









-
Floral scent is a key ornamental trait giving flowers their distinctive sensory and aesthetic value. It also plays essential roles in pollinator attraction, chemical/ecological defenses, and human utilization[1,2]. Precise and efficient characterization of floral volatile profiles is therefore important for both basic research and breeding applications[3]. The electronic nose (E-nose), or artificial olfactory system, is a VOC-sensing platform composed of a multisensor array. The array mimics biological olfaction and enables rapid, non-destructive detection of complex odor mixtures[4,5]. Compared with conventional approaches such as GC–MS, coupled with solvent extraction and chromatographic separation, E-nose analysis measures headspace volatiles directly, features high throughput, simple operation, and portability. Moreover, an E-nose could avoid contamination from organic solvents and provide more objective and reproducible results than human sensory evaluation[5,6]. Consequently, E-nose technology has been widely used in quality control and authenticity identification in the food and fermentation industries (e.g., Chinese Baijiu, coffee, and dairy products) and has also been introduced into the fragrance assessment of several ornamentals, including rose, iris, crabapple, and peach[7−13].
Clematis L. (Ranunculaceae) comprises woody or herbaceous climbing species[14]. China is one of the main centres of diversity, and many species have subsequently been introduced to Japan, Korea, and Europe, leading to a wide global distribution. More than 300 Clematis species have been reported worldwide, over 140 of which occur in China[15,16]. The Royal Horticultural Society classifies Clematis plants based on flowering time, growth habit, and pruning type. The horticultural groups include the Early Large-Flowered, Late Large-Flowered, Florida, Forsteri, Texensis, Integrifolia, and Viorna groups[17]. The Clematis plants are referred to as the ‘queen of climbers' owing to the rich flower forms, wide colour range, strong adaptability, and suitability for vertical greening, landscape planting, pot culture, and cut-flower use[18−20]. Now, they have become an important germplasm resource in ornamental horticulture. The research of Clematis plants primarily focuses on its medicinal properties, cultivation and growth conditions, cytology, morphological diversity, and interspecific hybridization. However, the fragrance trait of Clematis has received relatively little attention, which impedes the targeted breeding of aromatic varieties[21−25]. In practice, Clematis cultivars differ markedly in scent intensity—from strongly fragrant to weakly fragrant or scentless. This trait has clear potential for the fresh-cut and landscape markets[26,27].
In this study, we selected Clematis × cartmanii 'Joe' as the model material using an orthogonal experimental design to evaluate the effects of headspace equilibration time, flowering stage, and sample temperature on E-nose detection of Clematis floral scent. On this basis, we optimized E-nose parameters and established a rapid detection procedure. The optimized system was applied to 97 Clematis germplasm resources from 11 horticultural groups to obtain their E-nose response profiles. This study constructed fragrance fingerprints to compare aroma response characteristics among groups, and inferred the key categories of volatile compounds involved.
-
From March to July 2025, 97 Clematis germplasm resources (three wild/native species and 94 cultivated varieties) representing 11 horticultural groups were collected from the Clematis germplasm nursery of the Jiangsu Vocational College of Agriculture and Forestry (Zhenjiang, Jiangsu Province, China; latitude: 31°37' N, longitude: 118°57' E) (Table 1). All plants were healthy and at normal flowering time. The cut inflorescences were sampled at 09:00–10:00.
Table 1. Names of 97 Clematis samples for testing.
No. Germplasm name Group No Germplasm name Group 1 Clematis × cartmanii 'Joe' Forsteri group (one) 50 Clematis 'Sea Breeze' Viticella group (15) 2 Clematis 'Rōguchi' Integrifolia group (nine) 51 Clematis 'Dark Eyes' 3 Clematis 'Sizaia Ptitsa' 52 Clematis 'Astra Nova' 4 Clematis 'Hanajima' 53 Clematis 'Madame Julia Correvon' 5 Clematis 'Arabella' 54 Clematis 'Buckland Pixie' 6 Clematis 'East River' 55 Clematis 'Krakowiak' 7 Clematis 'Hakurē' 56 Clematis 'Emilia Plater' 8 Clematis 'Star River' 57 Clematis 'Étoile Violette' 9 Clematis 'Petit Faucon' 58 Clematis 'Rosalyn' 10 Clematis 'Inspiration' 59 Clematis viticella 'Betty Corning' 11 Clematis texensis 'Duchess of Albany' Texensis group
(three)60 Clematis 'Fairydust' 12 Clematis 'Gravetye Beauty' 61 Clematis × integrifolia 'Super Night' 13 Clematis 'Princess Diana' 62 Clematis 'Sunny Side' 14 Clematis 'Utopia' Florida group (nine) 63 Clematis 'Super Cute' 15 Clematis 'Alba Plena' 64 Clematis 'Bijou' Early Large-Flowered group (34) 16 Clematis florida 'Shamshel' 65 Clematis 'Guernsey Cream' 17 Clematis 'Shishimaru' 66 Clematis 'The Countess of Wessex' 18 Clematis 'Best Wishes' 67 Clematis 'Yukiokoshi' 19 Clematis 'Viennetta' 68 Clematis patens 'Ruriokoshi' 20 Clematis 'Cassis' 69 Clematis 'Doctor Ruppel' 21 Clematis 'Taiga' 70 Clematis 'Hakuokan' 22 Clematis 'Light Lnk' 71 Clematis 'Hayachine' 23 Clematis 'Rubens Superba' Montana group (one) 72 Clematis 'Parisienne' 24 Clematis 'Amethyst Beauty' Late Large-Flowered group
(14)73 Clematis 'Diamantina' 25 Clematis 'Lucky Charm' 74 Clematis 'Isabella' 26 Clematis' Moonfleet' 75 Clematis 'Moonglow' 27 Clematis 'Shimmer' 76 Clematis 'Ai-nor' 28 Clematis 'Reflections' 77 Clematis 'Samaritan Jo' 29 Clematis 'Polish Spirit' 78 Clematis 'HF Young' 30 Clematis 'Ville de Lyon' 79 Clematis 'Dorothy Tolver' 31 Clematis 'Comtesse de Bouchaud' 80 Clematis 'Giselle' 32 Clematis 'Grunwald' 81 Clematis 'Empress' 33 Clematis 'Danuta' 82 Clematis 'Viva Polonia' 34 Clematis 'Forever Friends' 83 Clematis 'Neva' 35 Clematis 'Ernest Markham' 84 Clematis 'Red Star' 36 Clematis 'Blue Angel' 85 Clematis 'Little Mermaid' 37 Clematis 'Basilisk' 86 Clematis 'Reiko' 38 Clematis 'Angel Pink' Viorna group
(seven)87 Clematis 'Blue Light' 39 Clematis 'Sonnette' 88 Clematis 'Haruhime' 40 Clematis 'Akanenotsubo' 89 Clematis 'Diamond Ball PBR' 41 Clematis 'Oshikiri' 90 Clematis 'Prinsesse Alexandra' 42 Clematis 'Odoriba' 91 Clematis 'Josephine' 43 Clematis 'Sophie' 92 Clematis 'Beautiful Bride' 44 Clematis 'Taigetsu' 93 Clematis 'Ooh La La' 45 Clematis 'Apple Blossom' Armandii group (one) 94 Clematis 'Lula' 46 Clematis finetiana Wild group
(three)95 Clematis 'Bees Jubilee' 47 Clematis cadmia 96 Clematis 'Bernadine' 48 Clematis terniflora 97 Clematis 'Rosemoor' 49 Clematis 'Minamonoyousei' Optimization of E-nose parameters for floral scent detection
-
Floral scent was analyzed using a portable PEN3 E-nose (AIRSENSE, Germany) equipped with a 10-sensor metal oxide (MOS) array (W1C, W5S, W3C, W6S, W5C, W1S, W1W, W2S, W2W, and W3S; Table 2). The instrument is capable of automatic adjustment/calibration and automatic enrichment. To minimise sensor drift, the array was cleaned and standardised before and after each measurement, which ensured the stability and reproducibility of the data.
Table 2. Characteristics of E-nose sensors.
Sensor no. Sensor name Sensitive components Reference 1 W1C Aromatic components Toluene, C7H8 2 W5C Highly sensitive to nitrogen oxides NO2 3 W3C Ammonia water, sensitive to aromatic components Benzene 4 W6S Mainly selective to hydrogen H2 5 W5S Alkanes, aromatic components Propane, C3H8 6 W1S Sensitive to methane CH3 7 W1W Sensitive to sulfides H2S 8 W2S Sensitive to ethanol CO 9 W2W Aromatic components, sensitive to organic sulfides H2S 10 W3S Sensitive to alkanes CH4 To identify the optimal detection conditions, we used Clematis × cartmanii 'Joe' as the test material and designed a three-factor, three-level orthogonal experiment. The three factors were: (1) flowering stage, (2) sample temperature, and (3) headspace equilibration time (Table 3); each treatment was performed with three biological replicates. Flowering stage was defined as: large bud stage (pre-anthesis), full flowering stage (2nd day after anthesis), and late full flowering stage (4th day after anthesis, when pollen was mature and dispersing). Fresh flowers were immediately placed into headspace vials and sealed, then incubated on a thermostatic magnetic stirrer (Labnet, USA). After reaching the preset equilibration time, samples were analyzed with the E-nose. Measurement parameters were: sampling interval 1 s, pre-sampling time 5 s, sampling time 60 s, auto-zero 10 s, sensor cleaning 40 s, internal flow 400 mL·min−1, and injection flow 400 mL·min−1.
Table 3. Factors and levels of the orthogonal design.
Level Flower stage (A) Samples temperature (B)/°C Equilibration time (C)/h 1 Large bud stage 20 0.5 2 Full flowering stage 40 2 3 Late full flowering stage 60 6 E-nose measurement of floral scent in 97 Clematis germplasm resources
-
The 97 Clematis samples were then tested under the optimal conditions determined above, so that floral-scent responses were directly comparable among groups and cultivars. For each sample, three biological replicates were measured using the same E-nose settings as in the orthogonal test.
Data analysis
-
For each run, the average sensor value at the stationary phase of the response curve was used as the characteristic value of that sample. Principal component analysis (PCA) and loading analysis were carried out using the instrument's built-in software WinMuster v1.6.2 (AIRSENSE, Germany). Radar plots were generated in Origin 2024 (OriginLab, USA). One-way analysis of variance (ANOVA) and multiple comparisons by Duncan's test of sensor data were performed in SPSS v19.0 (IBM, USA), and differences were considered significant at p < 0.05.
-
The 10 MOS sensors of the PEN3 showed distinct responses to the floral volatiles of Clematis × cartmanii 'Joe', with average signals ranging from 1.04 to 1.61. Sensor W2W gave the strongest signal and was significantly higher than all others (p < 0.05). W1W and W1S did not differ from each other but both responded more strongly than the remaining seven sensors, followed by W2S, whereas W1C, W3C, and W5C showed the lowest (and mutually non-significant) responses (Fig. 1a, b). PCA of the 10 sensors showed that PC1 and PC2 explained 93.12% and 4.50% of the variance, respectively, for a cumulative 97.62%, indicating that the first two components sufficiently represented the odor information. In the loading plot, W5S (alkanes, aromatic components) was farthest from the origin on PC1, and W2W (aromatic components, organic sulfides) and W1W (sulfides) also contributed strongly, while W1S (methane) had the highest loading on PC2 (Fig. 1c). These patterns indicate that the main volatile classes perceived in C. × cartmanii 'Joe' were organic/sulfur-containing compounds and aliphatic volatiles based on sensor characteristics.
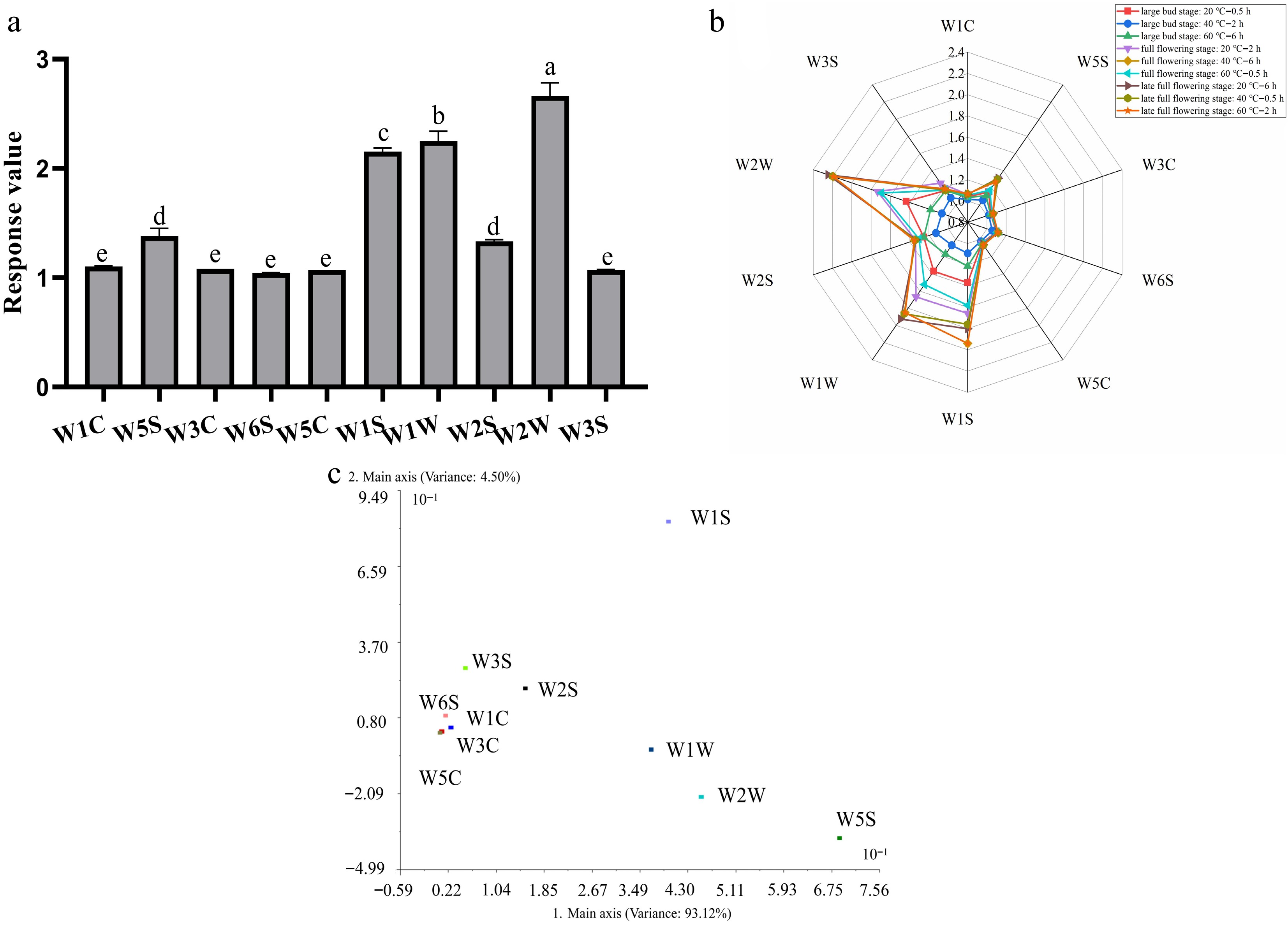
Figure 1.
(a) The variance analysis, (b) radar chart construction, and (c) loading analysis of response values of floral scent components of C. × cartmanii 'Joe' under nine treatments using E-nose technology.
Based on this, four sensors with the highest discriminative capacity (W2W, W1W, W1S, and W2S) were used to analyze the effects of flowering stage, headspace equilibration time, and sample temperature in an L9 (33) orthogonal design (Table 4). Range analysis showed that the influence of the three factors on E-nose response followed the order: flowering stage (A) > headspace equilibration time (C) > sample temperature (B), meaning that flowering stage was the key factor for volatile detectability, while temperature had the weakest effect. At level A3 (late full flowering stage), the responses of W1S, W1W, and W2W were all significantly higher than at A1 (large bud stage) and A2 (full flowering stage) (p < 0.01), indicating that C. × cartmanii 'Joe' emits the strongest and most readily detectable fragrance several days after anthesis. Differences among the levels of B and C were smaller and often non-significant.
Table 4. Different factors affecting the response of the E-nose.
Sensors Level A (flowering stage) B (sample temperature) C (headspace equilibrium time) W1S 1 1.17 ± 0.05c 1.46 ± 0.04ab 1.53 ± 0.06a 2 1.41 ± 0.10b 1.39 ± 0.04b 1.39 ± 0.04b 3 1.77 ± 0.12a 1.50 ± 0.02a 1.43 ± 0.06ab Range 0.60 0.10 0.15 W1W 1 1.17 ± 0.06b 1.52 ± 0.06 1.57 ± 0.10a 2 1.39 ± 0.11b 1.42 ± 0.08 1.35 ± 0.01b 3 1.82 ± 0.16a 1.44 ± 0.03 1.46 ± 0.08ab Range 0.66 0.10 0.22 W2S 1 1.19 ± 0.04b 1.25 ± 0.02ab 1.27 ± 0.05 2 1.27 ± 0.02a 1.24 ± 0.02b 1.22 ± 0.04 3 1.32 ± 0.04a 1.28 ± 0.02a 1.28 ± 0.02 Range 0.14 0.05 0.05 W2W 1 1.18 ± 0.07c 1.65 ± 0.08 1.75 ± 0.09a 2 1.50 ± 0.15b 1.57 ± 0.10 1.47 ± 0.01b 3 2.15 ± 0.21a 1.59 ± 0.06 1.60 ± 0.12ab Range 0.97 0.08 0.28 Different letters (a)–(c) indicate significant differences (p < 0.05) among different levels by Duncan's test. In the orthogonal table, treatments 7 and 8 (both using flowers at the late full-flowering stage with longer equilibration) produced the highest responses for the four key sensors (Table 5). Taking both statistical performance and operability into account, this condition was adopted as the optimal parameter set for subsequent measurements.
Table 5. Sensor response value analysis for C. × cartmanii 'Joe' under various factors and levels.
Treatments Factors and levels Sensors A B C W1S W1W W2S W2W 1 1 1 1 1.20 ± 0.06c 1.26 ± 0.12bcd 1.17 ± 0.05bc 1.27 ± 0.12cd 2 1 2 2 1.09 ± 0.08c 1.07 ± 0.06d 1.13 ± 0.11c 1.07 ± 0.06d 3 1 3 3 1.21 ± 0.09c 1.17 ± 0.07cd 1.25 ± 0.02ab 1.18 ± 0.08d 4 2 1 2 1.37 ± 0.12bc 1.37 ± 0.16bcd 1.25 ± 0.04ab 1.44 ± 0.19bcd 5 2 2 3 1.28 ± 0.09c 1.28 ± 0.09bcd 1.25 ± 0.03abc 1.36 ± 0.13cd 6 2 3 1 1.58 ± 0.15ab 1.52 ± 0.14bc 1.30 ± 0.01a 1.69 ± 0.25bc 7 3 1 3 1.80 ± 0.19a 1.93 ± 0.25a 1.33 ± 0.04a 2.25 ± 0.36a 8 3 2 1 1.81 ± 0.28a 1.92 ± 0.37a 1.34 ± 0.10a 2.29 ± 0.47a 9 3 3 2 1.70 ± 0.15a 1.63 ± 0.16ab 1.29 ± 0.05ab 1.90 ± 0.22ab Different letters (a)–(d) indicate significant differences (p < 0.05) among different levels by Duncan's test. Floral-scent fingerprinting of 97 Clematis germplasm resources
-
Using the optimized condition, we measured the floral-scent signals of 97 Clematis germplasm resources from 11 horticultural groups and constructed an E-nose–based fragrance fingerprint. The total sensor responses across materials ranged from 1.01 to 16.33. W1W, W2W, and W5S consistently showed higher signals, whereas W1C, W2S, and W6S remained low (Fig. 2a). Because W1W/W2W are mainly sensitive to sulfides and organic sulfides, and W5S to alkanes and aromatic volatiles, this pattern suggests that sulfur-containing and aliphatic compounds are the dominant contributors to fragrance differences among Clematis cultivars based on sensor characteristics.
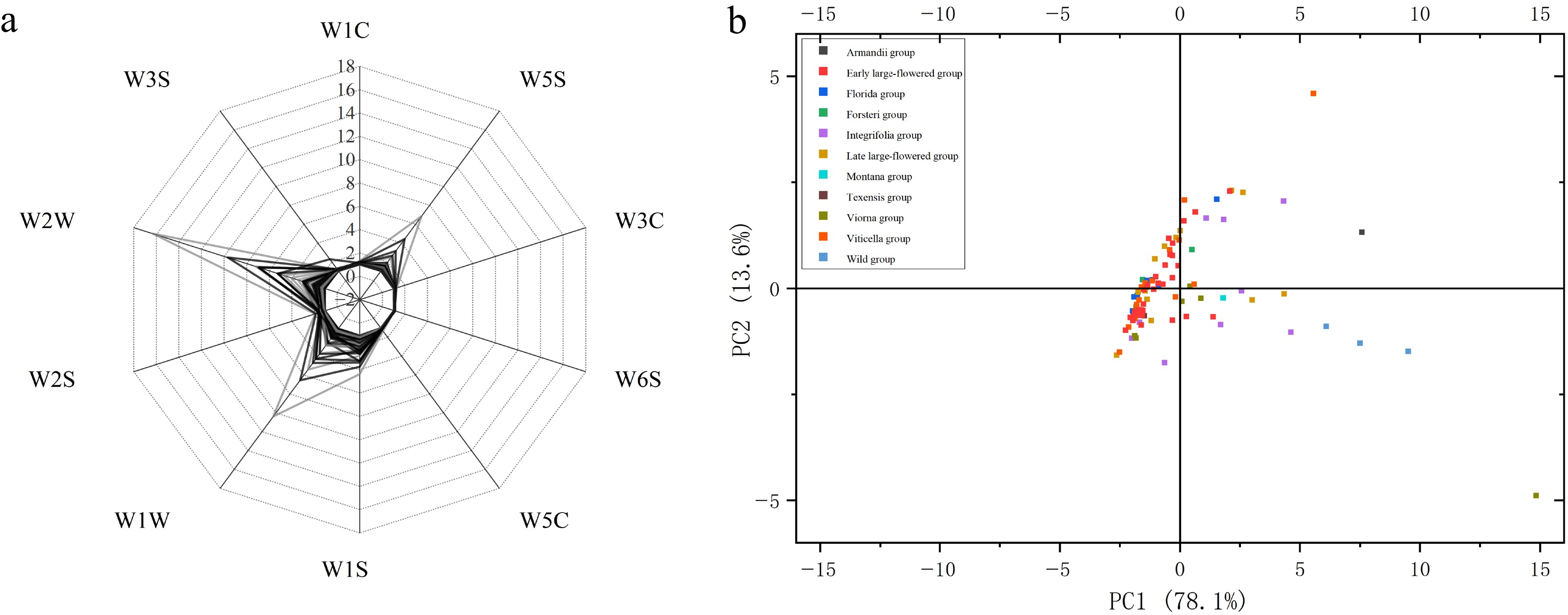
Figure 2.
(a) Fingerprint map construction, and (b) principal component analysis of floral fragrances from 97 Clematis varieties based on E-nose technology. The key groups were Viorna, Wild, Armandii, Integrifolia, and Late Large-Flowered groups.
PCA, based on the 10-sensor data showed that PC1 and PC2 explained 78.4% and 13.4% of the variance, respectively, for a cumulative 91.8% (Fig. 2b). Most groups (Early Large-Flowered, Montana, Late Large-Flowered, Wild, Viticella, Integrifolia, and Viorna) were clearly separated or loosely clustered, indicating that their fragrance profiles could be discriminated by the E-nose. According to their response ranges and PCA distribution, the 11 groups could be divided into three aroma-response types: (1) a high-response/complex type (Viorna, Wild, Armandii, Integrifolia, and Late Large-Flowered groups) with a very wide response interval (1.01–16.33) and a scattered distribution on PC1; (2) a medium-response/balanced type (Montana, Viticella, Early Large-Flowered groups) with moderate, relatively concentrated responses (about 1.02–5.35); and (3) a low-response/simple type (Forsteri, Florida and Texensis groups) with uniformly low values (1.02–1.53), and light or inconspicuous fragrance (Supplementary Fig. S1).
Screening of highly fragrant of Clematis germplasm resources
-
To identify materials with breeding value for scent, we compared germplasm resources within the major groups (≥3 samples). In the Viorna group, response values were 1.02–16.33, and 'Taigetsu' was clearly the strongest-scented cultivar (Fig. 3a, Supplementary Table S1). In the Wild group, the three native species showed 1.06–7.87, with C. cadmia and C. terniflora outperforming the third taxon (Fig. 3b). In the Integrifolia group (1.02–7.20), 'Rōguchi', 'Sizaia Ptitsa' and 'Hanajima' responded much higher than the others (Fig. 3c). In the Late Large-Flowered group (1.01–6.94), 'Comtesse de Bouchaud', 'Ernest Markham', and 'Lucky Charm' ranked at the top (Fig. 3d). In the Viticella group (1.10–5.24), 'Buckland Pixie', 'Betty Corning' and 'Dark Eyes' were stronger (Fig. 3e). The Early Large-Flowered group (34 cultivars) mostly showed low, clustered responses (1.02–4.08), with only 'Moonglow', 'Rosemoor', 'Viva Polonia' and 'Red Star' being relatively higher (Fig. 3f). By contrast, the Florida group (nine cultivars) and the Texensis group (three cultivars) showed only weak signals (about 1.02–2.23 and 1.03–1.53, respectively), consistent with faint fragrance (Fig. 3g, h).
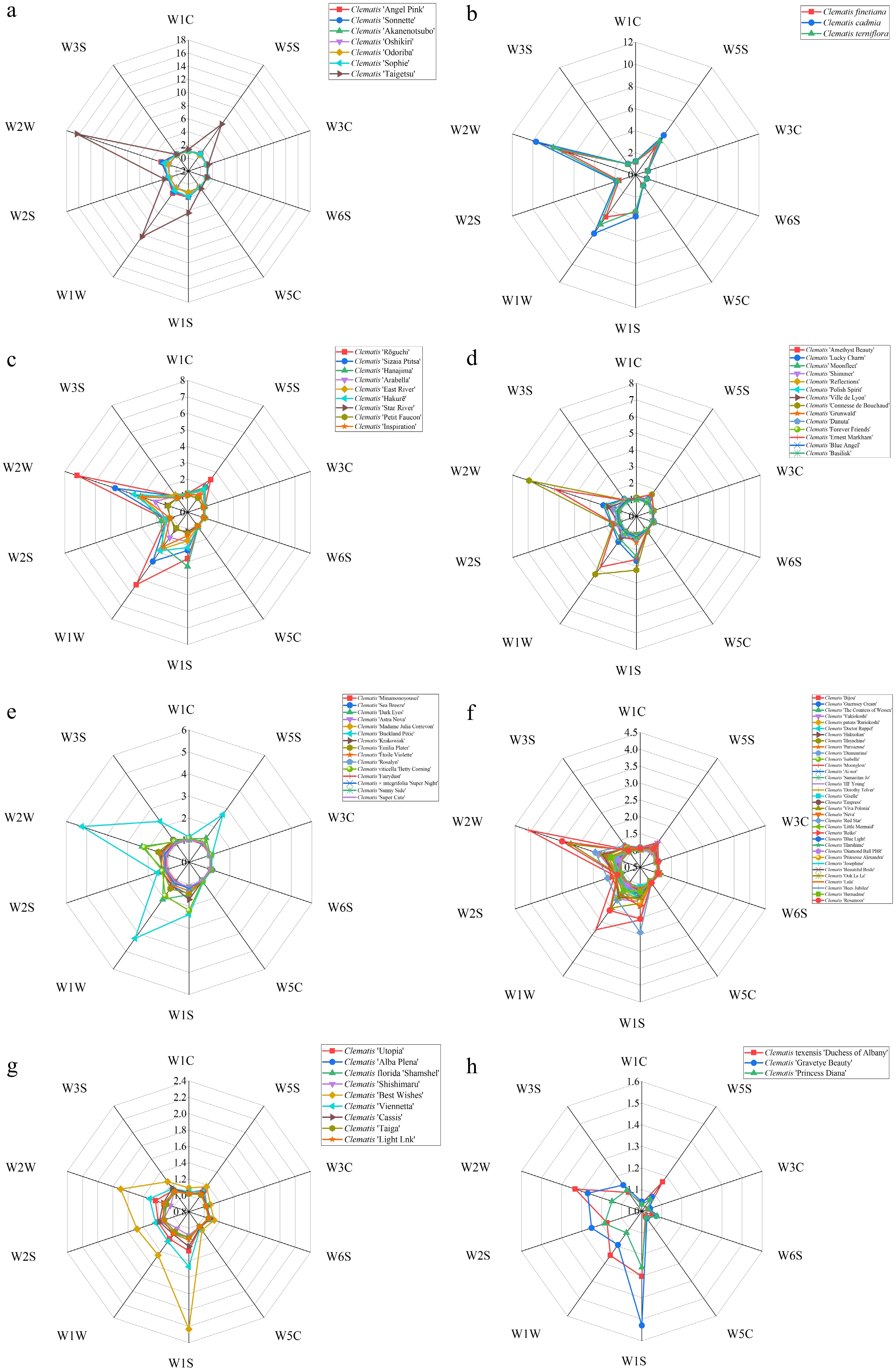
Figure 3.
Radar charts of floral fragrance responses for eight groups of Clematis. (a) Viorna group; (b) Wild group; (c) Integrifolia group; (d) Late Large-Flowered group; (e) Viticella group; (f) Early Large-Flowered group; (g) Florida group; and (h) Texensis group.
Integrating the 10-sensor signals of all 97 samples, we finally identified 15 Clematis germplasm resources with outstanding aroma performance. Ranked from highest to lowest total response, they were: 'Taigetsu', C. cadmia, C. terniflora, C. finetiana, 'Apple Blossom', 'Rōguchi', 'Comtesse de Bouchaud', 'Buckland Pixie', 'Ernest Markham', 'Sizaia Ptitsa', 'Hanajima', 'Moonglow', 'Rubens Superba', 'Hakurē', and 'East River' (as listed in the original dataset). All of these elite materials showed strong signals on W1W, W2W, and W5S, supporting the conclusion that Clematis floral scent is mainly characterized by sulfide- and alkane-related volatiles. Overall, most cultivated Clematis in this collection emitted only low to moderate levels of fragrance, highlighting the breeding value of the few highly scented accessions identified here.
-
Fragrance is an important yet underexplored ornamental trait in Clematis, and it has clear value for future aroma-oriented breeding. In this study, we established the first rapid E-nose-based detection system for Clematis floral scent to our knowledge and applied it to 97 germplasm resources from 11 horticultural groups. The results demonstrate that an array of 10 metal-oxide sensors can capture the integrated volatile profile of Clematis flowers quickly and non-destructively, and that the resolution is sufficient to distinguish both fragrance intensity and aroma type among groups. This confirms that E-nose technology is a feasible, high-throughput alternative to labor-intensive GC-MS when the goal is germplasm screening or pre-breeding evaluation.
The orthogonal test showed that flowering stage was the dominant factor affecting E-nose responses, followed by headspace equilibration time and sample temperature (A > C > B). Consistent with the research results of cherry blossoms (flowering period > headspace equilibrium time)[26]. The optimal signal was obtained at the late full flowering stage, after petals were fully expanded and pollen was dehiscing. This suggests that Clematis releases the highest amount of volatiles toward the end of the display period. A similar pattern has been reported in other ornamentals, where visual cues dominate at anthesis, while scent emission increases later to maintain pollinator attraction as floral color or turgor declines. This is consistent with Raguso's view that plants can shift from visual to olfactory signaling when pollinator return rates fall[28]. The release of floral aroma in lilies[27] and gardenia[29] mainly occurs during the peak flowering period, which may be related to the fact that they did not set the end of the peak flowering period, or it may be due to differences between species. Under our conditions, equilibration at 20 °C for 6 h allowed volatiles to diffuse sufficiently in the headspace without heat-induced loss of aroma compounds, and therefore the A3B1C3 combination (late full flowering stage × 20 °C × 6 h) was selected as the recommended parameter set.
The 10 sensors differed markedly in their usefulness for Clematis[30]. W1W, W2W, W5S, and W1S contributed most to PC1 and showed the highest discrimination, whereas W1C, W3C, and W5C had low sensitivity to Clematis volatiles and added little to classification. Because W1W and W2W are sensitive to aromatic/organic sulfides and W5S and W1S to alkanes and methane, the data indicate that sulfide- and alkane-type volatiles are the key components underlying fragrance differentiation in Clematis based on sensor characteristics. This agrees with reports in cherry blossoms and grape hyacinth that MOS sensors targeting sulfides, alcohols/ethanol, and low-carbon alkanes capture a large share of floral scent variation[26,31]. It also means that, for future large-scale screening, the sensor set could be simplified around these four high-information channels to reduce redundancy and improve throughput. The correlation between E-nose response signals and specific volatile compounds still requires verification by chemical methods
According to the characterization of aroma-active volatiles in three Chinese bayberry (Myrica rubra) cultivars, the E-nose could be used as a quick analysis tool that can be used to distinguish between samples from different cultivars by measuring the concentration of volatile compounds. And, the findings were in good agreement with the results obtained by GC-MS-O as they both separated the samples into the different groups. This confirmed the usefulness of the E-nose as a bayberry fruit identification tool[32]. In addition, the volatile compounds analysis of Tremella aurantialba fermentation via E-nose and HS-SPME-GC-MS showed that the results of PCA by GC-MS were more similar with that of the E-nose[33]. Therefore, the E-nose combined has a high potential to quickly distinguish and classify different cultivars.
PCA of the 97 samples revealed clear group-level patterns, and on that basis we proposed an E-nose fragrance classification for Clematis: (1) a high-response, complex type including the Viorna group, Wild group, Armandii group, Integrifolia group and Late Large-Flowered group, which showed wide response ranges and richer volatile signals; (2) a medium-response, balanced type including the Montana, Viticella and Early Large-Flowered groups, which showed moderate but stable responses; and (3) a low-response, simple type including the Forsteri, Florida and Texensis groups, which were weakly scented and poorly separable. The Forsteri group and Armandii group include only one cultivar; the low within-group replication may influence the fragrance classification for Clematis. Within these, 15 accessions—such as 'Taigetsu', C. cadmia, C. terniflora, C. finetiana, 'Apple Blossom', 'Rōguchi', and 'Comtesse de Bouchaud'—consistently triggered high responses on W1W, W2W, and W5S, and can therefore be regarded as elite fragrance donors for breeding. Among them, only three accessions are large-flowered types, while most of them with great floral fragrance are small-flowered types. Compared to large-flowered types, the high fragrance of small-flowered types enhances their attractiveness to pollinators. While E-nose fingerprinting is rapid, the sensor array responds to total ionisable mass rather than to specific odorants. In the next step of the research, we will analyse the key aroma compounds of 15 Clematis germplasm resources with outstanding fragrance performance by GC-MS technology.
-
This work establishes a practical, standardized E-nose protocol for Clematis floral-scent evaluation, identifies the key volatile categories perceived by the sensor array (sulfides and alkanes) based on sensor characteristics, and provides an aroma-based classification of 11 horticultural groups. By rapidly pinpointing 15 strongly scented germplasm resources, the study offers direct material support and technical guidance for breeding Clematis with improved fragrance, and it shows that E-nose-based fingerprinting can be extended to aroma differentiation in other ornamental species.
-
The authors confirm their contributions to the paper as follows: resources, project administration and writing – original draft: Liu H, Song W; data curation and software: Xu W, Xu M; methodology: Mao X; investigation and validation: Chen R; conceptualization and writing – review & editing: Yuan M. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed in the current study are available from the corresponding author on reasonable request.
-
This work was supported by the Technology Plan Project of the Jiangsu Vocational College of Agriculture and Forestry (2022kj07), and the Revitalization of Seed Industry Project of Jiangsu Province (JBGS[2021]092).
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/opr-0026-0011.
- Supplementary Table S1 Ranges of floral fragrance response values for eleven groups of Clematis.
- Supplementary Fig. S1 Ranges of floral fragrance response values for 11 groups of Clematis.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu H, Xu W, Xu M, Mao X, Chen R, et al. 2026. Establishment of a determination method and screening of aromatic germplasm in Clematis L. using electronic nose technology. Ornamental Plant Research 6: e022 doi: 10.48130/opr-0026-0011 

Establishment of a determination method and screening of aromatic germplasm in Clematis L. using electronic nose technology
- Received: 06 November 2025
- Revised: 09 April 2026
- Accepted: 17 April 2026
- Published online: 05 June 2026
Abstract: Fragrance is an important ornamental trait in Clematis (Ranunculaceae), yet rapid and standardized evaluation methods are still lacking. In this study, we established an electronic nose (E-nose)-based protocol for efficient fragrance detection in Clematis and characterized the volatile response profiles across diverse germplasm. Using an orthogonal experimental design, we optimized E-nose operating conditions by analysis of variance (ANOVA) and multiple comparisons. Three factors significantly affected sensor responses: flowering stage, headspace equilibration time, and sample temperature. The optimal detection combination was A3B1C3, i.e., sampling at the late full flowering stage, and equilibrating for 6 h at 20 °C. Among the sensors, W2W, W1W, and W1S showed significantly higher response values than the others (p < 0.05). It indicated that Clematis floral scent is likely dominated by aromatic sulfides, alkanes, and other S-containing compounds. We then applied the optimized system to 97 Clematis germplasm resources from 11 horticultural groups. They were classified into three E-nose fragrance types: (1) high-response/complex type; (2) medium-response/balanced type; and (3) low-response/simple type. Finally, 15 germplasm resources with outstanding fragrance performance were identified. This work establishes a rapid, reproducible, E-nose detection system for Clematis floral scent and provides a practical tool for early selection of fragrant parents in breeding. The fragrance fingerprints generated here further suggest that sulfides and alkanes are key discriminating volatiles in Clematis based on sensor characteristics, supporting both cultivar classification and aroma-oriented breeding.
-
Key words:
- Clematis /
- Floral scent /
- Scent-oriented breeding