









-
Drought and salinity are significant abiotic factors that often occur together in natural ecosystems, affecting plant growth and development[1]. Drought is caused by water scarcity in arid regions and irregular precipitation patterns, while salinity is mainly caused by saline soil, irrigation with saline water, evaporation, and the precipitation cycles[2,3]. Drought and salinity in soil and water are recognized to diminish the water-absorbing capacity of plants, leading to loss of osmotic equilibrium, cell dehydration, and eventual death[4]. In addition to osmotic imbalance under salinity, ionic toxicity and nutritional imbalances also exist[1]. Therefore, the physiological alterations caused by drought and salinity stresses are influenced by shared or distinct stress signaling pathways that regulate downstream gene expression and function across various metabolic pathways, thereby facilitating plant adaptation to these environmental challenges.
Grasses (Poaceae), which are vital for agriculture and ecosystems, respond to combined drought and salinity stress by integrating the molecular mechanisms underlying tolerance to both stresses[1,5,6]. For instance, drought and salinity tolerance simultaneously enhanced by increasing the antioxidant enzyme activity (e.g., peroxidase [POD], catalase [CAT], and superoxide dismutase [SOD]) following the exogenous application of polyamines (e.g., putrescine, spermidine, and spermine) in bermudagrass (Cynodon dactylon L.)[6]. Similarly, the induction of salt and drought-responsive genes occurred upon exogenous application of γ-aminobutyric acid (GABA) in creeping bentgrass (Agrostis stolonifera L.), involving Ca2+, and reactive oxygen species (ROS) signaling as well as calcium-dependent protein kinases (CDPKs)-mediated signal transduction[7−9]. These dual improvements suggest the involvement of a common regulatory mechanism that confers tolerance to both stresses. Recent omics studies in several common turfgrasses under drought and salt stress revealed that both plant hormone signaling pathways and mitogen-activated protein kinase (MAPK) signal transduction pathways were significantly enriched[10−16].
Grasses also have unique responses to salt and drought stress. In switchgrass (Panicum virgatum L.), 152 transcription factors (TFs) belonging to families such as WRKY, BES1, NF-X1, and Nin-like were differentially regulated in response to drought, and 31 TFs from families including AP2, ZF-HD, HSF, and LBD were exclusively upregulated in response to salt stress[5]. However, 87 TFs, including members of the bHLH, bZIP, NAC, G2-like, MYB-related, GRAS, and ERF families, were upregulated in response to both salt and drought stresses[5]. Furthermore, a greater number of differentially expressed genes were identified under drought stress than under salt stress in switchgrass, consistent with the pattern observed in Arabidopsis[5,17]. Suggesting that drought stress imposes a more substantial physiological challenge, requiring broader and more complex cellular responses than salt stress[5,18].
Grasses exhibit both shared and distinct molecular responses to drought and salinity stress compared with grass and non-grass model plants. For example, among all differentially expressed unigenes in tall fescue (Festuca arundinacea Schreb.) in response to drought stress, 16.46% showed no detectable homology to any of the three model plant genomes (Arabidopsis, O. sativa, and Brachypodium distachyon), wherease 83.54% were putative homologues of these model plants[19]. Recent findings indicate that in the ABA-induced MAPK cascade associated with abiotic stress responses by regulating antioxidant defense and ROS homeostasis, the MAPKKK17/18–MKK3 module is conserved between Arabidopsis and red fescue (Festuca rubra L.). At the same time, the downstream MAPK component differs[16]. These differences likely reflect the unique evolutionary history and physiological adaptations of grasses. In particular, the emergence of specialized photosynthetic strategies, such as C4 photosynthesis and, in rare cases, CAM-like pathways, may have contributed to lineage-specific regulatory and stress-response mechanisms[20]. Therefore, stress responses in grass species warrant dedicated investigations and classification.
In this review, the convergent and divergent roles of key regulatory modules, the abscisic acid (ABA) and MAPK pathways, are examined, along with their interplay with other signaling components, transcription factors, and post-transcriptional mechanisms coordinating plant adaptation to drought and salinity stress. Accordingly, ABA and MAPK pathways have been conceptualized as a central regulatory hub that integrates diverse upstream stress-derived signals and coordinates downstream transcriptional reprogramming. Although multiple signaling modules, including Ca2+, ROS, GABA, transcription factors, and non-coding RNAs, contribute to stress responses, they do not operate as isolated pathways. Instead, these modules either converge on, modulate, or partially bypass the ABA-MAPK core to fine-tune stress-responsive transcription. This conceptual framework enables the identification of signals integrated into the ABA-MAPK axis in grasses in response to salt and drought stress.
-
Signaling networks in plants comprise various interconnected modules, each of which plays a distinct role in perceiving, transmitting, and executing responses to environmental and developmental signals[15,21]. The ABA signaling pathway, mediated by a key isoprenoid hormone[22], regulates growth and stress adaptation by maintaining a dynamic balance among ABA biosynthesis, degradation, and transport[1,13]. Additionally, the MAPK cascade, a conserved eukaryotic signaling module, transmits stress signals to downstream target genes[23] (Fig. 1). Transcriptome analysis of carpetgrass (Axonopus compressus L.) under drought stress and the halophytic grass Puccinellia nuttalliana under salt stress revealed that the most significantly enriched signaling pathways were hormone signaling pathways and the MAPK signaling pathway, which can be either ABA-dependent or independent[24,25]. The ABA and MAPK signaling pathways do not function in isolation. Still, they are intricately interconnected with other signaling pathways, including Ca2+, ROS, and phytohormones, forming a system that fine-tunes plant stress responses[10,22,26−30].
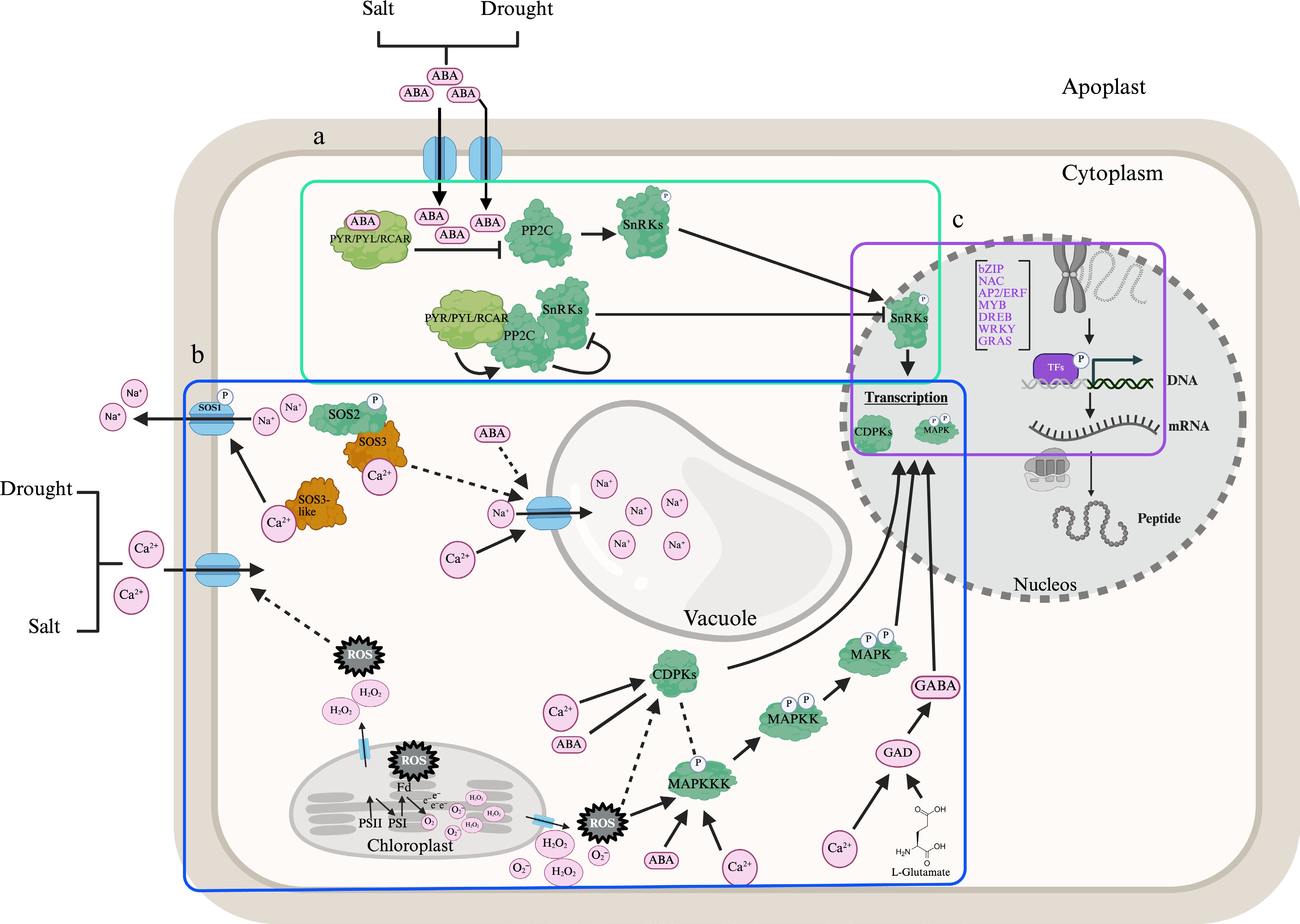
Figure 1.
Schematic representation of ABA and MAPK signaling pathways integrated with CDPK, ROS, and GABA signaling pathways regulating transcriptional responses under salt and drought stress. Stress signaling pathways are divided into three main modules, indicated in the figure. (a) ABA perception and signal initiation (the cyan rectangle). Under drought and/or salinity stress, endogenous ABA levels increase[13,31], which promotes the binding of ABA to PYR/PYL/RCAR receptors[32]. This causes PYR/PYL/RCAR to recruit protein phosphatase 2C (PP2C[17]) and to inhibit PP2C phosphatases, leading to activation of SNF1-related protein kinases (SnRK2s[33,34]). In the presence of ABA, activation of SnRK2s leads to phosphorylation of downstream target proteins[35], which subsequently regulate transcription. (b) MAPK cascade activation, ROS, SOS, Ca2+, and GABA signaling (the blue rectangle). Stress signals (e.g., ABA, Ca2+, H2O2, O2−, GABA) activate a conserved MAPK cascade[35] involving sequential phosphorylation of MAPKKKs, MAPKKs, and MAPKs[11,13,14]. CDPK, MAPKs (e.g., MAPK1), and ion transporters such as SOS1 have been associated with drought and salt stress[1,10,29]. Stress-induced cytosolic Ca2+ signals activate the SOS signaling pathway. ABA induces changes in chloroplast activity and increases ROS production that further contribute to MAPK cascade activation and antioxidant enzyme responses[36]. Low cytosolic pH and high Ca+2 concentration activate glutamate decarboxylase (GAD), which catalyzes GABA biosynthesis from L-glutamate[36]. (c) Transcriptional regulation (the violet rectangle), activated SnRK2s, CDPKs, and MAPKs phosphorylate transcription factors (e.g., bZIP, NAC, AP2/ERF, MYB, DREBs, WRKY, GRAS, and AREBs/ABFs), which bind to cis-regulatory elements in promoters of stress-responsive genes. The target genes mediate protective responses, including osmolyte accumulation, ROS detoxification, and ion homeostasis[37]. Arrows indicate directional signaling. Solid lines represent experimentally established interactions. Dashed lines indicate hypothetical interactions. Arrow connections represent positive regulation, and block connections indicate negative regulation. Different modules of the ABA signaling pathway are color-coded for clarity. Blue: channels and transporters. Pink: signaling molecules. Light green: ABA receptors. Dark green: phosphatases and kinases. Purple: Transcription factors. Orange: Ca2+ receptors. Key interactions and gene functions were compiled from the following studies: (AsCDPK, AsSOS1, MAPKs under salinity[1,21,29,38−40]) (CDPK26 and MAPK1 in drought responses[10,12,13,17]) (SnRK2s in ABA-dependent pathways[33,41]), (PYL/RCAR5 and SnRK2s in ABA signaling under salt and drought stress[19,32,42,43]). The hypothetical interaction between ROS and Ca2+ influx is supported by evidence from model species showing that ROS elevates cytosolic Ca2+ concentration in guard cells[44]. The proposed hypothetical CDPKs-MAPKKKs interaction is inferred from their concurrent transcriptional induction under salt stress in P. nuttalliana and previously reported signaling cross-talk in Arabidopsis[24,45]. The putative link between CDPK-ROS signaling and between SOS3-NHX is based on evidence from wheat[46,47]. At the same time, the hypothetical interaction between ABA and NHX is supported by studies in Brachypodium[48].
-
Abiotic stress responses in plants are triggered by sensing primary stress signals and subsequently amplified by secondary signaling molecules. While salt stress is characterized by ionic imbalance, drought stress is mainly associated with osmotic stress[13,34,49]. Secondary stress signals can include small molecules (glutamate, proline, and GABA), ion fluxes (Ca2+), ROS, reactive nitrogen species, and phytohormones (e.g., ABA, indoleacetic acid [IAA], cytokinin [CK], salicylic acid [SA], gibberellic acid [GA], ethylene [ET], jasmonic acid [JA]), depending on the species, genotype and stress type[28,34,50,51]. Since directly elucidating the involvement of secondary messengers in stress responses remains challenging, their roles are frequently investigated by exogenously applying specific substances. For instance, exogenous application of Ca2+, H2O2[10], and GABA[7,52] was tightly associated with drought tolerance in creeping bentgrass. Moreover, spermidine pretreatment in creeping pentgrass improved drought tolerance[53], and exogenous application of Ca2+, spermidine (in zoysiagrass [Zoysia japonica Steud.] and creeping bentgrass)[50,54], and GABA (in creeping bentgrass)[8,55] has been linked to improved ion homeostasis in response to salt stress. Interestingly, the dual application of SA and Ca2+ improved both drought and salt responses in bermudagrass, a C4 grass[49]. These studies suggest that signaling molecules can overlap between salt and drought stress.
-
Calcium signaling is integrated into the ABA-MAPK pathway, functioning as both a secondary messenger and a regulatory node in stress signal transduction under drought and salt stress across different turfgrasses[29]. In response to external signals such as salt and osmotic stress, the cytoplasmic calcium content temporarily increases[28]. Molecules such as calmodulin (CaM), calcineurin-B-like proteins (CBLs), and CDPKs detect calcium changes and transmit them to downstream pathways by conformational changes in CaM and CBLs, relieving autoinhibition and activating CDPKs upon Ca2+ binding[56], thereby affecting the ABA-MAPK regulatory hub[57]. Improved salt tolerance through the salt overly sensitive (SOS) signal transduction pathways, which are responsible for exporting excessive Na+ out of cells, was observed upon exogenous application of Ca2+ (Fig. 1)[1,58]. Accordingly, stress-induced cytosolic Ca2+ signal induces the salt overly sensitive (SOS) signaling pathway in switchgrass[56], and bentgrass[29] under salt stress. Also, in bermudagrass, plasma membrane Ca2+-ATPase transporters (PMCA) were induced 1 h after salt stress, emphasizing the key role of Ca2+ signaling in the early stages of salt stress[21]. Moreover, salt stress induced the expression of CBL-interacting protein kinases (CIPK6, CIPK23, CIPK29) involved in K+ transport in halophyte grass Aeluropus lagopoides[59]. These studies highlight the contribution of Ca2+ signaling to Na+ exclusion and compartmentalization, thereby diminishing salt toxicity in grass cells under salt stress. Another ABA-MAPK signal transduction-related pathway, comprising CDPK26 and mitogen-activated protein kinase 1 (MAPK1), was tightly associated with drought response mechanisms in creeping bentgrass[10] and bermudagrass[11] (Fig. 1), suggesting critical roles in these mechanisms. In rough bentgrass (Agrostis scabra 'NTAS'), an AsCDPK and ion homeostasis-related proteins, including AsSOS1, AsNHX1, and AsNHX2, were significantly induced by salt stress[29] (Table 1, Fig. 1), which are the downstream regulations of Ca2+ signaling[10,29,50].
Table 1. Differentially expressed transcription factors under salt stress.
Turfgrass species Accession/unigenes number Responsive gene/signal compound Downstream targets Related function/description Up/down regulation Tissue Study Ref. Festuca arundinacea (Schreb.) CL19340.Contig2_All NAC021 AIR3 (Auxin-induced
in root cultures)/DBPRegulating auxin-responsive genes/enhancing root architecture under salinity Up Leaf RNA-seq [15] CL914.Contig6_All ERF1 (Ethylene response factor 1) GSTU5 (Glutathione
S-transferase Tau 5),
PRX34 (peroxidase 34)Regulating ROS signaling CL5384.Contig1_All WRKY20 ABI5 (ABA insensitive 5),
NHX1 (Na+/H+ antiporter 1)Regulating ABA signaling CL12389.Contig1_All WRKY46 P5CS1 (Δ1-pyrroline-5-carboxylate synthetase 1),
ABI5 repressionOsmotic stress response, lateral root development Unigene16967_All NAC67 − Salt tolerance is maintained by maintaining cell membrane stability and chlorophyll content − MYB38 DFR (Dihydroflavonol
4-reductase), ANS (anthocyanidin synthase)Regulating axillary meristem formation qPCR Lolium arundinaceum/Festuca arundinacea − FabZIP69 − Stress response transcription factor Up Leaf qPCR [1] Cynodon dactylon (L.) Pers. − C3H − TF regulates stress response through hormone regulation Up Root RNA-seq [21] − ABFs RD29B, RAB18 (osmoprotectants),
HKT1 (Na+ exclusion)ABA-responsive TFs Agropyron elongatum L. MK203870.1 SnRK2.4 PIP2;1, AREB1, bZIP17 Salt stress response by regulating aquaporins and root hydraulics, ABA signaling Up Leaf and root qPCR [34] KC625489.1 NAC9 LRX1, PIN3, NHX3 Salt stress response by induction of lateral root development, ion sequestration The SOS signaling pathway is comprised of a downstream sensor, kinase, and ion homeostasis channel, including SOS3/SOS3-like calcium-binding protein 8 (SCaBP8), SOS2 protein kinase, and SOS1 (Na+/H+ antiporter), respectively[56]. The crosstalk between the ABA and SOS pathways involves several key interacting proteins and phosphorylation events, as revealed by studies in the model plant Arabidopsis. For instance, both ABA and SOS pathways converge on regulating ion transporters, where protein phosphatase 2C (PP2C) ABA-insensitive 2 (ABI2) physically interacts with SOS2 kinase and inhibits SOS1 phosphorylation and activation[60]. Additionally, evidence suggests potential shared phosphorylation targets or regulatory interactions between SnRK2 and SOS2 kinases. For example, recent findings indicate that SOS2 can regulate the transcription factors PLETHORA1/2 (PLT1/2) post-translationally to maintain root meristems under salt stress, impacting root growth[61]. However, the exact mechanisms in grasses require further investigation. Transcriptome analyses of bermudagrass roots under salt stress have shown the simultaneous upregulation of genes involved in ABA biosynthesis and signal transduction, such as 9-cis-epoxycarotenoid dioxygenase (NCED), PP2C, and ABA-responsive element (ABRE)-binding factors (ABFs), together with key components of the SOS pathway, including SOS3/CBLs[21]. These concurrent expression patterns suggest a potential interaction or coordination between ABA-related signaling and the SOS module in grasses under salinity stress, although direct mechanistic evidence is still unclear.
GABA is an amino acid that can impact ABA-MAPK signal transduction and gene regulation by modulating Ca2+-dependent signaling pathways in oxidative stress induced by either salt or drought stress[27,37]. Exogenous application of GABA enhanced drought tolerance through osmoprotectants biosynthesis (e.g., free proline and water-soluble compounds [WSCs]) to facilitate osmotic adjustment, H2O2, and MDA level reduction to enhance cell membrane stability and redox equilibrium in creeping bentgrass[7]. GABA also significantly improved the salt tolerance of creeping bentgrass under prolonged salt stress by inducing salt-tolerance genes linked to critical metabolic pathways, including saccharide, amino acid, and lipid metabolism[55]. Investigating salt tolerance mechanisms in creeping bentgrass indicated that exogenous spermidine improved salt tolerance by increasing intermediate metabolites of the GABA shunt and TCA cycle, which play crucial roles in energy metabolism, signal transduction, and antioxidant defense[54]. Therefore, modulation of GABA biosynthesis and its application in grass species demonstrate significant potential to enhance stress tolerance, particularly under drought and salt conditions.
Furthermore, water deficit and salinity stress increase ROS, such as singlet oxygen (1O2), H2O2, O2·−, and the hydroxyl radical (HO·), which are toxic byproducts of the electron transport chain but also function as stress signal transduction molecules[34,62,63]. Drought stress in Kentucky bluegrass (Poa pratensis L.) (cv. Midnight II) triggers ROS accumulation (O2·− in leaves and H2O2 in leaves and roots), which subsequently increases antioxidant enzyme activities in the leaf samples (including ascorbate peroxidase [APX], monodehydroascorbate reductase [MDHAR], dehydroascorbate reductase [DHAR]), and root samples (including catalase [CAT], glutathione reductase [GR], and MDHAR). It changes their gene expression patterns[64]. In this study, MDHAR was upregulated in both leaves and roots during drought and remained elevated after recovery, suggesting its sustained involvement in antioxidant defense in Kentucky bluegrass[64]. Also, under salt stress, the expression of antioxidant enzymes, including CAT, SOD, POD-1, and POD-2 in bermudagrass was up-regulated[65], indicating an active molecular response to salinity stress in this grass. The same pattern was observed in perennial ryegrass, which showed increased oxidative damage under drought stress, as indicated by elevated H2O2 and lipid peroxidation. Meanwhile, heat shock proteins (HSP70, HSP90-6, and HSP26.2) and several antioxidant genes (e.g., APX, GR, CAT, Cu/ZnSOD, Trx, 2-Cys Prx) were upregulated[66] (Supplementary Table S1). In a comparative study on tall wheatgrass (Agropyron elongatum), the salt tolerance ecotype significantly contributed to a decrease in oxidative stress markers such as O2−, H2O2, and MDA, along with higher expression of NHX1, NHX2, HKT1.4, and sucrose non-fermenting 1-related protein kinase 2 (SnRK2.4)[34]. These studies suggest that ROS signaling is involved in both salt and drought stress and in coordinated regulation with HSPs, forming an integrated defense mechanism contributing to drought tolerance.
-
Phytohormones are ubiquitous compounds that can reprogram the transcriptome and, therefore, metabolism to mitigate stress-induced damage[33]. Salt and drought stresses mainly cause a cascade of responses in grasses through phytohormone pathways, including ABA, CK, ET, and JA[28,31,67]. The integration of CK biosynthesis and ABA signaling plays a crucial role in improving drought and salt tolerance[31]. ABA and CK pathways share intermediate proteins (PP2C, SnR kinases, and transcription factors) and therefore exhibit cross-talk in stress responses[32,68,69]. Salt stress can hinder plant root development by reducing root length and branching[51], while drought stress prompts root depth[70]. However, halophytes and salt-tolerant grass species like bermudagrass can stimulate root growth in response to salt[51,71], suggesting that the stress response is species-specific. The root/shoot ratio is primarily controlled by xylem ABA content, which maintains the functional equilibrium between below-ground and above-ground organs and facilitates water and nutrient absorption by regulating other pathways[25,70]. It is also worth noting that a high CK content, together with enriched ABA transporter pathways, was associated with vigorous multi-rhizome growth in Kentucky bluegrass[67].
ABA is an endogenous developmental signal that is produced in varying amounts under different environmental stresses[31,72], and indirectly initiates stress adaptation processes in flowering plants (angiosperms)[73]. Inside the cell, ABA binds to PYR/PYL receptors and activates SnRK2 protein kinase and alters the expression of genes that regulate stress response (e.g., root growth and architecture)[13,41] (Fig. 1). An ABA-independent mechanism activates SnRK2.4 and SnRK2.10 in response to salt stress in switchgrass, while upregulation of ABA-responsive SnRK2.6 and SnRK2.8 was also detected[35]. In perennial ryegrass (Lolium perenne L.), most of the LpSnRKs (SAPKs) were responsive to salt and drought stress and hormonal (ABA, cytokinin, and ethylene) treatments[74]. Exceptionally, the LpSAPK9 (SnRK2.x) that was identified as a key gene in drought tolerance was downregulated by either ABA, cytokinin (6-BA), or ethephon (ET) treatments[74]. Therefore, SnRK2 (SAPKs) are regulated by hormonal signaling pathways and alter the downstream genes in response to drought and salt stresses[25,35,74].
In the ABA signaling pathway, several well-known protein regulators play central roles, among which phosphatases (most PP2C members) function as negative regulators, whereas kinases (SnRK2s) act as positive regulators[75]. In general, multiple protein kinase families regulate gene expression in grasses under drought[10,25,32,76,77] and salt stresses[15,21,29,33−35]. Among these, the plant-specific SNF1-related protein kinase family SnRK (Sucrose non-fermenting 1-related protein kinase) is known as a key regulator of osmotic stress responses[41]. As illustrated in Fig. 1, drought and salt stress-induced ABA buildup[13,31] leads to the activation of SnRKs (SnRK2.6 and SnRK2.8) via the PYR/PYLs-PP2Cs regulatory module[32,42]. A comparable pattern was observed under salt stress, where SnRK2 genes were upregulated. In contrast, PYR/PYL genes were downregulated, thereby activating downstream transcription factors in smooth bromegrass (Bromus inermis L.)[43]. The upregulation of SnRK2 has been observed in other turfgrasses under drought stress (e.g., in short rhizome and rhizome node samples of tall fescue[32], and in leaf samples of red fescue[16]) and to salt stress (e.g., in leaf samples of Phalaris arundinacea L.[33], and in mixed leaf and root samples of tall wheatgrass)[34]. These findings collectively highlight the consistent upregulation of SnRK2 genes across various grass species in response to drought and salinity stress, suggesting a pivotal role in mediating adaptive responses to these stresses. Moreover, an increased PP2C : PYR/PYL expression ratio was observed alongside elevated RCAR3 transcript levels under salt stress, suggesting complex transcriptional regulation of ABA signaling components rather than direct receptor activation[42].
Focusing on the transcript levels of PP2C in various grass species highlights its critical role in regulating salt and drought stress response (Fig. 1). Molecular responses of P. arundinacea, a forage grass with rapid growth and high biological yield under salt stress, identified a multitude of DEGs associated with hormone signaling, including PP2C family gene[33]. Moreover, upregulation of PP2C has been reported in the belowground organs of tall fescue[32], and in the leaves of red fescue[16] under drought stress, as well as in the roots of bermudagrass[21], and in leaves of P. arundinacea in response to salt stress[33], underscoring the crucial role of PP2C as a key components of ABA signaling that mediate responses to both drought and salinity stresses.
In addition, Aux and CK have antagonistically and/or synergistically regulatory roles on the ABA-MAPK hub in the induction of drought and salt responses. CK signaling networks in grasses involve crosstalk with MAPK pathways through ROS signaling[26,78], and auxin-responsive genes often exhibit antagonistic linkage with ABA through MAPK-mediated[38]. Elevated antioxidant activity (particularly CAT and superoxide dismutase [SOD]) and reduced oxidative damage in ipt over-expressed perennial ryegrass was observed, associated with enhanced CK levels, water content, and photosynthetic rates[78]. In creeping bentgrass overexpressing the ipt gene under the control of the SAG12 promoter, ROS accumulation was reduced while the activity of SOD, CAT, APX, and DHAR increased under drought conditions[79]. Overexpression of the ipt gene that increases endogenous CK level has improved stress tolerance in various turfgrasses, including tall fescue[80], creeping bentgrass[81], and perennial ryegrass[78,79]. These ipt transgenics enhanced drought tolerance by improving osmotic adjustment, water-use efficiency, sustained photosynthetic rates, and root viability[79,81]. Moreover, overexpression of MtIPT in creeping bentgrass improved drought tolerance through altering the transcription of auxin signaling-related genes (including ARFs, IAAs, SAURs, AUX1, ILR1-like proteins, and MAPs), as well as cytokinin signal transduction genes (CRE1, B-ARR)[81]. Salt stress induced the expressions of TAA/YUCCA and CYP735A involved in the biosynthesis of Aux (indole-3-acetic acid) and CK (trans-zeatin riboside, DHZR) in the root of salt-tolerant bermudagrass. It was associated with improved root growth and root/shoot ratio. At the same time, the corresponding phytohormones were reduced in the root of the salt-sensitive line[31]. Similarly, increasing the transcript levels of Aux/IAA family protein Z. macrostachya under salt stress might be the reason for high endurance to salt stress[42]. Additionally, in ipt-overexpressed creeping bentgrass, TFs such as bHLH148, MYB4/4-like, and WRKY28/53/71, which contribute to enhanced RNA degradation and protein modification through the MAPKKK pathway and JA signaling, were upregulated. At the same time, ABA-mediated responses were suppressed[26].
In addition to CK and AUX, other phytohormones (e.g., ET, JA, SA, melatonin) are also involved in salt and drought stress responses. Under salt stress, P. arundinacea, a fast-growing forage grass with high biological yield, showed significantly different gene expression associated with JA signaling-related genes (e.g., jasmonic acid–amino synthetase), ET (ethylene-insensitive protein family gene EIN2 and EIN), and IAA (auxin-responsive protein gene IAA)[33]. Plant hormones and stress signals may modulate stress response signaling in a species-dependent manner in grasses. Under combined drought and salt stress conditions, exogenous SA and Ca2+ enhanced antioxidant defense activities (SOD, POX, APX, and GR), in the photosynthetic tissues of bermudagrass and subsequently enhanced recovery capacity by prevent increasing oxidative stress, however in perennial ryegrass SA and Ca2+ application during salt and drought stress was associated with elevated H2O2 and TBARS (assay-derived indicators reflecting lipid peroxidation) levels[49], emphasizing the modulation of stress response through cross-talk of Ca2+ and ROS signaling. Also, exogenous melatonin improved drought tolerance in tall fescue and salt stress in Festuca elata[82,83], likely through the ROS pathway. Recent studies on phytohormones suggest that the interplay of various hormones, including ABA, CK, AUX, SA, and melatonin, plays a crucial role in mediating drought and salinity stress responses of grasses.
Tall fescue can tolerate long-term and severe drought stress and has been widely used to study drought tolerance mechanisms[1,19,72,76,77,82]. Transcriptomic analysis of tall fescue belowground tissue (the short rhizome or b-type rhizome) samples under drought stress revealed that phytohormone regulation induced rhizome inhibition; however, rhizome initiation was observed after recovery. In this study, the expression of 79 genes involved in the ABA signaling pathway was upregulated by drought stress, along with increased ABA levels. In contrast, genes in the signaling pathways of other phytohormones, such as indole-3-acetic acid (IAA) (29 genes), CK (seven genes), and gibberellin A4 (GA4) (11 genes), were downregulated, leading to a reduction in auxin and gibberellin levels[32]. Similarly, in another study, the expression level of ABA synthase-related genes was higher in tall fescue leaf, pseudostem, root, and crown tissues under water stress (Supplementary Table S2)[76]. Also, in other turfgrasses (e.g., Kentucky bluegrass and bermudagrass), high ABA levels under drought and salt stress have been linked to enhanced stress tolerance[31,84,85]. Briefly, in one study, in the root sample of bermudagrass, the moderate accumulation of ABA content in the salt-tolerant genotype (C43) was associated with higher root activity, while higher ABA content in the salt-sensitive genotype (C198) led to lower root growth under high NaCl conditions[31]. Similarly, in another study, in bermudagrass, a lower ABA content in the drought-tolerant cultivar (Guanzhong), associated with downregulation of PP2C and therefore higher activity of SnRK2, was reported, while higher ABA content in the drought-sensitive cultivar (Yangjiang) was observed in response to drought stress[85]. Therefore, enhancing ABA levels to some extent could improve drought and salt tolerance by promoting root growth.
-
The mitogen-activated protein kinase (MAPK) pathway is one of the most important signal transduction pathways in plants, orchestrating environmental signals[14]. MAPK modules consist of at least three functionally linked serine/threonine kinases that are hierarchically phosphorylated[39]. In this cascade, first MAPK Kinase Kinases (MAPKKKs) are activated by various stress signals (including osmotic stress, cytosolic Ca2+, and ABA) and subsequently phosphorylate MAPK Kinases (MAPKKs), which in turn activate MAP Kinases (MAPKs)[10−13]. Activated MAPKs then phosphorylate a range of downstream target genes, including transcription factors, enzymes, and other signaling proteins, thereby mediating stress-responsive signaling pathways[11,14,39] (Fig. 1). Transcriptomic and metabolomic analyses of two bermudagrass genotypes (drought-sensitive and tolerant) under drought stress revealed that both the plant hormone signal transduction and the MAPK signaling pathway were prominently enriched in the two bermudagrass genotypes[11]. This highlights the pivotal roles of hormone signal transduction and the MAPK signaling pathway in the drought response mechanisms in both genotypes of bermudagrass. Although genotype-specific responses, including changes in hormone signaling, metabolism, and stress-related metabolites like ABA, can be seen.
In recent studies, MAPKKKs were identified as integrated regulators in signal transduction in response to both drought and salt stresses. MAPK signaling in Kentucky bluegrass[13], sheepgrass (Leymus chinensis)[12], creeping bentgrass[10], and tall fescue[77] contributes to drought-induced osmotic adjustment. To explain briefly, MAPKKK18-18 and MAPKK3 regulated drought stress by the ABA signaling pathway in red fescue[16]. In the drought-tolerant forage species sheepgrass, all MAPK-related genes (e.g., MAP2K1, MAPK1-2, and MAPK1-3) were upregulated, along with Ca2+ signaling, and plant hormone transduction pathways being enriched under drought stress[12]. In creeping bentgrass, MAPK1 and CDPK26, as essential protein kinases, were involved in signal transduction pathways in drought response mechanisms[10]. Moreover, transcriptome analysis of the pool of leaf, shoot, root, and inflorescence parts of tall fescue also emphasized the upregulation of protein kinases (MAPK/ CDK/ AKT) under drought stress[77] (Table 2).
Table 2. Differentially expressed transcription factors under drought stress.
Turfgrass species Accession/unigene number Responsive gene/signal compound Downstream target genes Related function Up/down regulation Tissue Study Ref. Festuca arundinacea (Schreb.) CL21940.Contig3_All ABF LEA (Late embryogenesis abundant) genes (e.g., dehydrins) – protect cellular structures. RD29B (responsive to desiccation) – osmotic adjustment. RAB18 (ABA-responsive) – dehydration tolerance. ABA-responsive element binding factors Up Rhizome/rhizome node RNA-seq [32] CL14001.Contig6_All ARF GH3 (auxin-responsive genes) – balances auxin homeostasis. SAUR (small auxin-up RNA) genes – control cell expansion. PIN (auxin efflux carriers) – adjusts root architecture for water uptake. IAA/auxin response factors Festuca arundinacea (Schreb.) − AP2/EREBP RD29A – osmoprotectant synthesis. COR15A (cold-regulated) – stabilizes membranes. LEA14 – protects macromolecules. Abiotic stress-responsive TFs Up Leaf, pseudostem, root, crown RNA-seq [76] − HSFs HSP70, HSP90, sHSPs Heat shock TFs − NAC TFs ERD1, SAG12, VIN2 Regulation of physiological process-related genes/senescence/stress-resilience genes − bZIP TFs ADH1, P5CS Regulation of physiological process-related genes/ABRE-containing genes − WRKY TFs PR1, GST, POD Regulating stress-responsive genes − 3helix TFs ELIPs, PSBS Triple helix transcription factors − bHLH TFs CHS, FERRITIN Regulation of physiological process-related genes − GARP RBCS, CAB G2-like transcription factor/photosynthesis-related genes Axonopus compressus L. − NAC TFs SAG113, ERD1, P5CS Regulation of physiological process-related genes Up Leaf RNA-seq [25] − ABI5 RD29B, LEA14, GST ABA-dependent transcription factor − MYB3 PAL, C4H, CER1 Stress-responsive TFs − WRKY1 PR1, APX2, HKT1 Abiotic stress-responsive TFs Agrostis stolonifera GR279830.1 MYB13 PAL/C4H/CHS Abiotic stress-responsive TFs qPCR [10] DV867719.1 WRKY75 APX2/HKT1/CAT1 Abiotic stress-responsive TFs In response to salt stress, MAPK-related genes were essential regulators of stress-related genes in switchgrass[35,86], bermudagrass[21], Lolium temulentum (a model grass species)[39], and seashore paspalum (Paspalum vaginatum)[40]. In brief, the expression of stress-responsive proteins like CAT1, SOD1, GSH1, APX1, and P5CS2 is promoted in switchgrass under prolonged salt stress by activated MAPKs and SnRK2 signals sent to their TFs[35]. Induction of MAPK2 1 h after salt stress in bermudagrass,[21] and also rapid activation of the 46 kDa MAPK (p46) after exposure to salt and other abiotic stresses in L. temulentum[39], suggesting the function of MAPK in early stress responses to salt stress (Table 1). In seashore paspalum, PvWRKY52 serves as a central hub gene in response to salt stress, predicted to interact with key elements of the MAPK pathway components such as MKS1, MPK3, and MPK4[40].
-
Transcription factors (TFs) play a pivotal role in regulating stress-responsive genes as important downstream components of the ABA-MAPK signaling network. Recent studies have shed light on the intricate networks of TF families in turfgrass, including bZIP, NAC, WRKY, MYB, and ubiquitin-related mechanisms, linked with MAPK and/or ABA signaling, which enhance plant adaptation to salt and drought stresses[10,17,21,25,59,62,76,87]. For instance, bZIP family proteins in grass respond to drought and salt stress[1,12,62,76] by functioning as transcriptional regulators that integrate upstream regulatory inputs and control the expression of downstream transcription factor families, including MYB, zinc finger proteins, NAC, and AP2/ERF members[62,88]. In a mixed sample comprising leaf-like stems and the crown tissues of tall fescue under drought stress, the bZIP TFs were upregulated[76]. Similarly, the bZIP69 transcription factor was upregulated in the leaf tissues of tall fescue under salinity stress[1]. Also, the transcription factor bZIP (homolog of Ghi016560) demonstrated high expression in the shoots of the halophyte grass Sporobolus virginicus under salinity; conversely, its expression was reduced in the root samples[62]. These studies emphasize the common yet tissue-specific regulation of bZIP transcription factors in response to salt and drought stress (Tables 1, 2).
ABFs, members of the bZIP TF family, bind ABREs and participate in ABA signaling pathways and ABA negative feedback regulation, playing critical roles in salt and drought stress responses in various grass tissues[24,30]. Under salinity stress, ABF expression is significantly upregulated in the roots of common bermudagrass (C. dactylon Pers.)[21], highlighting its importance in ABA-mediated signaling for root adaptation to high-salinity environments. Furthermore, ABF expression levels are elevated in the belowground organ of tall fescue during drought conditions[32], suggesting their pivotal role in maintaining growth and resilience under limited water availability and ionic stress. Also, under drought stress, increased ABF expression has been observed in the leaves of bentgrass[10], indicating its involvement in regulating leaf-specific responses to water deficit as well. These findings underscore the tissue-specific regulation of ABFs in response to abiotic stress and their critical function in orchestrating adaptive mechanisms in different grass species.
Moreover, NAC transcription factors, a large plant-specific gene family modulating versatile biological processes, are involved in salt and drought stress response in grasses (e.g., orchardgrass [Dactylis glomerata L.], perennial ryegrass, tall wheatgrass, sheepgrass, switchgrass)[12,34,63,89,90]. Notably, the expression of NAC genes is altered in coordination with ABA- and/or MAPK-related genes and signaling pathways[12,34,63,90]. LpNAC22 alleviated drought stress symptoms by directly binding to the promoter of LEA family genes in perennial ryegrass[89]. In tall wheatgrass, the salt tolerance feature was associated with the NAC9 transcription factor, along with a decrease in ROS[34]. Moreover, PvNAC1 (switchgrass NAC transcription factor 1) was induced by multiple stress factors, including salt, dehydration, H2O2, and ABA in switchgrass[63]. It is worth noting that overexpression of PvNAC1 significantly improved plant growth and salt tolerance, as evidenced by higher scavenging of ROS, reduced Na+ accumulation, and increased K+ levels in both roots and shoots of switchgrass[63]. Collectively, these studies indicated the important function of NAC transcription factors in dehydration and salt stress adaptation in grasses.
Several WRKY transcription factors have been linked with the ABA-MAPK regulatory core in various grass species in response to salt and drought stress[10,17,18,76,87]. In addition to activation of MAPK and WRKY, other TFs belonging to AP2/ERF, bHLH, and MYB families were identified as potential key regulators of the drought response in sheepgrass[12]. Moreover, multiple WRKY transcription factors and ABA biosynthesis and signaling were upregulated in all tissue types, comprising leaf, pseudostem, and crown tissues of tall fescue under drought stress[76]. Similarly, under drought conditions, the expression of WRKY75 and WRKY1 was induced, respectively, in leaf tissues of creeping bentgrass and carpet-grass with ABA-MAPK regulation[10,25]. In creeping bentgrass, exogenous Ca2+ application induced the expression of WRKY75 and MAPK1, along with increased expression of drought-responsive genes encoding antioxidant enzymes such as SOD, MDHAR, and glutathione reductase (GR)[10]. While WRKY1 was co-expressed with both MAPK1 and NCED4 (9-cis-epoxycarotenoid dioxygenase), an ABA biosynthesis enzyme in carpetgrass[25]. HvWRKY38 has also been implicated in abiotic stress signaling, as overexpression of this TF in transgenic bahiagrass (Paspalum notatum Flügge) improved survival, biomass accumulation, water retention, root regeneration, and photosynthetic efficiency under severe dehydration stress[87], which might be associated with ABA signaling. Under salinity stress, WRKY20 and WRKY46 were upregulated in leaves of tall fescue[15], plus in seashore paspalum, WRKY52 was significantly expressed in root and interacting with MKS1, MPK3/MPK4 in MAPK signaling[40]. In the halophyte A. lagopoides, a C4 perennial grass native to Iran, the expression of WRKY33 was induced in response to salt stress, accompanied by enrichment of the MAPK signaling pathway and thereby modulating the downstream functional genes to maintain osmotic and ionic balance[59]. In bermudagrass, WRKY2 negatively regulates lateral root growth under salt stress through AtMAPK3 function that ultimately regulates ABA signaling and auxin homeostasis[38]. In switchgrass, certain WRKY genes show no significant change under salt stress, while under drought, they are strongly regulated at early timepoints after stress exposure[5]. These studies indicate that the WRKY transcription factor family is mediating responses to both drought and salinity stresses across diverse grass species through the ABA-MAPK regulatory hub (Tables 1, 2). However, WRKY expression can be selectively and tissue-specifically induced under drought or salinity conditions.
Ubiquitin-mediated protein degradation or ubiquitination is a critical post-translational regulatory mechanism in plant stress responses. Different types of ubiquitin ligase enzymes (E3 ligases), which determine the fate of target proteins in signaling pathways, are facilitating the ubiquitination process[91]. The RING-type E3 ligases (DRIP1 and DRIP2) regulate the degradation of the DREB2A TF under non-stress conditions[92]. Additionally, F-box proteins (a subclass of SCF E3 ligases) modulate substrate specificity in plant development and stress responses[93]. The expression levels of multiple RING-type E3 and E3 SCF ligases were induced under drought stress in bermudagrass[94]. In zoysiagrass, members of the plant U-box (PUB) family of ubiquitin ligases were induced in roots under salt stress, and the ABRE motif was the most abundant cis-element in the promoter regions of ZjPUB genes, indicating a broad involvement of ABA in the transcriptional regulation of ZjPUBs[95]. These studies suggest the contribution of the 26S proteasome pathway in removing damaged proteins during stress conditions through the ABA signaling pathway.
The initial stress response in plants is coordinated by a rapid shift in certain TF activity through the ABA-MAPK signaling pathway. For instance, following just 1 h of salt stress, early-responsive TFs (e.g., C3H, ARF, NAC, and Trihelix) were strongly induced, accompanied by consistent and continuous up-regulation of genes involved in ABA biosynthesis and signaling (e.g., NCED, PP2C, ABFs) in bermudagrass[21]. Suggesting the established role of ABA signaling in regulating first-line transcriptional responses that activate downstream defense genes in bermudagrass[21]. Beyond the immediate shock, the SPL (SQUAMOSA promoter-binding protein-like) family is central to managing abiotic stresses by growth regulation. In centipedegrass (Eremochloa ophiuroides [Munro] Hack.), 19 EoSPL genes displayed distinct activity in leaves and responded differentially to salt and drought stresses, and all EoSPL genes harbored ABRE elements linked to ABA-responsive regulation, except for EoSPL10[96].
A study on switchgrass identified multiple LBD TFs harboring ABRE and MBS cis-elements implicated in ABA-mediated stress responses. Among these, overexpression of PvLBD12 enhanced salt stress tolerance in transgenic plants, highlighting its potential for genetic improvement of stress tolerance[97]. Similarly, the introduction of the AtDREB1A gene into Kentucky bluegrass significantly boosted drought tolerance. This enhancement was linked to higher ABA and IAA content, water-soluble carbohydrates, proline, and proteins[98].
-
In response to abiotic stress, long non-coding RNAs (lncRNAs), microRNAs (miRNAs), and small interfering RNAs (siRNAs) function as a sophisticated regulatory network of non-coding RNAs that fine-tune gene expression, enabling plants to rapidly adapt by adjusting their physiology and activating protective mechanisms[4,99]. Transcriptional response of switchgrass to primary and secondary dehydration stresses highlighted the pivotal role of lncRNAs in stress adaptation and memory. The study identified numerous novel lncRNAs involved in ABA and ET biosynthesis, signaling pathways, as well as starch and sucrose metabolism. A distinct pattern of lncRNAs upregulation was observed in the upstream of ABA biosynthesis, and ABFs mirrored the induction of their cognate targets during repeated dehydration[99]. Conversely, the lncRNAs located upstream of ET biosynthesis genes and ethylene-responsive transcription factors (ERFs) that exhibited expression patterns similar to their associated genes were predominantly downregulated under drought conditions[99].
In transgenic creeping bentgrass expressing elevated levels of the rice miR319 gene (Osa-miR319a), significant morphological and physiological modifications were observed through the regulation of TEOSINTE BRANCHED 1/CYCLOIDEA/PCF (TCP), which are plant-specific TFs, and AsNAC60[4]. These included reduced tiller numbers, broader and thicker leaves, larger stems, an increased weight-to-area ratio, and enhanced wax coverage, which improved drought and salt tolerance, likely through reduced sodium uptake and enhanced structural resilience[4]. Given the regulatory role of TCP TFs in drought stress via the ABA-related stress gene ABA1[100], and the rapid induction of OsNAC3 expression by ABA and salt stress[101], it can be inferred that TCP and NAC TFs may control salt and drought stress-responsive genes in grasses through ABA signaling. However, this hypothesis requires experimental validation in grasses.
Additionally, a recent study on salt-tolerant switchgrass demonstrated the role of N6-methyladenosine (m6A) modification, a dynamic and reversible modification of eukaryotic mRNA, in regulating salt-responsive genes and pathways. A reduction in m6A modifications positively regulated salt-stress-related genes, including MPK6, CAT1, and Glutathione synthetase 1 (GSH1), as well as osmoregulatory pathway genes such as P5CR and delta 1-pyrroline-5-carboxylate synthase 2 (P5CS2). Interestingly, the expression of key salt resistance pathway genes (SOS1 and SCaBP8/CBL10, associated with Ca2+ signaling) increased independently of m6A modifications[56]. These findings position m6A modification as an important post-transcriptional regulator of salt stress responses in switchgrass, partly through modulation of MAPK-related signaling components such as MPK6.
Similarly, in the halophyte cordgrass (Spartina alternifolia Loisel), miRNAs have been reported as critical regulators of plant responses to salinity stress[102]. Under salt stress, Sal-miRNA397a was strongly upregulated, while Sal-miRNA397b exhibited moderate elevation. In addition, novel miRNAs, including Sal-miR740 and Sal-miR528, exhibited tissue- and time-specific expression patterns, indicating potential involvement in salinity adaptation[102]. The observation that stress-responsive miRNAs target both positive and negative stress tolerance regulators reflects the complex control dynamics of miRNA-mediated stress responses. Although the direct involvement of specific signaling pathways was not addressed, such regulatory features are consistent with the multilayered control observed in major stress signaling systems, including ABA and MAPK pathways[13,39,44].
In switchgrass seedlings exposed to severe drought stress, conserved miRNAs displayed dose-dependent changes in expression[103], which may reflect the dynamic nature of stress-responsive signaling. Under high drought stress, miR156, associated with developmental processes and biomass accumulation, was up-regulated, whereas miR162, linked to miRNA biogenesis and stress responses, exhibited a down-regulation[103]. Taken together, these findings underscore the importance of miRNAs such as miR156, miR162, miR319, Sal-miRNA397a, and Sal-miR528 in improving drought and salinity tolerance in turfgrass species[4,102,103].
-
Abiotic stress resistance in plants, including various grass species, is a complex trait regulated by a multitude of genes (Tables 1, 2, Supplementary Tables S1, S2). Targeting transcription factors, which influence the expression of numerous downstream genes, is more practical and effective than manipulating individual functional genes[104]. Therefore, understanding the interaction of stress-responsive regulatory systems, such as signaling pathways, transcription factors, and their associated functional genes, is of critical importance. These regulatory mechanisms significantly influence hormonal signaling and metabolic pathways, including carbon and protein metabolism, which are essential for optimizing internal processes under stress conditions[6,29,54].
The Ca2+, GABA, and ROS signaling are shared pathways modulating drought and salinity stress responses, through ABA and MAPK signaling[5,19,55,64,84]. Those signal molecules further induce regulatory networks, such as transcription factors and small RNAs, which coordinate plants' shared response to salinity and drought stress. Furthermore, understanding the crosstalk between drought- and salt-induced signaling pathways could guide the development of grass cultivars resilient to both stresses, which often occur concurrently in natural environments. A synergistic approach that leverages both osmotic adjustment through improved water retention, reduced ion toxicity in the cytoplasm, and metabolite accumulation associated with drought tolerance, combined with effective vacuolar salt compartmentalization linked to cytoplasmic protection, ion homeostasis, and ultimately salinity tolerance, is essential (Fig. 2). This integrated approach could support growth and metabolic functions under simultaneous stress conditions.
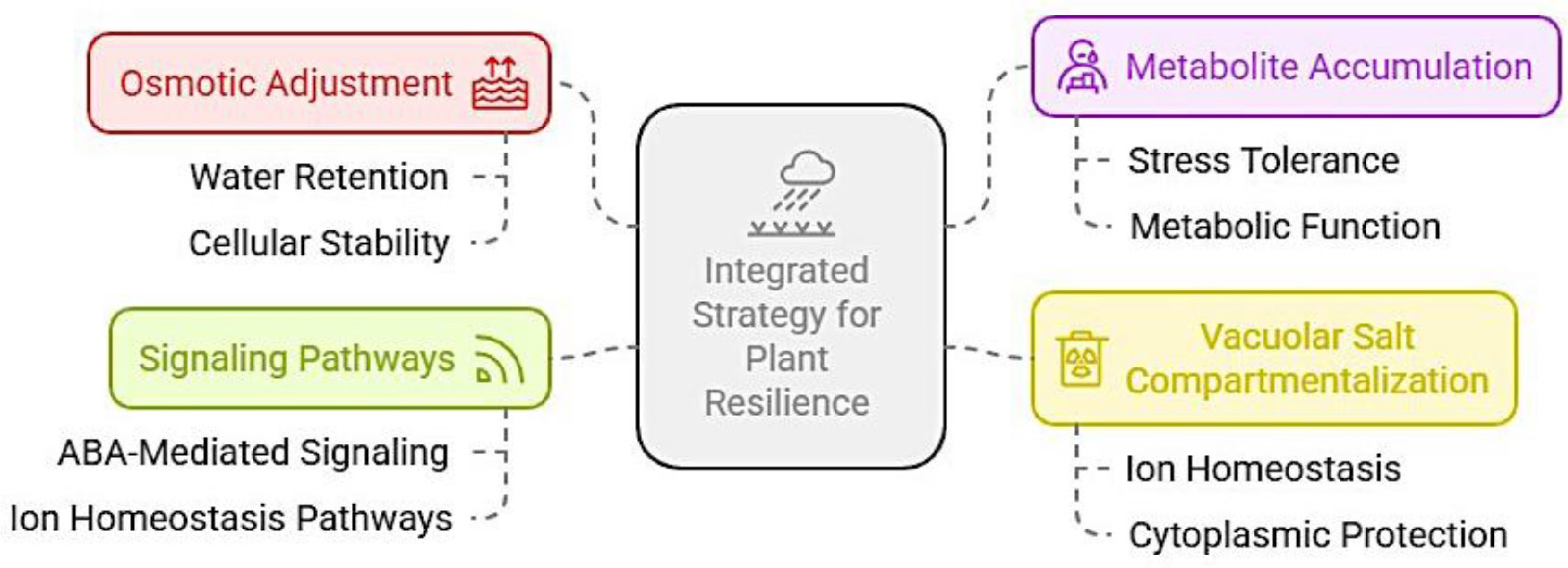
Figure 2.
Integrated strategy for plant tolerance to drought and salinity stresses.
-
This study highlights the complex regulatory components governing grass responses to drought and salinity stress, with a particular focus on ABA and MAPK roles in transcriptional regulation. During which some research gaps are also identified, which may help future studies. For instance, the cross-talk points between the ABA and SOS pathway (SOS2 in Arabidopsis) in grasses are not well characterized. Many other questions remain. For example, how does the crosstalk between ABA, Ca2+, and ROS influence plant tolerance to both drought and salinity? How can the synergistic approach be effectively implemented in agricultural practices to enhance plant resistance? Are there any potential trade-offs or drawbacks in combining the mechanisms for drought and salinity stress responses?
While available transcriptomics studies mainly focused on evaluating single stress responses, there is a noticeable gap in information on molecular responses to a combined set of salt and drought stress. Dual stress transcriptome studies will cast light on the interaction of regulatory frameworks, including upstream signaling pathways and transcriptional interaction networks. Moreover, moving beyond descriptive transcriptomics toward an integrated understanding of the regulatory mechanisms and their cross-talks that drive dual stress adaptation in non-model grass species is necessary.
-
Future studies should aim to fill critical knowledge gaps in our understanding of gene regulatory networks and signaling cross-talk in grasses exposed to simultaneous drought and salinity stress. Special attention should be given to the functional validation of key transcription factors and signaling components using molecular genetic tools or overexpression systems in relevant grass species. Combining transcriptomic data with proteomic, epigenomic, and metabolomic approaches will also be instrumental in unraveling multilayered regulatory networks. Additionally, detailed characterization of ABA and MAPK pathway components in response to drought and salinity stress in common tolerant grass species is needed. Ultimately, such integrative efforts will support molecular breeding strategies for developing multi-stress-tolerant grass cultivars suited for changing climate scenarios.
The authors thank all the colleagues who reviewed and proofread the manuscript. This work was not supported by any funding agency.
-
The authors confirm their contributions to the paper as follows: study conception and design, draft manuscript preparation: Esmailpourmoghadam E, Esmaeili S, Salehi H, Huang B. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that there is no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/grares-0026-0006.
- Supplementary Table S1 Differentially expressed target genes under salt stress.
- Supplementary Table S2 Differentially expressed target genes under drought stress.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Esmailpourmoghadam E, Esmaeili S, Salehi H, Huang B. 2026. ABA and MAPK signaling-mediated transcriptional regulation of drought and salinity responses in grasses. Grass Research 6: e015 doi: 10.48130/grares-0026-0006 

ABA and MAPK signaling-mediated transcriptional regulation of drought and salinity responses in grasses
- Received: 12 December 2025
- Revised: 02 February 2026
- Accepted: 19 March 2026
- Published online: 02 June 2026
Abstract: The adverse effects of drought and salinity stress are associated with various stress-signaling pathways and transcription factors, which can be unique or shared by both salt and drought stresses. This article provides an overview of recent advances in grasses, focusing on hormonal signaling, the mitogen-activated protein kinase (MAPK) cascade, stress signal transduction molecules (calcium, reactive oxygen species [ROS], γ-aminobutyric acid [GABA]), and transcriptional and post-transcriptional regulation under drought and salt stress. Studies suggest that signaling molecules can overlap between salt and drought stress; therefore, studying the subsequent signal transduction pathways is of great importance. Transcriptomic studies highlight stress-specific gene regulation in hormone signaling (e.g., ion transporters are strongly upregulated under salt stress, whereas abscisic acid [ABA]-related genes, the growth-promoting hormones, including indoleacetic acid [IAA] and gibberellic acid [GA], are downregulated under drought). ABA signaling, MAPK cascade, ROS pathway, and calcium signaling are central to both stresses. However, salt stress mostly involves ABA signaling, rapid MAPK activation, and ion homeostasis. Studies highlighted genotype-specific responses and tissue-specific regulation of hormone signal transduction and MAPK signaling. The integration of these pathways underscores the significance of molecular crosstalk in drought and salinity adaptation, offering opportunities for genetic engineering and breeding programs to enhance stress tolerance in grass species.
-
Key words:
- Abscisic acid /
- Drought /
- MAPK cascade /
- Salinity