









-
Rose (Rosa spp.) is one of the most important ornamental flowers worldwide, with significant economic value. Flower color, a key ornamental trait, substantially influences its market value[1,2]. Anthocyanins, a class of natural pigments widely distributed in both the vegetative and reproductive organs of plants, serve as the primary coloring compounds in rose petals[3,4]. The biosynthesis of anthocyanins occurs via the flavonoid pathway, catalyzed by a series of enzymes including chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), flavonoid 3′-hydroxylase (F3′H), dihydroflavonol 4-reductase (DFR), anthocyanidin synthase (ANS), and uridine diphosphate (UDP)-glucose: flavonoid 3-O-glucosyltransferase (UFGT)[5,6]. The expression of structural genes in the anthocyanin pathway is primarily regulated by three families of transcription factors: MYB, basic helix–loop–helix (bHLH), and WD40 Repeat protein (WDR). Among them, MYB and bHLH transcription factors can function independently or form the MBW protein complex with WDR to exert regulatory roles[7,8]. In addition to the canonical MYB regulators, other transcription factors such as WRKYs, NAM/ATAF/CUC Domain Transcription Factors (NACs), Ethylene Response Factors (ERFs), and HY5 have been reported to participate in the regulation of anthocyanin biosynthesis. These factors may act through binding directly to the promoters of anthocyanin structural genes, by regulating MYB expression, or through the formation of multicomponent complexes with MBW (MYB–bHLH–WDR) components[9−12]. On one hand, synthesis and accumulation of anthocyanin can be induced under various stress conditions, such as the light's quality and intensity, temperature, nutrient status (e.g., sugars, water, hormones, metal ions), and pathogen infection[13]. On the other hand, anthocyanins contribute to stress mitigation by chelating heavy metal ions, participating in redox reactions, and scavenging reactive oxygen species (ROS)[14].
WRKY transcription factors represent one of the largest families of transcriptional regulators in plants, and are extensively involved in stress resistance and immune responses[15,16]. They mediate adaptations to diverse biotic and abiotic stresses[17,18]. A defining characteristic of WRKY proteins is the presence of a highly conserved N-terminal WRKY domain (featuring the WRKYGQK motif), which enables specific binding to the W-box cis-element [(T)TGAC(C/T)], and a C-terminal zinc finger motif. These zinc fingers are primarily classified as C2H2 or C2HC types. According to the number of WRKY domains and the zinc finger structure, the family is categorized into three major groups: Group I (two WRKY domains and C2H2-type zinc fingers), Group II (one WRKY domain and C2H2-type zinc fingers, further divided into subgroups IIa–IIe), and Group III (one WRKY domain and C2HC-type zinc fingers)[19,20].
Accumulating evidence implicates WRKY members from various groups in the regulation of anthocyanin biosynthesis. Documented examples include the Group I members MdWRKY10 from apple (Malus domestica)[21], PyWRKY26 from pear (Pyrus pyrifolia)[10], and homologs of AtTTG2 in Arabidopsis thaliana, such as PhPH3 in petunia (Petunia hybrida)[22], StWRKY13 in potato (Solanum tuberosum)[23], LhWRKY44 in lily (Lilium hybrida)[24], and PpWRKY44 in peach (Prunus persica)[25]; the Group IIa member MdWRKY40[26]; the Group IIb members McWRKY71from bitter melon (Momordica charantia)[27] and PyWRKY31[10]; the Group IIc members MdWRKY50[28], FaWRKY71 from strawberry (Fragaria × ananassa)[29], and homologs of AtWRKY75[30] such as MdWRKY75[31] and CitWRKY75 from Citrus spp.[32]; the Group IId members MdWRKY11[33] and NtWRKY11 from tobacco (Nicotiana tabacum)[34]; and Group III members StWRKY70[35] and BnWRKY41-1 from rapeseed (Brassica napus)[36]. Despite extensive studies on WRKY-mediated anthocyanin regulation in other plants, the molecular mechanism underlying WRKY transcription factors regulating anthocyanin biosynthesis in Rosa chinensis remains unclear, which limits our understanding of the formation of flower color and stress adaptation in rose.
WRKY transcription factors regulate anthocyanin accumulation in plants through both transcriptional and post-transcriptional pathways. They can directly activate anthocyanin-related structural genes and/or upstream R2R3-MYB regulators, or they can interact with components of the MBW complex to form higher-order regulatory complexes. For instance, in lily, LhWRKY44 not only directly activates the promoters of LhF3H and LhGST but also physically interacts with the LhMYBSPLATTER protein[24]. In sweet potato (Ipomoea batatas), IbWRKY44 directly binds to the IbDFR promoter and interacts with both IbMYB1 and IbbHLH1/2[37]. PyWRKY26 directly transactivates PyMYB114 and its related protein also interacts with the PyMYB114 protein[10]. MdWRKY10 transcriptionally activates MdF3'5'H, MdUFGT, and MdMYB10, while also interacting with MdTTG1[21]. Conversely, MdWRKY11 is involved in regulating MdUFGT, MdMYB10, and MdMYB11 but does not interact with the core MBW complex[38].
In this study, we report, for the first time, that the Group IIc WRKY transcription factor RcWRKY71 not only modulates anthocyanin accumulation and drought tolerance in Rosa chinensis, but also enhances its transcriptional activation of downstream anthocyanin-related MYB regulators by forming homodimers via its leucine (Leu) zipper domain.
-
The rose cultivars used in this study were cultivated at the Germplasm Resource Center of Shanghai Normal University. Petals from rose varieties with different flower colors, including the white-flowered 'Mount Shasta', the yellow-flowered 'Yellow Leisure Liness', the pale purple-flowered 'Libellula', and the red-flowered 'Angela' and 'Yan Li', were sampled on the day of anthesis. Petals were collected from 10 individual plants for each cultivar and divided into three biological replicates. All samples were immediately frozen in liquid nitrogen and stored at −80 °C. Nicotiana benthamiana and N. tabacum were grown in a controlled growth chamber under the following conditions: 23 °C with a 16-hour light/8-hour dark photoperiod.
Cloning of RcWRKY71 and promoters of RcMYBs and anthocyanin biosynthesis genes
-
The full-length sequences of RcMYBs, RcbHLH42, and RcTTG1 were cloned in a previous study, as described therein[4]. The full-length coding sequences of RcWRKY71 were cloned from cDNA of Rosa chinensis 'Old Blush'. The promoters of RcMYB1 and anthocyanin pathway structural genes were cloned in a previous study, as described therein[4]. The promoters of RcMYBs were cloned from the genomic DNA of Rosa chinensis 'Old Blush'. All primers used are listed in Supplementary Table S1. To investigate the functional role of the Leu zipper domain in the RcWRKY71 protein, site-directed mutagenesis was performed on its coding sequence. The mutated gene fragment was chemically synthesized by Sangon Biotech (Shanghai, China). The nucleotide sequence is as follows: CAATCACAGGGTTTGATGTTCCTTCATACATGAACACCAGCTTCACAGAGTACAACTCGCATGCAAGCGGCTTTCGGCTCATCGTCTTCACCCTCCGATCACGAAGTATTTTCCTCCATTGACGAAGGCAGCCATCAAAAGCCTGTTGATAATCACGGATACTCA. Subsequently, this synthesized fragment was used to replace the corresponding wild-type sequence in the original vector to construct the mutant expression vector.
Sequence alignment and phylogenetic analysis
-
The amino acid sequences of RcWRKY71 and its homologs were obtained from the NCBI database, whereas the amino acid sequences of the Arabidopsis WRKY family members were retrieved from UniProt. Multiple sequence alignment was performed using DNAMAN 9.0. Phylogenetic analysis was conducted with MEGA11 (
https://megasoftware.net ) using the maximum likelihood method, and the resulting tree was visualized and annotated using iTOL (https://itol.embl.de ).Reverse transcription quantitative polymerase chain reaction
-
Total RNA was extracted using a Polysaccharide Polyphenol Plant Total RNA Extraction Kit (TIANGEN; DP441). First-strand cDNA was synthesized from the extracted RNA using the Hifair® AdvanceFast 1st Strand cDNA Synthesis Kit (YEASEN; 11149ES60). Quantitative polymerase chain reacton (qPCR) was performed using Hieff® qPCR SYBR Green Master Mix (No Rox) (YEASEN; 11201ES08) on a Thermo Fisher Scientific QuantStudio 5 Real-Time PCR System. RcActin was used as the internal reference gene, and the relative expression levels were calculated using the 2−ΔΔCᴛ method. The qPCR primers of RcMYB1, RcbHLH42, RcTTG1, and structural genes of the anthocyanin pathway were cloned in a previous study, as described therein[4]. The qPCR primers of stress-responsive genes were cloned in a previous study, as described therein[39]. All primers used for reverse transcription (RT)-qPCR are listed in Supplementary Table S2.
Subcellular localization
-
The coding sequence (CDS) of RcWRKY71 (without the stop codon TGA) was cloned into the pCAMBIA2300 vector to generate the RcWRKY71–GFP fusion construct, which was then introduced into Agrobacterium tumefaciens strain GV3101. The Agrobacterium was resuspended in an infiltration buffer (10 mM MgCl2, 10 mM MES [2-(N-morpholino)ethanesulfonic acid], and 200 μM acetosyringone [AS]) and adjusted to an optical density (OD600) of 1.0. After incubation in the dark for 2 hours, the suspension was infiltrated into leaves of 6-week-old N. benthamiana plants. The infiltrated plants were kept in the dark for 24 hours and then transferred to a normal light–dark cycle. After 3 days, leaf discs were stained with 10 μg/mL 4′,6-diamidino-2-phenylindole (DAPI) for 15 min. Green fluorescent protein (GFP) and DAPI fluorescence signals were observed and captured using Olympus FV3000 confocal scanning microscope with excitation wavelengths of 488 and 340 nm, respectively. All primers used are listed in Supplementary Table S3.
Transient overexpression in Rosa hybrida 'Mount Shasta' and N. tabacum
-
The full-length CDS of RcWRKY71 (including the stop codon) was cloned into the pCAMBIA2300 vector. Agrobacterium strains harboring either the empty pCAMBIA2300 vector or the pCAMBIA2300–RcWRKY71 construct were prepared and adjusted to an OD600 of 1.0 as described above. Petal discs (6 mm in diameter) were prepared from nearly open flowers of Rosa hybrida 'Mount Shasta' using a cork borer. The discs were infiltrated with the Agrobacterium suspension under a vacuum of −50 kPa for 3 min. After infiltration, the discs were rinsed, kept in the dark for 24 hours, and then placed on filter paper moistened with a 0.5% sucrose solution in Petri dishes to maintain humidity. The petal discs were incubated at 18 °C under light for 7 days before anthocyanin extraction, dry air treatment, or RNA extraction. The experiment was repeated at least three times independently. For each biological replicate, discs were collected from at least five different flowers, with the total number of discs per group exceeding 100. For transient expression in tobacco (N. tabacum) leaves, Agrobacterium strains carrying pCAMBIA2300–RcMYB1 were mixed in a 1:1 ratio with strains carrying either the empty pCAMBIA2300 vector or pCAMBIA2300–RcWRKY71. The mixed suspensions were infiltrated into tobacco leaves, which were then observed for phenotyping and used for anthocyanin extraction after 7 days of cultivation. All primers used are listed in Supplementary Table S3.
Yeast one-hybrid assay
-
The CDSs of RcWRKY71 and mRcWRKY71 were cloned into the pB42AD vector. The promoters of anthocyanin-related structural genes and activator-type MYB genes were cloned into the pLacZ-2μ vector; a subset of these plasmids was constructed as described previously[4]. The yeast strain EGY48 (Saccharomyces cerevisiae) was co-transformed with 1 μg each of pB42AD, pB42AD-RcWRKY71, and their respective pLacZ-2μ recombinant vectors, using the Polyethylene Glycol/Lithium Acetate (PEG/LiAc) method. Transformed yeast cells were selected and grown on synthetic dropout (SD) medium lacking tryptophan and uracil (SD/-Trp/-Ura), with or without X-Gal, as indicated. All primers used are listed in Supplementary Table S3.
Yeast two-hybrid assay
-
The CDSs of RcWRKY71 and mRcWRKY71 were cloned into the pGBKT7 vector. The CDSs of activator-type MYBs were cloned into the pGADT7 vector, a subset of these plasmids was constructed as described previously[4]. The pGBKT7–RcWRKY71 and pGADT7 recombinant vectors were co-transformed in 1-μg quantities into the yeast strain Y2HGold (S. cerevisiae) using the PEG/LiAc method. Transformed yeast cells were first selected on SD medium lacking leucine and tryptophan (SD/-Leu/-Trp). Positive clones were then assayed for protein interaction on quadruple dropout medium (SD/-Trp/-Leu/-His/-Ade) supplemented with 3-AT, Aureobasidin A (AbA), and X-α-Gal. All primers used are listed in Supplementary Table S3.
Bimolecular fluorescence complementation assay
-
The CDS of RcWRKY71 was cloned into a vector containing the N-terminal fragment of yellow fluorescent protein (YFP) (YN), generating RcWRKY71–YN. The CDS of activator-type MYBs and mRcWRKY71 were cloned into a vector containing the C-terminal fragment of YFP (YC), generating RcMYBs–YC. Agrobacterium strains carrying RcWRKY71–YN (or the empty YN control) were mixed in a 1:1 ratio with strains carrying RcMYBs–YC (or the empty YC control). The mixtures were infiltrated into N. benthamiana leaves as described previously. YFP and mCherry fluorescence signals were detected and imaged using an Olympus FV3000 confocal scanning microscope with excitation wavelengths of 505 and 587 nm, respectively. All primers used are listed in Supplementary Table S3.
Split-luciferase complementation assay
-
The CDS of RcWRKY71 was cloned into an nLUC vector, generating RcWRKY71–nLUC. The CDSs of activator-type MYBs were cloned into a cLUC vector, generating cLUC–RcMYBs. Agrobacterium strains carrying RcWRKY71–nLUC (or the empty nLUC control) were mixed in a 1:1 ratio with strains carrying cLUC–RcMYBs (or the empty cLUC control). The mixtures were infiltrated into N. benthamiana leaves. After 5 min of spraying the abaxial leaf surface with 150 μg/mL D-luciferin and subsequent dark adaptation, luminescence images were captured using a cooled low-light Charge-Coupled Device (CCD) imaging system (Tanon-4200, China). All primers used are listed in Supplementary Table S3.
Dual-luciferase reporter assay
-
The CDSs of RcWRKY71 and mRcWRKY71 were cloned into the pGreenII 62-SK effector vector. The promoters of RcMYB1, RcMYB113c, and RcMYB114a were cloned into the pGreenII 0800-LUC reporter vector. Agrobacterium strains carrying the empty pGreenII 62-SK or pGreenII 62-SK-RcWRKY71 effector were mixed in a 1:1 ratio with strains carrying the respective pGreenII 0800-LUC-proMYB reporter construct. The mixtures were infiltrated into N. benthamiana leaves. Luminescence was detected as described for the split-luciferase complementation assay (LCA), using a cooled low-light CCD imager (Tanon-4200, China) after application of the substrate. All primers used are listed in Supplementary Table S3.
Anthocyanin extraction and quantification
-
Anthocyanin content was determined following an established protocol with modifications[40]. Fresh petal discs (0.2 g) were homogenized in 2 mL of acidified methanol (1% HCl, v/v) and incubated at 4 °C for 24 hours in the dark. After centrifugation at 12,000 rpm for 1 min (4 °C), the supernatant was collected, and its absorbance was measured at 530 and 657 nm using a UV-2600 spectrophotometer (Techcomp, China). Anthocyanin levels were expressed as relative units per milligram fresh weight, calculated as: [(A530 − A657) × Dilution factor/Sample weight (mg)] × 1,000.
Chromatin immunoprecipitation quantitative polymerase chain reaction
-
The CDS of RcWRKY71 (without the stop codon TGA) was cloned into the pCAMBIA2300 vector to generate the RcWRKY71–Flag fusion construct. Rose petal discs were infiltrated with Agrobacterium strains harboring either the RcWRKY71–Flag or the Flag-EV (empty vector) construct using the vacuum infiltration method described above. Samples were collected at 3 days post-infiltration (dpi). Chromatin immunoprecipitation (ChIP) assays were performed as described by Bowler et al.[41], with adjustments. Briefly, 5 g of leaf tissue from RcWRKY71–Flag and control (Flag-EV) transgenic plants was crosslinked with 1% formaldehyde. Chromatin was extracted, fragmented by sonication, and immunoprecipitated using an anti-Flag antibody (Abcam, ab290). After reversing the crosslinks, the enriched DNA was purified and analyzed by quantitative polymerase chain reaction (qPCR) with gene-specific primers (Supplementary Table S4). Each experiment included three biological replicates.
-
To identify the WRKY genes responsible for anthocyanin accumulation in rose, we first screened differentially expressed WRKY genes from our previous transcriptome data taken from different floral developmental stages. A gene expression heatmap revealed that WRKY genes within the cluster marked with a red asterisk were highly expressed during the S2 and S3 stages, suggesting a potential association with anthocyanin accumulation (Fig. 1a). Furthermore, an analysis of the expression of these candidate genes in rose cultivars with varying flower colors showed that Chr5g0002561 and Chr5g0013131 were highly expressed in the two red-flowered varieties (Fig. 1b). Since drought treatment was previously found to induce anthocyanin accumulation in rose petal discs, we further analyzed the expression of these WRKY genes under such conditions. The results demonstrated that Chr5g0013131, Chr5g0002561, and Chr2g0151681 were significantly induced by drought stress. Chr5g0013131 exhibited the highest relative expression level under drought stress, suggesting its dominant role in drought-induced anthocyanin accumulation (Fig. 1c). On the basis of these collective findings, we selected Chr5g0013131 as the candidate WRKY gene for further investigation into its role in regulating drought-induced anthocyanin accumulation.
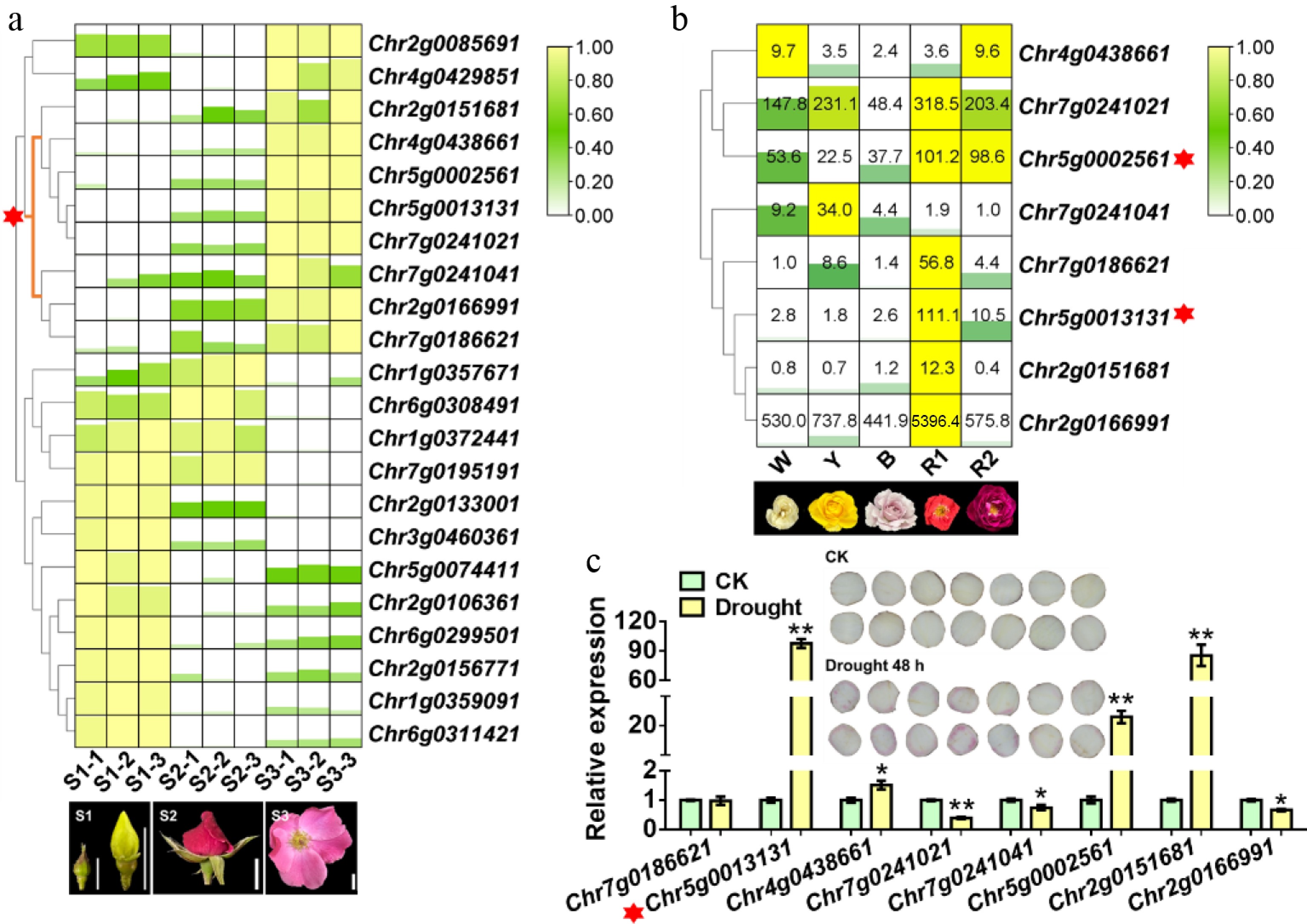
Figure 1.
Identification of a WRKY gene involved in drought-induced anthocyanin accumulation in Rosa chinensis. (a) Heatmap showing differentially expressed WRKY genes from the transcriptome of R. chinensis 'Old Blush' at different flower developmental stages. Representative petal images for each stage are shown. Scale bar: 1 cm. The red asterisk marks the cluster of WRKY genes highly expressed at the S2 and S3 stages, which are critical for anthocyanin accumulation in rose petals. (b) Expression patterns of candidate WRKY genes in rose cultivars with different flower colors. W, white-flowered cultivar (R. hybrida 'Mount Shasta'); Y, yellow-flowered cultivar (R. hybrida 'Yellow Leisure Liness'); B, blue-purple-flowered cultivar (R. hybrida 'Libellula'); R1 and R2, red-flowered cultivars (R. hybrida 'Angela' and R. hybrida 'Yan Li', respectively). Representative petal images are shown. Scale bar, 1 cm. The red asterisk indicates the WRKY genes with relatively high expression in red petals. (c) Relative expression levels of selected WRKY genes in response to drought stress. The phenotype of anthocyanin accumulation in R. hybrida 'Mount Shasta' petal discs with (Drought) or without (CK) 48 hours of drought treatment is shown. The red asterisk marks the stronger response of Chr5g0013131 to drought stress. Data are presented as the mean ± standard deviation (n = 3). Asterisks indicate significant differences (* p < 0.05; ** p < 0.01; Student's t-test).
RcWRKY71 is a transcription factor with transcriptional activation activity
-
A phylogenetic tree constructed with Chr5g0013131 and Arabidopsis WRKY family members revealed that Chr5g0013131 shares high sequence homology with AtWRKY71 and clusters within the Group IIc subfamily; hence it was designated RcWRKY71 (Fig. 2a). Multiple sequence alignment showed that RcWRKY71 contains the canonical WRKY domain and a C2H2 (CX4-5CX22-23HXH) zinc finger motif (Fig. 2b). To assess the transcriptional activity of RcWRKY71, we generated an effector plasmid, pBD-RcWRKY71, using pBD-VP16 as a positive control. The reporter construct contained the firefly luciferase (LUC) gene, driven by a minimal cauliflower mosaic virus 35S promoter upstream of GAL4-binding elements (Fig. 2c). Co-infiltration of pBD-RcWRKY71 with the reporter construct into N. benthamiana leaves resulted in significantly higher LUC activity compared with the pBD control (Fig. 2c), indicating that RcWRKY71 functions as a transcriptional activator. To determine the subcellular localization of RcWRKY71, we constructed a RcWRKY71–GFP fusion vector and transiently expressed it in tobacco leaves. The GFP fluorescence signal overlapped with DAPI staining, demonstrated that RcWRKY71 is localized in the nucleus (Fig. 2d). Collectively, these results confirmed that RcWRKY71 is a nuclear-localized transcription factor with transcriptional activation activity.
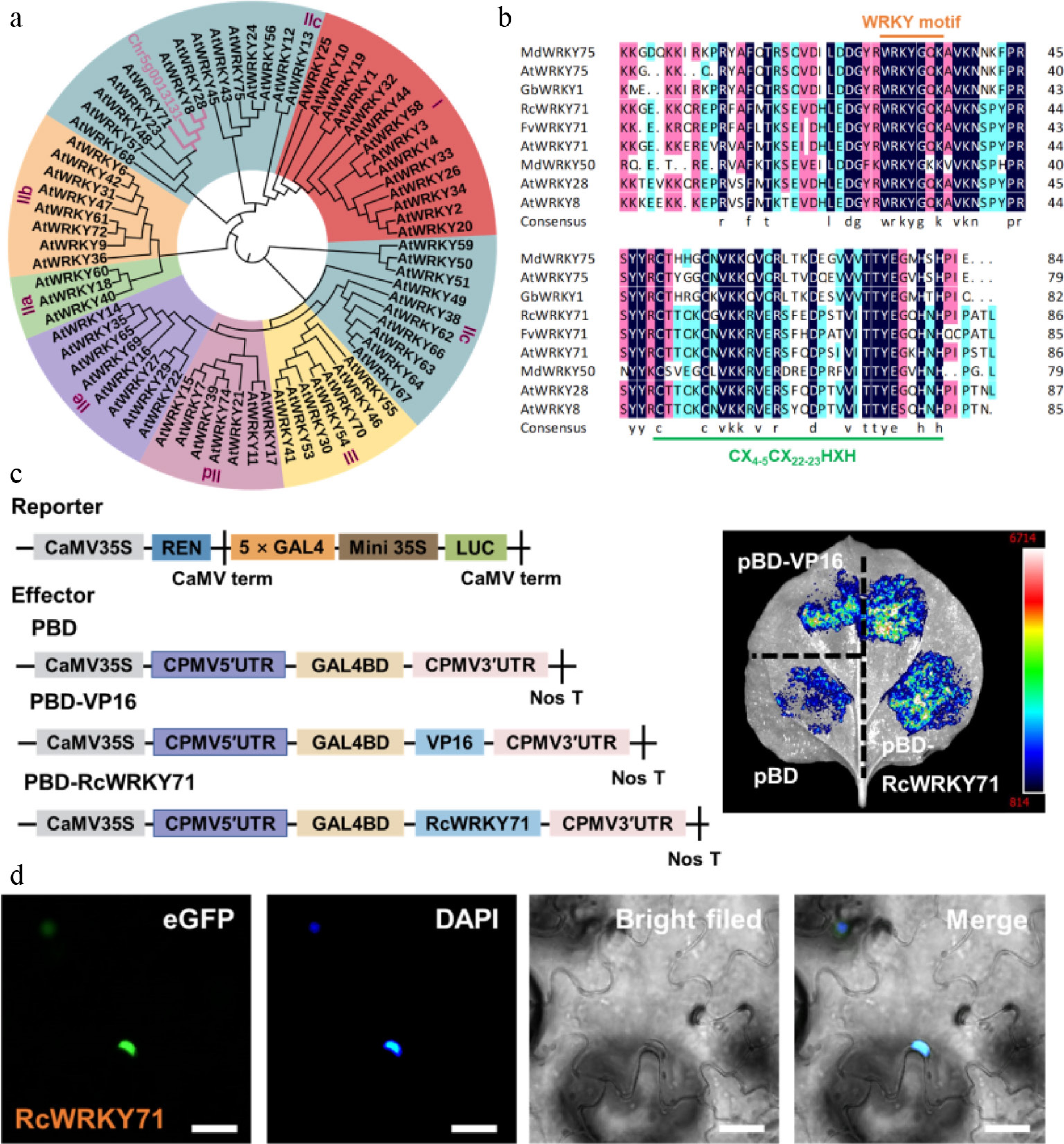
Figure 2.
Phylogenetic analysis, multiple sequence alignment, transcriptional activity assay, and subcellular localization of RcWRKY71. (a) Phylogenetic tree constructed from the full-length amino acid sequences of RcWRKY71 and WRKY family members from Arabidopsis thaliana. (b) Multiple sequence alignment of RcWRKY71 and its homologs from different species. The conserved WRKY domain and zinc finger motif were indicated. (c) Transcriptional activity of RcWRKY71 assessed using the GAL4-based luciferase (LUC) reporter system. The effector and reporter constructs are schematically illustrated. (d) Subcellular localization of RcWRKY71. The RcWRKY71–GFP fusion protein (green) localizes to the nucleus, as shown by co-staining with DAPI (blue). Scale bars, 20 μm.
RcWRKY71 promotes anthocyanin accumulation in R. chinensis
-
To investigate whether RcWRKY71 is involved in regulating anthocyanin biosynthesis in rose, we first performed transient overexpression assays in tobacco leaves. The results demonstrated that co-expression of RcWRKY71 with RcMYB1 and RcbHLH42 significantly enhanced anthocyanin accumulation two days after infiltration, compared with the expression levels of the empty vector (EV) with RcMYB1 and RcbHLH42 (Fig. 3a). We also conducted transient overexpression assays in petal discs of the white-flowered rose cultivar 'Mount Shasta'. Overexpression of RcWRKY71 alone induced visible anthocyanin pigmentation and significantly increased anthocyanin content in the petal discs (Fig. 3b, c). Reverse transcription–qPCR (RT-qPCR) analysis revealed that transient overexpression of RcWRKY71 markedly upregulated the expression levels of all tested structural genes in the anthocyanin pathway. Among the MYB genes known to regulate anthocyanin accumulation in rose, the expression levels of RcMYB1, RcMYB114a, and RcMYB114b were significantly induced (Fig. 3d). These findings collectively indicate that RcWRKY71 acts as a positive regulator of anthocyanin accumulation in rose.
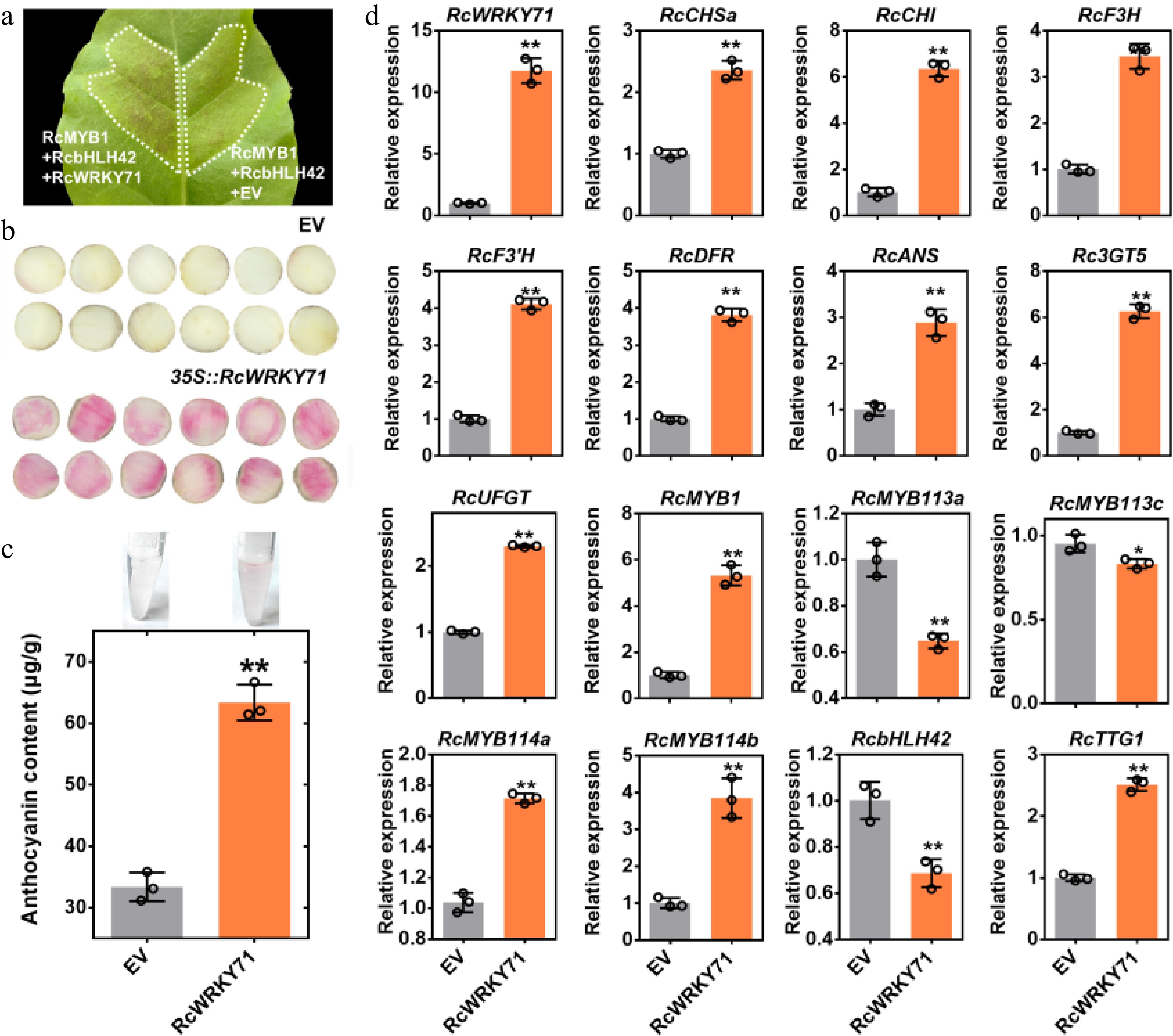
Figure 3.
Phenotypic and molecular analysis of the transient overexpression of RcWRKY71 in tobacco leaves and rose petal discs. (a) Phenotype of tobacco leaves following transient overexpression of RcWRKY71. (b) Phenotype of rose petal discs of the white-flowered rose cultivar 'Mount Shasta' after transient overexpression of RcWRKY71. (c) Anthocyanin content measurement in 'Mount Shasta' petal discs transiently overexpressing RcWRKY71. Data are presented as the mean ± standard deviation (n = 3). Asterisks indicate a significant difference (** p < 0.01; Student's t-test). (d) RT-qPCR analysis of the relative expression levels of RcWRKY71, anthocyanin structural genes, and regulatory genes in 'Mount Shasta' petal discs after transient overexpression of RcWRKY71. Data are presented as the mean ± standard deviation (n = 3). Asterisks indicate a significant difference (* p < 0.05, ** p < 0.01; Student's t-test).
Overexpression of RcWRKY71 enhances drought tolerance in rose via anthocyanin accumulation
-
Previous research has demonstrated that RcWRKY71 positively regulates drought resistance in rose by modulating stomatal movement and ROS scavenging[39]. Given that anthocyanin accumulation effectively enhances elimination of ROS and confers resilience to various abiotic stresses[14], we hypothesized that RcWRKY71-mediated anthocyanin accumulation contributes to improved drought tolerance in rose petals. Petal discs of the 'Mount Shasta' cultivar were subjected to drought treatment 3 days after infiltration. The results showed that petal discs overexpressing RcWRKY71, which accumulated anthocyanins, exhibited less wrinkling than the control and recovered better after rewatering (Fig. 4a). Furthermore, relative electrolyte leakage measurement, along with 3,3'-diaminobenzidine (DAB) and nitroblue tetrazolium (NBT) staining, revealed that leaves overexpressing RcWRKY71 accumulated less hydrogen peroxide and superoxide anions, indicating reduced oxidative damage (Fig. 4b, c). Quantitative analysis showed that the expression levels of RcABF2 from the abscisic acid pathway, as well as the stress-responsive genes RcDREB1, RcPR4, RcSPCH, and RcLEA14, were significantly upregulated (Fig. 4d). These results indicate that RcWRKY71 enhances drought stress tolerance, at least partially, by promoting anthocyanin accumulation.
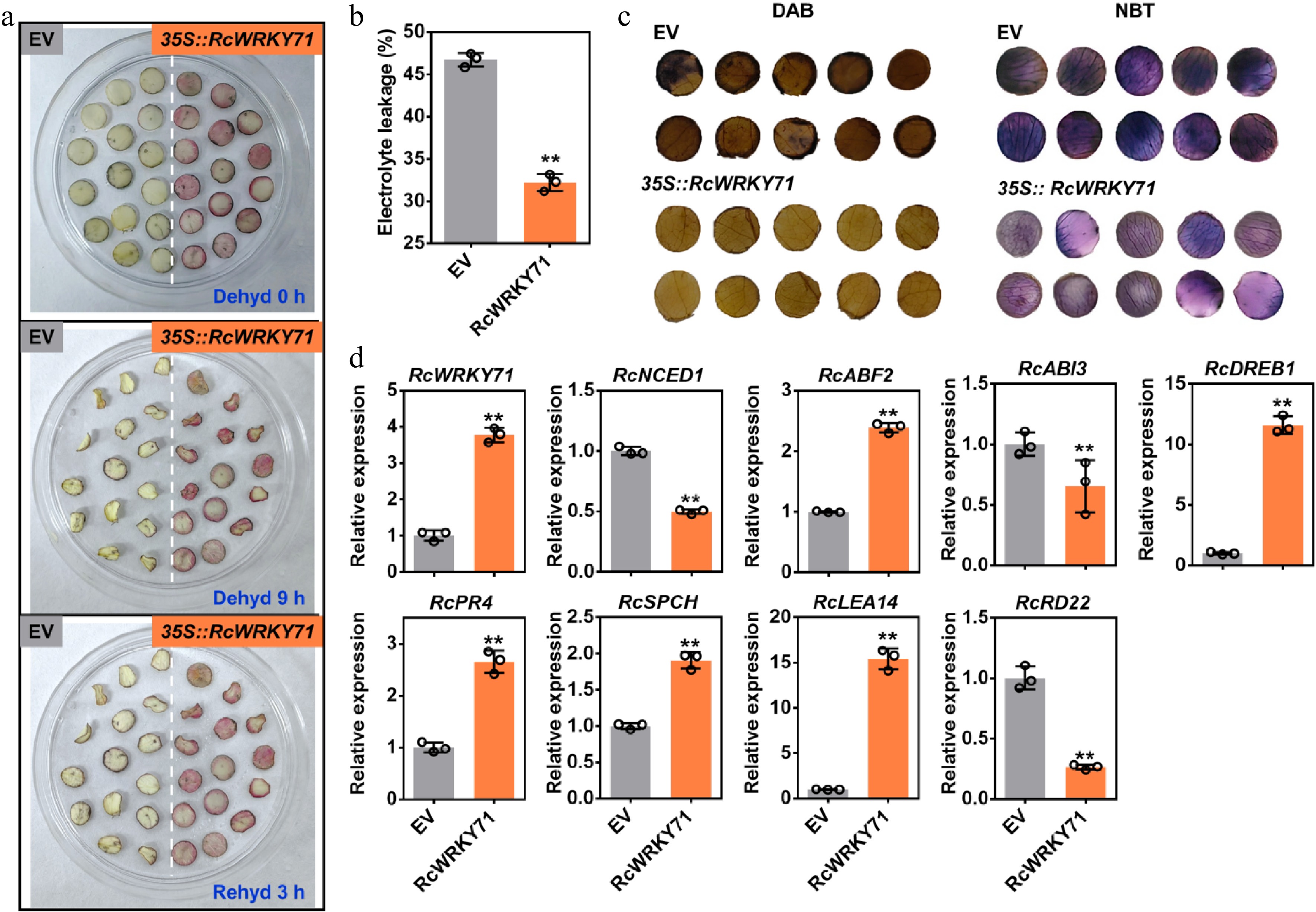
Figure 4.
Physiological and molecular analysis of rose petal discs overexpressing RcWRKY71 under drought stress. (a) Phenotype of petal discs after 5 days of transient overexpression of RcWRKY71 followed by drought treatment. (b) Relative electrolyte leakage measured in petal discs after 5 days of transient overexpression of RcWRKY71 and subsequent drought stress. Data are presented as the mean ± standard deviation (n = 3). Asterisks indicate significant differences (** p < 0.01; Student's t-test). (c) Histochemical detection of hydrogen peroxide (H2O2) and superoxide anion (O2−) using DAB and NBT staining, respectively, in petal discs under the same conditions as in (a). (d) RT-qPCR analysis of the relative expression levels of stress-responsive genes in petal discs after 5 days of transient overexpression of RcWRKY71. Data are presented as the mean ± standard deviation (n = 3). Asterisks indicate significant differences (** p < 0.01; Student's t-test).
Silencing of RcWRKY71 reduces anthocyanin accumulation and impairs drought tolerance in petal discs
-
To further elucidate the function of RcWRKY71, virus-induced gene silencing (VIGS) was performed in rose petal discs, followed by drought treatment 7 days after infiltration. Compared with the control, RcWRKY71-silenced petal discs exhibited significantly reduced anthocyanin accumulation and showed more severe wilting after 6 hours of drought stress (Fig. 5a). Measurements of anthocyanin content, relative electrolyte leakage, and histochemical staining with DAB and NBT further confirmed that silencing of RcWRKY71 led to decreased anthocyanin levels and increased drought-induced damage (Fig. 5b–e). RT-qPCR analysis revealed that silencing of RcWRKY71 significantly downregulated the expression of anthocyanin biosynthetic genes; the activators RcMYB1, RcMYB113c, and RcMYB114a; and stress-responsive genes including RcABF2, RcDREB1, RcPR4, RcSPCH, and RcLEA14 (Fig. 5f). These results demonstrate that RcWRKY71 acts as a positive regulator of both anthocyanin accumulation and drought stress tolerance.
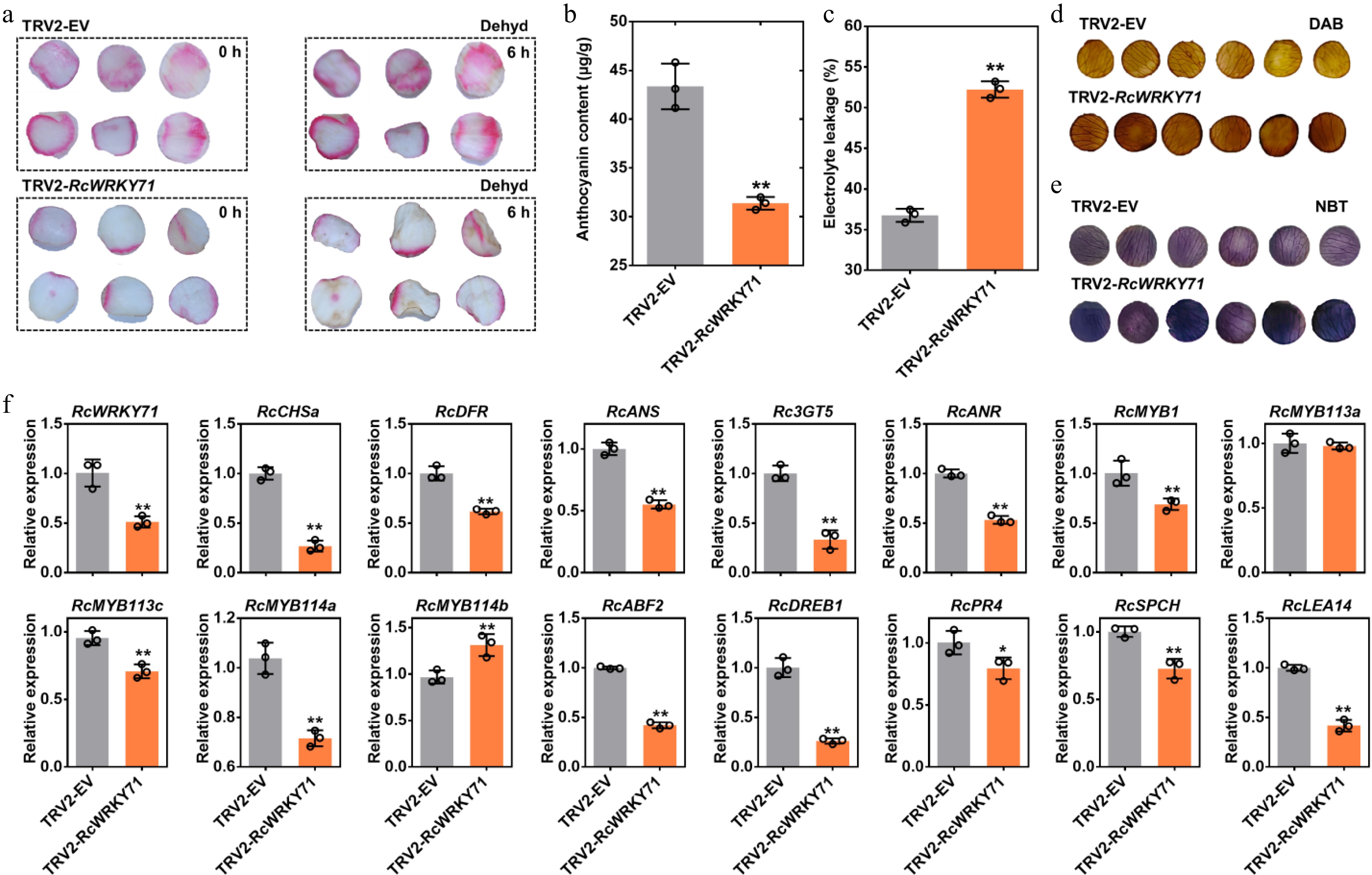
Figure 5.
Phenotypic, physiological, and molecular analysis of RcWRKY71-silenced rose petal discs. (a) Anthocyanin accumulation phenotype and drought stress phenotype in petal discs after RcWRKY71 silencing. (b) Anthocyanin content measurement in RcWRKY71-silenced petal discs. Data are presented as the mean ± standard deviation (n = 3). Asterisks indicate significant differences (** p < 0.01; Student's t-test). (c) Relative electrolyte leakage in RcWRKY71-silenced petal discs under drought stress. Data are presented as the mean ± standard deviation (n = 3). Asterisks indicate significant differences (** p < 0.01; Student's t-test). (d) Detection of hydrogen peroxide using DAB staining in RcWRKY71-silenced petal discs under drought stress. (e) Detection of superoxide anion using NBT staining in RcWRKY71-silenced petal discs under drought stress. (f) RT-qPCR analysis of the relative expression levels of anthocyanin biosynthesis/regulatory genes and stress-responsive genes in RcWRKY71-silenced petal discs under drought stress. Data are presented as the mean ± SD (n = 3). Asterisks indicate significant differences (** p < 0.01; Student's t-test).
RcWRKY71 promotes anthocyanin biosynthesis by directly binding to the promoters of RcMYB1, RcMYB113c, and RcMYB114a
-
WRKY transcription factors regulate anthocyanin biosynthesis at both the transcriptional and post-transcriptional levels. Transcriptional regulation involves direct binding to the promoters of anthocyanin-related structural genes or activator-type MYB genes, whereas post-transcriptional regulation occurs through interactions with one or more components of the MBW protein complex[10,21,24,37,38]. To further investigate the molecular mechanism by which RcWRKY71 regulates anthocyanin accumulation in rose, we first performed a yeast one-hybrid assay to examine whether RcWRKY71 functions at the transcriptional level. The results showed that RcWRKY71 does not directly bind to the promoters of anthocyanin-associated structural genes (Fig. 6a) but it binds specifically to the promoters of the activator-type MYB genes RcMYB1, RcMYB113c, and RcMYB114a (Fig. 6b). Bioinformatic analysis revealed that all three promoters (proRcMYB1, proRcMYB113c, and proRcMYB114a) contain potential WRKY71-binding sites (Fig. 6c). Subsequent dual-luciferase (LUC) reporter assays demonstrated that RcWRKY71 significantly activates the transcription of RcMYB1, RcMYB113c, and RcMYB114a (Fig. 6d, e). Furthermore, ChIP-qPCR confirmed the significant enrichment of RcWRKY71 at specific regions of these promoters: P1 of proRcMYB1, P2 of proRcMYB113c, and P4/P5 of proRcMYB114a (Fig. 6f). Collectively, these results demonstrate that RcWRKY71 promotes anthocyanin biosynthesis by directly binding to and transactivating the promoters of RcMYB1, RcMYB113c, and RcMYB114a.
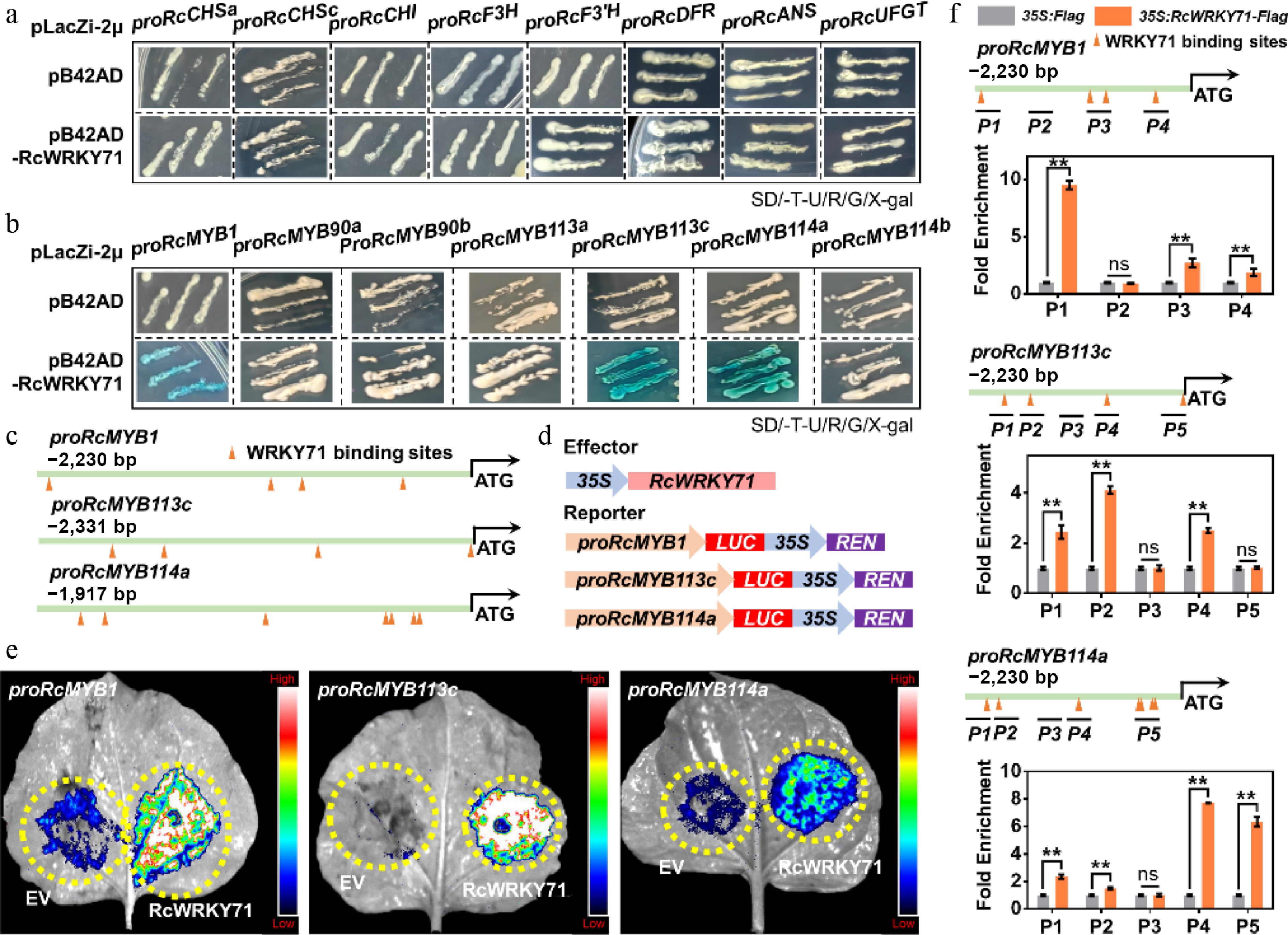
Figure 6.
Transcriptional regulatory mechanism of RcWRKY71 in anthocyanin accumulation in R. chinensis. (a) Yeast one-hybrid assay testing the binding of RcWRKY71 to the promoters of anthocyanin structural genes. (b) Yeast one-hybrid assay examining the binding of RcWRKY71 to the promoters of SG6 subgroup MYB genes that regulate anthocyanin accumulation. (c) Prediction of WRKY71-binding sites in the promoters of RcMYB1, RcMYB113c, and RcMYB114a using JASPAR. (d) Schematics of the dual-luciferase (LUC) reporter constructs. (e) Transactivation activity of RcWRKY71 on the promoters of RcMYB1, RcMYB113c, and RcMYB114a measured by dual-luciferase assays. (f) ChIP-qPCR analysis of RcWRKY71 binding to the promoters of RcMYB1, RcMYB113c, and RcMYB114a. Data are presented as the mean ± SD (n = 3). Asterisks indicate significant differences (** p < 0.01; Student's t-test).
RcWRKY71 does not interact with the MBW complex but forms homodimers
-
To investigate whether RcWRKY71 regulates anthocyanin biosynthesis in rose at the post-translational level by interacting with the MBW protein complex, we first performed a bimolecular fluorescence complementation (BiFC) assay. The results showed that RcWRKY71 does not interact with RcMYB1, RcMYB113c, RcMYB114a, RcbHLH42, or RcTTG1. However, it interacts with itself to form homodimers (Fig. 7a). The absence of an interaction with the MBW complex's components and the self-interaction of RcWRKY71 were further confirmed by yeast two-hybrid and split-luciferase complementation assays, respectively (Fig. 7b, c).
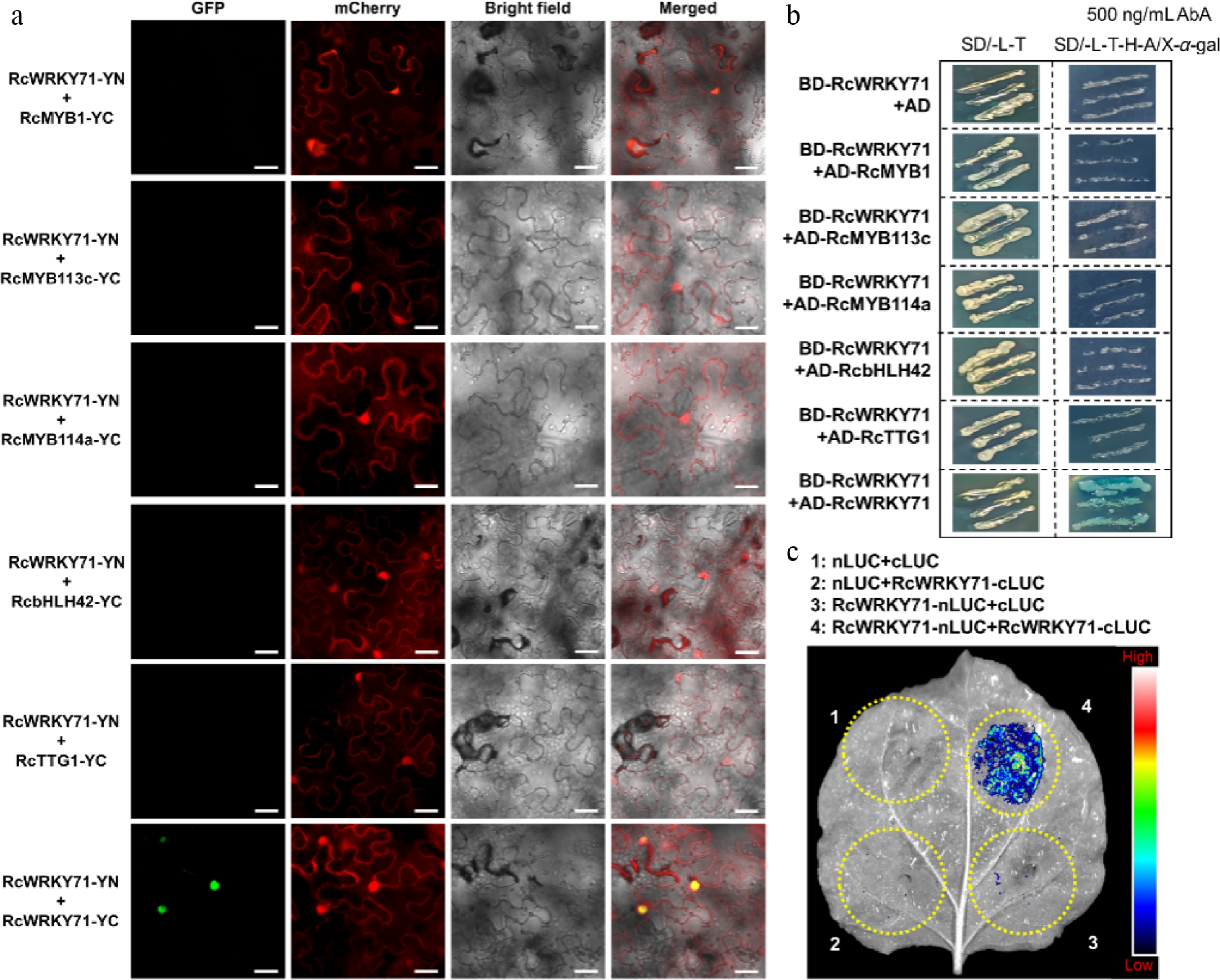
Figure 7.
Post-translational regulatory mechanism of RcWRKY71 in anthocyanin accumulation in R. chinensis. (a) BiFC assay examining the protein interaction between RcWRKY71 and components of the anthocyanin-related MBW complex. Scale bars, 20 μm. (b) Yeast two-hybrid assay testing the protein interaction between RcWRKY71 and MBW complex members. (c) Split-luciferase complementation assay confirming the homomeric interaction of RcWRKY71.
Leucine zipper-mediated homodimerization enhances RcWRKY71 function
-
To determine whether the function of RcWRKY71 relies on homodimer formation, we first mapped its interaction domain. Structural analysis revealed that RcWRKY71 contains a Leu zipper motif at its N-terminus, a key domain for protein homo- or heterodimerization. Mutagenesis of conserved Leuresidues within this motif, followed by yeast two-hybrid assays, showed that RcWRKY71 interacts with fragments containing the Leu zipper but not with fragments lacking it. Moreover, mutation of the Leu residues abolished this interaction. BiFC assays further confirmed that the Leu zipper is essential for RcWRKY71's homodimerization (Fig. 8a, b).
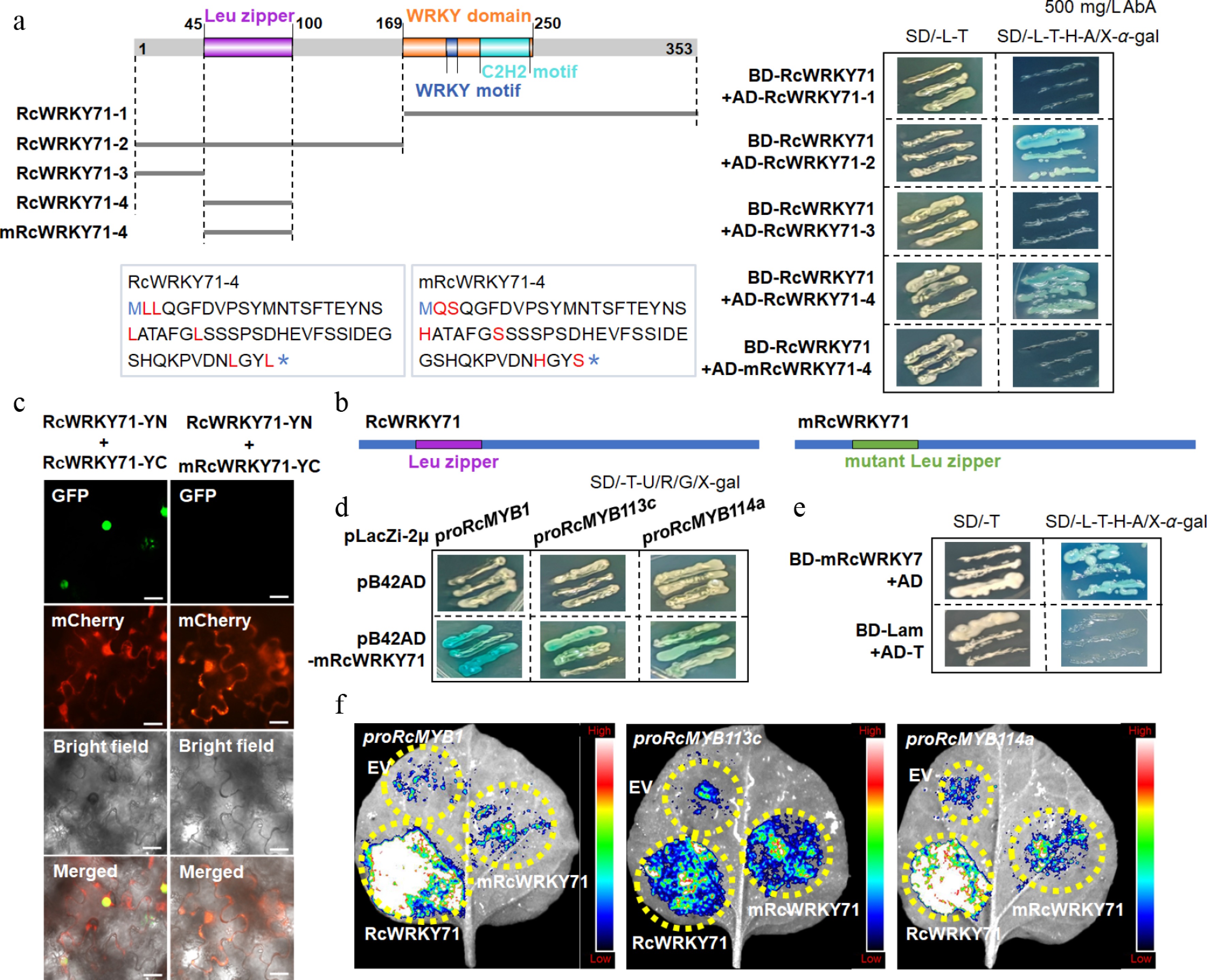
Figure 8.
Effect of Leu-zipper mutation on homodimer formation and function of RcWRKY71. (a) Yeast two-hybrid assay showing the interaction between different truncated versions of RcWRKY71 and the effect of Leu-zipper mutation on protein interaction. (b) Schematic diagram of RcWRKY71 and the mutant mRcWRKY71. (c) BiFC assay confirming that the Leu-zipper mutant mRcWRKY71 fails to interact with wild-type RcWRKY71. Scale bars, 20 μm. (d) Yeast one-hybrid (Y1H) assay demonstrating that the Leu-zipper mutant mRcWRKY71 retains the ability to bind to the promoters of downstream genes. (e) Yeast two-hybrid assay showing that the Leu-zipper mutant mRcWRKY71 maintains its transcriptional activation activity. (f) Dual-luciferase (Luc) assay showing that the transactivation ability of mRcWRKY71 toward downstream gene promoters is significantly attenuated.
We further found that the mutant RcWRKY71 (mRcWRKY71) with a disrupted Leu zipper retained its ability to bind to the promoters of RcMYB1, RcMYB113c, and RcMYB114a (Fig. 8c, d). Yeast cells harboring BD-mRcWRKY71 grew normally on SD/-L-T-H-A/X-α-gal medium without AbA, whereas the negative control failed to grow, indicating that the Leu zipper mutation does not affect the intrinsic transcriptional activity of RcWRKY71 (Fig. 8e). However, dual-luciferase assays demonstrated that although mRcWRKY71 could still activate the transcription of downstream MYB genes, its transactivation activity was significantly lower than that of the wild-type RcWRKY71 (Fig. 8f).
These results indicate that although the Leu zipper mutation does not impair the DNA-binding ability or intrinsic transcriptional activity of RcWRKY71, the disruption of homodimer formation significantly attenuates its capacity to activate downstream gene expression.
-
WRKY transcription factors constitute a large family of transcriptional regulators in plants, classified into three major groups (I, II, III), with Group II further subdivided into five subgroups (IIa–IIe), depending on the number of WRKY domains and the structure of their zinc finger motifs[15]. An increasing body of evidence has demonstrated that WRKY members from various groups participate in the regulation of anthocyanin biosynthesis. For instance, within Group I, homologs of AtTTG2 function as positive regulators in multiple species[21,23−25]. In Group III, StWRKY70 promotes anthocyanin accumulation in potato tubers, whereas BnWRKY41-1 and AtWRKY41 act as negative regulators in the leaves[35,36]. Notably, among the Group II subgroups, members from IIa, IIb, IIc, and IId have been implicated in anthocyanin regulation, with IIc members like MdWRKY50 and FaWRKY71 being prominent examples[28−32]. In this study, we identified RcWRKY71 from Rosa chinensis as a member of the Group IIc subgroup (Fig. 2a). Its expression pattern strongly correlated with anthocyanin accumulation during petal development and across cultivars with different flower colors, positioning it as a key candidate regulator in rose (Fig. 1).
The interplay between anthocyanin accumulation and abiotic stress tolerance is widely documented, with anthocyanins serving as potent ROS scavengers. Correspondingly, several WRKY transcription factors are known to mediate stress-induced anthocyanin accumulation. Examples include MdWRKY17 and MdWRKY50 in apple under drought, McWRKY71 in Momordica charantia under ozone stress, and FaWRKY71 in strawberry under multiple stresses[27−29]. Our results confirm that RcWRKY71 enhances drought tolerance, at least partly, through promoting anthocyanin accumulation (Fig. 6), as anthocyanins can scavenge ROS and reduce oxidative damage (Figs. 3, 4). However, the previously reported regulation of stomatal movement and ROS-scavenging genes by RcWRKY71 may also contribute to drought resistance, indicating a multipathway regulatory network mediated by RcWRKY71[39].
WRKY proteins use diverse molecular strategies to regulate anthocyanin synthesis. These include forming protein–protein interactions with components of the MBW complex (e.g., StWRKY70 with StAN1[35], MdWRKY40 with MdMYB1[26], and PhPH3 with PhAN11[22]) or directly binding to the promoters of anthocyanin-related structural genes or their upstream MYB regulators (e.g., MdWRKY50 targeting MdCHS and MdCHI[28], or CitWRKY75 activating CitRuby1[32]). Our comprehensive interaction and transactivation assays revealed that RcWRKY71 functions primarily through direct transcriptional activation of specific MYB genes (Fig. 6b, f). It does not directly bind to the promoters of core anthocyanin-related structural genes (Fig. 6a), nor does it physically interact with the MBW complex's components (RcMYBs, RcbHLH42, RcTTG1) (Fig. 7a, b). This delineates a clear and specific regulatory path for RcWRKY71, distinguishing its mechanism from that of other reported WRKY proteins.
Extensive protein–protein interactions exist among WRKY transcription factors. To date, experimental evidence has demonstrated interactions between members belonging to at least five out of the seven WRKY subgroups[40,41]. For instance, in Arabidopsis, subgroup IIa members WRKY18, WRKY40, and WRKY60 form heterodimers via their N-terminal Leu zipper motifs[42]. In rice (Oryza sativa), the Subgroup IIa protein OsWRKY71 can form not only homodimers but also heterodimers with the Subgroup IId protein OsWRKY51[43]. Furthermore, heterodimerization has been reported between the Subgroup IIb members WRKY6 and WRKY42 in Arabidopsis[44], as well as between the Subgroup III protein WRKY30 and either WRKY53, WRKY54, or WRKY70[45]. Notably, interactions across subgroups have also been observed, such as the heterodimer formed between Group IIa WRKY18 and Group I WRKY25[41].
In this study, we demonstrate for the first time that the Group IIc WRKY transcription factor RcWRKY71 forms homodimers through its N-terminal Leu zipper domain; disruption of this domain abolishes the interaction (Fig. 8). Previous studies have suggested that although a canonical Leu zipper is predominantly found at the N-terminus of Group IIa members, the N-terminal regions of WRKY proteins from other Group II subgroups and Group III also contain multiple Leu, isoleucine, or valine residues that may form similar secondary structures mediating dimerization[40,42].
The formation of homo- or heterodimers is crucial for modulating WRKY proteins' function, either enhancing or repressing their activity[46,47]. In our study, disrupting the Leu zipper in RcWRKY71 did not affect its ability to bind to the promoters of the downstream MYB genes, but significantly attenuated its transcriptional activation of RcMYB1, RcMYB113c, and RcMYB114a (Fig. 8). This indicates that homodimerization of RcWRKY71 primarily amplifies its transcriptional regulatory potency rather than determining DNA-binding specificity. This mechanism differs from other reported cases: In Arabidopsis, the WRKY18–WRKY60 heterodimer enhances the DNA-binding ability of WRKY18, whereas the WRKY60–WRKY40 dimer reduces the binding capacity of WRKY40[42]; WRKY25 may act as a redox switch, activating the expression of WRKY53 by forming a heterodimer with WRKY18, whereas WRKY18 homodimers strongly suppress WRKY53, thereby finely tuning the progression of senescence[41]. Similar functional enhancement via dimerization has been observed in other species; for example, the rice OsWRKY51–OsWRKY71 heterodimer substantially strengthens the binding of OsWRKY71 to the Amy32b promoter[43], and the apple MdWRKY50-MdWRKY17 heterodimer boosts the transcriptional activation of MdCHS and MdCHI[28].
In summary, our study not only expands the current knowledge by revealing that Group IIc WRKY proteins can form homodimers, but also elucidates a distinct mechanism through which Leu zipper-mediated dimerization enhances transcriptional activity. This finding provides a fresh perspective for understanding how WRKY transcription factors finely regulate downstream target genes via oligomerization to coordinate secondary metabolism and stress responses in plants.
-
In summary, our study reveals that the WRKY transcription factor RcWRKY71 promotes anthocyanin biosynthesis in Rosa chinensis by binding to and transactivating three activator-type MYB genes: RcMYB1, RcMYB113c, and RcMYB114a. Furthermore, we provide the first evidence that a Group IIc WRKY protein can form homodimers, which enhances its regulatory function.
-
The authors confirm their contributions to the paper as follows: study conception and design: He G, Ming F; data collection: Zheng R, Zou D, Yuan X; analysis and interpretation of results: Zheng R, He G, Ming F, Lou Y; draft manuscript preparation: Zheng R, He G. All authors reviewed the results and approved the final version of the manuscript.
-
All relevant data in this study are provided in the article and its supplementary file. The raw RNA-seq data have been submitted to the NCBI Sequence Read Archive (SRA) under accession number PRJNA1400243.
-
This work was supported by Shanghai Young Science and Technology Talent Sailing Program, China (23YF1431100); the Agriculture Research System of Shanghai, China (Grant No. 202208); and Shanghai Plant Germplasm Resources Engineering Research Center, 17DZ2252700.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primers used in full length sequence cloning of RcWRKY71 and Promoters of RcMYBs.
- Supplementary Table S2 Primers used in RT-qPCR.
- Supplementary Table S3 Primers used in vectors construction.
- Supplementary Table S4 Primers used in ChIP-qPCR.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zheng R, Zou D, Yuan X, Lou Y, Ming F, et al. 2026. RcWRKY71, a Group IIc transcription factor, enhances anthocyanin accumulation and drought tolerance in Rosa chinensis by activating MYB regulators and forming homodimers. Ornamental Plant Research 6: e016 doi: 10.48130/opr-0026-0007 

RcWRKY71, a Group IIc transcription factor, enhances anthocyanin accumulation and drought tolerance in Rosa chinensis by activating MYB regulators and forming homodimers
- Received: 30 November 2025
- Revised: 15 January 2026
- Accepted: 21 January 2026
- Published online: 22 April 2026
Abstract: Anthocyanins are vital secondary metabolites in plants, playing key roles in defending against diverse biotic and abiotic stresses. WRKY transcription factors are known to participate in stress-induced anthocyanin accumulation; however, the mechanism by which WRKY transcription factors regulate anthocyanin biosynthesis in rose (Rosa chinensis) remains unreported. In this study, we identified RcWRKY71, a Group IIc WRKY transcription factor containing the conserved WRKYGQK heptapeptide and a C2H2-type zinc finger motif. Subcellular localization analysis revealed that RcWRKY71 is localized in the nucleus and possesses transcriptional activation activity. Functional characterization demonstrated that RcWRKY71 promotes anthocyanin accumulation in rose petals and enhances drought tolerance. Further mechanistic studies showed that RcWRKY71 activates anthocyanin biosynthesis by directly binding to the promoters of three R2R3-MYB transcription factors: RcMYB1, RcMYB113c, and RcMYB114a. Additionally, RcWRKY71 forms homodimers via its leucine (Leu) zipper domain, which is essential for its functional enhancement. Our findings reveal a novel regulatory mechanism by which WRKY transcription factors modulate anthocyanin accumulation and confer drought resistance in rose, highlighting the role of Leu zipper-mediated homodimerization in enhancing transcriptional activity.
-
Key words:
- RcWRKY71 /
- Rosa chinensis /
- Anthocyanin biosynthesis /
- Drought tolerance /
- MYB regulators