










-
Cotton is a globally important cash crop. As an eco-friendly variety, naturally colored cotton (NCC), exhibits desirable traits such as stress and disease resistance, environmental friendliness, energy efficiency, antibacterial properties, and UV protection, offering significant potential for the textile industry[1]. Brown fiber is the most common NCC type, but its application is limited by lower yield and inferior fiber quality compared to white cotton[2,3]. Therefore, elucidating the molecular regulatory mechanisms of pigment biosynthesis and understanding the interplay between pigmentation and fiber development are critical for overcoming these bottlenecks.
Previous studies have established that the pigments in brown cotton fibers are primarily proanthocyanidins (PAs), whose biosynthesis is dependent on the flavonoid pathway. Silencing GhCHS2, GhLAR1, GhANR, or GhDFR1 resulted in reduced pigment accumulation and light fiber color, whereas overexpression of GhANR or GhLAR enhanced PA accumulation[4,5]. Notably, LAR and ANR are key enzymes determining the specific biosynthesis of PAs[6].
PA biosynthesis is primarily regulated at the transcriptional level. Among major plant transcription factor families, MYB proteins play the most critical role in modulating the flavonoid pathway. The first identified MYB regulator of PAs was Arabidopsis Transparent testa 2 (AtTT2)[7]. MYB proteins typically form a ternary MBW complex with bHLH and WD40 proteins[8−11]. In this complex, MYB proteins (e.g., AtTT2) play a central role, capable of activating PA pathway gene expression either independently or synergistically with bHLH, while WD40 proteins primarily serve as a scaffold, facilitating complex assembly[7,12,13]. To date, numerous MYB transcription factors have been identified as core regulators of anthocyanin and PA biosynthesis in various plants. For instance, in peach, PpMYB18 inhibited the activity of the MBW activation complex by competing for bHLH binding, thereby preventing excessive pigment accumulation[14]. In banana, MaMYBPA1/2 formed functional MBW complexes with MaMYCs and MaTTG1 to promote PA synthesis, while MaMYBPR1-4 interacted with MaMYCs to form inhibitory complexes; notably, activators could induce inhibitor expression, regulating PA synthesis through competitive binding[15]. In Vaccinium, expression of VaMYBPA1.1 required direct activation by VaMYBA1 and VaMYBPA2[16]. Additionally, in kiwifruit, the MYB activator AcMYB110 was targeted and suppressed by miR828, indirectly modulating anthocyanin accumulation[17]. In cotton, GhmiR858 negatively regulated PA accumulation by targeting GhTT2L[18]. Collectively, these findings demonstrate that PA biosynthesis in plants is governed by a highly sophisticated regulatory network involving a multitude of transcriptional activators and repressors.
In cotton, two MYB transcriptional activators, GhTT2-3A and GhMYB113 have been reported to regulate PA biosynthesis. GhTT2-3A, a homolog of Arabidopsis TT2 (AtTT2), cooperates with GhbHLH130D to activate PA biosynthetic genes and modulate brown pigmentation[19]. The Re gene, responsible for the cotton red foliated mutant phenotype, was identified as GhMYB113. When expressed under the control of the CaMV 35S promoter, the transgenic plants produced red leaves but white fibers; however, fiber-specific expression of GhMYB113 led to brown fiber formation. GhMYB113 interacted with PIF4/TT8 to activate the anthocyanin/PA biosynthetic pathway[20]. To comprehensively identify genes involved in pigment regulation in cotton, extensive studies have been conducted using a range of omics-based approaches. For instance, Canavar et al. analyzed the expression levels of flavonoid synthesis-related genes in three naturally colored cotton fibers[21]. Xiao et al. compared differentially expressed genes between developing brown and white cotton fibers using digital gene expression profiling, quantitative real-time PCR, and liquid chromatography-mass spectrometry (LC-MS)[22]. Sun et al. performed a comparative transcriptomic and biochemical analyses between wild brown-fiber cotton (Gossypium stockii), and white-fiber cotton (Gossypium arboreum)[23]. Li et al. conducted metabolomic and gene expression analyses on green, brown and white cotton fibers, while another study performed comparative proteomic analysis on near-isogenic lines of brown and white cotton fibers at 12, 18, and 24 d post-anthesis (DPA)[24]. Zhang et al. conducted metabolomic and gene expression analyses on brown, and white cotton fibers[25]. Wang et al. integrated proteomic and metabolomic analyses of fibers (10, 20, and 30 DPA) from the brown cotton variety 'Zong 1-61' (Z161) and white cotton variety 'RT'[26]. These multi-omics analyses have identified numerous transcription factors potentially involved in pigment regulation, such as MYB, bHLH, NAC, WRKY, TCP, and bZIP, etc. However, direct experimental evidence displaying their actual involvement in pigment synthesis remains lacking.
Given the pivotal role of R2R3-MYB proteins in regulating the flavonoid pathway, this study focused on MYB transcription factors. By integrating multi-omics data, we identified MYB genes differentially expressed between brown and white cotton fibers. Transient expression assays in tobacco leaves further identified 10 MYB genes capable of altering pigmentation. Functional characterization demonstrated that multiple MYB repressors and activators coordinately regulate PA synthesis. Our findings enrich the regulatory network of PA synthesis in cotton, provide new insights into its molecular control, and offer valuable genetic resources for improving fiber quality in brown cotton.
-
Tobacco (Nicotiana benthamiana) seeds were surface-sterilized and sown on half-strength Murashige and Skoog (½MS) medium. They were cultured in a growth chamber under controlled conditions (16 h light/8 h dark photoperiod, at 25 ± 1 °C) for 5−7 d. The seedlings were then transplanted into soil and grown for 4–6 weeks before Agrobacterium infiltration. Seeds of cotton (Gossypium hirsutum cv. Coker 312) were surface sterilized through sequential treatments: immersed in 70% (v/v) ethanol for 1 min, followed by 10% (v/v) hydrogen peroxide treatment for 1.5 h. After two rinses with sterile distilled water, the seeds were incubated at 28 °C overnight. The sterilized seeds were initially sown on ½MS medium plates and cultured for 3 d. Subsequently, the seedlings were transplanted into pots containing nutrient soil and grown in a growth room under controlled conditions (28 ± 2 °C, 16 h light/8 h dark photoperiod) for 7 d, after which they were transferred to the field.
Phylogenetic analysis
-
GhMYB protein sequences were retrieved from the Cotton database (CottonMD,
https://yanglab.hzau.edu.cn ). Arabidopsis MYB protein sequences were obtained from the TAIR database (TAIR,www.arabidopsis.org ), and MYB protein sequences from other species were acquired from GenBank (NCBI,www.ncbi.nlm.nih.gov ). The multiple sequence alignments were performed by Clustal W. A neighbor-joining phylogenetic tree of cotton MYB proteins, and homologous MYB proteins from other plants, was constructed using MEGAX with 1000 bootstrap replications. The tree was then optimized and visualized using iTOL (https://itol.embl.de/itol.cgi ).RNA extraction and quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis
-
Cotton bolls were tagged on the day of flowering and designated as 0 d post-anthesis (DPA). Bolls at 5, 10, 15, and 18 DPA were harvested. Fibers were rapidly separated from cotton ovules and immediately frozen in liquid nitrogen and stored at −80 °C. Total RNA was extracted from fibers using the RN53-EASYspin Plus Polysaccharide & Polyphenol/Complex Plant RNA Rapid Extraction Kit (Aidlab, Beijing, China). Total RNA was isolated from cotton calli using RNA Isolater Total RNA Extraction Reagent (Vazyme, Nanjing, China), according to the manufacturer's protocol. For each sample, 1 μg of RNA was reverse transcribed into cDNA using HiScript II 1st Strand cDNA Synthesis Kit (Vazyme, No. R211). qRT-PCR was performed using SYBR Green Reagent (Vazyme, Nanjing, China). The cotton gene GhUBI1 served as an internal reference gene. Each qRT-PCR value is the mean plus standard deviations (SD) of three independent experiments with three biological replicates. The primers used in this study are provided in Supplementary Dataset S1.
Subcellular localization and transcriptional activation activity analysis
-
The coding sequences of GhMYB308_A01, GhMYB3_A08, and GhMYB5_A11 were individually cloned into the pCAMBIA-2300-eGFP vector. The constructs were transformed into the Agrobacterium tumefaciens strain GV3101 individually. The Agrobacterium cells were resuspended in infiltration solution (10 mM MgCl2, 10 mM MES [2-(N-morpholino) ethanesulfonic acid] pH 5.7, and 150 mM acetosyringone) to an OD600 (optical density at 600 nm) of 1.0. The agrobacterial suspension was incubated at room temperature for 1−3 h and then infiltrated into the abaxial side of tobacco leaves using a needleless syringe. After 72 h of culture, GFP fluorescence in tobacco leaf epidermal cells was detected under a Leica TCS SP8 laser confocal microscope.
For transcriptional activation analysis in yeast, the full-length open reading frame (ORF) of GhMYB308_A01, GhMYB3_A08, and GhMYB5_A11 genes was individually cloned into yeast two-hybrid vectors pGBKT7 and pGADT7, respectively. Each pGBKT7-MYB recombinant vector was individually introduced into the yeast strain Y2H-Gold. The transcriptional activation activity assay was performed according to the BD Matchmaker Library Construction & Screening Kits User Manual (BD Biosciences Clontech, Palo Alto, CA, USA).
Plant transformation
-
For transient expression of GhMYB in tobacco, the coding sequence of each GhMYB was inserted into the pGreenII 62-SK vector under the control of the CaMV 35S promoter. Each vector was then transferred into the Agrobacterium tumefaciens GV3101 strain carrying the helper plasmid pSoup-P19 by electroporation. Agrobacterium cells were resuspended in an infiltration medium containing 10 mM MgSO4, 10 mM MES at pH 5.5, 150 μM acetosyringone to an optical density at 600 nm (OD600) of 1.0. The suspensions were placed at room temperature for 1.5 h, and then infiltrated into the abaxial side of tobacco leaves as previously described[27].
To construct the GhMYB overexpression vector, each coding sequence was subcloned into the pBI121 vector under the control of the CaMV 35S promoter, and then transferred into the Agrobacterium tumefaciens LBA4404 strain. Agrobacterium-mediated cotton transformation was performed as described previously[28]. The positive transgenic calli were confirmed by PCR.
Transactivation assay (TAA)
-
Sequences of 2-kb upstream of the open reading frames (ORFs) of PA pathway genes (GhLAR, GhDFR, and GhANR) were amplified from cotton genomic DNA. Reporter constructs were generated by inserting the promoters of these genes into the pGreen II 0800-LUC vector, with the Renilla (REN) luciferase (LUC) driven by the CaMV 35S promoter on the same vector serving as a reference to normalize transfection efficiency. The full-length CDS of GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 were cloned into the pGreenII 62-SK vector individually and used as effector vectors. The reporter and effector constructs, together with the helper plasmid pSoup-P19, were co-transformed into the Agrobacterium tumefaciens strain GV3101. Agrobacterium-mediated transformation was carried out on Nicotiana benthamiana leaves. After 72 h of culture, infected leaf regions were harvested, and LUC and REN activities were measured with a Dual-Luciferase Reporter Assay Kit (Promega, E1910). The detection of promoter activity was performed as previously described[29].
Yeast two-hybrid (Y2H) assay
-
Prior to performing the yeast two-hybrid (Y2H) assay, the BD-gene fusion construct was individually transformed into the Y2H-Gold yeast strain for the yeast autoactivation assay. The positive yeast transformants that grew normally on SD/-Trp dropout medium were spotted onto SD/-Trp/-His/-Ade triple dropout medium supplemented with X-α-Gal for growth and color development analyses. For strains with autoactivation activity, an appropriate concentration of 3-AT was screened and added to inhibit the autoactivation, after which the subsequent yeast two-hybrid assay was conducted. The coding sequence of GhMYB308_A01, GhMYB3_A08, and GhMYB5_A11 was individually cloned into the pGBKT7 vector to serve as a bait vector. Meanwhile, the pGADT7 vectors containing the full-length sequences of GhbHLH130D, GhMYB113, or GhTT2-3A respectively, were used as prey vectors. The bait vector and prey vector were co-transformed into the yeast Y2H-Gold strain. Positive clones were first identified by PCR and then plated on double dropout medium (SD/-Leu/-Trp, SD/-LW). The transformants were further spotted on quadruple dropout medium (SD/-Trp/-Leu/-His/-Ade, SD/-LWHA) to test their growth ability, following the method previously described[30].
The luciferase complementation imaging (LCI) assay
-
The luciferase complementation imaging (LCI) assay was performed as previously described[27]. The coding sequence of GhMYB308_A01, GhMYB3_A08, GhMYB5_A11, GhTT2-3A, GhMYB113, and GhbHLH130D was cloned into the JW771 and JW772 vector (BioVector NTCC, Beijing, China), respectively. Each ORF was fused to the N-terminus of luciferase (nLUC) or the C-terminal half of luciferase (cLUC) to generate GhMYB3_A08-nLUC, GhMYB5_A11-nLUC, GhMYB308_A01-nLUC, GhTT2-3A-nLUC, GhMYB113-nLUC and GhbHLH130D-nLUC, or GhMYB3_A08-cLUC, GhMYB5_A11-cLUC, GhMYB308_A01-cLUC, GhTT2-3A-cLUC, GhMYB113-cLUC, and GhbHLH130D-cLUC, respectively.
-
Wang et al. and Hinchliffe et al. conducted multi-omics studies using near-isogenic lines of brown and white cotton[26,31]. We focused on the R2R3-MYB transcription factors identified in these two studies that were potentially involved in the regulation of anthocyanin and PA biosynthesis. The two studies reported 96 and 21 MYB transcription factors, respectively (Fig. 1a; Supplementary Dataset S2). Furthermore, numerous MYB transcription factors regulating anthocyanin and PA biosynthesis have been documented in other plant species. Given the functional conservation of MYB proteins, we performed BlastP searches against the cotton genome using 78 anthocyanin- and/or PA-regulating MYB proteins from 34 other plant species (Supplementary Dataset S3). Regulatory functions of these 78 MYBs were classified into six major categories (Supplementary Datasets S4-S9). We identified 65 MYB homologs with > 50% amino acid identity (Fig. 1a; Supplementary Dataset S2). In total, we compiled a set of 145 candidate MYB proteins potentially involved in anthocyanin and PA regulation. Phylogenetic analysis further classified these 145 MYB proteins into subgroups SG1–26 and G1–6 (Supplementary Fig. S1).
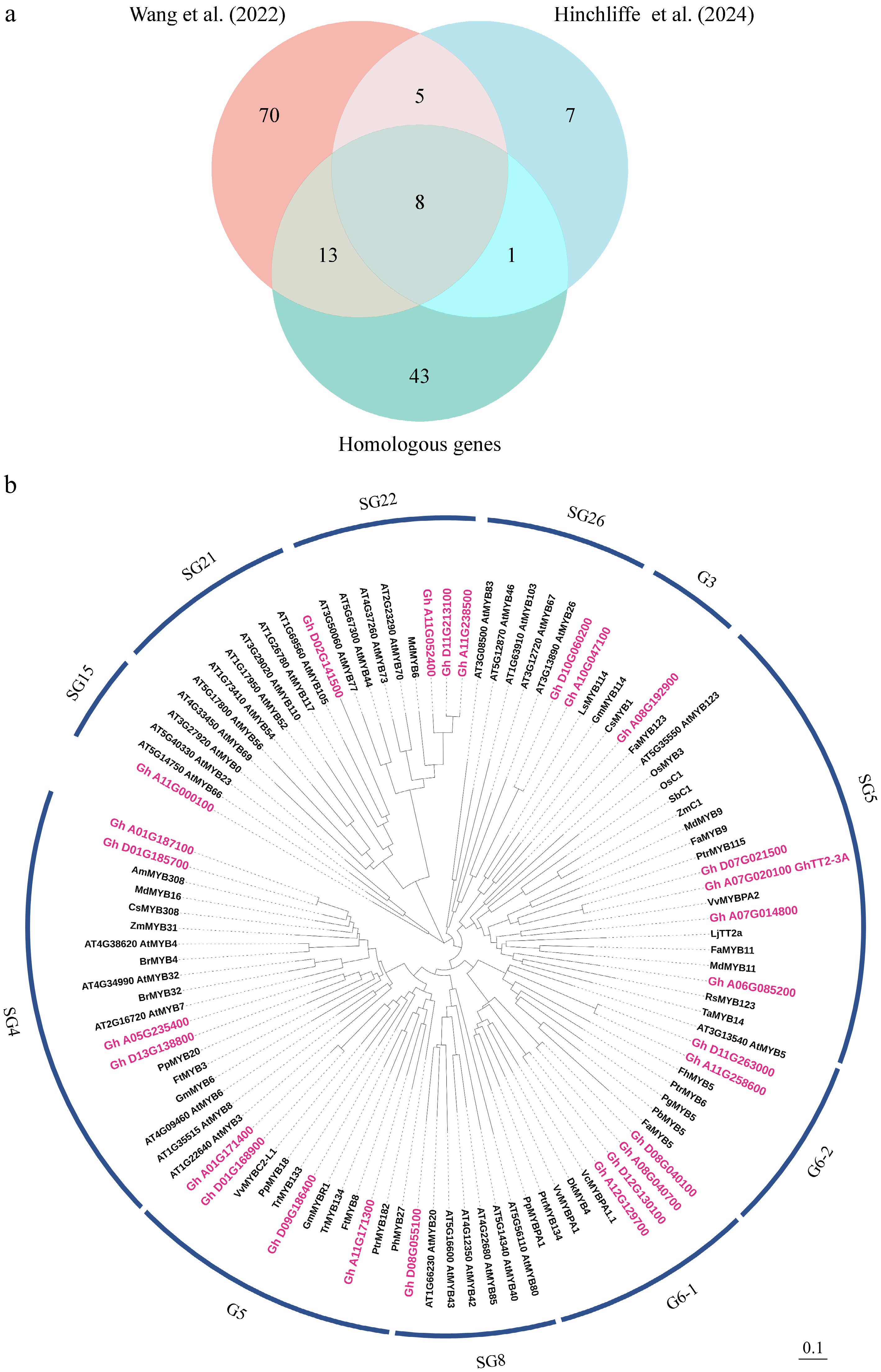
Figure 1.
Identification and phylogenetic relationship analysis of MYB genes related to anthocyanin/proanthocyanidin (PA) biosynthesis. (a) Venn diagram showing intersecting and overlapping candidate MYB transcription factors regulating anthocyanin/PA biosynthesis. MYB genes are derived from Wang et al.[26], and Hinchliffe et al.[31]. Homologous genes represent cotton homologs showing over 50% amino acid identity with known MYBs involved in the regulation of anthocyanin/PA biosynthesis from 34 other plants (Supplementary Dataset S3). (b) Phylogenetic analysis of 27 candidate cotton MYB proteins and reported anthocyanin/PA-related proteins from other plants. Candidate cotton MYB proteins are marked in red. Different clades are labeled based on homology with Arabidopsis proteins (SG1–26). G1 to G6 represent new subgroups. GenBank accessions of protein sequences are provided in Supplemental Dataset S2, S3. Scale bar indicates branch length.
To prioritize strong MYB regulators, we performed pairwise comparisons among the three MYB gene sets and selected intersecting and overlapping candidates. As shown in Fig. 1a, 27 MYB transcription factors were commonly identified across all comparisons. Phylogenetic analysis revealed that these 27 MYBs belong to distinct subgroups: SG4 (4), SG5 (4), SG8 (1), SG15 (1), SG21 (1), SG22 (3), G3 (1), G5 (4), G6-1 (4), and G6-2 (2) (Fig. 1b; Supplementary Table S1). These 27 MYBs represent strong candidates for further studies.
Expression analysis of 27 GhMYBs in white and brown cotton fibers
-
To investigate whether these genes are indeed associated with pigment synthesis, we first analyzed the expression of these 27 MYB genes in cotton fibers of brown and white near-isogenic lines at different developmental stages. Given that some genes belong to the A or D subgenomes of cotton, and considering the high sequence similarity between coding sequences of A and D subgenome gene pairs, we ultimately designed 19 pairs of primers to detect the expression of these genes respectively (Fig. 1a; Supplementary Dataset 1).
We first examined the expression of these genes in Z161 (brown cotton) and its near-isogenic line RT (white cotton) at 5, 10, 15, and 18 DPA fibers (Fig. 2a). qRT-PCR results revealed significant differential expression of these candidate MYB genes across over two developmental stages in RT/Z161 fibers, suggesting their potential roles in pigment regulation. The expression profiles displayed five patterns (I–V): six gene pairs showed downregulation in brown cotton at 2−4 stages (I), implying they are potential repressors; three gene pairs (including GhTT2-3A) showed upregulation in brown cotton at 3−4 stages (II), suggesting they are potential activators; four gene pairs showed initial upregulation followed by downregulation (III); four gene pairs showed initial downregulation followed by upregulation (IV); and three gene pairs exhibited a downregulation-upregulation-downregulation trend (V) (Fig. 2b). Very strangely, GhMYB113 expression was not detected in RT/Z161.
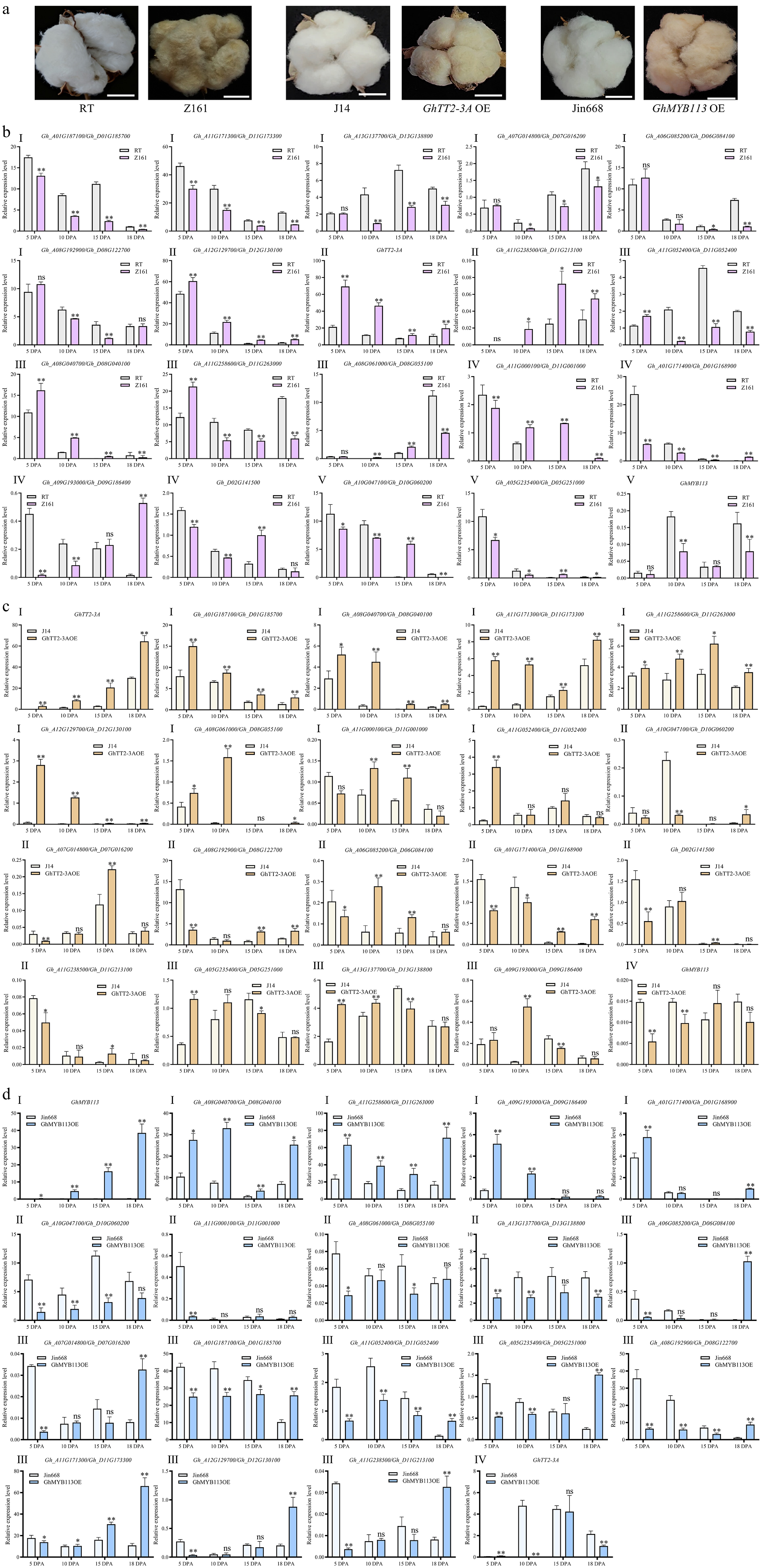
Figure 2.
Twenty-seven GhMYB genes exhibited differential expression in fibers of white vs brown cotton across four developmental stages (5–18 DPA). (a) Mature fibers of RT and its near-isogenic line Z161, J14, and GhTT2-3A- overexpressing line (GhTT2-3A OE), and Jin668 and GhMYB113- overexpressing line (GhMYB113 OE). Scale bar = 20 mm. (b) Expression analysis of 27 GhMYBs in RT (white cotton) and its near-isogenic lines Z161 (brown cotton) across the four fiber stages. The expression profiles displayed five patterns (I−V). (c) Expression analysis of 27 GhMYBs in J14 (white cotton) and GhTT2-3A overexpression line (brown cotton). The expression profiles were grouped into four types (I−IV). (d) Expression analysis of 27 GhMYBs in Jin668 (white cotton) and GhMYB113 overexpression line (brown cotton). The expression profiles were categorized into four types (I−IV). The cotton ubiquitin gene GhUBI1 was used as a reference gene for normalization. Data are presented as mean ± standard deviation (n = 3). Different letters above the bar graphs indicate significant differences determined by independent two-tailed t-test (GraphPad Prism 10). Independent two-tailed t-test was used for statistical analysis, where p < 0.05 is denoted by *, p < 0.01 is denoted by **, and no significant difference (p ≥ 0.05) is denoted by ns.
To determine whether these genes have regulatory relationships with GhTT2-3A, we also examined the expression of the 19 gene pairs in GhTT2-3A overexpression (OE) cotton (brown fiber), and its transformation recipient J14 (white fiber) across four fiber stages. The expression profiles were categorized into four types (I−IV): eight gene pairs showed upregulation at 1−4 stages (I) in brown fiber, seven gene pairs showed initial downregulation followed by upregulation (II), and three gene pairs showed the opposite pattern of initial upregulation followed by downregulation (III). Among these, eight gene pairs exhibited co-expression with GhTT2-3A, suggesting they are potentially regulated by GhTT2-3A (Fig. 2c). We further examined the expression of the 19 gene pairs in GhMYB113 OE cotton (brown fiber), and its transformation recipient Jin668 (white fiber) across four fiber stages (Fig. 2d). The expression profiles were categorized into four types (I-IV): four gene pairs showed upregulation at 2−4 stages (I) in brown fiber; four gene pairs showed downregulation at 1−4 stages (II), suggesting these eight gene pairs may co-regulate PA synthesis with GhMYB113; and nine gene pairs showed initial downregulation followed by upregulation (III) (Fig. 2d). As GhMYB113 expression was not detected in RT/Z161, we performed additional qPCR assays to assess GhMYB113 expression across multiple white and brown cotton varieties. Our results indicate that, unlike GhTT2-3A and GhbHLH130D, GhMYB113 transcript remains consistently low or nearly undetectable in fibers of both white and brown cotton varieties. In contrast, GhTT2-3A and GhbHLH130D display significantly higher expression in brown fibers than in their white counterparts across multiple developmental stages, suggesting that GhTT2-3A likely functions as a major PA-activating MYB transcription factor (Supplementary Fig. S2).
Overall, 12 genes were selected for subsequent analyses based on two criteria. First, their expression in cotton fibers across different developmental stages in RT (white cotton) was consistently upregulated or downregulated compared to those in Z161 (brown cotton). Second, their expression showed co-expression with either GhTT2 or GhMYB113.
Transient color assay of candidate MYB in tobacco leaves
-
To investigate the functional roles of the 12 candidate MYBs, we transiently expressed each MYB in Nicotiana benthamiana leaves and assessed anthocyanin or PA accumulation. Transient co-expression assays revealed that co-infiltration of GhTT2-3A with GhbHLH130D induced PA deposition, visualized as brown pigmentation after DMACA staining (Fig. 3). Expression of GhMYB113 alone or in combination with GhbHLH130D produced visible magenta pigmentation (anthocyanins) prior to staining, and brown pigmentation (PAs) after DMACA staining (Fig. 3a-d), consistent with their roles as activators of anthocyanin/PA biosynthesis. Based on qRT-PCR analyses, Gh_A08G040700, Gh_A11G258600, Gh_A11G000100, and Gh_A08G061000 displayed coincident expression patterns in GhTT2-3A and GhMYB113 OE fiber lines. We first expressed these four MYBs individually in tobacco. The staining results showed that individual expression or co-expression with GhbHLH130D of the four MYB genes did not induce detectable pigment accumulation (Fig. 3a-d). However, when co-expressed with established regulatory complexes, distinct functional effects emerged. Gh_A08G040700 significantly enhanced pigment accumulation induced by GhTT2-3A + GhbHLH130D, and also intensified the brown deposition mediated by GhMYB113 alone or GhMYB113 + GhbHLH130D (Fig. 3a), indicating that Gh_A08G040700 acts as a positive regulator of both anthocyanin and PA biosynthesis, likely by potentiating MBW (MYB–bHLH–WD40) complexes formed by GhTT2-3A or GhMYB113 with GhbHLH130D. Conversely, Gh_A11G258600 (Fig. 3b), Gh_A11G000100, (Fig. 3c), and Gh_A08G061000 (Fig. 3d) markedly suppressed pigment deposition when co-expressed with these activator complexes, suggesting they function as repressors that interfere with MBW complex activity to inhibit anthocyanin and PA accumulation.
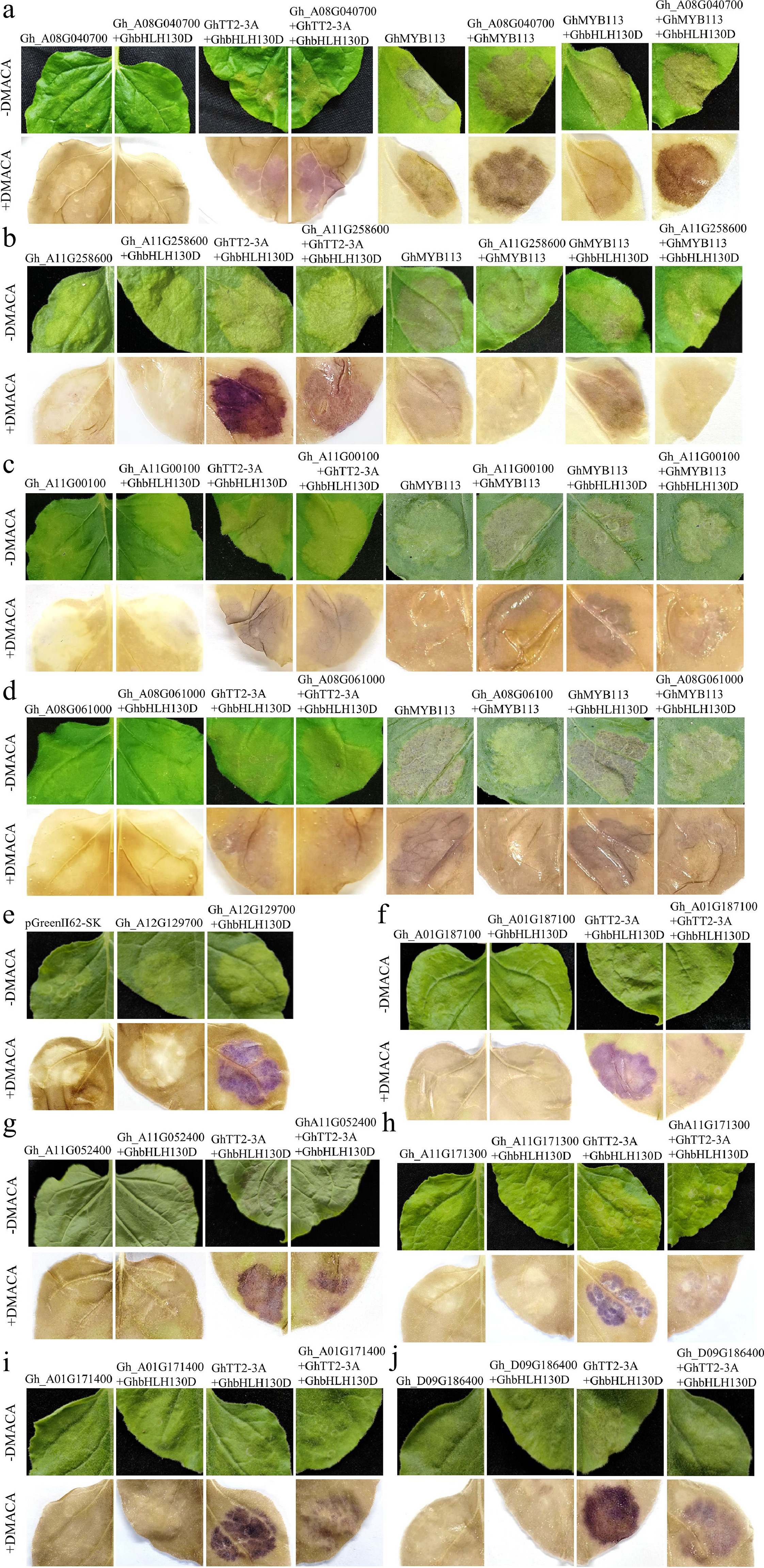
Figure 3.
Transient expression of GhMYB in tobacco leaves. (a) Co-expression of Gh_A08G040700 with GhTT2-3A and GhbHLH130D significantly enhanced pigment deposition. Gh_A08G040700 also augmented pigment accumulation induced by GhMYB113 alone or in combination with GhbHLH130D. Transient expression of GhTT2-3A alone or together with GhbHLH130D in tobacco leaves resulted in brown pigmentation following DMACA staining. Similarly, expression of GhMYB113 alone or in conjunction with GhbHLH130D leads to the production of light red anthocyanins, with brown substances becoming visible upon DMACA staining. The empty vector pGreenII 62-SK was used as the negative control. (b) Gh_A11G258600 significantly suppressed pigment accumulation induced by the co-expression of GhTT2-3A and GhbHLH130D. Co-expression of Gh_A11G258600 with either GhMYB113 alone, or in combination with GhbHLH130D also reduced pigment deposition. (c) Gh_A11G00100 reduced pigment accumulation resulting from the co-expression of GhTT2-3A and GhbHLH130D, and also suppressed pigment deposition induced by either GhMYB113 alone, or in combination with GhbHLH130D. (d) Gh_A08G061000 also significantly inhibited pigment deposition. (e) Co-transformation of Gh_A12G129700 with GhbHLH130D could induce pigment deposition. (f) Gh_A01G187100 significantly inhibited PA formed by GhTT2-3A and GhbHLH130D. (g) Gh_A11G052400 inhibited the deposition of PAs. (h) Gh_A11G171300 inhibited the deposition of PAs. (i) Gh_A01G171400 inhibited the pigment formed by GhTT2-3A and GhbHLH130D. (j) Gh_D09G186400 inhibited the pigment formed by GhTT2-3A and GhbHLH130D. Each experiment was repeated 3–5 times. The photographs were taken 7 d after infiltration. Original images are presented as Supplementary Fig. S3.
Among the remaining eight candidates, only Gh_A12G129700 induced pigment deposition when co-expressed with GhbHLH130D (Fig. 3e). Notably, co-expression of Gh_A01G187100 (Fig. 3f), Gh_A11G052400 (Fig. 3g), or Gh_A11G171300 (Fig. 3h) with GhTT2-3A + GhbHLH130D suppressed PA accumulation, implying their roles as repressors of PA biosynthesis. Similarly, Gh_A01G171400 (Fig. 3i) and Gh_D09G186400 (Fig. 3j) significantly attenuated pigment formation. In contrast, no significant effect on pigment accumulation was observed for Gh_A10G047100 (Supplementary Fig. S3k) and Gh_A13G137700 (Supplementary Fig. S3l) under the experimental conditions tested in this study. Overall, among the 12 candidate MYB transcription factors involved in anthocyanin and/or PA biosynthesis, we identified 2 activators, 8 repressors, and 2 had no detectable effect on pigment accumulation under our experimental conditions.
Subcellular localization and transcriptional activation analyses of GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01
-
To validate the functional roles of GhMYBs in regulating anthocyanin and proanthocyanidin biosynthesis, based on the phylogenetic relationship, expression profile, and transient tobacco expression results, we selected representative Gh_A08G040700 and Gh_A11G258600 from two distinct subclades within the G6 subgroup, and Gh_A01G187100 from the SG4 subgroup for further characterization.
Based on the phylogenetic tree and homology alignment shown in Fig. 1, we designated Gh_A01G187100 as GhMYB308_A01, Gh_A08G040700 as GhMYB3_A08, and Gh_A11G258600 as GhMYB5_A11, respectively. All three GhMYB proteins contain two consecutive R2 and R3 repeats, classified as typical R2R3-MYB transcription factors, and harbor the conserved motif required for interaction with GhbHLH130D. Additionally, GhMYB3_A08 possesses a C-terminal PA-Clade1 motif (Fig. 4a), GhMYB5_A11 contains a C1 motif (Fig. 4b), and GhMYB308_A01 harbors both C1 and C2 motifs (Fig. 4c). Subcellular localization assays confirmed that all three proteins are localized exclusively in the nucleus (Fig. 4d). Yeast activation assays further revealed that GhMYB3_A08 and GhMYB5_A11 exhibit transcriptional activation activity, whereas GhMYB308_A01 lacks autoactivation capability in yeast (Fig. 4e).
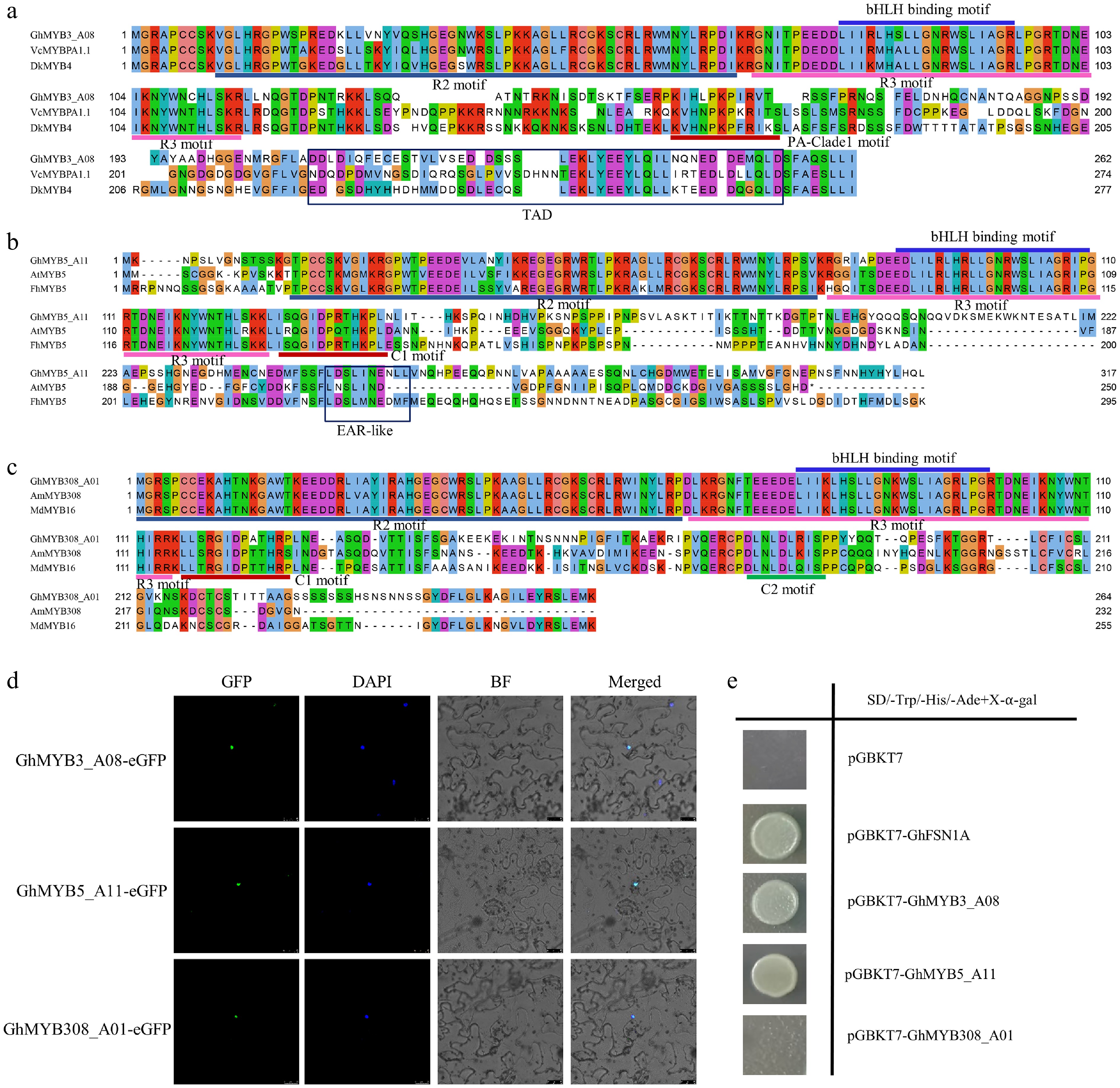
Figure 4.
Subcellular localization and transcriptional activation assays of GhMYB proteins. (a)–(c) Multiple sequence alignment analysis of GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 with other known plant MYB regulators related to anthocyanin/PA biosynthesis. Conserved residues and partial conservation are shown in different colors. The N-terminal R2 and R3 motifs, bHLH binding motif, and C-terminal C1 and C2 motifs are indicated with lines in different colors. TAD and EAR-like domains are indicated with black boxes. Protein sequences of VcMYBPA1, DkMYB4, AtMYB5, FhMYB5, AmMYB308, and MdMYB16 are listed in Supplementary Dataset S3. (d) Subcellular localization of GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 in tobacco leaves. GFP: green fluorescent protein. DAPI: nuclear dye. BF: bright field. Merged indicates overlaid image of GFP, DAPI, and bright field. Scale bar = 25 μm. (e) Transcriptional activation analysis of GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 in yeast. pGBKT7 serves as the negative control, and pGBKT7-GhFSN1A as the positive control.
Overexpression of GhMYB3_A08 promotes PA synthesis, whereas overexpression of GhMYB5_A11 or GhMYB308_A01 inhibits PA accumulation in cotton
-
We constructed overexpression vectors of GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01, respectively, and transformed each of them into cotton. Following selection on kanamycin-containing media and confirmation of transgene expression, we analyzed the phenotypes of transgenic calli lines and the control. DMACA staining revealed that GhMYB3_A08-overexpressing calli lines exhibited significantly darker pigmentation than the control, whereas GhMYB5_A11-overexpressing calli showed reduced staining intensity. Notably, GhMYB308_A01-overexpressing calli displayed markedly diminished staining, with pigment accumulation nearly abolished (Fig. 5c, e).
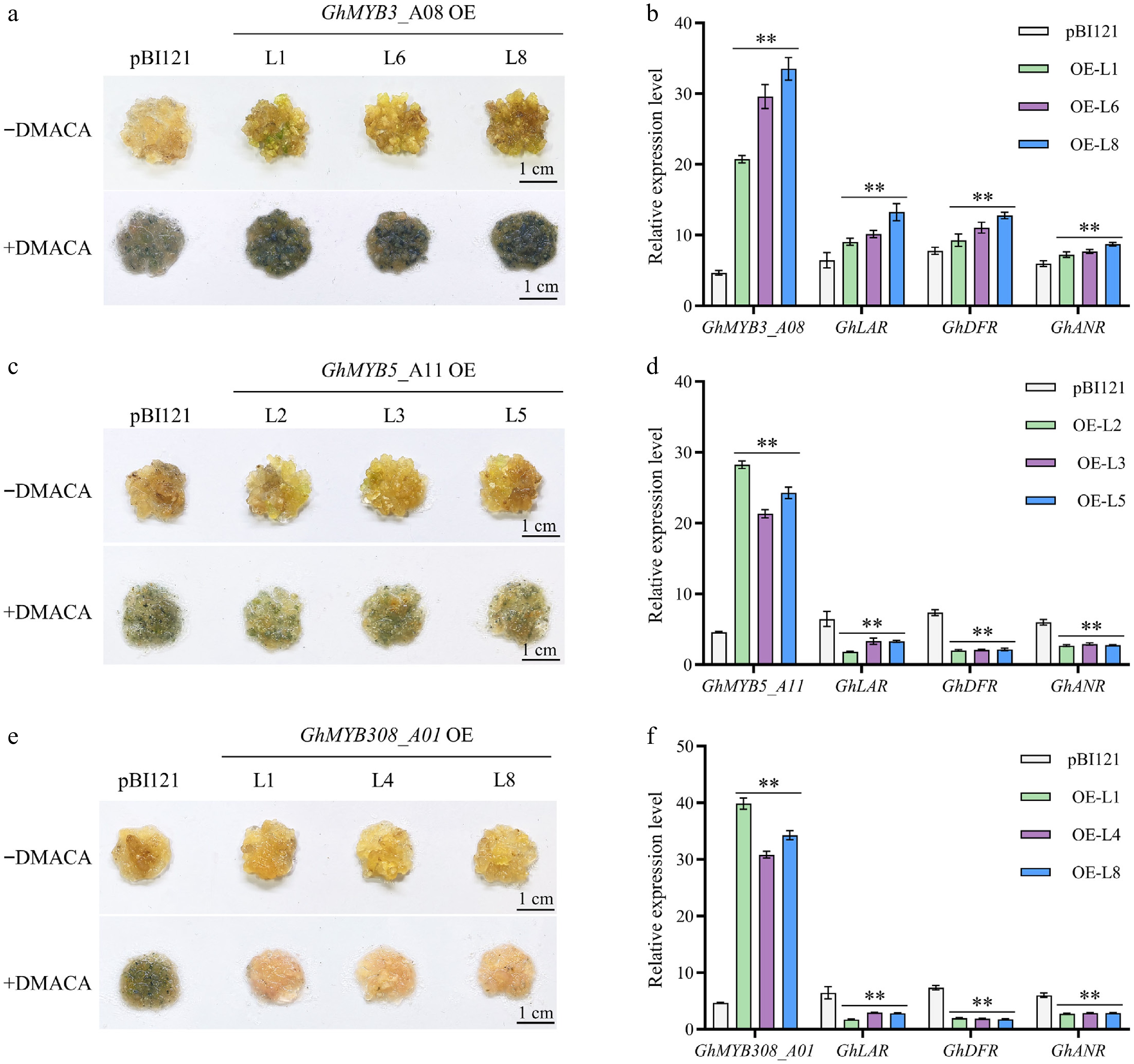
Figure 5.
Functional analysis of GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01. (a) Expression of GhMYB3_A08 in cotton calli led to enhanced PA accumulation compared to the control. −DMACA: callus before staining; +DMACA: callus after staining with 0.1% DMACA for 5 min. OE L1/L6/L8 represent three independent GhMYB3_A08 overexpression transgenic lines. pBI121 represents the empty vector control. (b) GhMYB3_A08 overexpression upregulated the expression of PA pathway genes GhLAR, GhANR, and GhDFR. (c) Overexpression of GhMYB5_A11 in cotton callus reduced PA accumulation. OE L2/L3/L5 represent three independent transgenic lines. (d) GhMYB5_A11 overexpression downregulated the expression levels of PA synthesis genes. (e) Overexpression of GhMYB308_A01 in cotton callus significantly reduced PA accumulation compared with the control. OE L1/L4/L8 represent three independent transgenic lines. (f) Expression of GhLAR, GhANR, and GhDFR were significantly decreased compared with the control. Values are presented as mean ± standard deviation, n = 3. Statistical analysis was performed using univariate t-test, and different letters indicate statistically significant differences (p < 0.05). Scale bar = 1 cm.
We further examined the expression of key PA biosynthetic genes GhLAR, GhDFR, and GhANR. In GhMYB3_A08-overexpressing lines, transcript levels of all three genes were significantly upregulated (Fig. 5b), indicating that GhMYB3_A08 promotes PA biosynthesis by activating downstream structural genes. In contrast, GhMYB5_A11- and GhMYB308_A01-overexpressing lines exhibited substantial downregulation of GhLAR, GhDFR, and GhANR compared to controls (Fig. 5d, f). Collectively, these results demonstrate that GhMYB3_A08 acts as a positive regulator, whereas GhMYB5_A11 and GhMYB308_A01 function as negative regulators of PA biosynthesis in cotton.
Transcriptional regulation of PA biosynthetic gene promoters by GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01
-
To further determine the transcriptional regulatory roles of GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 on promoters of cotton PA biosynthesis pathway genes, we performed transcriptional activation assays in Nicotiana benthamiana using a dual-luciferase reporter system. Results showed that GhMYB5_A11 and GhMYB308_A01 exerted varying degrees of transcriptional repression on the GhDFR promoter, and GhMYB5_A11 also suppressed the GhANR promoter. When co-expressed with GhMYB113, GhMYB3_A08 significantly enhanced the transcriptional activation of the GhLAR, GhDFR, and GhANR promoters compared to GhMYB113 alone, indicating a synergistic activation effect. In contrast, co-expression of GhMYB5_A11 with GhMYB113 markedly attenuated GhMYB113-mediated activation of the GhLAR, GhANR, and GhDFR promoters. Similarly, GhMYB308_A01 co-expression with GhMYB113 significantly reduced GhMYB113-driven activation of the GhLAR and GhANR promoters, demonstrating clear repressive effects (Fig. 6a, b). These findings indicate that distinct MYB transcription factors differentially regulate PA biosynthetic genes, likely through physical or functional interactions with GhMYB113, thereby either enhancing or suppressing promoter activity.
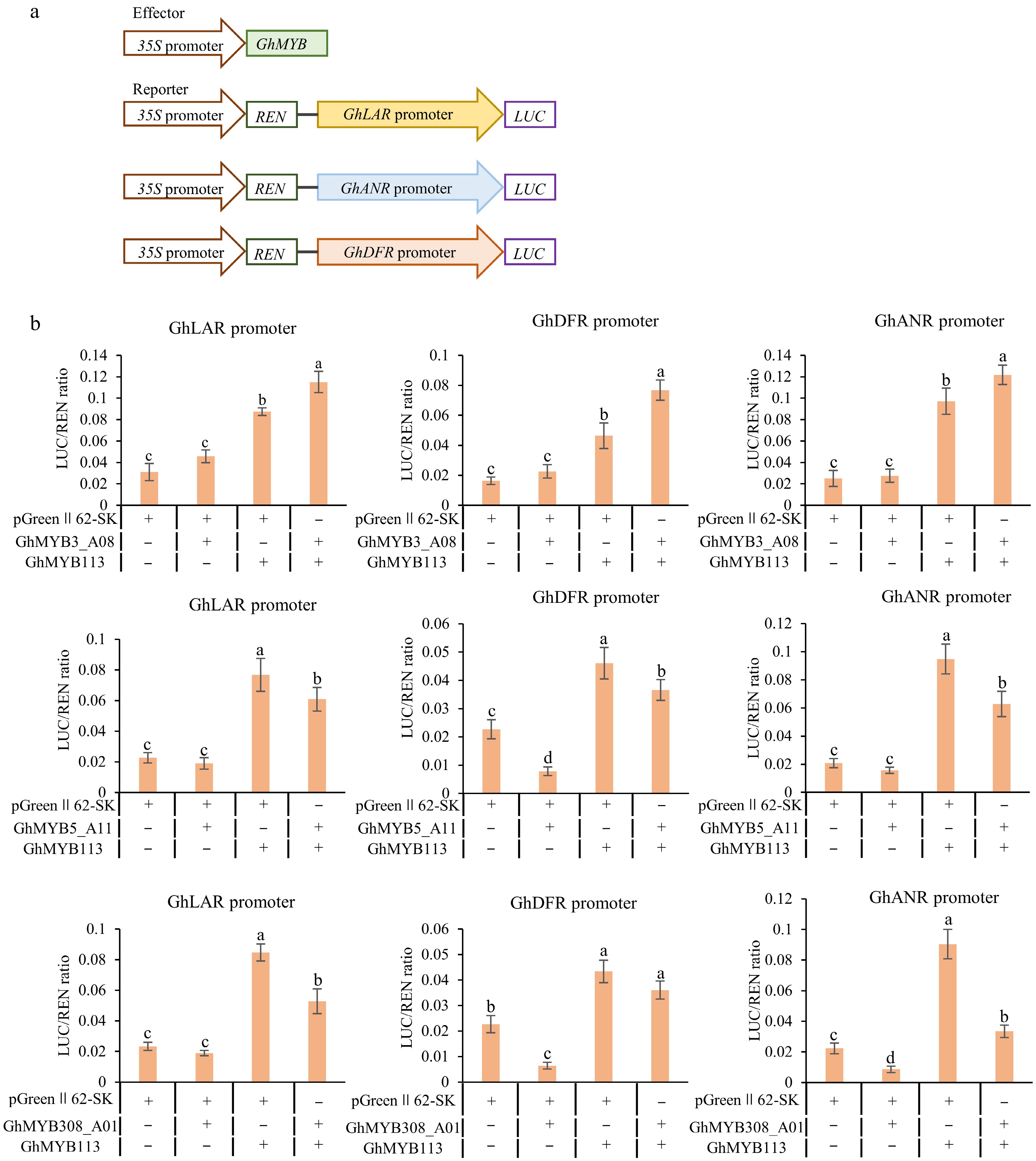
Figure 6.
Transcriptional activation analysis of GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01on promoters of GhLAR, GhANR, and GhDFR. (a) Schematic diagram of effector and reporter vectors used in transactivation assays. (b) Activation/repression effect of GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 on the promoters of three PA genes GhLAR, GhANR, and GhDFR. Values are presented as the mean ± standard deviation (SD) of three biological replicates. Statistical analysis was performed using one-way ANOVA combined with Tukey's HSD multiple comparison test, and different lowercase letters indicate significant differences between groups (p < 0.05).
Protein–protein interactions between MYB transcription factors and GhbHLH130D
-
The above results suggest that the three MYB proteins may exert their regulatory functions by interacting with the GhTT2-3A–GhbHLH130D or GhMYB113–GhbHLH130D complexes. To further elucidate the molecular mechanisms underlying the roles of GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 in the PA biosynthetic regulatory network in cotton, we first examined their protein–protein interactions using a yeast two-hybrid (Y2H) system. GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 were cloned into the pGBKT7 vector containing the DNA-binding domain (BD), while GhbHLH130D, GhTT2-3A, and GhMYB113 were cloned into the pGADT7 vector containing the activation domain (AD). The respective BD constructs were transformed into the yeast Y2H-Gold strain and plated on SD/-Trp/-His/-Ade medium supplemented with 25 mM 3-AT (Supplementary Fig. S4). Y2H results revealed that GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 each physically interacted with both GhTT2-3A and GhMYB113 (Fig. 7a, b). Additionally, GhMYB3_A08 and GhMYB5_A11 interacted with GhbHLH130D, whereas no interaction was detected between GhMYB308_A01 and GhbHLH130D (Fig. 7b), indicating functional divergence among these GhMYB members in their ability to associate with bHLH partners. Luciferase complementation imaging (LCI) assays further corroborated these interactions in planta. GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 all interacted with GhTT2-3A and GhMYB113, while GhMYB3_A08 and GhMYB5_A11 formed complexes with GhbHLH130D (Fig. 7c). Collectively, these results demonstrate that GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 modulate PA biosynthesis by forming protein complexes with GhTT2-3A and/or GhMYB113, with differential capacities to engage the bHLH partner GhbHLH130D.
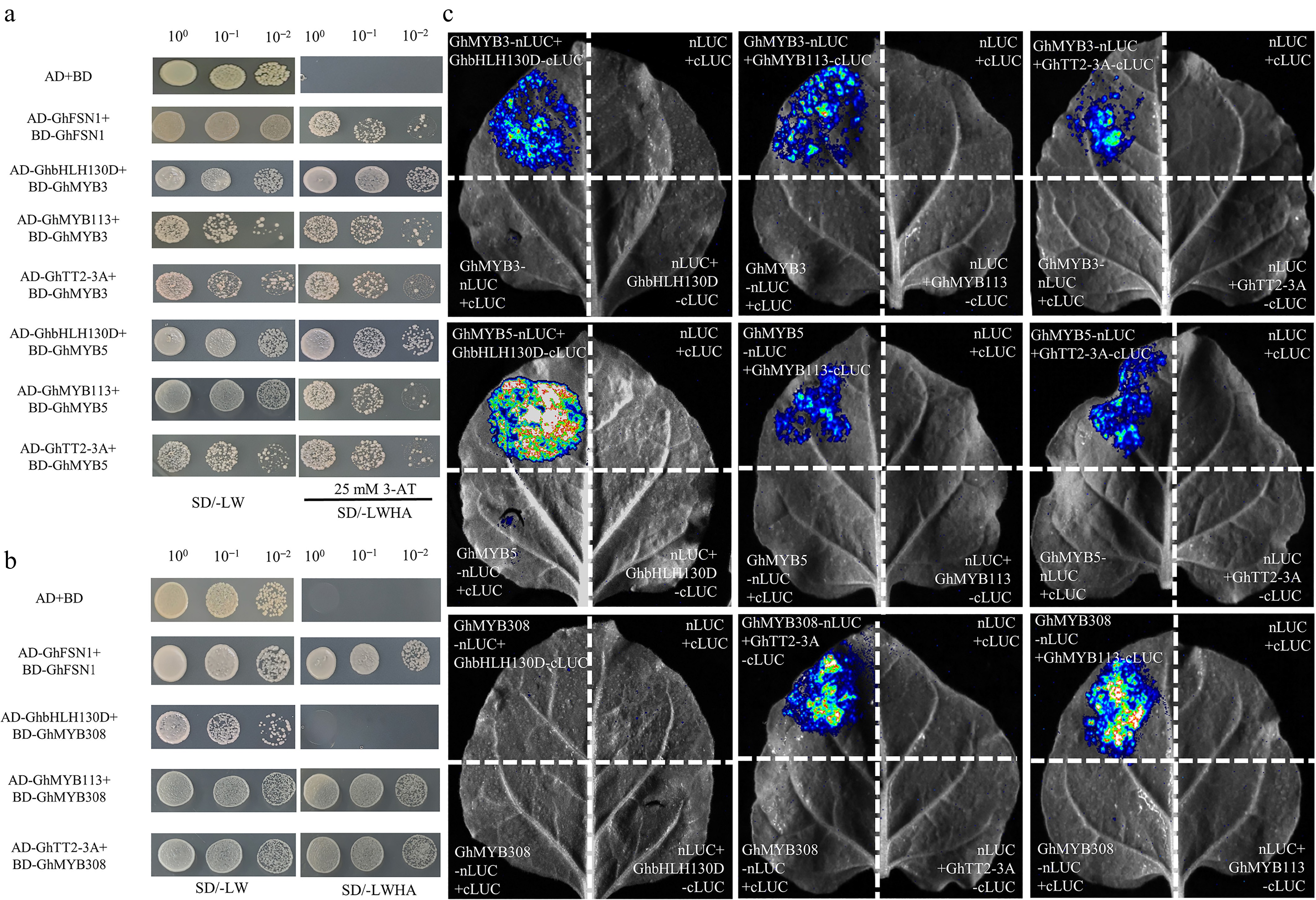
Figure 7.
Protein-protein interactions among GhMYB3, GhMYB5, GhMYB308, GhbHLH130D, GhTT2-3A, and GhMYB113. (a) Yeast two-hybrid (Y2H) assay revealing interactions among GhMYB3, GhMYB5, GhTT2-3A, GhMYB113, and GhbHLH130D. The combination of pGBKT7 (BD) + pGADT7 (AD) served as the negative control. AD-GhFSN1 + BD-GhFSN1 was used as the positive control. (b) Y2H demonstrated GhMYB3 and GhMYB5 interacted with GhbHLH130D, whereas no interaction was detected between GhMYB308 and GhbHLH130D. (c) Verification of in vivo protein-protein interactions by Luciferase Complementation Imaging (LCI) assay. Negative controls include nLUC + cLUC, GhMYB-cLUC + nLUC, and GhMYB-nLUC + cLUC. GhMYB3_A08, GhMYB5_A11, and GhMYB308_A01 are abbreviated as GhMYB3, GhMYB5, and GhMYB308, respectively.
-
Integrating multi-omics data, this study screened differentially expressed MYB genes between brown and white cotton fibers and identified eight MYBs that inhibit anthocyanin/PA deposition and two MYBs that promote pigment synthesis through functional analysis.
In various plant species, MYB repressors modulate anthocyanin/PA biosynthesis through diverse mechanisms[32,33]. For instance, PpMYB18 employs a dual repression strategy. First, it competes with MYB activators for binding to the bHLH interaction motif; second, it directly suppresses transcription of anthocyanin/PA biosynthetic genes via C-terminal C1/C2 repression motifs[14,34]. GhMYB5_A11 likely operates through a similar mechanism. MYB transcription factors in the SG4 subgroup primarily function as repressors of proanthocyanidin synthesis[35]. GhMYB308_A01, a member of the SG4 subgroup, contains a canonical bHLH-interaction motif; however, we did not detect physical interaction between GhMYB308_A01 and GhbHLH130D. Whether it interacts with other bHLH partners remains to be investigated[36]. This protein may instead repress PA biosynthetic genes via its C-terminal C1/C2 repression motifs. Notably, GhMYB308_A01 also interacts with GhTT2-3A and/or GhMYB113, potentially disrupting the function of the GhMYB113/GhTT2-3A–GhbHLH130D activator complex. Consistent with our findings, a recent study demonstrated that Chinese cabbage BrMYB32 attenuates anthocyanin biosynthesis by interfering with the formation of the MYB–bHLH–WD40 (MBW) activation complex[37] and repressing transcription of anthocyanin pathway genes, highlighting a fine-tuned regulatory mechanism mediated by activator–repressor interactions[38−41].
It is evolutionarily advantageous for plants to develop regulatory mechanisms that spatiotemporally and quantitatively modulate anthocyanin biosynthesis in response to diverse developmental, environmental, and hormonal signals[42]. This explains why multiple independent regulatory modules have evolved upstream of the MBW complex, each responsive to distinct stimuli. Such modular architecture prevents pleiotropic unnecessary anthocyanin production while preserving evolutionary flexibility. However, the question remains: why so many repressors? Specifically, why has evolution not favored a comparable number of independent activators to respond to internal and external cues? As noted by LaFountain & Yuan, the answer lies in the regulatory logic of a 'double-negative' mechanism: an input signal represses or degrades a repressor of the anthocyanin-activating complex, thereby activating biosynthesis[33]. Compared to simple or sequential positive regulation (e.g., direct activation of the MBW complex or its upstream activators), this design offers at least two advantages. First, positive regulation would necessitate that each MBW gene acquire numerous cis-regulatory elements or enhancers to integrate multiple signals, a constraint that is evolutionarily difficult to overcome. Double-negative logic circumvents this by allowing distinct repressive modules to evolve for stimulus-specific control, leaving the MBW complex itself unchanged. Second, double-negative regulation frequently involves the degradation or sequestration of repressors. De-repression at the protein level enables a significantly faster response to environmental or hormonal signals than transcriptional positive regulation, as it bypasses the need for de novo transcription and translation. Thus, the prevalence of repressors in anthocyanin regulation reflects an elegant evolutionary solution that balances precision, modularity, and responsiveness. In fact, the prevalence of MYB repressors is also observed in several other species. For instance, Deng et al. screened anthocyanin regulatory genes from leaves of sweetpotato and identified three MYB activators (IbMYB1, IbMYB2, and IbMYB3) and five MYB repressors (IbMYB27, IbMYBx, IbMYB4a, IbMYB4b, and IbMYB4c)[43]. Furthermore, in the dissection of the transcriptional regulatory network that fine-tunes PA biosynthesis in bananas, a total of six PA-specific MYB transcription factors were identified, four of which functioned as repressors[15].
As shown in Fig. 1 and Supplementary Fig. S1, the G3, G5, and G6 clades represent newly defined evolutionary subgroups that warrant attention, potentially possessing specialized roles in regulating anthocyanin/PA synthesis. Notably, genes within the G6 subgroup, including PpMYBPA1, PtrMYB134, and VvMYBPA1, have been shown to upregulate PA biosynthetic genes[44−46]. Similarly, Arabidopsis AtMYB5 and AtTT2 positively regulate PA accumulation in seed coats[47], suggesting that the six cotton MYBs clustered in this clade may share similar functions. Consistent with this, our results showed GhMYB3_A08 synergistically activates anthocyanin and PA accumulation (Fig. 1b), whereas GhMYB5_A11, also belonging to the G6 subgroup, inhibits pigment deposition. This functional divergence within the same evolutionary clade is very intriguing. First, the sequence alignment of the two proteins revealed distinct variations in their C-terminal regions. GhMYB5_A11 harbors a conserved EAR-like repressor motif (LxLxL) in its C-terminus, a canonical domain known to recruit co-repressors (e.g., TOPLESS family proteins) and suppress downstream gene expression[48,49]. By contrast, GhMYB3_A08 lacks this EAR motif; instead, its C-terminus contains a transactivation domain (TAD) enriched with acidic amino acids (e.g., Glu/E, Asp/D), which is critical for interacting with transcriptional co-activators (e.g., MEDIATOR complex subunits) and promoting gene transcription[50,51]. The functional divergence between GhMYB3_A08 and GhMYB5_A11 is likely attributable to differences in their C-terminal domain motifs. Additionally, given that GhTT2-3A and GhbHLH130D form an activation protein complex to synergistically promote the transcription of the promoter of structural genes for PA biosynthesis, thereby increasing PA content. A Luciferase complementation imaging (LCI) assay was performed to assess the impact of GhMYB3_A08 or GhMYB5_A11 on the interaction affinity between GhTT2-3A and GhbHLH130D. As shown in Supplementary Fig. S5, coexpression with GhMYB5_A11 resulted in significantly weaker luminescence compared to the control, indicating that GhMYB5_A11 attenuates the in vivo interaction between GhTT2-3A and GhbHLH130D. In contrast, coexpression with GhMYB3_A08 led to markedly stronger luminescence, suggesting that GhMYB3_A08 enhances the interaction between these two proteins (Supplementary Fig. S5). These results suggest that GhMYB3_A08 enhances PA synthesis by promoting the assembly of the activation complex, while GhMYB5_A11 decreases PA synthesis by inhibiting its formation. Based on this finding, together with protein-protein interaction and TAA results, we propose a working model to explain how activators and repressors interact to regulate PA biosynthesis. In this model, GhTT2-3A/GhMYB113 acts synergistically with GhbHLH130D and GhWD40 proteins to bind directly to the promoter regions of target genes, thereby activating PA pathway gene transcription. GhMYB3_A08 can enhance the transcriptional activation activity. In contrast, GhMYB5_A11 attenuates the interaction between GhTT2-3A and GhbHLH130D, and inhibits the formation of the activation MBW complex, ultimately resulting in the repression of target gene expression (Supplementary Fig. S6).
In the G3 subgroup, CsMYB1 promotes catechin biosynthesis[52], GmMYB114 indirectly influences anthocyanin synthesis, and LsMYB114 regulates flavonoid pathway genes[53,54], implying that Gh_A08G192900 in this subclade may perform a similar regulatory role. In the G5 subgroup, VvMYBC2-L1 and PtrMYB182 negatively regulate anthocyanin accumulation[55,56], while PpMYB18 represses both anthocyanin and PA synthesis. Consistently, our results demonstrated that Gh_A01G171400, Gh_D09G186400, and Gh_A11G171300 in the G5 subgroup significantly inhibited anthocyanin or PA deposition, supporting functional conservation within this subgroup.
Natural brown cotton derives its pigmentation from the accumulation of PAs. Recent studies have shown that enhancing GhTT2-3A expression in cotton results in shorter fibers, reduced yield, and darker brown pigmentation, whereas suppressing its activity leads to longer fibers and lighter coloration[25]. The MYB activators and repressors identified in this study provide precise genetic targets for genetic improvement of cotton fiber. Specifically, fiber-specific expression of activators could intensify pigmentation, while moderate induction of repressors may alleviate the inhibitory effect of PAs on fiber elongation, offering a promising strategy to simultaneously optimize both color and fiber quality.
To balance pigment biosynthesis and fiber development, PA synthesis must be governed by a sophisticated transcriptional regulatory network comprising both activators and repressors. Our findings significantly expand the known regulatory landscape of PA biosynthesis in cotton, and provide novel insights into the molecular mechanisms underlying this balance.
-
The authors confirm their contributions to the paper as follows: conceived and designed the research: Xu W; performed the experiments: Luo J, Luo Y, Gao Y, Cai J, Sun X; analyzed the data: Xu W, Liu L, Luo J; wrote the manuscript: Luo J, Xu W. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
-
This work was supported by the Key Research and Development Program of Xinjiang Uygur Autonomous Region (2024B02006-2) and National Natural Science Foundation of China (Grant No. 32372104).
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 The 27 MYB candidate genes identified as putative regulators of anthocyanin/PA biosynthesis.
- Supplementary Fig. S1 Phylogenetic analysis of cotton (Gossypium hirsutum) GhMYBtranscription factors and anthocyanidin/proanthocyanidin(PA)-related MYB proteins from other representative plant species.
- Supplementary Fig. S2 Expression profiling of GhMYB113, GhTT2-3A and GhbHLH130D in different cotton tissues and developing fibers.
- Supplementary Fig. S3 Transient expression of GhMYBin tobacco leaves.
- Supplementary Fig. S4 Optimization of autoactivation inhibition conditions prior to verifying protein-protein interactions between GhMYB3_A08, GhMYB5_A11, GhMYB308_A01 and GhbHLH130D, GhTT2-3A, GhMYB113.
- Supplementary Fig. S5 Protein–protein interaction affinities in N. benthamianawere analyzed by LUC complementation imaging assays.
- Supplementary Fig. S6 Proposed working model for the interplay between MYB activators and repressors in regulating proanthocyanidinbiosynthesis in cotton.
- Supplementary Dataset S1 Primers used in this study.
- Supplementary Dataset S2 Summary of candidate MYBs involved in anthocyanin/PA biosynthesis.
- Supplementary Dataset S3 Reported MYBs involved in regulating anthocyanin/PA biosynthesis from 34 plant species.
- Supplementary Dataset S4 MYB proteins that positively regulate anthocyanins biosynthesis and their cotton homologs.
- Supplementary Dataset S5 MYB transcription factors that negatively regulate anthocyanin biosynthesis and their cotton homologs.
- Supplementary Dataset S6 MYB proteins that positively regulate PA biosynthesis and their cotton homologs.
- Supplementary Dataset S7 MYB proteins that negatively regulate PA biosynthesis and their cotton homologs.
- Supplementary Dataset S8 MYB proteins that positively regulate both anthocyanins and PA biosynthesis and their cotton homologs.
- Supplementary Dataset S9 MYB proteins that negatively regulate both anthocyanins and PA biosynthesis and their cotton homologs.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Luo J, Luo Y, Liu Y, Gao Y, Cai J, et al. 2026. Multiple MYB repressors and activators coordinately modulate proanthocyanidin biosynthesis in cotton. Seed Biology 5: e014 doi: 10.48130/seedbio-0026-0010 

Multiple MYB repressors and activators coordinately modulate proanthocyanidin biosynthesis in cotton
- Received: 16 November 2025
- Revised: 27 January 2026
- Accepted: 12 February 2026
- Published online: 17 April 2026
Abstract: Naturally colored cotton (NCC) is a valuable germplasm resource for breeding, but brown cotton, the most common type, typically exhibits lower yield and inferior fiber quality than white cotton. Understanding the regulatory mechanisms of pigment biosynthesis is therefore crucial for improving fiber traits. Proanthocyanidins (PAs) in the flavonoid pathway are the primary pigments in brown fibers, and their biosynthesis is mainly regulated at the transcriptional level. Here, integrated transcriptomic and metabolomic analyses identified 145 MYBs potentially involved in pigment biosynthesis. Combining these with 78 known flavonoid-related MYBs from 34 other plant species, we narrowed down 27 candidate regulators. Expression profiling revealed 12 MYBs differentially expressed between white and brown fibers at different developmental stages, and strongly correlated with the expression of the known pigment synthesis activators GhTT2-3A and/or GhMYB113. Transient expression in tobacco revealed eight MYBs suppressed, two promoted, and two had no detectable effect on pigment accumulation under our experimental conditions. Functional validation with three representative MYBs in cotton demonstrated that GhMYB3_A08 enhanced PA deposition, while GhMYB5_A11 and GhMYB308_A01 suppressed it. GhMYB3_A08 enhanced GhMYB113-mediated activation of GhLAR and GhANR promoters, whereas the other two MYBs antagonized this activation. Protein interaction assays revealed that all three MYBs formed heterodimers with GhTT2-3A or GhMYB113, and GhMYB3_A08 and GhMYB5_A11 also interacted with GhbHLH130D. Together, our findings reveal that distinct MYB proteins act as activators or repressors of PA biosynthesis in cotton. Our results provide new insights into the transcriptional control of pigment accumulation and identify key genetic targets for improving brown fiber.
-
Key words:
- Cotton /
- MYB transcription factors /
- Proanthocyanidins /
- Repressors /
- Activators /
- Protein complex