









-
Baccaurea ramiflora Lour. is a tropical fruit tree of the Baccaurea genus in the family Phyllanthaceae, alias Burmese Grape (English), Mafai (Thai), Latkan (Hindi), Bhubi or Lotka (Bengali), Leteku (Assamese), Lerko (Bodo)[1]. The genus name was derived from the Latin word 'Baccaurea', which refers to the golden yellow color of the fruit[2]. It is an economic forest fruit originating from northeastern India to southern China and peninsular Malaysia, and is found in Nepal, India, Myanmar, Bangladesh, southern China, the Indochina Peninsula, Thailand, and Malaysia, with its main distribution in China being in Hainan, Lianjiang in Guangdong, southern Guangxi, and southwestern Yunnan[3]. Due to its unique taste, nutritional and therapeutic values, B. ramiflora has long been exploited as a wild fruit tree, and is an important ethnobotanical plant for the residents of areas where B. ramiflora grows[4], as well as an important food source plant for the wildlife of the southern subtropical forests[5]. Scientific names, as well as common names used in other countries, are listed in Table 1.
Table 1. Different names of B. ramiflora[6].
Language Name Scientific name Baccaurea ramiflora Lour. Synonyms Baccaurea flaccida Müll.Arg, Baccaurea oxycarpa Gagnep, Baccaurea pierardi Wall, Baccaurea propinqua Müll.Arg, Baccaurea sapida (Roxb.) Müll.Arg, Baccaurea wrayi King ex Hook.f, Gatnaia annamica Gagnep, Pierardia sapida Roxb. English Baccaurea, Burmese Grape, Lantern Tree, Mafai. Nepali Kala Bogoti Hindi Khattaphal Bangla Latkan, Kusumtenga. Thai Mafai Indonesia Tampoi Kuning Chinese Mu Nai Guo, San Ya Guo, Huo Guo. Malaysia Pupor, Tempui. Vietnamese Giau Gia Dat, Giau Tien. As an under-exploited wild fruit tree, B. ramiflora has potential economic and social value that is gradually gaining attention and recognition[7]. Fruit development and quality regulation form the foundation for the further commercial development and utilization of B. ramiflora. In existing research, Luo et al. surveyed B. ramiflora germplasm resources in Guangxi, initially screening seven superior individual plants based on criteria including high yield, large fruit size, vibrant fruit color, and excellent taste and flavor[8]. Kong et al. used the number of days of fruit development and peel color as harvesting criteria, and selected color index, edible rate, and titratable acid content as representative indicators for comprehensive evaluation of the quality of B. ramiflora, which could determine the quality differences of B. ramiflora at different maturity levels[9]. The skin color of B. ramiflora was often used as an important indicator for screening of genetically improved progeny because of its phenotypic distinctiveness and correlation with quality traits[10]. Existing research on the conservation and development of B. ramiflora germplasm resources was still very limited, and its germplasm resources have not been collected and conserved. Most of the high-quality germplasm resources rely on selective retention by local residents, resulting in unstable yields and inconsistent quality of high-quality B. ramiflora varieties[11]. Its commercial fruits can only be disseminated on a small scale and are not well cultivated commercially[3]. Therefore, it was necessary to summarize the research on the regulation of fruit development and quality of B. ramiflora, which can effectively promote the screening and breeding of new varieties of B. ramiflora with higher nutritional value and better taste.
The breeding objectives of B. ramiflora include yield stability, nutritional quality, flavor, appearance color, and shelf life. With the publication of the B. ramiflora genome (NCBI: PRJNA845134), the breeding research of B. ramiflora has entered a new phase at the molecular level[3]. In this paper, we review the research progress on fruit development and quality regulation of B. ramiflora to provide a reference for molecular breeding and improvement of desirable agronomic traits in this species. The focus of the paper includes the study of nutrition and key metabolites of B. ramiflora, flavor changes during fruit ripening, and the formation mechanism of fruit color diversity in B. ramiflora.
-
In this study, the database of Web of Science core set was selected, and the different names in Table 1 were used as the subject words for retrieval. Ninety-three articles published before 1 December 2025 were obtained, and full-text and citation references were selected to export plain-text files. With reference to the approach of Lin et al., CiteSpace 6.4. R1 software was used to pretreat and further analyze the data to obtain the research trend of Muqiguo (Fig. 1)[12]. Research on B. ramiflora has undergone more than a decade of gradual accumulation and development. In 2007, Yang et al. were the first to investigate the bioactive phenolic compounds in the leaves of the milk-fruit tree and isolated two new phenolic compounds from its leaves[13]. Research activity gradually intensified after 2014 and entered a peak period after 2020 (Fig. 1a). To date, Web of Science has indexed a total of 93 publications related to B. ramiflora. Research on B. ramiflora exhibits distinct geographical relevance, with research efforts primarily concentrated in South and Southeast Asia—regions where the species occurs naturally or is traditionally cultivated (Fig. 1b). India and China stand as the two major centers of knowledge production, followed by Malaysia and Bangladesh. The keyword network takes the species name Baccaurea ramiflora as its core node, which serves as a key connector between different research themes (Fig. 1c). Antioxidant activity emerges as a critical node following the species name, closely associated with keywords such as extracts and radical scavenging capacity.
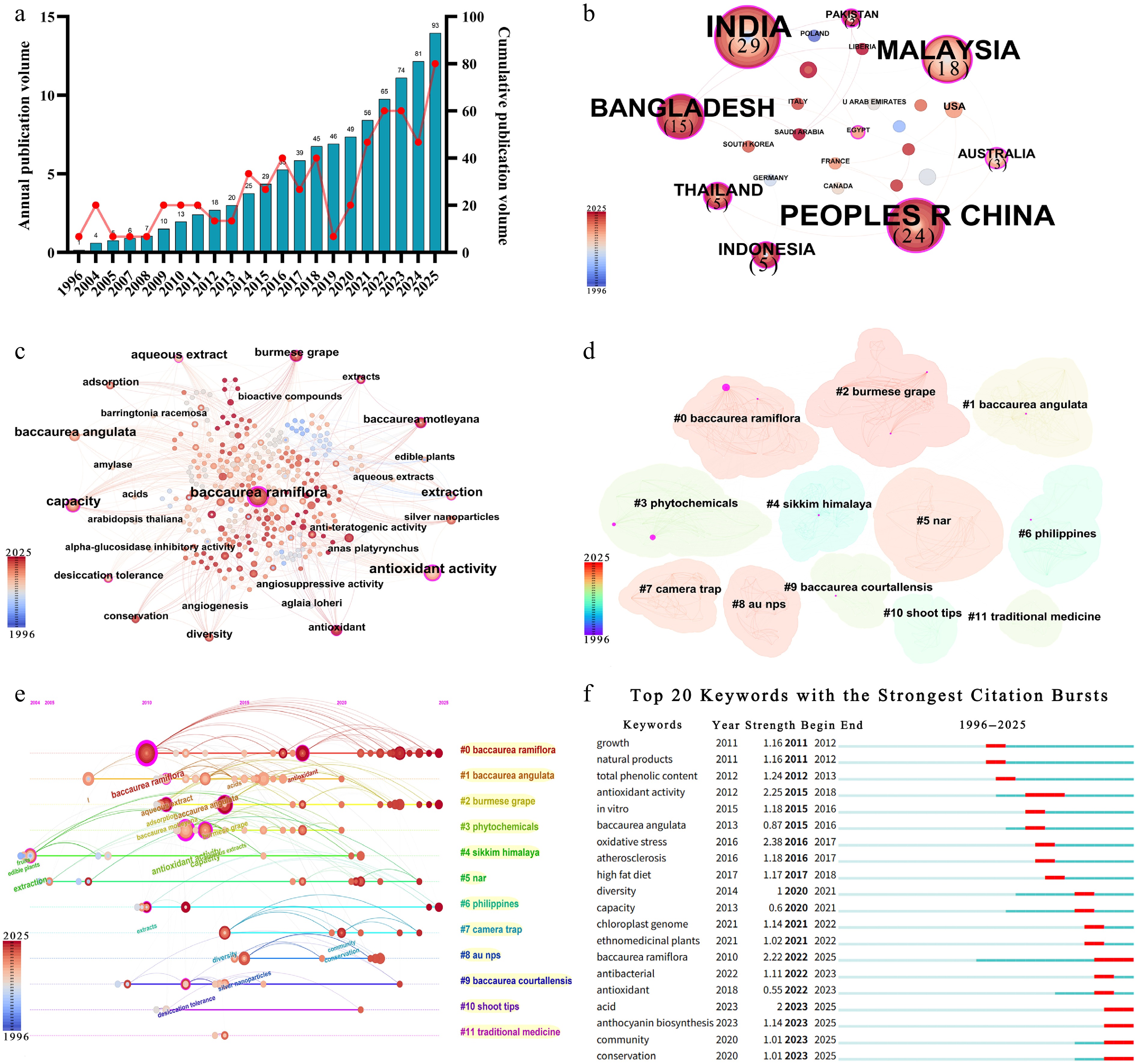
Figure 1.
Visual analysis of research trends in B. ramiflora. (a) Publication volume analysis. (b) Country-publication co-occurrence. (c) Keyword co-occurrence. (d) Keyword clustering. (e) Keyword timeline. (f) Keyword emergence map.
Cluster analysis of keywords yielded 12 clusters: #0 Baccaurea ramiflora, #1 Baccaurea angulata, #2 Burmese grape, #3 Phytochemicals, #4 Sikkim Himalaya, #5 Nar, #6 Philippines, #7 Camera trap, #8 AU NPS, #9 Baccaurea courtallensis, #10 Shoot tips, and #11 Traditional medicine (Fig. 1d). The timeline of clustered keywords is shown in (Fig. 1e), revealing a foundational layer centered on species chemistry and bioactive evaluation (#0, #1, #2, #3, #4, and #9); an application layer radiating toward multiple directions such as drug development (#11) and biotechnology (#8); and a support layer ensuring sustainable development through germplasm conservation (#10), ecological research (#5), and resource protection (#7). Detailed information on the clusters is provided in Supplementary Table S1.
Further analysis of keyword salience reveals shifts in research hotspots and emerging trends (Fig. 1f). Early-stage studies focused on exploring fundamental biological activities and mechanisms of action. Keywords with high salience during this period—such as antioxidant activity, total phenolic content, oxidative stress, and atherosclerosis—signaled a transition from simple component identification to validation of physiological functions in vivo and in vitro. Particularly, the potential for addressing metabolic diseases emerged as a key focus. Research frontiers from 2020 to the present have exhibited a dual expansion toward molecular mechanisms and resource management. The emergence of keywords like chloroplast genome, anthocyanin biosynthesis, and organic acid metabolism indicates that a growing body of studies is delving into molecular foundations—such as genetic backgrounds and secondary metabolite biosynthetic pathways—through genomics and metabolomics technologies. The persistent prominence of ethnomedicinal plants, community, and conservation—combined with studies on antibacterial activity—signals an expanded research perspective encompassing ethnobotany and sustainable utilization.
Overall, research on B. ramiflora has gradually gained attention, establishing a solid foundation in areas such as nutritional value, medicinal functions, and resource development. However, studies on the physiological and biochemical changes during fruit development, key regulatory factors influencing quality formation, and the impact of environmental factors on fruit quality remain insufficiently explored, and the research field lacks a stable, focused research direction. Furthermore, advances in omics technologies have propelled B. ramiflora research toward the in-depth exploration of molecular foundations, including genetic backgrounds and secondary metabolite biosynthetic pathways.
-
Fruit growth and development constitute a biological process governed by complex regulatory mechanisms, which ultimately determine fruit quality formation and are one of the core factors influencing the commercial value of fruits[14]. The widespread adoption of whole-genome sequencing over the past decade has advanced our understanding of traits related to fruit growth and development, including yield stability, fruit development dynamics, varietal domestication, and epigenetic regulation of fruit size[15]. These studies provide developmental directions and insights into the regulation of fruit growth and development in B. ramiflora. The phenological characteristics of B. ramiflora are shown in Fig. 2. B. ramiflora is a typical cauliflorous tropical dioecious plant, with apetalous flowers arranged in racemose panicles. Both male and female flowers are yellow with four to six sepals; male flowers contain four to eight stamens, and female flowers have a three-loculed ovoid ovary (Fig. 2)[7]. The fruiting trees started to bud in early February, and male plants flowered first, with white inflorescences, from February to March[8]. Female plants flowered later, with larger, dark red inflorescences, from mid-March to mid-April[8]. The young fruit developmental stage spans from mid-April to mid-July, with a total fruit development period of approximately 90 d. Fruits mature in mid- to late July, with a ripening period of 20–30 d; the earliest ripening occurs in early July, and the latest in early August[8]. Most B. ramiflora are propagated from seedlings, which generally take five to seven years to flower and fruit, entering the full fruiting period after ten years. The yield of a single plant varies relatively widely, with a yield of 45–500 kg per plant[8]. The fruit is ellipsoid to round, turning yellow or yellow-brown at maturity, and is a typical berry. Its edible part is the aril, and the leathery pericarp encloses three to four seeds[16]. Yang et al. proved that the ripening of B. ramiflora started 73 d after flowering and reached the ripening standard in 93 d[17]. Kong et al. used the number of days of fruit development and skin color as harvesting standards, and suggested that the fruit should be harvested at 91–98 d after anthesis, at which time the skin is pink or yellowish-pink, and the fruit is of good quality and flavor for fresh eating, and the fruit at 98 d after anthesis is of good quality and high storability, which is more suitable for the fresh marketing and storage of B. ramiflora after harvesting[9]. In addition, it has been documented that B. ramiflora in Hainan and Southeast Asia ripened about 45 d earlier than inland China[3]. Existing research has yielded relatively clear findings on the critical periods of fruit growth and development in B. ramiflora, but the underlying molecular regulatory mechanisms remain poorly understood.
-
Studies on B. ramiflora fruits have advanced steadily from the identification of its biological activities to the in-depth exploration of its metabolites[7,11]. To date, 138 compounds have been reported from B. ramiflora, which are categorized into 105 unique compounds after statistical analysis, with 22 duplicate entries detected. These compounds have been isolated and identified from different tissues of B. ramiflora: 40 compounds in the stem, 38 in the leaves, 35 in the roots, 24 in the fruits, and one in the seeds (Supplementary Table S2). The pulp of B. ramiflora is sweet, sour, and tender and can be eaten fresh as well as processed into juices, preserved fruits, jams, and fruit wines[1]. B. ramiflora also has better medicinal functions, such as anti-inflammatory, anti-tumor, and antioxidant activity[20]. B. ramiflora is rich in nutrients with 49.2% palatability, 84.7% water content, 0.06% fat, 0.29% fiber, 1.57 mg/100 g vitamin C, 1.99% titratable acid, and 11.87% total sugar, making it a good source of sugar, vitamin C, phenolics, and amino acids[21]. The pulp of B. ramiflora was low in fat, ash, and protein, but rich in dietary fiber, ascorbic acid (vitamin), and mineral elements such as calcium, magnesium, and phosphorus[22,23]. Chen et al. analyzed the metabolites of the pulp during the ripening stage of B. ramiflora by non-targeted metabolomics, and in the fully ripened stage of B. ramiflora, a total of 541 metabolites were identified, with 361 positive ion patterns and 201 negative ion patterns, of which 21 were duplicated[11]. The main components affecting fruit flavor include primary metabolites: 12 carbohydrates, three organic acids, seven amino acids, and eight amino acid derivatives, two vitamins, and 41 fatty acids; and secondary metabolites: 42 flavonoids, eight phenols, six phenolic acids, 26 phenylpropanoids, four steroids and five steroid derivatives, and 75 terpenoids[11]. The main components and differential metabolites are shown in Supplementary Table S3. Compared with previous studies, non-targeted metabolomics has enabled more systematic identification of chemical components in B. ramiflora pulp. The progress of research on compound identification to metabolite identification and analysis of B. ramiflora was shown in Fig. 3.
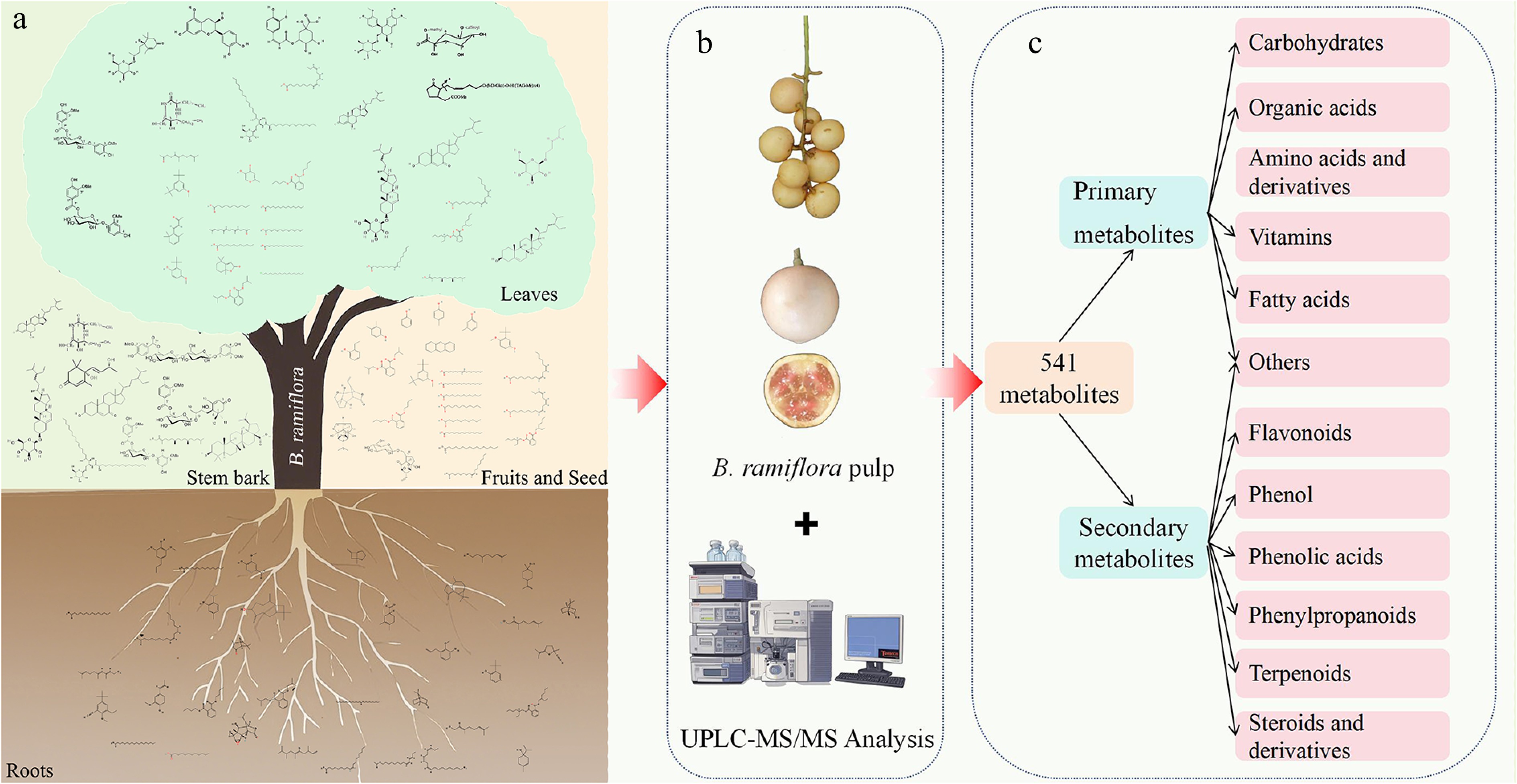
Figure 3.
Advances in the study of compound and metabolite diversity. (a) Compounds from different parts of the B. ramifora. (b) UPLC-MS/MS analysis of the pulp of the B. ramifora. (c) Main metabolite components of the pulp.
Fruit flavor
-
Taste and flavor are the most critical determinants of fruit quality. To stay competitive, new cultivar introductions increasingly require excellent sensory properties, resulting in the integration of sensory and consumer research into many fruit and vegetable breeding programs[24]. In the fruit metabolome, key flavor-differentiating metabolites determine fruit flavor quality by affecting aroma, sweetness, acidity, and bitterness. The genetic and domestication mechanisms underlying these metabolites are critical for fruit quality improvement and molecular breeding[25,26]. Chen et al. revealed the key components of the flavor of B. ramiflora through the non-targeted metabolomics, with D-(+)-glucose and citric acid determining its sweetness and sourness, respectively[11]. Eighteen differential metabolites were annotated to the flavor formation metabolic pathway in B. ramiflora, including L-Sorbose, D(−)-Salicin, 2-isopropylmalic acid, citric acid, L-Tryptophan, 5-hydroxyindole-3-acetic acid, spermine, dehydroascorbic acid, N-acetyl-L-phenylalanine, tryptamine, salidroside, L-Tyrosine, pantothenic acid, γ-linolenic acid, linoleic acid, 9-OxoODE, and jasmonic acid[17,18]. Yang et al. revealed differences in the quality metabolism of B. ramiflora through non-target metabolomic analyses of Baccaurea ramiflora 'LR' cultivar at different developmental periods[17], and hypothesized that the decline in citric acid content during ripening is a core driver of sourness reduction in B. ramiflora fruits. Another study by Yang et al. showed that L-sorbose and 5-hydroxyindole-3-acetic acid were closely correlated with the sugar-acid ratio and titratable acidity during fruit development, and confirmed these two metabolites as potential key flavor biomarkers of B. ramiflora via non-targeted metabolomic analysis of the B. ramiflora 'BR' at different developmental stages[18]. A significant rise in the key taste-differentiating metabolite L-sorbose affected a significant decline in the downstream substance 5-hydroxyindole-3-acetic acid, and a significant decline in citric acid affected a significant rise in the downstream substance Spermine. The results indicated that the elevation of the major sugars and the decrease of the major acids affected the major taste changes in the fruit of B. ramiflora.
Chen et al. obtained 39 key candidate genes for the quality of ripening fruit in different strains of B. ramiflora by comparative transcriptome analyses of ripening fruits of different strains of B. ramiflora[27]. Among them, AMY, BGL, and BG were associated with the increase of pulp sugar content, ALDH2 was associated with the inhibition of pulp de-astringency, PME and PL were important in promoting fruit softening and juice content, and ADH, TSB, and DHQ-SDH were associated with the accumulation of fruit aromatic substances[27]. A metabolic pathway for flavor formation in B. ramiflora fruit was mapped by integrating the related studies (Fig. 4).
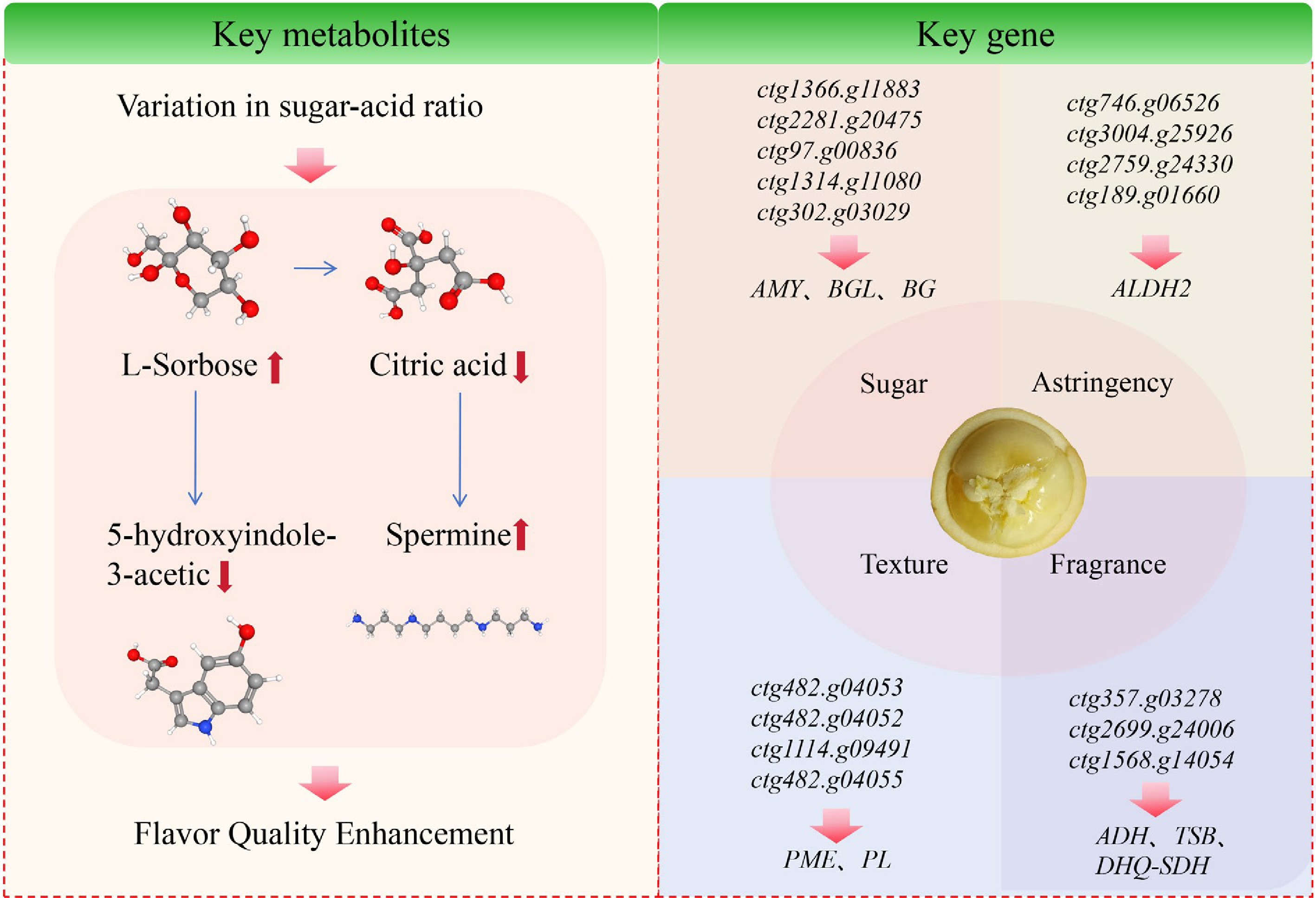
Figure 4.
Key metabolites and genes associated with fruit flavor in B. ramiflora. Notes: changes in key metabolites were comprehensively mapped by comparison of metabolites from five different periods of B. ramiflora 'LR' and 'BR'[17,18]. The key genes were mapped for the comparison of fruit quality between B. ramiflora 'LR' and 'BR' at maturity[27]. The full names of the key genes for B. ramiflora quality can be found in Supplementary Table S4. ↑ = significantly up, ↓ = significantly down (p < 0.01).
Fruit color diversity
-
Fruit color is a key trait influencing consumer preference, and fruits exhibit diverse color phenotypes, including red, pink, reddish-purple, purple, and blue, which are mainly determined by the accumulation of flavonoids (especially anthocyanins) and carotenoids[28]. Flavonoids are widely found in plants and are secondary metabolites synthesized in the phenylalanine pathway. Anthocyanins are a major subclass of flavonoids, which have been extensively studied as water-soluble natural pigments. The six most common anthocyanidins (anthocyanin aglycones) are pelargonidin, cyanidin, petunidin, malvidin, peonidin, and delphinidin[29]. Cyanidin is the most widespread in fruits, typically conferring pink to red hues; pelargonidin produces orange-red coloration; peonidin yields purplish-red tones; while petunidin, malvidin, and delphinidin usually give bluish-purple color, which are less abundant in most edible fruits. Anthocyanins retain the core biological activities of flavonoids, with proven efficacy in preventing cardiovascular diseases[30], diabetes[31], anti-cancer and heart disease[32,33]. Therefore, the colors presented by flavonoids in fruits and their values are gradually being discovered, which has led to their particular popularity among consumers and can be used as an important indicator for fruit quality studies.
In recent years, researchers have increasingly focused on the formation mechanism of pigment accumulation in plant tissues, and the anthocyanin biosynthetic pathway has been extensively studied worldwide. It is well established that the anthocyanin biosynthetic pathway is the downstream branch of the flavonoid metabolic pathway, and its biosynthetic framework is highly conserved across different plant species[34,35]. Both anthocyanosides and flavonoids are synthesized by the phenylalanine pathway, which undergoes three main stages: the early stage of phenylalanine cleavage, the middle stage of flavonoid metabolism, and the late stage of anthocyanoside synthesis[34,36]. A total of 45 structural genes related to the flavonoid and anthocyanin biosynthesis pathways were identified in the pulp of B. ramiflora, and 38 genes were differentially expressed[37]. The Anthocyanin biosynthetic pathway and key genes in B. ramiflora are shown in Fig. 5.
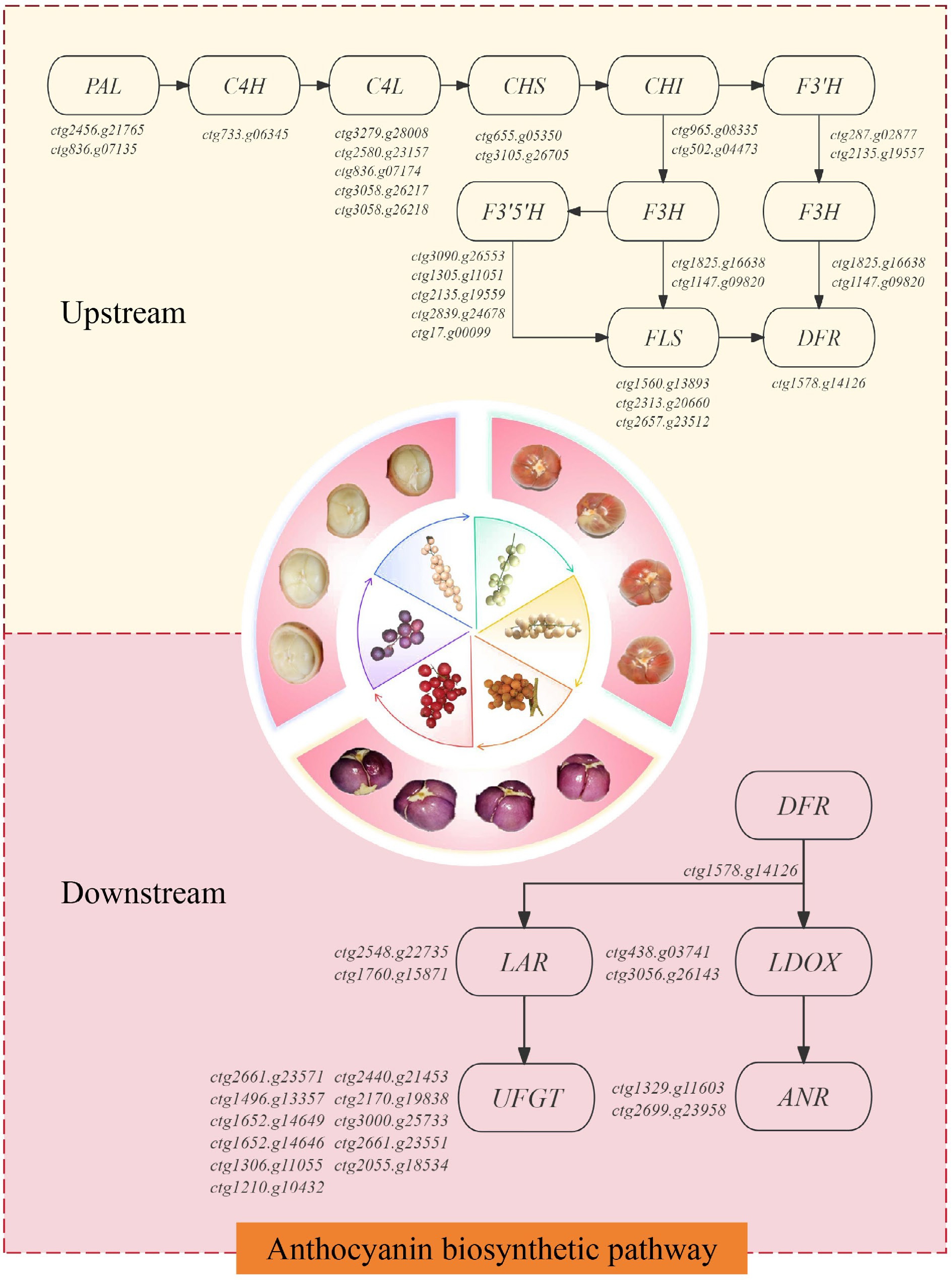
Figure 5.
Anthocyanin biosynthetic pathway in B. ramiflora. Notes: The pathways in yellow represent upstream genes of the Anthocyanin biosynthetic pathway; the pathways in red represent downstream genes of the Anthocyanin biosynthetic pathway. The full names of the key genes for B. ramiflora quality can be found in Supplementary Table S4.
Differences in fruit color of B. ramiflora are important indicators for quality screening of B. ramiflora, and previous studies have only provided assistance for quality screening of B. ramiflora from a phenotypic perspective[38]. Through integrated multi-omics analysis, progress has been made in elucidating the mechanisms underlying fruit color formation in B. ramiflora, providing a scientific basis for quality improvement of this species[18]. As a wild fruit tree, the fruit color of B. ramiflora was one of the important features for the discrimination between pollinators and seed dispersers[5,39]. Field surveys found that B. ramiflora are mostly light-colored at low latitudes and low altitudes, and dark-colored at high latitudes and high altitudes[38]. Hainan and Guangdong mostly showed yellow pericarp; Guangxi Fangchenggang and Dongxing were dominated by yellow, white, and pink pericarp, and Longzhou to Jingxi were dominated by pink, red, and purple pericarp; and Yunnan was dominated by red and purple pericarp with pink or purple flesh[38]. Similar to the study by Shi et al., fruit color was significantly correlated with different altitudes; apples exhibited a deeper red color at higher altitudes, and this color change was primarily attributed to the accumulation of anthocyanins[26]. It was found that the concentration of flavonoids such as root bark glycosides, huangqi glycosides, and hesperidin varied significantly among different varieties of B. ramiflora, which might be the reason for the pink color of B. ramiflora fruits, and that cornflower pigments might be the key metabolites for the pink color[11]. Existing studies have provided directions for the study of fruit color in B. ramiflora, but there is still no complete study on the formation mechanism of fruit color diversity in B. ramiflora fruits, and the key substances and candidate genes for coloring in different varieties and colors of B. ramiflora still need to be further explored.
Key pathway research
-
During the development of B. ramiflora, the expression of key genes in sugar-acid metabolism and anthocyanin synthesis pathways exhibits continuous yet stage-specific regulatory patterns (Fig. 6). The synergistic changes in these two metabolic pathways may jointly influence the formation of fruit flavor and color. In the sugar metabolism pathway, Stage I features upregulation of genes such as HK and PFK9, promoting glucose conversion to fructose-1,6-bisphosphate. Glycolysis shows pckA upregulation and PK downregulation, while organic acid metabolism exhibits IDH downregulation, suggesting this stage prioritizes sugar breakdown with reduced organic acid synthesis. In Stage II, INV is downregulated while SPS, FK, and some HK genes are upregulated, promoting sucrose synthesis and rapid fructose conversion. PK is upregulated in glycolysis, while ACO, IDH, and MDH are downregulated, and NADP-ME is upregulated in organic acid metabolism, collectively driving organic acid accumulation in the pulp. In Stage III, multiple sugar metabolism genes (INV, SUS, SPS, HK) are significantly upregulated, promoting rapid sugar accumulation; PFK9 is upregulated in glycolysis, while MDH and NADP-ME expression increases in organic acid metabolism. In Stage IV, only sugar metabolism remains active, with INV and HK continuously upregulated to enhance phosphorylation processes. In Stage V, INV exhibits differential regulation, while HK and FK are upregulated to promote sucrose breakdown and hexose interconversion. Upregulation of pckA and some PKs in glycolysis, coupled with IDH downregulation in organic acid metabolism, indicates diminished organic acid synthesis as sugar accumulation peaks[19].
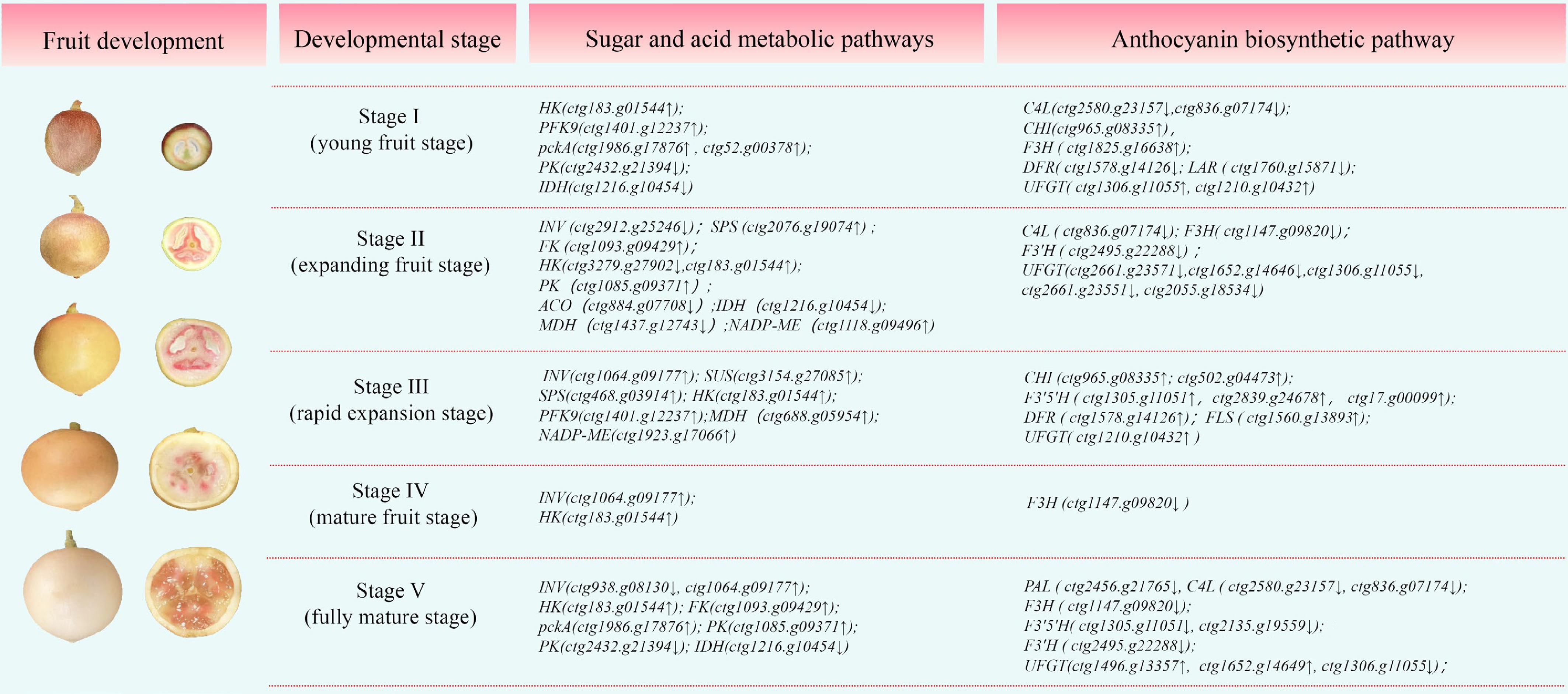
Figure 6.
Key genes at different developmental stages. Notes: ↑ = significantly up, ↓ = significantly down (p < 0.01). The full names of the key genes for B. ramiflora quality can be found in Supplementary Table S4.
In the anthocyanin pathway, structural gene expression changes across developmental stages significantly influence flavonoid and anthocyanin synthesis. Downregulation of genes like C4L in Stage I may initiate flavonoid synthesis pathways; Stage II downregulation of C4L, F3H, and multiple UFGT/3GT genes inhibits anthocyanin accumulation. Stage III witnessed upregulation of key genes, including CHI, F3'5'H, DFR, FLS, and UFGT, potentially promoting flavonoid degradation and anthocyanin conversion. Stage IV showed downregulation of only F3H, likely inhibiting anthocyanin synthesis. In Stage V, multiple genes, including PAL, C4L, and F3H, are downregulated, accompanied by upregulation of F3'5'H and some UFGT/3GT genes. Notably, C4L and F3H remain downregulated throughout development, suggesting they may indirectly promote anthocyanin accumulation by inhibiting competing pathways[18].
-
In recent years, as a wild fruit tree with potential economic value, some progress has been made in the study of its fruit quality formation. However, compared with other well-studied tropical fruit trees, molecular breeding studies on B. ramiflora remain in their infancy[40]. In this paper, we systematically summarized the developmental dynamics, nutritional and metabolic characteristics, and the regulatory mechanisms of flavor and fruit color of B. ramiflora, and revealed the genetic basis of its quality diversity, but there are still many key issues to be solved.
Germplasm resources are the material basis for genetic breeding, and the level of genetic diversity represents the genetic richness of the species in a particular environment[41]. Currently, high-quality B. ramiflora germplasm resources rely on sporadic, informal preservation by local farmers, with insufficient systematic germplasm collection and genetic diversity characterization, and a lack of a standardized collection and evaluation system[11]. To date, only two studies have reported the genetic diversity of B. ramiflora: Luo et al. analyzed the genetic diversity of 37 B. ramiflora germplasms from different sources using SCoT markers[42]. Zhou et al. analyzed the genetic diversity of 63 B. ramiflora germplasm resources from Guangxi, Yunnan, Hainan, and Guangdong, and constructed the corresponding DNA fingerprints[43]. For germplasm collection and conservation, there is still no unified technical standard for B. ramiflora germplasm resources, nor has a specialized germplasm repository been established. Therefore, the top priority for future B. ramiflora research is to strengthen the systematic collection and in-situ/ex-situ conservation of its germplasm resources. Through the extensive collection of wild and cultivated germplasm of B. ramiflora with different geographic distributions, the establishment of a standardized preservation system, and the construction of a comprehensive evaluation model by combining phenotypic and genotypic data, we can screen the high-quality resources with strong resistance and wide adaptability, and lay the material foundation for the subsequent development.
DNA sequence variation identified by genome resequencing studies provides a valuable resource for breeding and crop improvement, which can be used to introduce specific alleles, predict hybridization performance, and design more effective breeding strategies[44]. The release of the B. ramiflora reference genome has laid a foundation for molecular marker development, but most of the existing studies have focused on single varieties or a small number of samples, making it difficult to comprehensively resolve their genetic diversity[3]. Using whole-genome resequencing to analyze the genetic diversity of B. ramiflora, we can develop highly specific molecular markers for rapid identification of germplasm resources and kinship analysis. Meanwhile, key functional genes can be identified via genome-wide association analysis, and a trait-gene association database can be established to provide theoretical support for molecular marker-assisted breeding. Drawing on the successful experiences of watermelon[45] and mulberry[46], we need to locate the key loci for fruit size, sugar-acid ratio, fruit color, and other traits through large-scale resequencing of germplasm resources combined with genome-wide association analysis (GWAS), so that we can provide the target points for molecular markers to assist breeding. Targets for molecular marker-assisted breeding.
Through accurate identification of trait-associated genes in B. ramiflora germplasms, we can construct a core germplasm collection for this species by selecting representative genetic materials that capture its full genetic diversity to support efficient breeding and research. We can also develop DNA fingerprints to accurately distinguish B. ramiflora accessions, prevent germplasm mixing, and provide a scientific basis for plant variety rights protection. Population genetics approaches can be applied to reconstruct the evolutionary history of B. ramiflora, elucidate its genetic structure and kinship among germplasms, and inform the rational utilization of resources and the development of conservation strategies. Furthermore, trait association analysis can be performed to clarify the correlation between candidate genes and key agronomic traits, providing critical information for molecular marker-assisted breeding and functional gene cloning.
In terms of genetic resources innovation, we will focus on making breakthroughs in nutritional quality improvement and improving the content of vitamins, minerals, antioxidants, and other nutrients in B. ramiflora by means of gene editing and molecular marker-assisted selection, so as to satisfy consumers' demand for healthy food. For flavor quality, excavate and utilize key genes controlling flavor components such as aroma, sweetness, and acidity of B. ramiflora, and cultivate new varieties with strong fruity aroma and suitable sweet-sour ratios, so as to enhance consumers' taste experience. For color, research on the gene regulatory mechanism that affects the formation of the color of B. ramiflora, and develop varieties with bright color and market appeal. In addition, B. ramiflora exhibits specific adaptability to low-light environments, giving it high potential for cultivation in understory economic systems. However, the differences in light adaptability across different seedling stages and seed sources, as well as the underlying regulatory mechanisms, remain poorly understood, and the associated candidate genes warrant further exploration[47]. At the same time, genetic resources innovation research should be actively carried out to improve the fruit size, shape and storability of B. ramiflora, so as to provide diversified and excellent varietal resources for the sustainable development of the B. ramiflora industry, and ultimately to realize the breeding goal of providing consumers with tastier, longer-lasting, healthier, and more beautiful B. ramiflora, so as to promote the development of the B. ramiflora industry in the direction of high-quality and high-efficiency.
Finally, there are still four core gaps in current research. First, the hormonal regulatory network governing fruit development and quality formation is completely uncharacterized: no studies have defined the roles of core phytohormones in fruit expansion, ripening initiation, sugar-acid accumulation, or color transition, nor have they elucidated the crosstalk between hormone signaling pathways and the sugar-acid and flavonoid metabolic pathways. Second, the effects of environmental factors on fruit quality and their underlying mechanisms lack systematic investigation: existing research only reports correlations between latitude/altitude and fruit phenotypes, with no systematic dissection of how key environmental variables regulate fruit sugar-acid ratio, flavor metabolite accumulation, and peel coloration. This gap prevents the development of region-specific cultivation management practices to optimize fruit quality. Third, postharvest physiology and storage mechanism research is severely deficient: beyond a preliminary assessment of harvest timing on storability, no studies have classified the respiratory climacteric type of B. ramiflora fruit, characterized the physiological and molecular mechanisms of postharvest softening, browning, flavor loss, and pathogen infection, or developed targeted postharvest preservation technologies for commercial distribution. Fourth, the genetic basis of quality traits and a functional molecular breeding system have not been established: there are no published QTL mapping or GWAS analyses for key agronomic traits, no validated trait-linked molecular markers, and no stable genetic transformation system for the species, leaving molecular breeding for B. ramiflora in its infancy relative to other tropical fruit crops.
-
As a fruit possessing both market potential and scientific research value, studies on the fruit development and quality regulation of B. ramiflora not only deepen our understanding of its unique biological characteristics but also provide scientific support for the sustainable development of the industry. In this study, using CiteSpace to conduct a bibliometric analysis of research trends, through systematic exploration of the regulatory mechanisms governing fruit formation, developmental processes, and quality traits such as flavor and color, we have identified key factors influencing B. ramiflora quality. These findings not only aid in optimizing cultivation management practices to enhance fruit quality and yield but also provide theoretical foundations and technical guidance for varietal improvement. Moving forward, efforts should intensify to study and utilize B. ramiflora genetic resources, deeply explore its superior genes, and combine modern biotechnological approaches to develop new varieties with excellent traits that meet market demands. Concurrently, strengthening academia-industry collaboration will drive the large-scale and standardized development of the B. ramiflora industry, enhance its overall market competitiveness, and facilitate the market penetration of this unique fruit to satisfy consumers' diverse demands.
We sincerely thank all the authors of the literatures cited in this paper, whose pioneering and fundamental research on Baccaurea ramiflora provided essential theoretical and data support for this work. We also thank the editors and anonymous reviewers for their constructive comments on this manuscript. This research was supported by the program for scientific research start-up funds of Guangdong Ocean University (Grant Nos 060302052305 and 060302052102), Forestry Science and Technology Innovation Project of Guangdong province (Grant No. 2018KJCX023), Guangdong Provincial Key Laboratory of Functional Substances in Medicinal Edible Resources and Healthcare Products (Grant No. 2021B1212040015).
-
The authors confirm their contributions to the paper as follows: study conception and design: Chen J, Ye X, Huang J; sampling: Yang C, Luo P, Ye X, Huang J; bioinformatics analysis: Wang H; data processing, draft manuscript preparation: Chen J, Yang C, Zhu R; manuscript polishing: Yang C; writing − review and editing: Ye X, Huang J. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed in the current study are available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
-
Received 5 December 2025; Accepted 30 March 2026; Published online 25 May 2026
- Supplementary Table S1 Keyword clustering information.
- Supplementary Table S2 Compounds isolated from different parts of B. ramiflora. (Repeat are shown in bold).
- Supplementary Table S3 The main metabolites and differential metabolites of B. ramiflora [1].
- Supplementary Table S4 Key genes affecting the flavor of B. ramiflora.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen J, Yang C, Zhu R, Wang H, Luo P, et al. 2026. Fruit development and quality regulation of Baccaurea ramiflora: a review. Tropical Plants 5: e015 doi: 10.48130/tp-0026-0014 

Fruit development and quality regulation of Baccaurea ramiflora: a review
- Received: 05 December 2025
- Revised: 21 February 2026
- Accepted: 30 March 2026
- Published online: 25 May 2026
Abstract: Baccaurea ramiflora Lour., an evergreen tree belonging to the genus Baccaurea (family Phyllanthaceae), is native to northeastern India, southern China, and the Malay Peninsula, with a natural distribution spanning South Asia, Southeast Asia, and southern and southwestern China. As an underutilized wild fruit tree, it possesses considerable economic, edible, ecological, and medicinal value. Research on B. ramiflora commenced in 1996, with study activity intensifying markedly after 2020. Early work focused primarily on bioactive components and metabolic profiling, before research expanded into molecular regulatory mechanisms and germplasm resource management. As a dioecious, cauliflorous species, B. ramiflora has a fruit development period of approximately 90 d. Fruit ripening initiates at approximately 70 d after anthesis, with the optimal harvesting window occurring 91–98 d post-anthesis, when fruit quality and storability are highest. Notably, fruit ripening of B. ramiflora grown in Hainan (China) and Southeast Asia occurs approximately 45 d earlier than that of plants grown in inland China. Currently, the pathways and regulatory factors involved in sugar-acid metabolism and anthocyanin metabolism during fruit development and quality formation are relatively well characterized. However, molecular breeding for B. ramiflora remains in early stages. Systematic collection and conservation of germplasm resources are lacking, and the molecular regulatory networks underlying fruit quality traits are not fully elucidated. Future research should prioritize systematic germplasm collection and evaluation, whole-genome resequencing coupled with molecular marker development, and the identification of functional genes governing fruit quality.
-
Key words:
- Baccaurea ramiflora /
- Fruit development /
- Quality control /
- Metabolic pathway
