








-
Apples are a rich source of phenolic, triterpene, fibre, and vitamin compounds that are known for their health-promoting properties[1]. The polyphenolic compounds in apple include flavonols (FLAVO), phenolic acids, flavanols (FLAVA), dihydrochalcones (DHCs), and anthocyanins. These compounds are reported to be linked to health benefits such as reduced risk of cardiovascular disease, certain cancers, and diabetes[2]. Among the polyphenols, DHCs exhibit an excellent blood sugar-lowering effect for the management of diabetes. For example, phloridzin, which is the prominent DHC in apple, exerts anti-diabetic effects by increasing the insulin sensitivity and inhibiting sodium glucose co-transporter-2 (SGLT-2)[3]. Human intervention trials suggest that the co-ingestion of high polyphenol-containing apples (or apple products) with foods having a high glycemic index (GI) can lower the GI of these foods and partially reduce the absorption of some of the ingested sugars and fats[4].
Phenolic compounds play a crucial role in plant adaptation to environmental stresses, contributing to both biotic and abiotic stress tolerance[5,6]. Genome-wide association studies (GWAS) in diverse resources of apple germplasm have identified genomic regions associated with DHCs, FLAVO, and hydroxycinnamic acids that also harboured disease-resistance genes, including CC-NBS-LRR and NB-ARC domain-containing proteins[7]. In addition to phenolics, organic acids such as ascorbic acid (AsA) contribute to plant tolerance against drought and heat stress[5,8]. Enhancing the AsA content in apple is desirable for improving both stress resilience and nutritional quality[9,10]. Genomic regions linked to the glycosylated derivative ascorbic acid 2-beta-glucoside (AA-2βG) also contain drought- and heat-responsive genes, suggesting coordinated genetic regulation of antioxidant metabolism and stress adaptation[7]. By interacting with aroma, taste, and texture, compounds such as terpenes, phenolics, and flavonoids shape sensory attributes such as sweetness (SWEET), bitterness, astringency (ASTR), and aroma, while also providing antioxidant and anti-inflammatory benefits that contribute to the prevention of chronic diseases[11,12].
Historically, apple breeding programmes have focused on the improvement of consumer traits (e.g., appearance, texture, taste, SWEET, and flavour) and production traits (e.g., yield, storability, and disease resistance), with no direct focus on polyphenol content or associated health benefits[13]. Several studies have shown that modern apple cultivars tend to have significantly lower fruit phenolic acid and FLAVA contents than those of wild species and heritage cultivars[14,15]. The phenolic content in cultivated apples was reported[16] to be 68% lower than in their wild progenitors (Malus sieversii). Kumar et al.[7] reported that Malus spp. accessions showed at least 2-fold higher average content of several metabolites (e.g., AsA, chlorogenic acid, phloridzin, and trilobatin) than M. domestica accessions. It has been suggested that selection against internal flesh browning and ASTR could contribute to lower phenolic content in commercial cultivars[17].
Apples have higher fructose content and less favourable fructose–glucose ratios (FGRAT) than some other common fruits such as oranges and bananas. Hence, people suffering from fructose malabsorption are advised against eating the fruit of most modern apple cultivars[18]. Although apple consumption is generally considered beneficial to health, there is growing interest in further enhancing the health benefits of apples. Breeding programmes are now beginning to consider improving apple's phytochemical content while maintaining or even improving fruit eating quality (EQUAL). Introgression of novel traits (e.g., phytochemicals) from primitive germplasm could be achieved by using a PBC breeding strategy. A dual-target (i.e., EQUAL and phytochemicals) breeding strategy requires an understanding of the genetic correlation between EQUAL traits and phytochemicals. The genetic architecture of several phenolic compounds is reported to be complex polygenic[7,19], suggesting that a genomic selection (GS) approach would be ideal. GS uses DNA marker information spread across the entire genome to predict the genetic value of an individual for polygenic traits even before those traits are expressed or measured[20]. Hence, it has the potential to fast-forward the introgression process, as the generation interval in apples is about 7 years.
Although the phytochemical makeup of apple fruit affects important traits such as nutrition, flavour, texture, disease resistance, and postharvest quality, phytochemical-oriented breeding strategies for improving the quality of apple are lacking. The main objectives of this study included estimation of the genetic control (heritability) of phytochemicals and the genetic relationships between phytochemical traits and conventional eating-quality traits, with the long-term goal of developing a breeding strategy for the simultaneous improvement of both trait groups. This study also investigated the potential of GS for fast breeding of new high-health cultivars.
-
Accessions of wild Malus species were used as paternal parents to hybridise with commercial cultivars to raise four families: 'PremA17' × M. trilobata, 'PremA17' × M. ioensis, 'Scilate' × M. trilobata, and 'Scifresh' × M. sieboldii. The number of seedlings representing each cross was 74, 16, 51, and 163, respectively. 'Royal Gala' and 'Braeburn' cultivars were involved in the pedigree of the seed parents, which provided genetic connectedness among the crosses. Two-year-old seedlings were propagated onto 'M9' rootstock and then planted into the orchard (Havelock North, New Zealand) in 2020 at 3.0 m × 0.5 m spacing for fruit evaluation. All trees received standard commercial management for nutrition, pesticides, fruit hand-thinning, and irrigation.
Fruit sampling and sensory evaluation
-
Fruit sampling was conducted over a period of two years (2023–2024), depending on fruit availability. Six fruits were harvested from 191 and 281 of the 304 seedlings during the 2023 and 2024 seasons, respectively, with 180 seedlings being common between the two seasons. We followed the fruit harvesting and storage protocols reported earlier[7]. Briefly, fruits were harvested once from each seedling when judged mature by visual inspection of skin background colour and the starch pattern index (SPI) rating[21]. Fruits were harvested when the SPI index was between 2 and 3 and then stored for 10 weeks at 0.5 °C, followed by a further 1 week at 20 °C before evaluation.
Thin wedges of cortical tissue were cut from two fruits from each seedling for sensory evaluation by a trained assessor. Several sensory traits, including SWEET, sourness (SOUR), ASTR, and overall EQUAL, were assessed on a scale of 0 (= lowest) to 9 (= highest). EQUAL was calculated as the average of the fruit texture (firmness, crispness, juiciness) and SWEET scores. Fruit size (SIZE) was also assessed visually on a scale of 1 (< 30 g) to 8 (> 210 g).
Phytochemical phenotyping
-
Fruit tissues were sampled following a previously published protocol[7]. For each apple accession, 16 tissue cores were taken—four from each of four apples—using a #5 corer (9 mm internal diameter). The cores were collected from four positions evenly spaced around the apple's equator and included both skin and cortex but excluded the core and seeds. If needed, cores were trimmed to approximately 1 cm in length. The total sample (~10 g) was placed into a 250-mL Schott bottle containing 100 mL of extraction solvent (80:20:2 ethanol : Milli-Q water : formic acid) and spiked with 1.05 mg/L of the internal standard naringenin. The mixture was homogenised using an Omni GLH850 homogeniser (Kennesaw, GA, USA), then left to extract overnight at 4 °C. An aliquot of the extract was later transferred to a centrifuge tube, centrifuged, and the supernatant was taken for analysis. For the analysis of polyphenolics, the supernatant was diluted 1:1 with methanol (actual volumes: 200 µL extract + 200 µL methanol) and then analysed by liquid chromatography–high resolution accurate mass–mass spectrometry (LC-HRAM-MS). Analysis of AA-2βG was conducted using ultra-high-performance liquid chromatography (UHPLC). The response was calibrated using AA-2αG (Fluka, USA) authentic standards; therefore, AA-2βG was quantified as AA-2αG equivalents[7]. The concentration of phytochemical compounds was expressed on a fresh weight basis.
For sugar quantification, a subsample of the extract was dried in a centrifugal evaporator and redissolved in 1 mL of ultra-pure water with a type 1 resistivity of 18.2 megaohm-cm (MΩ-cm). Samples were separated and quantified for soluble mono-, di-, and tri-saccharides using a DIONEX ICS5000+ PA20 column with an amino trap. A separation gradient was applied from 9 to 100 mM KOH using an eluent generator and saccharides were detected using an electrochemical detector. Individual saccharides were quantified against HPLC-grade saccharides sourced from Sigma-Aldrich.
Phenotypic data analysis
-
Since the seedlings were evaluated over two consecutive years, a mixed-effects model incorporating permanent environmental effects was applied to each trait[20]:
$ {y} = {Xb} + {Za} + {Zp} + {e} $ (1) where, y is the vector of observations; b is the vector of fixed effects (overall mean, year); a, p, and e are vectors of random additive genetic effects, permanent environment effects, and residual effects, respectively. X is the incidence matrix for the fixed effects and Z is the incidence matrix linking observations to seedlings. The variances associated with the random effects a, p, and e were
$ \sigma _{a}^{2} $ $ \sigma _{p}^{2} $ $ \sigma _{e}^{2} $ $ \sigma _{a}^{2} $ $ \sigma _{a}^{2} $ $ \sigma _{p}^{2} $ $ \sigma _{e}^{2} $ DNA sequencing and variant calling
-
DNA was extracted using a CTAB-based protocol and was subjected to a second round of clean-up using 2 volumes of AMPure XP beads (Beckman Coulter), binding for 5 min at room temperature, and washing twice with 200 μL of 80% ethanol for 30 s; all traces of ethanol were then removed before eluting in 30 μL TE (pH 7.5) buffer. About 100–200 ng of DNA from each sample was randomly fragmented for 9 min according to the protocol from NEBNext®Ultra™ II FS DNA Library Prep (Cat. # E6177L New England Biolabs), but using half reaction volumes. The fragmented DNA had an average size of 300 bp, as analysed by capillary electrophoresis (Fragment Analyzer, HS NGS method 1–6,000 bp). Barcodes were introduced by amplifying the library construct with NEBNext Multiplex Oligos for Illumina (96 Unique Dual Index Primer Pairs, cat. # E6441A, E6443A, E6445A, E6447A). Unused indexed oligos were removed using magnetic beads (AMPure XP at 0.9X). The amplified library fragment distribution had an average size of 450 bp. Equal amounts of each library were pooled based on the amount of DNA quantified by fluorescence (Qubit HS dsDNA kit, ThermoFisher), one pool per plate. To refine the fragment distribution, a second AMPure XP magnetic bead size selection was performed on each pool, at 0.6/0.2X ratio. The samples were sequenced in pair-end 150 bp mode using the NovaSeq platform at Novogene. We used a low-coverage whole-genome resequencing approach for genotyping, generating about 3.6 Gb data per seedling.
The data quality was checked using FastQC v0.11.7 (
www.bioinformatics.babraham.ac.uk/projects/fastqc ) and the data were cleaned with bbduk in the BBMap/38.33 package (https://sourceforge.net/projects/bbmap ). The cleaned reads were mapped to GDDH13 v1.0 reference genome[23] using hisat2/2.2.1 (with '--sensitive --no-spliced-alignment -I 100' options)[24]. The alignment sam files were sorted and indexed using samtools/1.12. Single-nucleotide polymorphisms (SNPs) and INDELs were then called with 'bcftools mpileup' and 'bcftools call' in bcftools/1.12[25]. The resulting compressed VCF file was indexed using tabix/0.2.6. INDEL and SNP features were further separated and filtered with vcftools/0.1.14[25] based on minor allele frequency (MAF) and the maximum MAF (--maf 0.05 --max-maf 0.9). Multiallelic SNPs (sites with more than two alleles) were excluded using PLINK2/v2.00a2lm (--max-alleles 2 --min-alleles 2)[26]. SNPs with missing data frequency > 2% were discarded, and the k-nearest neighbour method was then used to impute the missing genotypes at the retained SNPs.Genomic prediction analysis
-
The following random regression BLUP model was used to evaluate the performance of genomic predictions for each trait:
$ {y} =\mu {1} _{ {n} } + {Xg} + {\varepsilon } $ (2) where, y is a vector of n BLUPs obtained from Eq. (1);
$ \mu $ $ \sigma _{g}^{2} $ $ \sigma _{\varepsilon }^{2} $ The predictive ability was evaluated using the "x.val" function in the R package PopVar[27]. The ten-fold cross-validation (nFold = 10) procedure was repeated 25 times (nFold.reps = 25). In each replication, the seedling population was split into 10 equal sets, each containing one-tenth of the seedlings, which was predicted by the remaining nine-tenths of the population. The accuracy of genomic prediction was expressed as the average correlation between the genomic predicted breeding value (GEBV) and observed BLUP values in the validation set across all folds and replications.
-
The concentrations of key phytochemical compounds detected in this study are presented in Table 1. The groups of compounds included chalcones (CHALC), cinnamic acids (CINNA), FLAVA, FLAVO, triterpenoids, and AA-2βG.
Table 1. Across-season distribution of the content (μg/g) of phenolics, triterpenoids, and organic acids in the seedling population.
Compound Group Min. Max. Mean SD Corr. 3-Hydroxyphloretin 2'-xyloglucoside Chalcone 0 39.7 2.5 4.7 0.89 3-Hydroxyphloridzin Chalcone 0 43.2 2.3 4.6 0.91 Phloretin-2'-O-xyloglucoside Chalcone 0 386 43.1 46.1 0.81 Phloridzin Chalcone 1 558.8 38.4 63.8 0.80 Sieboldin Chalcone 0 290 9.3 35.6 0.80 Trilobatin Chalcone 0 107.2 4.2 13.0 0.54 5-Caffeoylshikimic acid Cinnamic acid 0 290 14 31.5 0.70 Chlorogenic acid Cinnamic acid 1.1 2,495.8 389 387.6 0.89 Neochlorogenic acid Cinnamic acid 0 27.6 1.4 3.6 0.85 trans-4-p-Coumaroyl quinic acid Cinnamic acid 0 463.9 32.1 49.3 0.91 trans-5-p-Coumaroyl quinic acid Cinnamic acid 0 276.4 20.8 31.5 0.77 Catechin Flavanol 0 318.2 11 25.0 0.97 Epicatechin Flavanol 1.6 932.1 140.1 154.5 0.95 Procyanidin B1 Flavanol 0 356.5 33.5 45.4 0.92 Procyanidin B2 Flavanol 4 1,358 248.2 282.9 0.93 Procyanidin B5 Flavanol 0.2 148 20.8 24.8 0.96 Procyanidin B7 Flavanol 0 78.2 12 13.5 0.94 Procyanidin C1 Flavanol 2.4 1,056 128.3 171.7 0.96 Quercetin 3-arabinopyranoside Flavonol 4.4 264.2 26.8 21.3 0.30 Quercetin 3-galactoside Flavonol 2.1 341.9 41.3 32.1 0.37 Quercetin 3-glucoside Flavonol 0.9 304 16.4 23.3 0.71 Quercetin 3-rhamnoside Flavonol 4.4 84.8 19 12.0 0.60 Annurcoic acid Triterpenoid 1.5 318 70.9 66.2 0.61 Betulinic acid Triterpenoid 1.3 17.1 4.6 1.8 0.53 Corosolic acid Triterpenoid 2.6 39.9 12.2 5.9 0.63 Euscaphic acid Triterpenoid 0.4 255.8 29.8 33.4 0.58 Maslinic acid Triterpenoid 2.5 44.6 11.2 6.2 0.69 Medicagenic acid Triterpenoid 0 116.3 3.7 10.3 0.17 Pomolic acid Triterpenoid 1.5 35.5 14.9 7.9 0.63 trans-Caffeoylbetulinic acid Triterpenoid 0 22.7 2.8 2.6 0.01 Ursolic acid Triterpenoid 10.6 31.9 18.9 3.8 0.26 Ascorbic acid 2-beta-glucoside Organic acid 0 1,317.5 122.7 236.2 0.87 Sorbitol Carbohydrate 1.1 31.7 8.3 5.2 0.74 Glucose Carbohydrate 3.3 37.3 13.5 6.7 0.77 Sucrose Carbohydrate 2.5 183.1 40.4 14.8 0.69 Fructose Carbohydrate 12.5 92.9 37.7 9.5 0.54 Min. = minimum. Max. = maximum. SD = standard deviation. Corr. = between-season correlation. The content (mg/g) of individual sugars in the fruit is also presented. Across the population, all compounds showed substantial variation among seedlings (Table 1). Among the phenolic compounds, the average concentration of chlorogenic acid was the highest (389 μg/g), followed by procyanidin B2 (248 μg/g) and epicatechin (140 μg/g). In general, the concentrations of FLAVA compounds were higher than those of other phenolic compounds. The concentration of annurcoic acid (70.9 μg/g) was the highest among all TRITP compounds measured in this study. The average concentration of AA-2βG was 122.7 μg/g, with a range between 0 and 1,317.5 μg/g (Table 1). Sucrose was the most abundant sugar compound, with an average concentration of 40.4 mg/g and ranging between 2.5 and 183.1 mg/g (Table 1). The between-season correlation was high (> 0.80) for CHALC and FLAVA, suggesting that these compounds are more consistent than FLAVO, TRITP, and carbohydrates (CARBO) (Table 1).
As the concentrations of individual compounds within each group (e.g., CHALC, FLAVA, FLAVO) were generally strongly correlated, group concentrations were used to simplify subsequent analyses. Strong correlations among compounds within a group suggest that these compounds are synthesised through shared biochemical pathways[28,29]; hence, the use of group concentrations could enable more efficient screening/selection in breeding programmes when the goal is to improve the overall antioxidant capacity[30]. For all subsequent analysis (i.e., genetic parameter estimation and GS), the concentrations of individual compounds within a group (see Table 1) were first summed to obtain the content of total CHALC, total CINNA, total FLAVA, total FLAVO, total triterpenoids (TRITP), and total CARBO for each seedling. The total phenolic (TPHEN) content was also calculated (= CHALC + CINNA + FLAVA + FLAVO) for each seedling.
The distribution of group total concentrations, along with other traits (AA-2βG, fructose-glucose ratio [FGRAT], SIZE, SWEET, SOUR, ASTR, and EQUAL) in the assessed seedlings are presented in Fig. 1. The average SOUR score was higher than those of other sensory traits, and there was large variation for all sensory traits among seedlings in this study. The CARBO content varied between 59 and 311 mg/g, and the FGRAT varied between 1.0 and 9.55, with an average of 99.8 mg/g and 3.51, respectively (Fig. 1). The average FLAVA content (594 μg/g) was the highest compared with other groups of phenolic compounds, and the average concentrations of CHALC and FLAVO were very similar (100 μg/g). TRITP content was the least variable group of phytochemical compounds, with an average of 169 μg/g and a range of 518 μg/g in this study population. The content of AA-2βG varied between 0 and 1,317 μg/g, with an average of about 123 μg/g (Fig. 1).
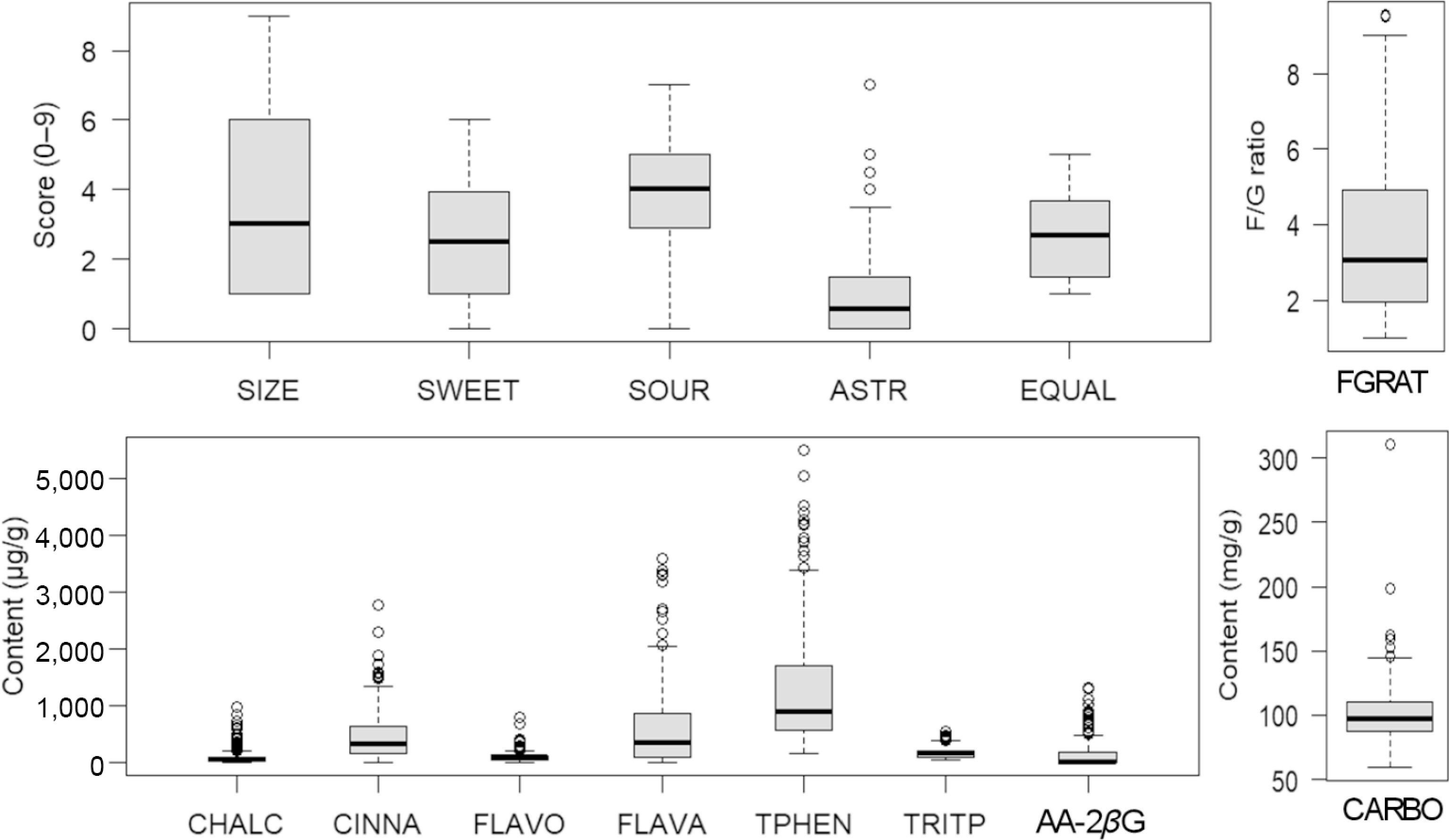
Figure 1.
Range and distribution of the content of phytochemicals (CHALC: chalcones, CINNA: cinnamic acids, FLAVO: flavonols, FLAVA: flavanols, TPHEN: total phenolics, TRITP: triterpenoids, AA-2βG: ascorbic acid 2-beta-glucoside), sensory traits (SIZE: fruit size, SWEET: sweetness, SOUR: sourness, ASTR: astringency, EQUAL: eating quality), and sugars (CARBO: carbohydrates, FGRAT: fructose–glucose ratio) in the fruits from all apple seedlings.
Genetic parameters
-
The 14 traits shown in Fig. 1 were considered for the estimation of genetic parameters. The additive genetic variance as a proportion of the total phenotypic variation is an estimate of the narrow-sense heritability (h2), and was the highest for SIZE (0.84), followed by FGRAT (Fig. 2). Several traits, such as ASTR, CINNA, FLAVA, SOUR, and TRITP, were found to be moderately heritable (h2 ≈ 0.50). FLAVO was the least heritable trait in this study (Fig. 2). The estimated heritability for AA-2βG (0.39) was very similar to that for total sugars (CARBO; 0.33) (Fig. 2).
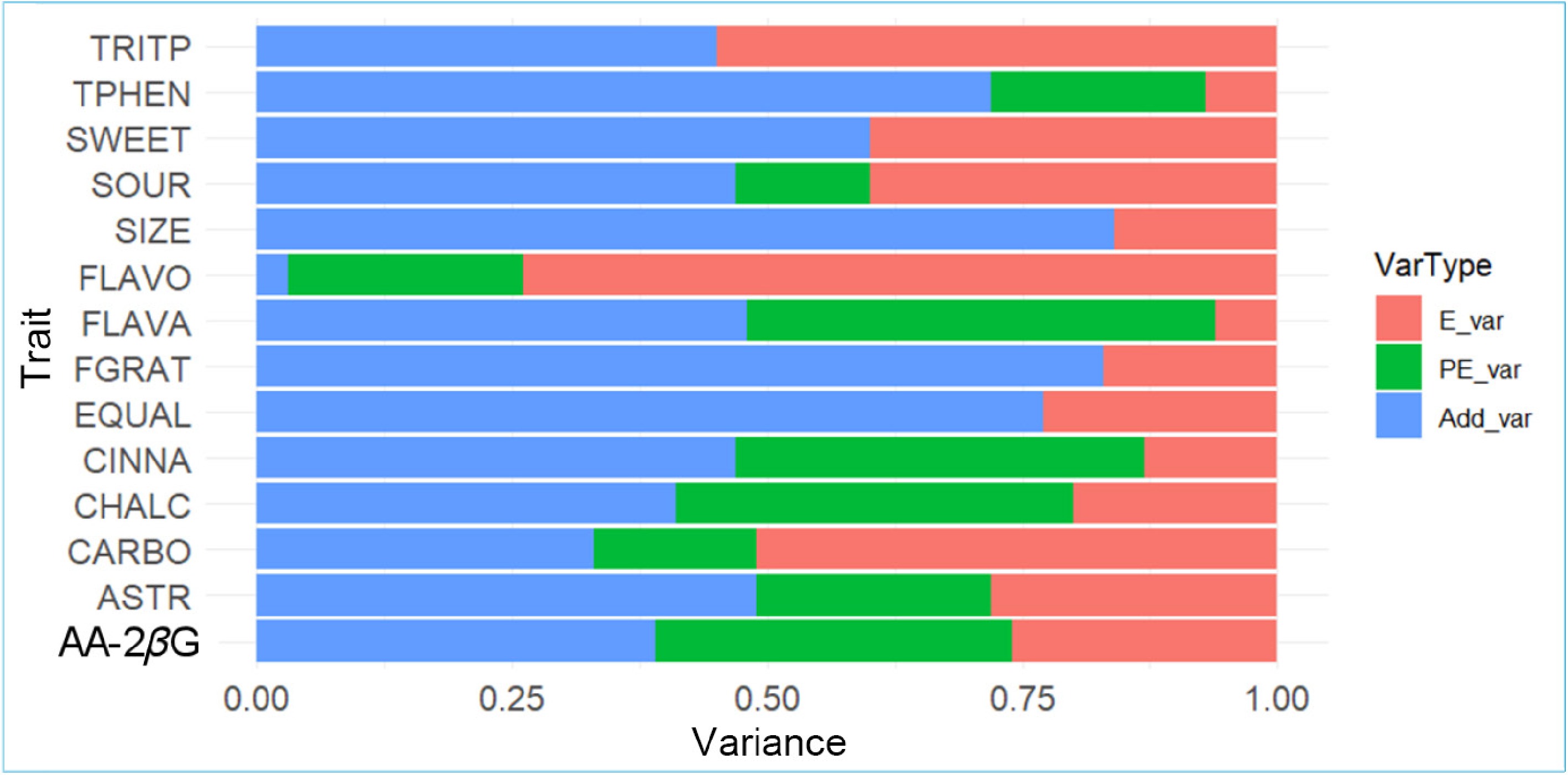
Figure 2.
Estimated proportion of the additive genetic (Add_var), permanent environment (PE_var), and environmental (E_var) variances for different traits (ASTR: astringency, CARBO: carbohydrates, CHALC: chalcones, CINNA: cinnamic acids, EQUAL: eating quality, FGRAT: fructose–glucose ratio, FLAVO: flavonols, FLAVA: flavanols, AA-2βG: ascorbic acid 2-beta-glucoside, SIZE: fruit size, SOUR: sourness, SWEET: sweetness, TRITP: triterpenoids, and TPHEN: total phenolics).
The variance attributed to the permanent environmental effects (i.e., consistent non-genetic differences between individuals across repeated records) was close to zero for SIZE, SWEET, EQUAL, FGRAT, and TRITP, but varied between 13% (SOUR) and 46% (FLAVA) for the other traits. For all groups of phenolic compounds (CHALC, CINNA, FLAVA, and FLAVO), there was substantial repeatable environmental variation (average 37%) at the individual plant level.
The nature of the phenotypic and genetic correlations was similar, but the magnitude of genetic correlations was somewhat higher than that of the phenotypic correlations (Fig. 3), because phenotypic correlations include not only genetic influences but also environmental influences, some of which may be uncorrelated or even negatively correlated between traits. EQUAL was inversely correlated with different groups of phenolic compounds (CHALC, CINNA, FLAVA, and FLAVO) but positively correlated with TRITP and CARBO. AA-2βG displayed positive correlations with ASTR, SOUR, and phenolics (Fig. 3). SIZE was negatively correlated with AA-2βG, phenolics, and ASTR, but a positive correlation was observed with CARBO, TRITP, and EQUAL. The moderate positive genetic correlation (0.40) between ASTR and TPHEN shows the trade-off required in selection. FGRAT was strongly correlated (> 0.70) with SWEET and EQUAL (Fig. 3).
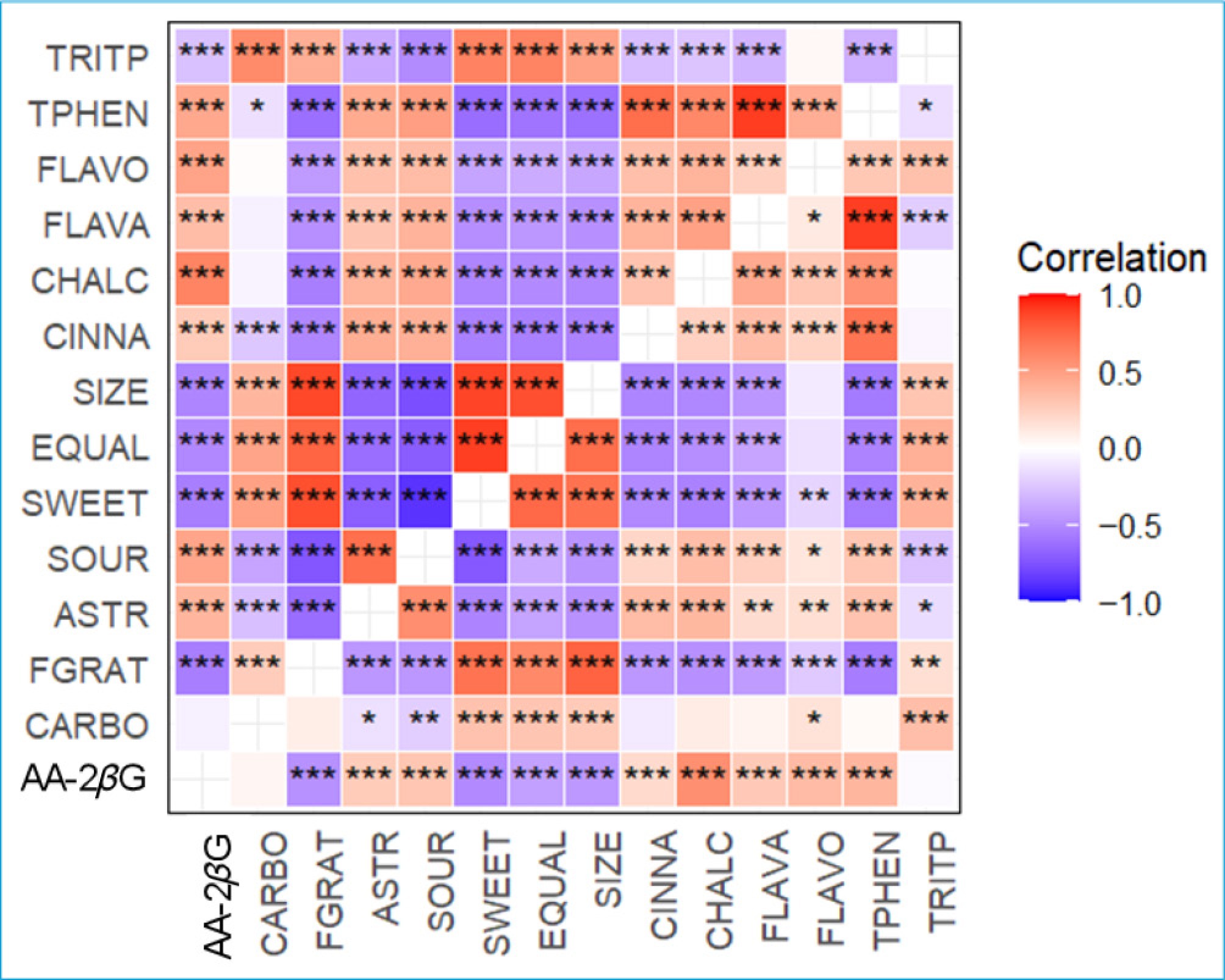
Figure 3.
Genetic (upper-left triangle) and phenotypic (lower-right triangle) correlations between pairs of traits (ASTR: astringency, CARBO: carbohydrates, CHALC: chalcones, CINNA: cinnamic acids, EQUAL: eating quality, FGRAT: fructose–glucose ratio, FLAVO: flavonols, FLAVA: flavanols, AA-2βG: ascorbic acid 2-beta-glucoside, SIZE: fruit size, SOUR: sourness, SWEET: sweetness, TRITP: triterpenoids, and TPHEN: total phenolics) in the seedling population. Significance levels of correlation coefficients: *** p < 0.001; ** p < 0.01, * p < 0.05.
The results presented in Fig. 4 show that the total phenolic concentration decreases as EQUAL increases. On the other hand, TRITP and FGRAT increased with an increase in EQUAL. A few seedlings with an EQUAL score of 4 displayed total phenolic and TRITP contents of approximately 1,100 and 195 μg/g, respectively, with an FGRAT of 3.0. Upon lowering the threshold for EQUAL to 3.0, we identified a seedling with total phenolics, TRITP, and FGRAT of 1,686 μg/g, 221 μg/g, and 1.42, respectively (Fig. 4), indicating a trade-off between EQUAL and phytochemical content.
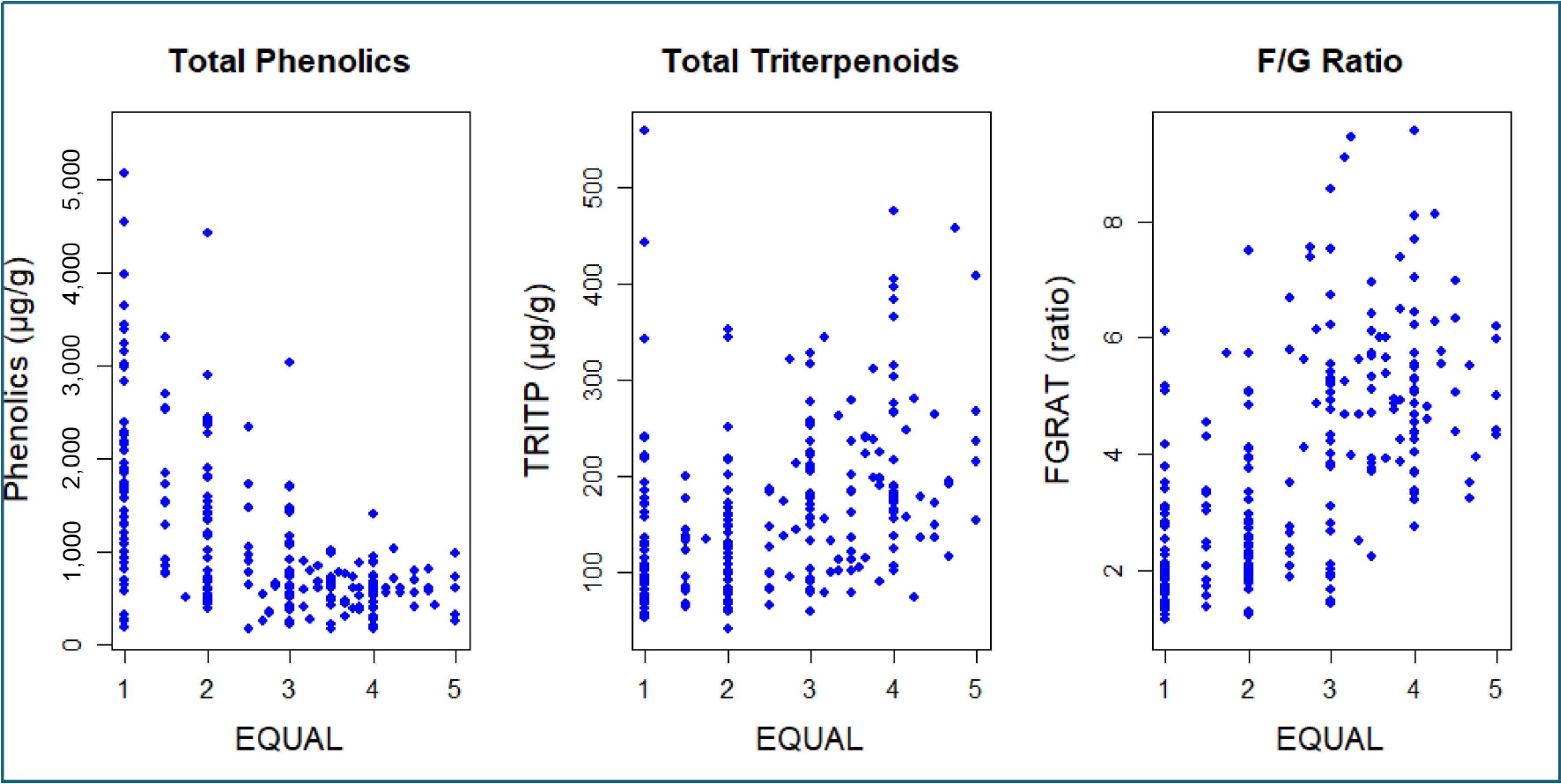
Figure 4.
Phenotypic covariation between the eating quality (EQUAL) and some phytochemicals in the seedling population.
Genomic predictions
-
A total of approximately 186,000 SNPs were retained following the quality control measures. The accuracy of genomic prediction for various traits ranged from 0.50 (FLAVO) to 0.92 (SIZE), with an average accuracy of 0.75 (Fig. 5a). Except for CHALC, the other phenolic groups (CINNA, FLAVA, and FLAVO) were among the traits with the lowest prediction accuracy. The standard deviations of prediction accuracies for ASTR (0.16) and FLAVO (0.14), calculated from the cross-validation replications, were among the highest of all traits. The degree of prediction accuracy was highly correlated (r = 0.72) with the heritability of traits (Fig. 5b).
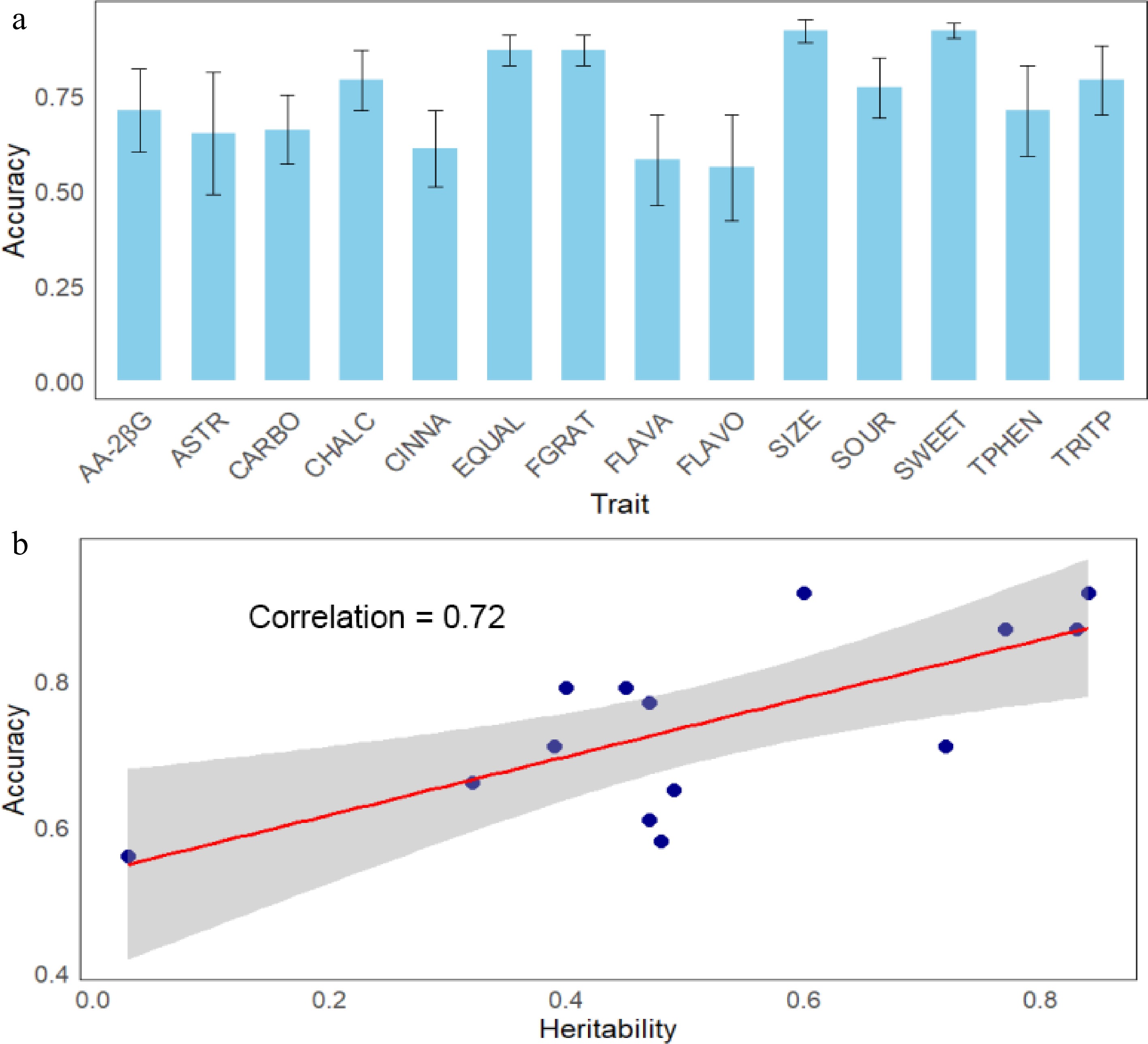
Figure 5.
Genomic prediction accuracies (± standard deviation) for various traits (a). AA-2βG: ascorbic acid 2-beta-glucoside, ASTR: astringency, CARBO: carbohydrates, CHALC: chalcones, CINNA: cinnamic acids, EQUAL: eating quality, FGRAT: fructose–glucose ratio, FLAVO: flavonols, FLAVA: flavanols, SIZE: fruit size, SOUR: sourness, SWEET: sweetness, TRITP: triterpenoids, TPHEN: total phenolics. Relationship between trait heritability and prediction accuracy (b). The grey area in (b) depicts the 95% confidence interval for the linear fit (red line).
-
Enhancing the phytochemical content in new apple cultivars is aimed at biofortifying the fruit with health-promoting compounds, while ensuring that EQUAL is maintained. Breeding of fruit crops has historically focused on the introgression of disease resistance, but efforts are now underway for the introgression of phytochemicals from wild species into commercial cultivars. Our study population represents the first generation of pre-breeding aimed at improving the phytochemical content and EQUAL simultaneously in future cultivars.
Historically, consumers' preference for sweet, non-astringent apples with slow browning probably led breeders to indirectly select for lower contents of some polyphenols[31]. Indeed, analyses show that the modern apple gene pool tends to have significantly lower phenolic content in the flesh than wild species and older heirloom cultivars[15,17]. Kumar et al.[7] showed that the average contents of phenolic compounds (e.g., phloridzin, chlorogenic acid, epicatechin, procyanidin B2, and procyanidin C1) were significantly lower in the cultivated Malus gene pool than in the wild species. The F1 seedlings, generated from hybridisation between the domesticated and wild species in this study, showed higher average concentrations of some phenolic compounds. For example, the average concentrations of phloridzin (38.4 μg/g), procyanidin B2 (248 μg/g), and procyanidin C1 (128 μg/g) were improved considerably compared with previously reported[7] concentrations in the domesticated gene pool.
Triterpenic compounds are present mainly in apple peel and its associated epicuticular wax[32]. The concentration of these compounds was previously reported to be similar between the domesticated and wild apple gene pools[7,15]. Over the past few decades, the incidence of chronic inflammatory diseases such as inflammatory bowel disease (IBD) has increased, and some studies have shown the ability of triterpenes and certain phenolic compounds to modulate inflammation processes[33]. Pentacyclic triterpenes are notably present in plant surfaces such as stem bark, leaves, and fruit waxes, where they protect the plant from biotic and abiotic stress factors[34]. Our study showed a favourable genetic correlation between TRITP concentration and overall EQUAL. The estimated moderate heritability of TRITP, along with a favourable correlation, suggests that a targeted genetic improvement of TRITP is feasible without compromising EQUAL.
Soluble sugars, mainly composed of sucrose, fructose, glucose, and sorbitol (an alcohol sugar), are an important component of sensory sweetness, which has been given high priority in apple breeding programmes. Only a few studies[35] have investigated the relationship between sensory sweetness and the concentration of individual sugars in apples, and apparently, there are no published reports of any relationship between sensory sweetness and FGRAT. Glucose content contributes lesser than fructose to perceived sweetness[36]; hence, apples with lower glucose but higher fructose (i.e., high FGRAT) may be rated as sweeter, even if the total sugar content stays the same. This makes FGRAT more important in consumer preference, flavour breeding, and marketability. Given a strong correlation between SWEET and FGRAT, modern apple cultivars generally display an unfavourable FGRAT of > 1.0, making them potentially unsuitable for patients suffering from fructose malabsorption problems[18]. Our results suggest that selection for apple cultivars with lower FGRAT scores would also enhance the phenolic content but would require innovative breeding strategies to balance the trade-off with EQUAL.
The sensory analysis was carried out by a single trained/experienced assessor, which limits the scope to account for variability in perception thresholds and liking preferences. Trained assessors regularly familiarise themselves with known reference standards for different sensory attributes. The standard apple cultivar breeding typically follows three stages[20]. The seedling population used for estimation of the genetic parameters in this study is part of Stage-1 evaluation, in which the main goal is mass elimination rather than a detailed characterisation of thousands of seedlings. The estimated heritability for the sensory traits ranged from 0.50 (ASTR) to 0.77 (EQUAL), and the between-season correlation ranged from 0.60 (SOUR) to 0.80 (EQUAL), suggesting that the assessor was consistent in evaluating several hundred seedlings. The selected seedlings are cross-checked by several assessors/researchers to ensure consistency before progressing to Stage-2 testing. Consumer studies are generally conducted on Stage-3 selections, which involve trained panellists and sometimes consumers as well. Because the Stage-2 and Stage-3 testing populations comprise only a few selections with limited genetic diversity, they are not ideal for estimation of genetic parameters.
Estimates of genetic parameters for sensory traits should be interpreted with caution as these traits are influenced by evaluator preferences and sensory protocols. While broad trends can be compared across studies, direct numerical comparisons should be made cautiously unless sensory evaluation protocols and analytical approaches are harmonised. Additionally, when sensory traits are assessed by a single evaluator, the resulting phenotypes may reflect not only underlying genetic differences but also consistent assessor-specific preferences, which can be confounded by genetic effects. This may lead to inflated heritability estimates and genetic correlations that partly capture the scoring bias rather than biological relationships, while genomic predictions may perform well within the same context but decline when applied in more diverse or independent settings. With multiple assessors, evaluator differences introduce additional variation that can reduce the heritability if unmodeled. Accounting for assessor and genotype interactions improves variance partitioning, yielding more robust and generalisable genetic parameter estimates.
Although the family sizes were unequal, some common cultivars were involved in the pedigree of the seed parents, which provided genetic connectedness among the seedlings from different crosses. In the individual-tree (animal) restricted maximum likelihood (REML)/BLUP model used in this study, each seedling is weighted according to its own information and the performance of its relatives, resulting in more precise and unbiased estimates of genetic variance. Moderate-to-high heritabilities of most phenolic compounds were consistent with those reported earlier[7,37]. The moderate heritability (≈ 0.40) and the high between-season correlation (≈ 0.90) observed for AA-2βG in this study are consistent with our previous research[7], providing further evidence of this compound's stability[38]. AA-2βG displayed a moderate negative correlation with EQUAL and a positive correlation with phenolic content (Fig. 3). Studies on berryfruits[39] and apples[40] have shown that AsA treatment increases the polyphenol content in the fruits, suggesting some positive association. A Japanese study showed that an increase in AsA content in apple juice resulted in reduced fruity and apple-like odours[41], which could pose challenges for a targeted improvement of this trait along with EQUAL.
Introgression of major gene traits (e.g., disease resistance), using pseudo-backcrossing (PBC), from wild apple species into elite lines or commercial cultivars is a common practice in apple breeding programmes[42]. In a conventional PBC design, foreground selection for monogenic traits is carried out using marker-assisted selection (MAS) and the resistant seedlings are then planted in the orchard for fruit quality evaluation. Studies in fruits[7] and other crops[43,44] have revealed the complex polygenic control of phenolics and other antioxidant compounds and suggested that a GS approach would be more effective for the introgression of polygenic traits (e.g., phytochemicals) from wild donors into elite lines[45].
In apple, GS could eliminate the requirement to evaluate seedling fruit, thus shortening the breeding cycle by several years. GS is actively being explored as a replacement for Stage-1 evaluation in apple breeding; evidence suggests that it can outperform traditional phenotypic selection in certain contexts, although the benefits depend on trait architecture, population structure, and breeding scenario[20,46]. In this study, GS accuracy ranged from 0.50 to 0.92 across different traits, and the across-trait average accuracy (0.74) was similar to that of phenotypic selection (0.70), providing further support for the use of GS in apple breeding. The introgression strategy would probably require multiple cycles of PBC with elite parents to minimise the non-desirable genomic content of the donors in selected parents[47], compounding the time benefits of GS and fast-forwarding the production of new cultivars with both increased concentrations of health-associated compounds and eating qualities. Further research with a larger sample size will help improve our understanding of the genetic relationships between phytochemicals and EQUAL traits, as well as enhance the accuracy of genomic predictions.
-
This investigation revealed a moderate degree of inverse correlations (≈ −0.45) between phenolic content and the overall eating quality (EQUAL) of the apples studied. The moderate-to-high heritability of these traits suggested that both traits may still be improved simultaneously. The positive association between EQUAL and triterpenoids (the main anti-inflammatory compounds in apples) suggested that faster gain can be made in TRITP without compromising EQUAL. The moderate heritability and high stability across seasons suggested that AA-2βG can be introgressed as a novel trait in future apple cultivars; however, the moderate negative correlation with EQUAL means a trade-off could be required in the selection process. The moderate-to-high genomic prediction accuracy for most traits suggests that GEBVs could be combined to develop selection indices for faster and simultaneous improvement of EQUAL traits and phytochemicals in pre-breeding programmes. Our study provides a valuable foundation for breeding apples with higher phytochemical content while simultaneously maintaining EQUAL.
This research was supported by the New Zealand Institute for Bioeconomy Science Limited's Technology Development – Pipfruit programme. We thank Helen Boldingh for conducting the sugar analysis. The constructive feedback from Richard Volz and Alastair Currie is thankfully acknowledged.
-
The authors confirm their contributions to the paper as follows: designed the project, analysed the data and prepared the first draft: Kumar S; coordinated the field sampling and collected the lab data: Molloy C, Stoklosinski H, Hunt M; coordinated DNA extraction and skim sequencing: Hilario E; conducted bioinformatics: Deng CH. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Kumar S, Deng CH, Hilario E, Molloy C, Hunt M, et al. 2026. Simultaneous improvement of phytochemical content and eating quality in apples: estimates of genetic parameters. Fruit Research 6: e021 doi: 10.48130/frures-0026-0013 

Simultaneous improvement of phytochemical content and eating quality in apples: estimates of genetic parameters
- Received: 03 November 2025
- Revised: 10 April 2026
- Accepted: 14 April 2026
- Published online: 04 June 2026
Abstract: Modern apple cultivars tend to have significantly lower phenolic content than wild species and heritage cultivars. Although the phytochemical makeup of apple fruit affects important traits, such as nutrition, flavour, texture and disease resistance, phytochemical-based breeding strategies for apple improvement are lacking. We studied the relationships between fruit phenolic content and sensory traits in a seedling population derived from crosses between the domesticated and wild apple species. We also investigated the potential of genomic selection for breeding new high-health cultivars faster. Phenolic content, sensory traits, sugar levels, and a stable vitamin C glycoside (ascorbic acid 2-beta-glucoside [AA-2βG]) were assessed to estimate the genetic parameters of these traits. Phenolic compounds (chalcones, flavanols [FLAVA], cinnamic acids), triterpenoids (TRITP), carbohydrates (CARBO), AA-2βG, and sensory traits (astringency [ASTR], sourness [SOUR]) were found to be moderately heritable (h2 = 0.35–0.50), while fructose–glucose ratio (FGRAT), fruit size (SIZE), and overall eating quality (EQUAL) were highly heritable (h2 > 0.75). FGRAT was strongly correlated (> 0.70) with sweetness (SWEET) and EQUAL. Phenolics and AA-2βG were positively correlated, but both displayed moderate negative correlations with EQUAL. TRITP content displayed a moderate positive correlation with EQUAL and negative correlation with SOUR and ASTR. The accuracy of genomic prediction ranged from 0.50 (FLAVA) to 0.90 (SIZE) and was highly correlated with trait heritability. The knowledge derived from this study can assist in designing DNA-based fast-breeding strategies for simultaneous improvement of phytochemicals and sensory traits.
-
Key words:
- Apple breeding /
- Genetic parameters /
- Phytochemicals /
- Phenolics /
- Genomic selection