








-
Agaricus subrufescens, also known as the almond mushroom, due to its distinctive almond-like flavor, was cultivated and consumed in the Atlantic United States from the late 1800s to the early 1900s[1]. The species was rediscovered in Brazil in 1960 and referred to as the 'Piedade mushroom', after the village in São Paulo where Takatoshi Furumoto collected it before sending specimens to Japan in 1965 for medicinal research. In 1967, Heinemann identified the Brazilian material as Agaricus blazei Murrill. Since that time, the mushroom has been referred to by several names, such as Himematsutake in Japan, Sun Mushroom® (Cogumelo do Sol) in Brazil, and Royal Sun Agaricus® in other regions[2]. A. subrufescens is an edible and medicinal mushroom cultivated all over the world for its culinary use and bioactive properties. Some of these compounds, such as polysaccharides, antioxidants, and immunomodulatory substances, have been shown to exhibit anti-tumor, anti-inflammatory, and antibacterial properties[1,2]. The rising demand for it has prompted research into improved cultivation methods, optimal strain selection, and plant crossbreeding to enhance quality, accelerate growth, and increase yields.
The production of A. subrufescens generally uses the procedures established for A. bisporus, which use composted lignocellulosic substrates. The compost is the fermented mixture of wheat straw, horse manure, and other organic residues[3]. The cultivation process includes inoculating spawn, the mycelium running in the compost, adding a casing layer, developing primordia, and producing the fruiting body. Environmental variables, including temperature, humidity, light, and CO2 concentrations, significantly influence mycelial growth, primordia development, and yield. However, strain variations are the principal factors affecting fruiting behavior and productivity[1,2]. Hybrid strains are increasingly studied due to their ability to combine advantageous characteristics from various parental lines, such as accelerated colonization, earlier primordia development, and increased yield[4].
Fungal extracellular enzymes are the primary catalysts for the degradation of substrates during culture. These enzymes are essential for nutrition assimilation and morphological development[5,6]. Hydrolytic enzymes, such as cellulases, hemicellulases, and β-glucosidases, facilitate the hydrolysis of complex polysaccharides, resulting in the release of sugars and essential nutrients required for mycelial growth and fruiting body formation[7−9]. Laccases and other oxidative enzymes decompose lignin and enhance the penetration of the mycelium. Chitinases and exochitinases break down the cell walls of fungi during their growth and primordia formation[4,10,11]. Previous research indicates that enzyme activity in basidiomycetes varies according to the strain and developmental stage, frequently achieving peak levels of laccase and cellulase activity during substrate colonization and primordia development. The study demonstrates a significant correlation between substrate utilization and stages of development[7,12]. Variations in extracellular enzyme activity have been documented during the cultivation period of A. subrufescens, highlighting the importance of enzyme dynamics in influencing its developmental and reproductive processes[13,14]. Oxidative degradation produces reactive oxygen species, including hydrogen peroxide (H2O2), which function as signaling molecules influencing morphogenesis and the development of fruiting bodies[15]. Understanding the relationship between strain-specific enzyme profiles, substrate degradation, and fruiting efficiency is important. Enhancing the growth of A. subrufescens, increasing its fruiting body production, and optimizing its biotechnological potential are necessary. Our knowledge about the variations in extracellular enzyme synthesis over the entire culture cycle remains limited, despite increased awareness of its health benefits. This indicates the need for additional research into the correlation between enzyme activity and specific developmental phases. This work aims to investigate the temporal dynamics of extracellular enzyme production over the cultivation cycle of A. subrufescens and determine the influence of these enzymatic activity patterns on substrate degradation and fruiting efficiency.
-
The four Agaricus subrufescens strains (CA454, CA487, CA918, and HB) used in this study were obtained from the Collection of Germplasms of Agarics in Bordeaux (CGAB), INRA, France. CA454 is a subculture of the collection strain A. blazei WC837; CA487 is a wild European strain; CA918 is a wild Thai strain collected by SC Karunarathna, P Callac, and S Rapior; and HB is a hybrid generated by crossbreeding the Brazilian strain CA454 and the French wild isolate CA487 (Table 1). For spawn preparation, mycelium cultivated on malt agar medium was transferred to sterilized rye grain, and incubated at 23 ± 2 °C until complete colonization occurred. The spawn was stored at 11 °C and acclimated to room temperature the day before spawning.
Table 1. Strains of A. subrufescens used in this study.
Strain Origin Remarks CA454 (= WC837) Brazil Subculture strain, also similar to ATCC 76739 CA487 France Wild isolate CA918 Thailand Wild isolate HB INRA Crossbred strain of CA454-3 × CA487-100 Mushroom cultivation
-
Strains were cultivated using compost and methods standard for the commercial production of A. bisporus[3]. The compost, supplied by Renault SA (Pons, France), consisted primarily of wheat straw and horse manure. Trays (17 cm height, 0.09 m2 surface area) were filled with 8 kg of compost and inoculated with 1% (w/w) spawn. Trays were incubated for 15 d within a climatic chamber maintained at 23 ± 0.5 °C and 85% ± 2% relative humidity. A casing layer (2.5 cm thick; limestone, ⅓ peat, ⅓ fine sand) was applied after incubation, and the trays were maintained under the same environmental conditions for 7 d (post-incubation). Fruiting was induced at 22–25 °C, a relative humidity of 95%–97%, and a low CO2 concentration (< 1,500 mg/m3). The number and fresh weight of mushrooms were recorded for up to 45 d post-casing (60 d after inoculation). Experiments were carried out using a completely randomized design with three replicates.
Compost sampling and crude enzyme extract preparation
-
Compost samples (150 g) were collected weekly from day 1 to day 42 post-inoculation. Samples for laccase and manganese peroxidase (MnP) tests were immediately milled using a household grinder. Crude enzymes were extracted by mixing 3 g of milled compost with 20 mL of distilled water and agitating the mixture for 20 min. Extracts were filtered and centrifuged at 12,000 × g for 10 min at 4 °C. For other enzyme assays, including fluorescein diacetate (FDA) hydrolysis, samples were lyophilized and ground. Crude extracts were prepared by mixing 0.7 g of powdered compost with 10 mL of distilled water and agitating the mixture for 30 min. Extracts were prepared as described above and used immediately for enzyme assays.
Enzyme assays
-
All enzyme assays were standardized using model substrates at 30 °C and the optimal pH for each enzyme. One unit (U) of enzyme activity is defined as the amount of enzyme releasing 1 μmol of product per min, per g of compost. All assays were performed in triplicate. Laccase (EC 1.10.3.2): the oxidation of syringaldazine (Sigma-Aldrich) was measured at 526 nm in 0.1 M phosphate buffer, pH 6.0[16]. Manganese peroxidase (MnP; EC 1.11.1.13): oxidative coupling of DMAB and MBTH (Fluka Chemica) in 0.1 M phosphate buffer, pH 6.0, with 0.05 M MnSO4 and 0.1 mM H2O2; absorbance measured at 590 nm[17]. Cellulase (EC 3.2.1.4) and xylanase (EC 3.2.1.8): the quantification of reducing sugars was performed using the DNS method, with CMC and xylan in a 0.05 M acetate buffer, pH 5.0, serving as substrates[18]. β-Glucosidase (EC 3.2.1.2), β-xylosidase (EC 3.2.1.37), and β-N-acetyl-glucosaminidase (EC 3.2.1.52) were evaluated using 4-nitrophenyl derivatives; the release of p-nitrophenol after 30 min of incubation was measured[19]. FDA hydrolysis: a lyophilized compost sample (0.1 g) was mixed with 5 mL of FDA working solution (0.01 mg/mL), incubated at 40 °C for 30 min with shaking, and the reaction was terminated with 50% acetone, followed by centrifugation at 11,000 rpm for 5 min at 15 °C, and absorbance was measured at 490 nm[20].
H2O2 assay
-
The content of hydrogen peroxide concentration in compost was determined according to the methodology outlined by Savoie et al.[15]. Briefly, 150 mg of lyophilized compost was mixed with 4 mL of 0.05 M phosphate buffer (pH 6.0) and incubated at 25 °C for 15 min. Extracts were centrifuged at 12,000 × g for 10 min at 4 °C and subsequently heated at 80 °C for 40 min to inactivate enzymes. After cooling, extracts were treated with 40 μL of 1% polyethyleneimine and centrifuged at 1,000 × g for 5 min. The reaction mixture contained 0.25 mL of extract, 0.25 mL of 20 U/mL horseradish peroxidase (Sigma), and 0.625 mL of chromophore solution (0.6 mM MBTH and 7.5 mM DMAB in 0.05 M phosphate buffer, pH 6.0). Oxidation was monitored at 590 nm after 3 min incubation at 30 °C.
Statistical analysis
-
All experiments were performed in triplicate, and results are presented as mean ± standard deviation (SD). Error bars in the figures represent the standard deviations of three replicates. Statistical significance was determined at p < 0.05.
-
All four A. subrufescens strains were successfully cultivated on commercial compost, resulting in the formation of fruiting bodies (Fig. 1). The mycelia of each strain fully colonized the compost trays within 15 d after applying a casing layer to induce fruiting. All four strains completely colonized the casing layer within 28 d under the studied conditions. However, significant differences were observed in the timing of primordia formation, fruiting, the number of fruiting bodies, and the total yield (Fig. 2). The French strain CA487 exhibited initial primordia formation at day 28, and developed the first fruiting bodies by day 35, potentially increasing its output, yielding approximately 830 fruiting bodies with a total fresh weight ranging from 12–12.5 kg (Fig. 2). The Brazilian strain CA454, the Thai strain CA918, and the hybrid strain (HB, a cross between CA454 × CA487) developed primordia by day 35, with the initial fruiting bodies forming on day 42. CA454 produced the lowest yield, fewer than 50 fruiting bodies, and had minimal fresh weight. CA918 produced approximately 150 fruiting bodies (~2 kg fresh weight), while the HB hybrid showed intermediate performance, producing over 200 fruiting bodies with a total fresh weight of 5.5 kg. These results indicate that genetic variables significantly influence fruiting characteristics and overall yield. Breeding programs can effectively combine desirable traits, as demonstrated by the improved productivity of the HB hybrid compared to its Brazilian parent. Variability in basidiomata morphology among strains was also observed, consistent with previous reports[2]. This aspect is likely influenced by both genotype and environmental factors. Since all strains were cultivated under conditions comparable to those used for A. bisporus, the observed differences in yield appear primarily genotype-driven. A. subrufescens strains exhibit a wide range of traits worldwide, including in the Americas, Asia, Europe, and Oceania[1,2]. The full scope of genetic diversity remains well understood; optimized cultivation practices can improve productivity[3]. This present study supports the importance of both genotypic selection and appropriate cultivation management for maximizing productivity.

Figure 1.
Overall cultivation cycle of A. subrufescens, using the French cultivar CA 487 as a representative, showing different developmental stages: (a) spawn-running period (day 14), (b) mycelial colonization in the casing layer (day 21), (c) fruiting induction (day 28), and (d) fruiting body formation (day 35). Scale bars: (c) = 1 cm, (d) = 1.5 cm.
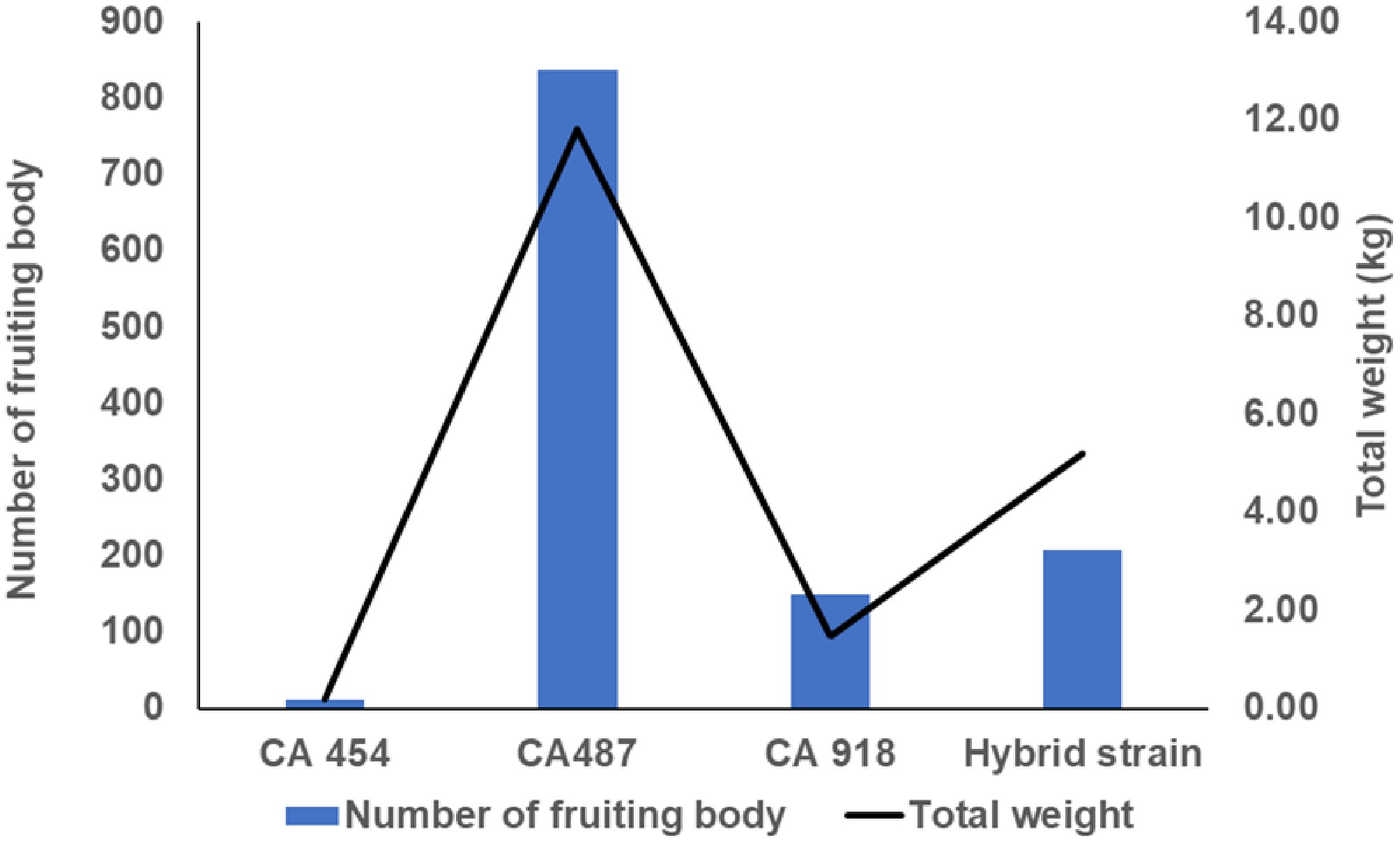
Figure 2.
Productivity of four strains of Agaricus subrufescens, showing the number of fruiting bodies and total weight (kg) for each strain.
Enzyme activities and fungal growth during cultivation
-
In general, enzymatic activity during A. subrufescens cultivation was strongly strain-dependent and closely linked to fruiting body development (Fig. 3). Laccase activity was undetectable at inoculation (day 0) but began to accumulate after 1 week, increasing steadily during spawn running and stabilizing or slightly declining after primordia formation. The Thai strain CA918 exhibited the highest peak activity (~2–2.3 U/g substrate; Fig. 3a). These trends align with findings in other basidiomycetes, where laccase production is associated with mycelial growth and the initiation of fruiting[4,21]. The activity of manganese peroxidase (MnP) was consistently low across all strains, showing a slight initial peak in CA454 before declining to near zero by day 21 (Fig. 3b). This pattern differs from the cyclical MnP production observed in A. bisporus, which is affected by the initial compost microbiota[22,23]. Cellulolytic and hemicellulolytic enzymes, including β-glucosidase, β-xylosidase, CMCase, and xylanas, exhibited distinct strain-specific temporal patterns. β-Glucosidase was detectable at inoculation, presumably indicating initial microbial activity (Fig. 3c). CA918 retained increased activity during later stages, whereas CA487 and the hybrid exhibited moderate, stable levels. β-Xylosidase revealed a comparable pattern, exhibiting an increase during fructification in CA454, CA918, and the hybrid, whereas CA487 showed a more cyclical pattern (Fig. 3d). CMCase activity reached its maximum at day 28 in CA454, CA487, and CA918, but decreased during primordia formation, whereas the hybrid obtained an earlier peak at day 14 and subsequently maintained relatively consistent activity (Fig. 3e). Xylanase activity followed the same pattern as CMCase. CA918 and CA454 maintained higher levels during later stages of incubation, whereas CA487 and the hybrid exhibited a decline (Fig. 3f). Fungal metabolic activity, measured through fluorescein diacetate (FDA) hydrolysis, showed a characteristic pattern. FDA activity was highest at day 0, then declined sharply by day 7 as the compost microbiota diminished and A. subrufescens became dominant. Activity subsequently stabilized or increased slightly by day 21, indicating resumed hyphal metabolism (Fig. 4). Exochitinase (N-acetyl-β-glucosaminidase), a marker of chitin remodeling during development, reached its highest level in CA918 during primordia formation (day 35) and declined during fruiting (day 42). Throughout the cultivation period, the concentration of other strains remained consistently low (Fig. 5). Comparable patterns were seen in the bacteriolytic enzyme activity, which was markedly increased in CA918 during primordia and fruiting formation, suggesting enhanced microbial interaction or substrate modification in this strain. The concentrations of hydrogen peroxide (H2O2), measured in mmole/g sample, increased for all strains during the cultivation period (Fig. 6). During the early phase (days 7–21), CA487 and the hybrid CA487 × CA454 had the most accelerated growth, with CA487 reaching approximately 1.05 mmole/g by day 21. In contrast, CA918 continuously exhibited low levels throughout the entire period (< 0.20 mmole/g). On day 28, the H2O2 concentration in the strains varied between 0.95 and 1.15 mmole/g. CA454 and the hybrid exhibiting higher levels than those of CA487. A notable decrease occurred at the primordia stage—particularly in CA454 (~0.43 mmole/g)—followed by a sharp increase during fruiting body development, when the hybrid and CA487 obtained approximately 1.3 mmole/g, and CA918 reached about 1.15 mmole/g. This biphasic pattern, characterized by an initial increase, a temporary decline at the primordia initiation, and a maximum during fruiting was consistent across all strains. In general, the enzymatic profiles suggest that the compost microbiota affects enzyme production in the early stage, whereas the physiological traits of specific strains predominantly regulate enzyme dynamics in the later stages of growth. The Thai strain CA918 consistently exhibited increased cellulolytic, hemicellulolytic, and lytic enzyme activities, especially during primordia development and fruiting, in comparison to the other strain assessed. In contrast, the hybrid and other strains exhibited decreased activity levels but indicated enhanced consistency. These differences likely contribute to the variations in how efficiently the strains utilize substrates, grow mycelium, and produce fruiting bodies.
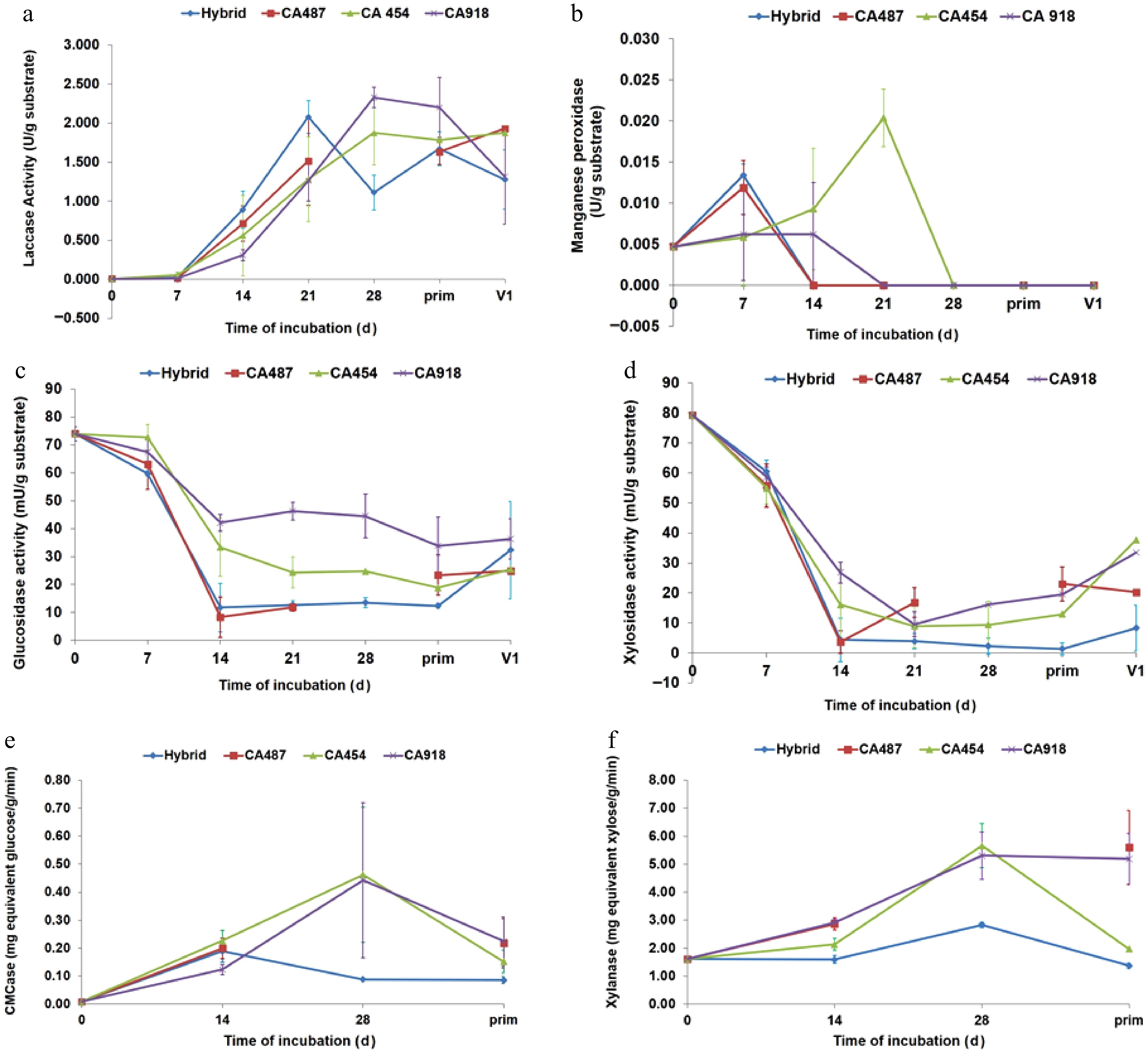
Figure 3.
Extracellular enzyme production by Agaricus subrufescens in compost: (a) laccase, (b) manganese peroxidase, (c) glucosidase, (d) xylosidase, (e) CMCase, and (f) xylanase. Symbols represent different strains: (▲) CA454, (■) CA487, (♦) hybrid CA487 × CA454, and (×) CA918. Error bars indicate the standard deviation from three parallel cultivations.
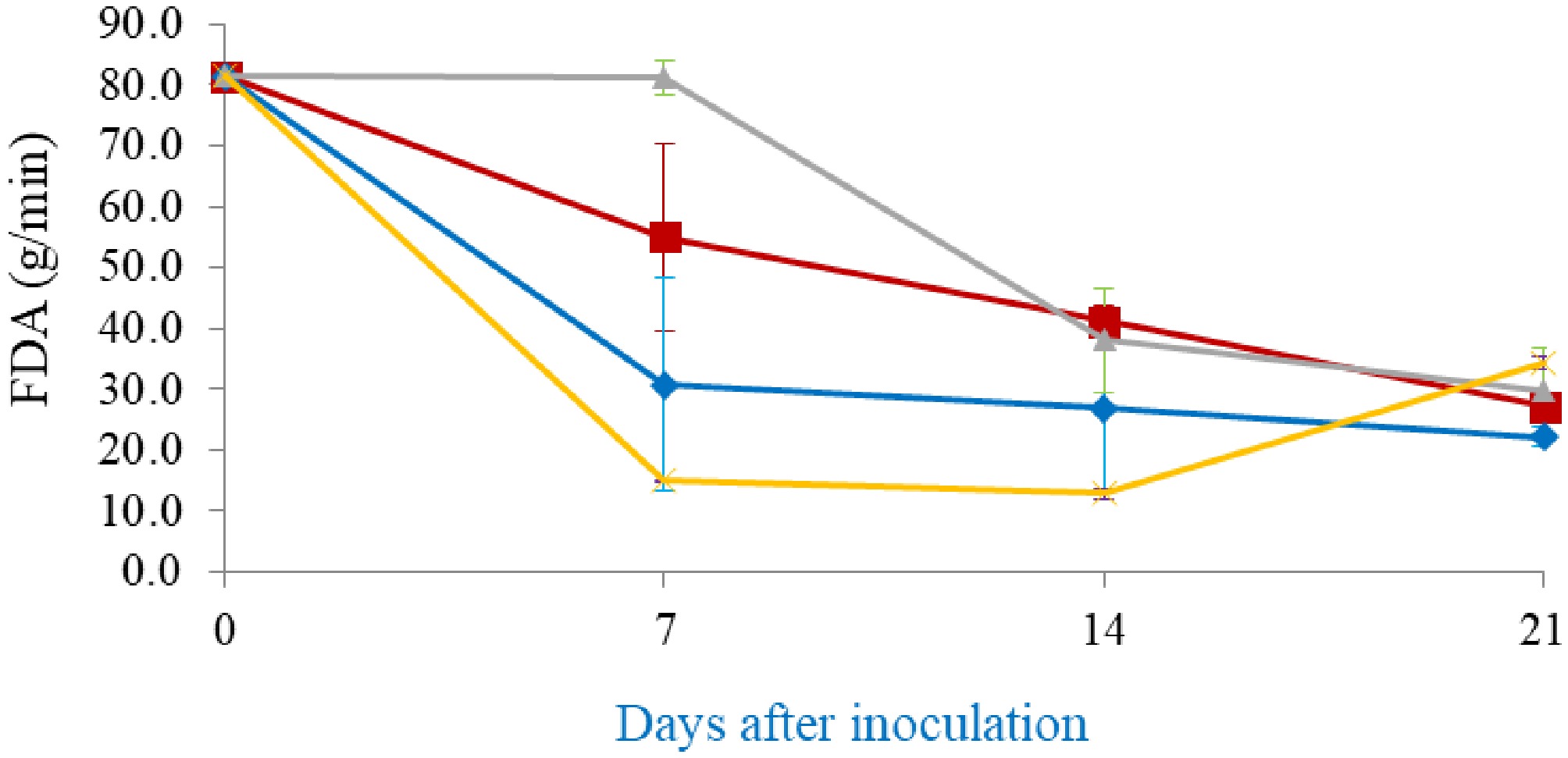
Figure 4.
Fluorescein diacetate (FDA) hydrolysis by Agaricus subrufescens strains: (▲) CA454, (■) CA487, (♦) hybrid CA487 × CA454, and (×) CA918. Error bars represent the standard deviation from three parallel cultivations.
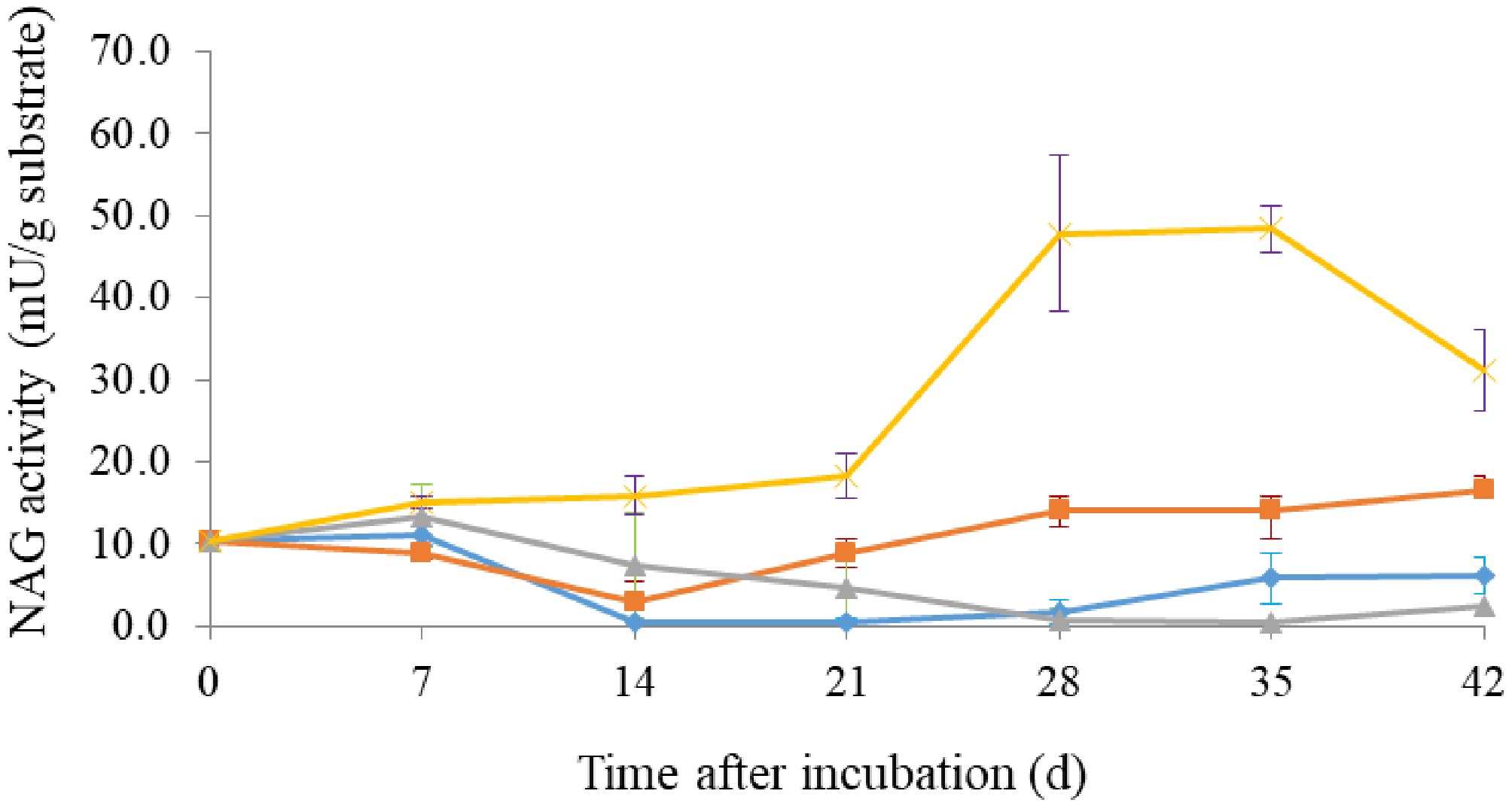
Figure 5.
N-acetyl-glucosaminidase (NAG) activity of Agaricus subrufescens strains: (▲) CA454, (■) CA487, (♦) hybrid CA487 × CA454, and (×) CA918. Error bars indicate the standard deviation from three parallel cultivations.
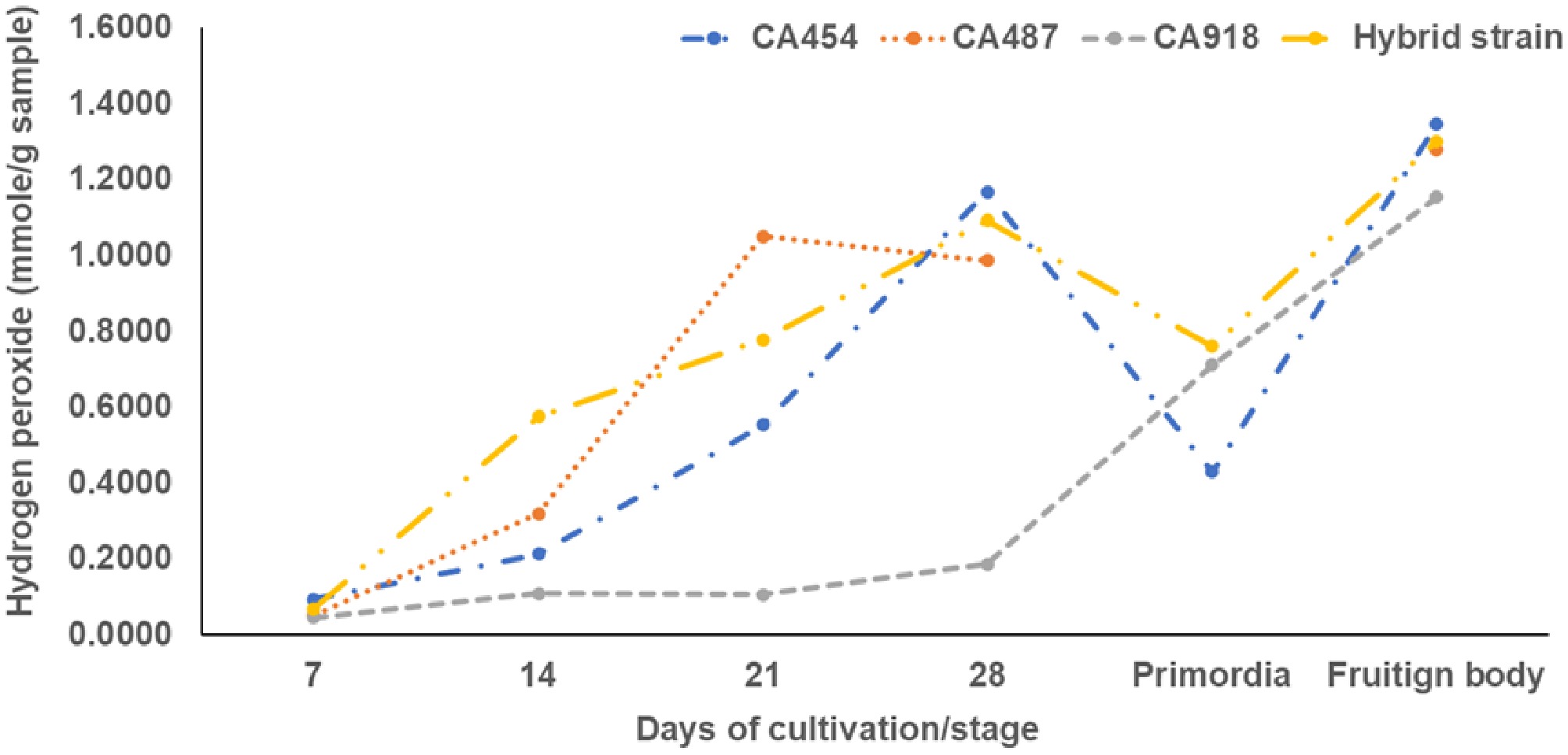
Figure 6.
Changes in H2O2 concentration in extracts from mushroom compost colonized by Agaricus subrufescens.
-
The current study provides a comprehensive analysis of the cultivation dynamics, substrate utilization, and enzymatic activity of four A. subrufescens strains, including both wild-type and hybrid isolates. Understanding these traits is essential for improving mushroom productivity and for progressing biotechnological applications, including the valorization of lignocellulose degradation from agricultural and industrial residues. Our results are consistent with previous research on A. bisporus and A. subrufescens, while introducing novel strain-level comparisons relevant to commercial cultivation. All four strains successfully colonized the commercial compost and produced fruiting bodies. Significant variation was observed in the timing of primordia formation, the beginning of fruiting, the number of fruiting bodies, and the total yield. The French CA487 strain exhibited the earliest formation of primordia and fruiting bodies, yielding the highest productivity, whereas the Brazilian CA454 strain yielded the lowest output. The hybrid strain exhibited intermediate performance, indicating that selective breeding can effectively combine advantageous traits, aligning with other studies that emphasize extensive genotypic and phenotypic diversity in A. subrufescens[1,2]. The strain-specific responses parallel the findings of Sousa et al.[14], who reported significant variation among A. subrufescens strains in terms of mycelial growth, productivity, and enzymatic activity, with performance in compost not consistently correlating with growth on conventional culture media. Our findings emphasize the importance of evaluating strains directly under production-relevant conditions.
The enzymatic profiles observed in this study revealed dynamic changes during substrate colonization and fruiting, incorporating trends previously recorded in both A. bisporus and A. subrufescens. Laccase activity was undetectable at inoculation but increased during spawn-running, peaking during primordia development—a pattern consistent with the findings of Sousa et al.[14], who indicated that A. subrufescens produces measurable laccase but produces minimal cellulase and no manganese peroxidase (MnP) or lignin peroxidase. Our study similarly found minimal MnP activity across all strains, further suggesting that MnP is not a major enzyme in A. subrufescens, unlike A. bisporus, where cyclic MnP production has been associated with early-stage compost microbial dynamics[22,23]. Hydrolytic enzyme activities—including β-glucosidase, β-xylosidase, CMCase, and xylanase—differed among strains, but often reached their peak during substrate colonization and the initial stage of primordia formation. This reflects the initial enzymatic peaks observed in A. bisporus[24], where N-acetyl-glucosaminidase and other polysaccharidases showed significant activity as the mushroom mycelium replaced indigenous microorganisms. Similar to Savoie's study, our FDA hydrolysis data exhibited an initial activity increase that likely reflects the transition from indigenous compost microbes to the dominance of A. subrufescens hyphae. Moreover, the consistently high β-glucosidase and exochitinase activity of the Thai strain (CA918) suggests an enhanced capability for degrading microbial residues and hemicelluloses—nutrient sources considered essential during the first colonization in A. bisporus compost[24]. The strong lignocellulolytic activity observed in some strains of A. subrufescens correspond with the findings of Farnet et al.[13], who demonstrated that various A. subrufescens strains can match or surpass A. bisporus in lignocellulose-modifying activity, hydrogen peroxide production, and substrate conversion under standard composting conditions. Importantly, they found that A. subrufescens growth did not significantly disrupt the functional diversity of compost microbial communities after 2 weeks—consistent with our observation that early microbial contributions affect initial enzymatic environments before strain-specific activities dominate during primordia development. Taken together, these findings highlight the interaction between compost microbial communities and the fundamental metabolic capacities of different A. subrufescens strains. During the stages of colonization, native microbes provide enzymatic functions and accessible nutrients, while the mushroom's hydrolytic enzymes increase as its mycelium asserts dominance. Laccase, CMCase, and xylanase activities subsequently reach their peak during primordia development and early fruiting, facilitating structural remodeling, nutrient mobilization, and morphogenesis—patterns consistent with findings from other basidiomycetes and white-rot fungi[7,8,10]. The coordinated enzymatic dynamics demonstrate how A. subrufescens efficiently adapts its metabolism according to substrate composition and developmental stage.
This study indicated that the interaction between genotypic features and culture conditions affects the productivity and enzymatic performance of A. subrufescens. The integration of our data with previous research on A. bisporus and A. subrufescens indicates that enzyme activity patterns may function as predictive indicators for growth, colonization efficiency, and fruiting capability. This knowledge can guide the selection and breeding of superior strains, the optimization of compost formulations, and the enhancement of cultivation procedures. The exhibited lignocellulolytic activities of A. subrufescens highlighted its potential for biotechnological applications, especially in the valorization of agricultural residues and towards the development of sustainable fungal bioprocesses.
-
There is a significant variety in growth rate, fruiting yield, and enzyme production among A. subrufescens strains. These variations are primarily due to their genetic composition. Developmental stages closely correlate with patterns of enzymatic activity. Laccase activity promotes the formation of primordia and fruiting bodies. Selective breeding and strain-specific cultivation methods designed for a certain strain can enhance productivity, while hydrolytic enzymes contribute to substrate breakdown. An in-depth analysis of enzyme dynamics can facilitate the determination of optimal growth conditions and the exploration of biotechnological applications for this commercially valuable mushroom. Selective breeding and strain-specific cultivation techniques may improve the production.
We are indebted to Nathalie Ferrer for her invaluable assistance during this experiment.
-
The authors confirm their contributions to the paper as follows: study conception and design: Kakumyan P, Savoie JM; data collection: Kakumyan P; analysis and interpretation of the results, review and editing: Kakumyan P, Chukeatirote E, Savoie JM; draft manuscript preparation: Kakumyan P, Chukeatirote E. All authors reviewed the results and approved the final version of the manuscript.
-
The authors confirm that the data supporting the findings of this study are available within the article.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Kakumyan P, Chukeatirote E, Savoie JM. 2026. Enzymatic dynamics during cultivation cycle of Agaricus subrufescens on commercial compost. Studies in Fungi 11: e014 doi: 10.48130/sif-0026-0012 

Enzymatic dynamics during cultivation cycle of Agaricus subrufescens on commercial compost
- Received: 24 December 2025
- Revised: 24 December 2025
- Accepted: 02 April 2026
- Published online: 20 May 2026
Abstract: Agaricus subrufescens is a valuable mushroom for commercial and health purposes due to its bioactive compounds and culinary use. Different genotypes exhibit distinct growth patterns, enzyme profiles, and fruiting strategies. This study evaluated four strains—French CA487, Brazilian CA454, Thai CA918, and a hybrid (CA454 × CA487)—cultivated on wheat straw–horse manure compost under controlled conditions to assess strain-specific development and enzymatic traits. Fruiting body formation, yield, and extracellular enzyme activities (manganese peroxidase, laccase, β-glucosidase, β-xylosidase, CMCase, xylanase, N-acetyl-β-glucosaminidase, and fluorescein diacetate hydrolysis) were monitored throughout the cultivation cycle alongside hydrogen peroxide (H2O2) accumulation. CA487 showed the earliest primordia and fruiting body development, producing ~830 fruiting bodies with a fresh weight of 12–12.5 kg, while CA454 had a limited yield. The hybrid displayed improved performance, suggesting targeted breeding can combine favorable traits. Enzyme activity was strongly influenced by strain and developmental stage: hydrolytic enzymes peaked during substrate colonization and primordia formation, whereas laccase activity was highest during fruiting body maturation. H2O2 levels initially increased during mycelial growth, slightly decreased during primordia formation, and peaked at full maturation. The Thai strain CA918 consistently showed high hydrolytic and lytic enzyme activity, reflecting versatile substrate utilization and developmental strategies. These results highlight the critical role of genotype and enzymatic regulation in optimizing A. subrufescens cultivation, improving yield, and maximizing its biotechnological potential.
-
Key words:
- Agaricus subrufescens /
- Enzymatic activity /
- Mushroom cultivation /
- Strain variation