scale deployment of hybrid trees. [94,95] × Chitalpa tashkentensis Hybrid sterility and embryo abortion confirmed by cytology and DNA evidence; fertility partially restored by induced polyploidy. Provided a model system showing that polyploid induction (oryzalin) + embryo rescue can restore fertility and generate advanced hybrid progeny (triploids, tetraploids). [91,96] × Gordlinia grandiflora Early hybrid attempts failed due to post-zygotic inviability; viable hybrids later obtained via controlled pollination. Demonstrated that emasculation + stored pollen, seed stratification, and clonal propagation can overcome hybrid inviability and maintain partially sterile hybrids. [93] Salix viminalis × Populus spp. Embryo abortion caused by defective endosperm development; hybrid embryos rarely develop naturally. Showed that embryo rescue at globular–cotyledonary stages can recover viable intergeneric hybrid plantlets. [83,89] Populus intersectional crosses (Leuce × Aigeiros; Leuce × Tacamahaca) Strong stigma-level rejection; pollen fails to germinate or tubes arrest early due to incompatibility. Identified species-specific incompatibility patterns and demonstrated effective use of mentor pollen and stigma solvent treatments to bypass pre-zygotic barriers. [86,87] Catalpa × Catalpopsis Severe pollen–stigma incompatibility prevents normal fertilization; natural hybridization almost impossible. Developed a multistep method involving grafting, temperature-controlled floral storage, mixed pollen under high temperature, and emasculation to achieve viable seedlings. [85] Populus alba × Populus euphratica Early embryo inviability due to endosperm failure; natural seed recovery very low. Demonstrated high success of ovule culture (> 90% plantlet recovery), highlighting ovule culture as a powerful tool for wide Populus crosses. [60] Eucalyptus interspecific hybrids Reduced hybrid viability in early generations due to developmental incompatibility. Provided evidence that post-zygotic inviability is widespread in distant tree hybrids; emphasized need for embryo rescue and large-scale selection. [11, 67] -
While progress in distant hybridization of forest trees has been substantial, several fundamental limitations continue to restrict its large-scale application. These challenges span biological, technical, ecological, and infrastructural domains[98].
Forest trees typically have long juvenile phases and extended generation times, which delay breeding cycles, hybrid evaluation, and trait stabilization. This severely limits the use of time-sensitive technologies such as CRISPR/Cas9, which require multiple generations to integrate and assess stable genetic modifications[8,9]. Although efforts to induce early flowering using transgenes like FT (Flowering locus T) have been explored in Populus and Eucalyptus, these approaches remain under development and are not yet widely transferable[7]. This bottleneck also complicates the use of molecular breeding tools, as phenotypic validation becomes difficult when traits are expressed late in the tree's life cycle.
In vitro techniques such as embryo rescue and in vitro fertilization (IVF) offer valuable tools for overcoming hybrid inviability in forest trees, particularly in wide or intergeneric crosses[78]. However, their broader application remains limited due to species-specific requirements, labor-intensive procedures, and technical complexities[70,89]. Challenges in transitioning these methods from lab to field, including the need for specialized infrastructure and the lack of cost-effective transfer protocols, further restrict scalability[99,100]. Potential solutions lie in automation, integration with genetic engineering and somatic hybridization, and a deeper understanding of underlying molecular mechanisms[101,102]. Continued innovation is crucial to improve the efficiency, accessibility, and ecological sustainability of in vitro forest tree breeding. Genomic progress is hindered by complex, repetitive, and heterozygous genomes, which complicate analysis and gene identification. While reference genomes exist for some species (e.g., Eucalyptus, Populus), many ecologically important species lack such resources[102].
In addition, the integration of genomic data into practical breeding remains weak due to gaps in genotype–phenotype correlation, limited marker validation, and the lack of scalable genomic selection frameworks. Expanding long-read sequencing, pan-genomic analysis, and trait-associated marker libraries could help address these challenges[13,102].
Ecological inviability limits hybrid success in natural environments. Hybrids that perform well under nursery conditions often show poor survival or maladaptation in natural ecosystems. Ecological inviability arises when hybrid phenotypes are intermediate, or fail to meet the selective demands of either parental habitat[15]. Long-term field trials are essential to assess hybrid fitness, yet these are often underfunded and time-consuming. Moreover, improper deployment of poorly adapted hybrids can result in ecological risks such as invasive potential, genetic introgression into wild populations, or reduced ecosystem stability. Integrating genomic and ecological data into predictive models could support more responsible deployment of hybrids under changing climatic conditions[16].
Despite all these limitations and gaps in research, the economic importance of overcoming these barriers cannot be overstated. Distant hybrids often exhibit heterosis, leading to faster growth, higher biomass, and improved yields under diverse environments[71,103]. In Populus and Eucalyptus, hybrids are used in plantations to increase wood production and reduce harvest cycles, supporting industries like timber, pulp, and bioenergy[104,105].
Disease resistance, such as to cypress canker or Dutch elm disease has been introgressed via hybridization, reducing chemical input needs and improving resilience[106,107]. Hybrids bred for drought, cold, and poor soil tolerance enable forestry on marginal lands, exemplified by Populus simonii × Populus nigra in northern China[108]. Their broader climate adaptability positions them as vital tools for future reforestation and restoration efforts[109]. Moreover, hybrid trees serve multiple industries, bioenergy, phytoremediation, landscaping, thanks to traits like fast growth and pollutant tolerance, making them ideal for short-rotation biomass crops and multifunctional plantations[110,111].
In brief, while the scientific and technical barriers to distant hybridization are significant, the economic and ecological payoffs justify continued investment and innovation. An integrative approach blending genomics, biotechnology, and ecological planning is essential to fully realize the potential of distant hybrids in modern forestry.
-
Advancing distant hybridization in forest trees requires a shift from isolated techniques toward a co-ordinated, interdisciplinary framework (Fig. 3). Genomic screening, including QTL mapping, GWAS, and marker-assisted selection can identify compatibility markers and adaptive traits at early stages[1,71,80]. These approaches not only accelerate parent selection, but also reduce the uncertainty of long breeding cycles. When coupled with biotechnological tools such as embryo rescue, in vitro fertilization, and polyploid induction, they provide direct means of bypassing reproductive barriers and improving hybrid viability[112]. In parallel, ecological modeling and adaptive trait mapping offer predictive insights into hybrid performance under diverse and changing environments[113]. Finally, accelerated breeding systems, incorporating early flowering induction, high-throughput phenotyping, and speed breeding, address the critical bottleneck of long generation times, and enable more rapid scaling of promising hybrids[114].
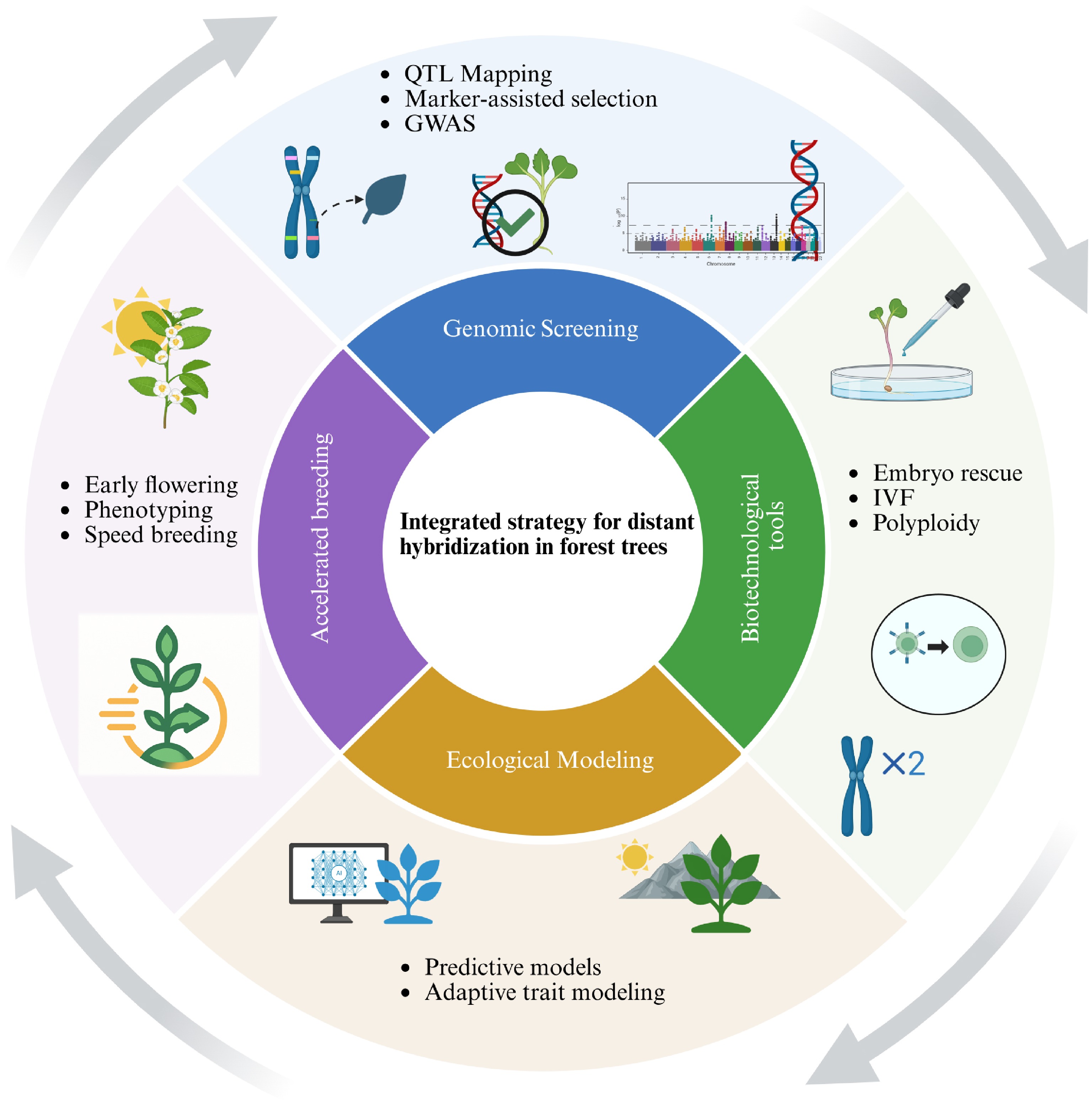
Figure 3.
Integrated strategy for distant hybridization in forest trees. This schematic outlines a multidisciplinary approach combining genomic screening (e.g., QTL mapping, GWAS, marker-assisted selection), biotechnological tools (e.g., embryo rescue, in vitro fertilization, polyploidy induction), ecological modeling (e.g., predictive and adaptive trait modeling), and accelerated breeding (e.g., early flowering, phenotyping, speed breeding). Together, these strategies enhance the success and efficiency of distant hybridization efforts in forest tree improvement programs.
The main challenge lies in integrating these diverse tools into a cohesive and scalable system that bridges experimental success with real-world applications. Despite promising advances, current progress remains fragmented, with limited predictive capacity across ecological contexts[113,115]. Persistent questions include why some species resist hybridization despite genomic compatibility, why hybrids that thrive in controlled settings often fail under natural conditions, and how to anticipate long-term ecological impacts[68]. The lack of integrative models that unify molecular, ecological, and climatic datasets continue to restrict reliable forecasting of hybrid performance, hindering practical deployment in forest management.
Future research must therefore prioritize the development of predictive, field-ready strategies that unite genomics, biotechnology, ecology, and accelerated breeding into a single operational framework (Fig. 3). Expanding genomic resources for non-model tree species, refining embryo rescue and IVF protocols for greater efficiency, and applying genome editing platforms such as CRISPR/Cas9 will be central to this effort. CRISPR applications in forest trees are still emerging, but genome-editing studies demonstrate strong potential for improving compatibility and accelerating breeding. CRISPR/Cas9 enables precise gene knockout in poplar, providing a direct route to modify reproductive traits and identify clear targets for future editing[116].
Equally important are long-term, multi-site field trials and decision-support systems that integrate molecular and climatic data, ensuring that hybridization strategies are ecologically viable as well as genetically innovative. Collaboration across disciplines, linking geneticists, ecologists, and forest managers will be crucial for transforming distant hybridization from a fragmented research endeavor into a forward-looking, climate-adaptive breeding paradigm capable of building resilient and genetically diverse forests.
-
The authors confirm their contributions to the paper as follows: study conception and design: Lan X; data collection: Bukhari S, Shi J, Gong M; analysis and interpretation of results: Bukhari S, Shi J; draft manuscript preparation: Bukhari S, Shi J, Chen S, Lan X. All authors reviewed the results and approved the final version of the manuscript.
-
All data analyzed during this study are included in this published article.
-
The authors declare that they have no conflict of interest.
-
#Authors contributed equally: Syeda Alia Zehra Bukhari, Jiaqi Shi
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Bukhari SAZ, Shi J, Gong M, Chen S, Lan X. 2026. Breaking barriers: advances and prospects for distant hybridization in forest trees. Forestry Research Advances 1: e004 doi: 10.48130/fra-0026-0001 

Breaking barriers: advances and prospects for distant hybridization in forest trees
- Received: 31 October 2025
- Revised: 17 December 2025
- Accepted: 06 January 2026
- Published online: 31 March 2026
Abstract: Distant hybridization, the crossing of genetically divergent tree species, genera, or families, offers transformative opportunities for enhancing genetic diversity, stress resilience, and climate adaptability. However, its large-scale application remains restricted by pre- and post-zygotic barriers that impede cross-compatibility and hybrid viability. This review synthesizes current understanding of these barriers, including reproductive asynchrony, pollen-pistil incompatibility, hybrid inviability, sterility, and ecological maladaptation. Here, conventional methods such as controlled pollination, mentor pollen, style manipulation, and polyploid induction are critically evaluated, alongside advanced biotechnological approaches including embryo rescue, in vitro fertilization, genomic selection, and genome editing. Case studies from Populus, Cupressus, and Catalpa illustrate how combining classical and molecular strategies has expanded the hybridization landscape while underscoring persistent species-specific challenges. Emerging genomic tools, predictive ecological modelling, and early flowering systems present new avenues to accelerate breeding cycles and improve hybrid performance under changing climatic conditions. This review proposes an integrative framework linking genomic screening, cytological interventions, and ecological modelling to overcome complex barriers and ensure ecological sustainability. Future research should focus on expanding genomic resources for non-model species, optimizing embryo culture protocols, and leveraging CRISPR-based gene editing to incompatibility systems. By bridging molecular genetics, physiology, and applied silviculture, distant hybridization can be transformed from a technical challenge into a strategic solution for forestry breeding.
-
Key words:
- Distant hybridization /
- Reproductive barriers /
- Forest tree breeding /
- Embryo rescue /
- Polyploidy /
- Hybrid vigor