








-
Soil heterogeneity refers to the nonuniform distribution of soil resources (e.g., water and nutrients), which significantly affects plant dynamics such as seed germination[1−4]. Germination rates have been shown to change when the soil heterogeneity is different[4,5], which, in turn, affects species' coexistence in plant communities[6]. Previous studies found that higher environmental heterogeneity could improve plants' coexistence through increasing spatial niche partitioning[7] and/or species sorting among heterogeneous niches[8]. Soil heterogeneity may increase in the future through the effects of both climate change and of human disturbance[9]. Thus, it is important to improve our understanding of the relationship between plants' coexistence and their responses to soil heterogeneity. However, exploring the effects of soil heterogeneity on seed germination under natural conditions is complex, because co-occurring factors such as soil legacies[10] and climate change[11] make it difficult to separate the effects of soil heterogeneity from those of other factors. Thus, experimental research is necessary to establish the cause-and-effect relationships. However, soil heterogeneity in many studies is only considered in two dimensions (i.e., either horizontally or vertically)[2,12], although natural soils vary both horizontally and vertically (i.e., in three dimensions)[5,13]. As two-dimensional approaches may obscure patterns that would appear in the field, here, we simulate soil heterogeneity in three dimensions.
Differences between soil patches can give rise to changes in plant growth. In an earlier experiment, perennial grasses tended to grow better on resource-rich patches compared with annual species because of stronger competitiveness of the former, whereas the opposite pattern was observed on resource-poor patches[4,14]. Specifically for seed germination, some plant species release allelochemicals to hinder the germination of the seeds of other coexisting species[15−17], or can enhance their own seed germination by associating with some types of microbes such as mycorrhizal fungi in mutualisms[18]. Moreover, the microclimate (especially temperature and moisture) also significantly affects germination[4]. These factors may change with soil heterogeneity, for example, because the microbial communities[18] or soil moisture[4] differ between soil patches.
Seed germination is the first step in plants' establishment[19,20], and differences in germination-stage responses influence the seasonal and competitive environments experienced by plants[21,22]. Environmental and biological factors collectively contribute to divergent responses of seeds during germination (i.e., different species exhibit distinct requirements for temperature and soil moisture during germination)[23], and plant competition may favor early germination to gain a competitive advantage[24]. Soil heterogeneity and species composition can significantly affect plant biomass[5], but studies focusing solely on biomass production cannot directly elucidate the effects of soil heterogeneity and species composition on the early plant establishment process. As the coexistence of species in a plant community depends on both germination and plant growth dynamics, it is important to establish the exact effects of soil heterogeneity and species composition on seed germination in plant communities.
To explore the effect of species composition and soil heterogeneity on seed germination of plant species, a controlled experiment was performed in which seven species compositions, derived from all the possible combinations of three forage grasses (i.e., Festuca elata, Bromus inermis, and Elymus breviaristatus) were crossed with four levels of soil heterogeneity. These soil heterogeneity levels were created by filling pots with two substrates (the resource-rich and resource-poor substrate) in an alternating pattern to simulate fine, small, medium, or large patch sizes (Fig. 1a, b). Specifically, we tested two hypotheses. (1) At the pot scale (whole system), seed germination is assumed to increase with increasing patch size, as plants tend to reduce germination to mitigate negative influences induced by strong variation in the soil conditions (including the microenvironment)[5], which we expect is likely to be more variable in pots with relatively smaller patch sizes[4]; we also expect this pattern to be modified by species composition (Fig. 1c)[5,25,26]. (2) At the substrate scale (within-pot patches), the positive association between patch size and germination rate (Hypothesis 1) is mainly observed on the resource-rich substrate because of the relatively better conditions for seed germination (e.g., higher soil moisture and temperature buffering) (Fig. 1d)[14].
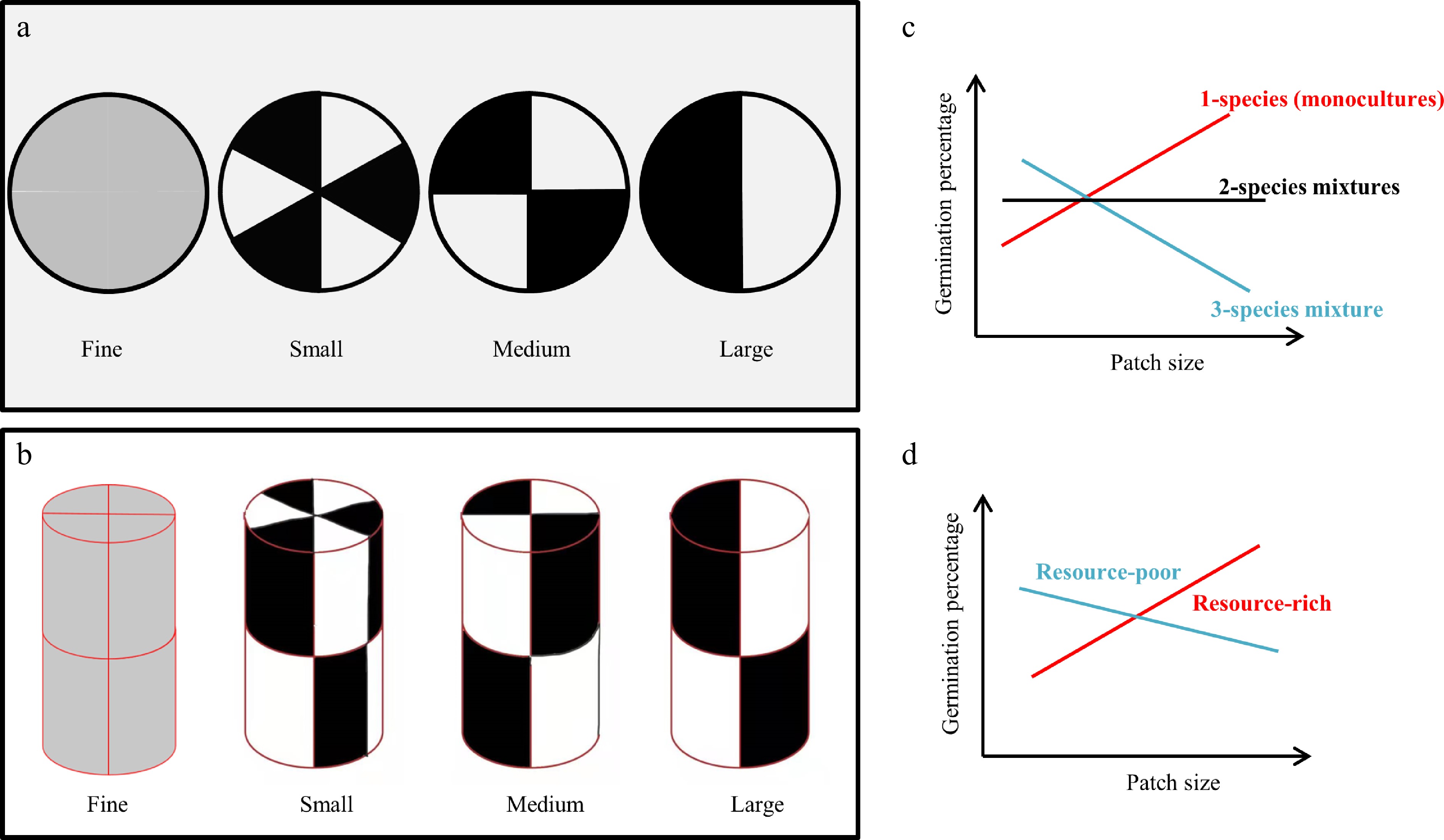
Figure 1.
Four soil heterogeneity levels were applied in this experiment, where each level was created by alternatively filling pots with resource-rich (black) and resource-poor (white) substrates, giving rise to fine, small, medium, and large patch sizes: (a) Top view; (b) lateral view. (c) Hypothesis 1 (at the pot scale): Seed germination is expected to increase with increasing patch size, and this pattern is supposed to be modified by species composition. (d) Hypothesis 2 (at the substrate scale): seed germination is hypothesized to increase in the resource-rich substrate with increasing patch size, whereas the opposite pattern is predicted to be found on the resource-poor substrate. Note that the one-species (monoculture), two-species mixtures, and three-species mixture in (c) could be any of the target species.
-
A controlled experiment was performed in a greenhouse at the Linze Pratacultural Research Station of Lanzhou University, China (39°14′30″ N, 100°3′25″ E) from May 15 to September 14, 2021. The station is characterized by a temperate continental arid monsoon climate, with a mean annual temperature and mean annual precipitation of 7.79 °C and 110.7 mm, respectively[27].
Soil heterogeneity in this study was experimentally created via the method developed by Liu et al.[13] in three steps. (1) A pot was vertically separated into two layers, and the space in each layer was divided into patches with equal size by placing plastic plates with slits in each layer. (2) These patches were filled with two substrates (i.e., the resource-rich and resource-poor substrate; Table 1) in an alternating pattern. (3) The plastic plates were subsequently removed, allowing plant roots to grow freely into all patches. It is worth mentioning that similar compression was applied when filling the substrates in each pot to ensure that the amount of soil resources was identical in all pots. In this experiment, soil heterogeneity was modified by varying the patch size (Fig. 1). The resource-poor and resource-rich substrates used in this experiment were created by mixing soil and sand in a ratio of 1:4 and 4:1, respectively. The sand was bought from a locally commercial supplier; the soil was collected from the local crop field. Characteristics of the resource-rich and resource-poor substrates (Table 2) were tested at the beginning of the experiment.
Table 1. Soil pH, electrical conductivity (EC), soil organic carbon (SOC), available phosphorus (AP), and total nitrogen of the resource-rich and resource-poor substrate at the beginning of the experiment.
Substrate pH EC (μS·cm−1) SOC (g·kg−1) AP (mg·kg−1) Total nitrogen (g·kg−1) Resource-rich 7.1 ± 0.02 1,469 ± 38 7.52 ± 0.15 4.04 ± 0.09 0.14 ± 0.01 Resource-poor 7.2 ± 0.07 1,120 ± 102 6.73 ± 0.25 3.36 ± 0.06 0.05 ± 0.01 Note that three replicates were used in these tests, and the values in this table are shown as the mean ± standard error (SE). Table 2. Effects of patch size (fine, small, medium, and large), species composition, and their interaction on seed germination percentage tested by a generalized linear mixed model at the pot scale.
df F p Patch size 3, 56 4.3 0.009 Species composition 6, 56 12.7 < 0.001 Patch size × species composition 18, 56 3.6 < 0.001 F-values, p-values, and degrees of freedom (dfbetween-groups, dfwithin-groups) are given, with significant results (p < 0.05) in bold. Identical pots (28.5 cm top diameter, 20.0 cm bottom diameter, 22.0 cm height) were used in this study, and they were separated into four groups by patch size (fine, small, medium, or large) (Fig. 1a, b). The pot size was based on the previous studies such as Liu et al.[14], and Liu and Hou[4]. The only difference among these pots was the patch size, with the fraction of resource-rich and resource-poor substrate always being 50% each. Six holes 10 mm in diameter were made in the central bottom of each pot to enable drainage and avoid standing water. Soil heterogeneity in this experiment was considered to increase with decreasing patch size, since the soil conditions vary more frequently (spatially) in pots with a smaller patch size[13,14,28].
We focused on three perennial grasses (i.e., F. elata, B. inermis, and E. breviaristatus) commonly used in cultivated grasslands and the restoration of degraded grasslands. Seeds used in this study were bought from a commercial company (Best, Beijing, China). Seven species compositions were created by using all the possible combinations of the three target species (i.e., F. elata, B. inermis, E. breviaristatus, F. elata + B. inermis, F. elata + E. breviaristatus, B. inermis + E. breviaristatus, and F. elata + B. inermis + E. breviaristatus). Thus, our experiment had 28 treatments (four levels of soil heterogeneity × seven species compositions), resulting in 84 pots in total with three replicates for each unique treatment. Seed density was based on our previous studies[13,28]. At the beginning of the experiment, all the seeds (in the appropriate proportion per species) were thoroughly mixed and evenly separated into two halves. Each half was randomly sown in each of the two substrates in each pot. A few millimetres of substrate were subsequently added to maintain soil moisture and temperature. During the experiment, water was added to each pot when necessary to avoid drought. All pots were randomly placed in a greenhouse.
Germinated seeds of each species were counted daily when they could be discerned as separate species, and the experiment was ended when no more new seeds had germinated for three consecutive days. Germinated seeds were counted separately in the resource-rich and resource-poor substrate in each pot. To quantify the seed germination percentage, we considered the amount of seeds sown; in the resource-rich (or resource-poor) substrate, this was half of the total number of seeds added in each pot. The data from the last record were used to calculate the seed germination percentage.
Data analysis and statistics
-
To test the first hypothesis, an analysis was performed with a generalized linear mixed model (GLMM) focusing on the pot scale (1), in which the effects of patch size (i.e., fine, small, medium, and large), species composition, and their interaction on seed germination percentage were explored, with pot identity as a random factor. To examine the second hypothesis, we used GLMMs at the substrate scale (2), investigating the effects of patch size (i.e., small, medium, and large), species composition, substrate, and their interactions on seed germination percentage, again with pot identity as a random factor. The fine patch size was not included in this second (patch-based) analysis, since the two substrates could not be accurately separated at this scale. Moreover, separate analyses were conducted for the resource-rich and resource-poor substrate (3), using GLMMs to study the effect of patch size, species composition, and their interaction on the seed germination percentage in the resource-rich and resource-poor substrate, with pot identity as a random factor. In all three analyses, post-hoc analyses (pairwise comparisons with least significant difference [LSD]) were performed to test the differences among the variables. To account for the increased Type I error risk with multiple comparisons, we also applied Tukey corrections, which resulted in identical outcomes regarding significant and nonsignificant differences, underlining the robustness of our findings. All statistical analyses were performed in SPSS 23.0 (IBM Corp., 2015).
-
In the first analysis, patch size, species composition, and their interaction all significantly affected the seed germination percentage (Fig. 2; Table 2). Specifically, the small patch size supported a higher seed germination percentage than the other patch sizes in the mixture with three species growing together (Fig. 2), and a similar pattern was found for the mixture of F. elata + B. inermis. However, a different pattern was found for the mixture of B. inermis + E. breviaristatus, where the fine and large patch sizes had higher seed germination percentages than the small and medium patch sizes. Such effects were not apparent in the mixture of F. elata + E. breviaristatus, where patch size had no significant effect on the seed germination percentage (Fig. 2). All these indicate the complex effects of patch size and species composition on seed germination.
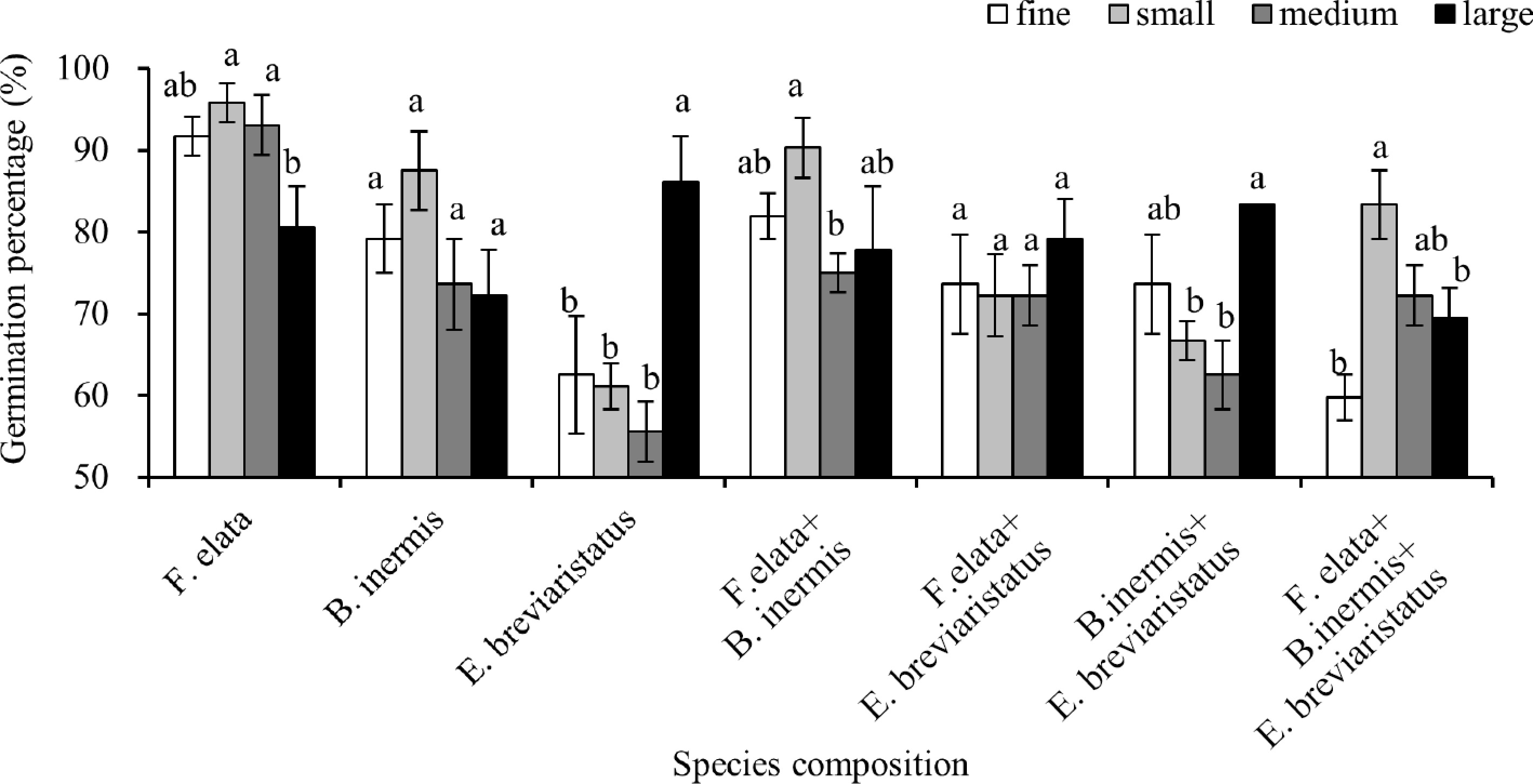
Figure 2.
Mean ± SE of seed germination percentage of the seven species compositions shown at the pot scale, separated by patch sizes (soil heterogeneity), i.e., fine, small, medium, and large. Significant differences between patch sizes for each species composition are labeled by different letters (post hoc analysis with LSD).
In the second analysis, patch size, species composition, and their interaction significantly affected seed germination percentage (Table 3). The substrate did not significantly affect the seed germination percentage as a single factor, but a marginally significant effect of patch size × species composition × substrate was found (Table 3).
Table 3. Effects of patch size (small, medium, and large), species composition, substrate, and their interactions on seed germination percentage were explored via GLMM at the substrate scale, separated by substrate.
df F p Patch size 2, 92 5.3 0.007 Species composition 6, 92 7.3 < 0.001 Substrate 1, 92 2.3 0.134 Patch size × species composition 8, 92 4.3 < 0.001 Patch size × substrate 2, 92 1.9 0.160 Species composition × substrate 6, 92 1.2 0.312 Patch size × species
composition × substrate8, 92 1.8 0.064 F-values, p-values, and degrees of freedom (dfbetween-groups, dfwithin-groups) are given, with significant results (p < 0.05) in bold. In the third analysis, a significant interactive effect of patch size and species composition was found in both the resource-rich and resource-poor substrates (Table 4). As a single factor, patch size and species composition showed different patterns in resource-rich and resource-poor substrates (Table 4). Specifically, the small patch size supported a higher seed germination percentage than the medium and large patch sizes with the three-species mixture grown on a resource-rich patch (Fig. 3a), but this effect disappeared in the resource-poor substrate (Fig. 3b).
Table 4. The effects of patch size, species composition, and their interaction on seed germination percentage investigated via GLMM.
Resource-rich Resource-poor df F p df F p Patch size 2, 46 4.6 0.015 2, 46 2.8 0.070 Species composition 6, 46 2.3 0.053 6, 46 5.9 < 0.001 Patch size × species composition 8, 46 2.4 0.016 8, 46 3.4 0.001 F-values, p-values, and degrees of freedom (dfbetween-groups, dfwithin-groups) are given, with significant results (p < 0.05) in bold. 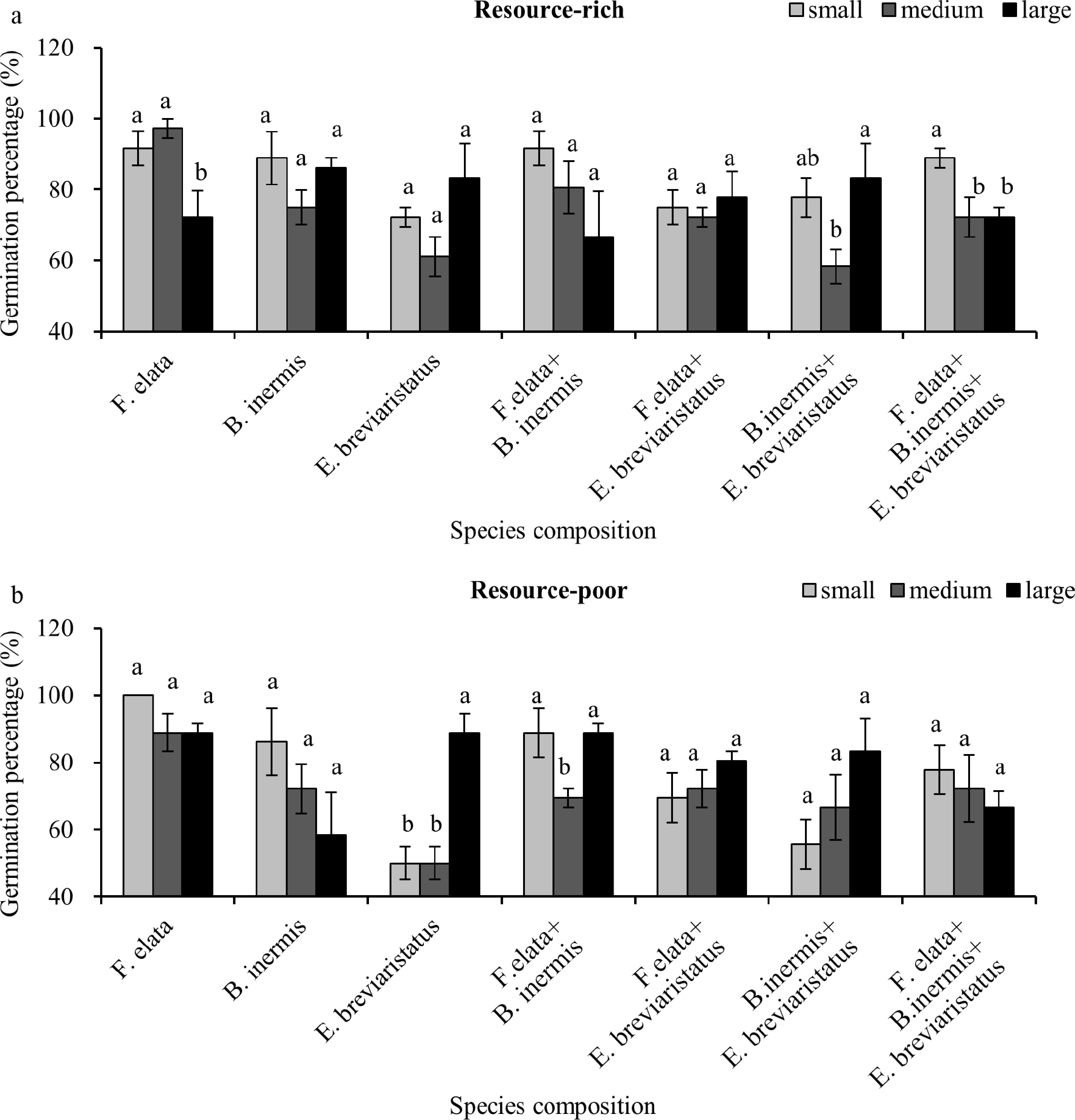
Figure 3.
Mean ± SE of seed germination percentage of the seven species compositions on (a) the resource-rich and (b) the resource-poor substrate, shown at the substrate scale and separated by patch size (soil heterogeneity; i.e., small, medium, and large). Significant differences between patch sizes for each species composition are labeled by different letters (post hoc analysis with LSD).
-
The first hypothesis was that the seed germination of plant species was interactively affected by species composition and soil heterogeneity. This was supported, since we recorded significant interactive effects of patch size (i.e., soil heterogeneity) and species composition on the seed germination percentage (Table 2). Moreover, a higher germination percentage was generally found in pots with small patches than in the pots with the other patch sizes, a pattern that was found in four of the seven species compositions (Fig. 2). This differs from earlier observations in monocultures[4], where seed germination was higher in pots with large patches than in pots with small patches. One possible reason may be that when multiple species germinate, they can affect each other's germination success, through, for example, association with specific microbes or through allelochemicals[17,18]. Previous studies found that the seed germination of a species can be either be impeded by the allelochemicals released by the neighboring species through inhibition or promotion of radicle and hypocotyl elongation[17]. In two of the seven communities (i.e., E. breviaristatus and B. inermis + E. breviaristatus), a higher seed germination percentage was observed in pots with large patches (Fig. 2). This is consistent with the finding of Maestre and Reynolds[29], where seed germination percentage decreased with increasing distance to a resource-rich patch. The varied results in our study highlight the need for further investigation into the exact mechanisms that govern seed germination in differing circumstances (soil heterogeneity and species composition in this case).
Our second hypothesis proposed that any increases in germination with patch size at the pot scale (the whole system) would mainly be derived from the resource-rich substrate but not the resource-poor substrate. This hypothesis was not supported, even though different germination patterns were observed on the resource-rich and resource-poor substrates. Seed germination percentage first decreased and then increased with increasing patch size in communities of B. inermis + E. breviaristatus growing on the resource-rich substrate (Fig. 3a), and in F. elata + B. inermis communities growing on the resource-poor substrate (Fig. 3b). Reasons for this pattern may be linked to allelochemicals affecting seed germination via processes such as inhibiting the growth of the plant embryo[17], whereas microbial association could drive seed germination differentially if their activity differs in resource-rich and resource-poor substrates[26,30]. However, these mechanisms remain speculative, as allelochemicals, microbial communities, and microclimatic variation were not directly quantified.
The results of this study should be extrapolated and interpreted with caution because of the following reasons. (1) The origin of the seeds (wild or commercial) may affect germination[31,32]. Seeds used in this study were bought from a seed company, but wild seeds may perform differently[4]. (2) Seed germination was only tested for mixtures of three forage grasses, and more species originating from different functional groups should be considered, since plant communities often feature a more diverse species mix[10]. (3) Allelochemicals and microbial associations were not measured in our study[5], which prevents us from exploring the underlying mechanisms pertaining to seed germination in more detail. In our study, the results from the substrate scale could not be extrapolated additively to the pot scale. Interactive and nonadditive effects are common in ecology, necessitating further research into the underlying mechanisms to enable extrapolation through modeling. Furthermore, extrapolation to natural environments (e.g., for restoration applications) will require further studies at larger scales, incorporating more sources of naturally occurring heterogeneity (seeds, species, soils, etc.).
-
The authors confirm their contributions to the paper as follows: study design: Liu Y; experiment, data collection, and analyses: Li H, Pang S, Ning Y, Yang Q, Zhu Q, Liu Y; manuscript drafting: Liu Y, Ying Z, De Boeck HJ. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
We thank Mingxia Wang, Guoe Li, Cheng Zhang, Wanhe Zhu, Weiqi Jiang, and Lijuan Ran for experimental assistance. This work was supported by the Longyuan Youth Talent Project of Gansu Province and the Fundamental Research Funds for the Central University (lzujbky-2025-14).
-
The authors declare no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li H, Pang S, Ning Y, Yang Q, Zhu Q, et al. 2026. Soil heterogeneity and species composition interactively affect the seed germination of three typical forage species. Grass Research 6: e018 doi: 10.48130/grares-0026-0009 

Soil heterogeneity and species composition interactively affect the seed germination of three typical forage species
- Received: 15 December 2025
- Revised: 20 March 2026
- Accepted: 27 March 2026
- Published online: 15 June 2026
Abstract: Soil heterogeneity has been shown to affect seed germination of plant species in monocultures, and plant species' interactions in a community may also impact seed germination. However, whether and how both factors interact to modulate seed germination is still unclear. To investigate these interactive effects, a controlled experiment was conducted in which seven species compositions derived from all the possible combinations of three forage grasses (Festuca elata, Bromus inermis, and Elymus breviaristatus) were factorially crossed with four levels of soil heterogeneity. This heterogeneity gradient was developed by applying resource-rich and resource-poor substrates in an alternating pattern in pots, creating fine, small, medium, and large patch sizes. The results showed that patch size and species composition interactively affected seed germination percentage. Specifically, the small patch size supported a higher seed germination percentage in the mixture with three species compared with other patch sizes (i.e., fine, medium, and large patch sizes). Moreover, this pattern was only found in the resource-rich substrate, not in the resource-poor substrate. A combination of abiotic (e.g., soil moisture and temperature) and biotic effects (e.g., allelopathic suppression) may have caused these varying patterns. Designs such as ours that vary soil heterogeneity in three dimensions should be combined with additional measurements of the underlying mechanisms to further elucidate the factors governing seed germination.
-
Key words:
- Bromus inermis /
- Elymus breviaristatus /
- Festuca elata /
- Patch size /
- Plant community /
- Soil heterogeneity /
- Seed germination