










-
Seed size is a fundamental agronomic trait that determines the crop yield potential and is governed by the coordinated growth and development of the pericarp, embryo, and endosperm[1]. The endosperm is the primary nutrient storage organ in wheat, constituting over 80% of the seed's dry weight in a fully developed mature wheat seed,[2] and is therefore the major sink for starch and storage proteins[3]. The degree of endosperm development determines the size and weight of wheat grains[4], directly affecting their yield and quality. Endosperm development in cereals follows the nuclear type of endosperm development, where the initial endosperm nuclei divide without immediate cytokinesis, forming a coenocyte (early endosperm)[5]. As development progresses, the endosperm enters the cellularization stage, where the endosperm nuclei are partitioned by newly formed cell walls to create cellularized endosperm cells[6,7]. Cellularization largely fixes the internal volume of the grain, so its timing strongly influences the final grain size[8−10].
Auxin acts as a developmental timer that governs the switch from the coenocytic (syncytial) endosperm to the cellularized endosperm stage. High auxin concentrations in the coenocytic endosperm are sustained by polar export to the surrounding pericarp; when auxin falls below a threshold, cellularization is triggered[11]. Flavonoids have been reported to regulate auxin transport or local concentration, affecting the timing of endosperm cellularization[12−14]. For instance, kaempferol and quercetin modulate auxin efflux by inhibiting PIN and P-glycoprotein (PGP) transporters and their kinase regulator PINOID (PID)[15−18]. Genetic studies in Arabidopsis corroborate this link: the tt4 mutant (defective in Chalcone synthase, CHS) displays elevated polar auxin transport, whereas overexpression of TT3 (DIHYDROFLAVONOL 4-REDUCTASE, DFR) or TT7 (FLAVONOID 3ʹ-HYDROXYLASE, F3ʹH) reduces it[19−22]. Because both flavonol and auxin pathways branch from phenylalanine, feedback between their metabolic nodes may fine-tune auxin distribution[21−24]. Yet, it remains unclear how this metabolic–hormonal interaction governs the timing of endosperm cellularization and, by extension, grain size in wheat.
We previously characterized TaPGS1 (Triticum aestivum Positive Regulator of Grain Size 1), a IIIf-clade bHLH transcription factor that is homologous to Arabidopsis TT8 (TRANSPARENT TESTA 8) and whose bHLH core domain is most similar to rice OsRc (Oryza sativa, red pericarp gene). While TT8 and OsRc influence pericarp pigmentation and dormancy[13,14], TaPGS1 overexpression enlarges wheat grains without altering the pericarp color and is accompanied by increased expression of flavonoid-pathway genes[25]. Beyond their developmental roles, flavonoids also consist of nutritional and functional quality components, with compounds such as kaempferol and quercetin contributing to the antioxidant capacity and potential health benefits. These observations suggest that TaPGS1 may modulate the flavonoid pathway to regulate auxin dynamics and cellularization, but the underlying mechanism remains unknown.
Here, we integrate the transcriptomic and metabolomic profiling of TaPGS1 overexpression lines to identify differentially expressed genes and metabolites in the flavonol pathway. We visualize flavonol and auxin localization in developing seeds, assess the effect of exogenous kaempferol on cellularization, and test direct TaPGS1 targets using dual-luciferase (LUC), yeast two-hybrid, and bimolecular fluorescence complementation (BiFC) assays. Our study uncovers a TaPGS1-centered regulatory module that links flavonol accumulation to auxin-mediated control of endosperm cellularization, providing a mechanistic insight into the genetic improvement of wheat grain size.
-
We used common wheat (Triticum aestivum) cultivar Fielder and two TaPGS1 overexpression lines (OE-166-6 and OE-166-39) in the Fielder background as study materials. Wild-type Fielder and the overexpression lines were grown at Wenjiang, China (30°72′ N, 103°87′ E). Field traits, plant height, spike length, tiller number, effective spike number, and grain number per spike were recorded. Spikes at the soft dough stage were harvested, air-dried, and manually threshed, and grain moisture was adjusted to ~10% with a near-infrared analyzer (DA 7250, PerkinElmer). For germination tests, fifty seeds per line (three replicates) were sown on moist filter paper, germination was scored daily for 7 d, and germination percentage was calculated. Mature seeds were harvested from the main spikes of five independent plants per line and naturally dried to ~10% moisture. Seed length, width, and thousand-grain weight were determined based on >100 seeds per replicate. Other wheat plants were grown in a greenhouse under a 16-h light / 8-h dark photoperiod with automated climate control (day/night setpoints 20/15 °C, with heating/cooling to maintain conditions near the setpoints; 55%–60% relative humidity). Nicotiana benthamiana plants were cultivated in a growth chamber under the same photoperiod at 25 °C.
RNA-seq differential expression analysis and quantitative real-time polymerase chain reaction
-
Total RNA was extracted from developing seeds of TaPGS1 overexpression lines and wild-type Fielder grown in a greenhouse at 5, 10, 15, and 20 d post-anthesis (DPA). Strand-specific complementary DNA (cDNA) libraries were constructed using the standard Illumina protocol and sequenced on an Illumina HiSeq 2500 platform. Raw reads were quality trimmed with fastp (v1.0.1)[26]. Transcript-level quantification was performed with Salmon (v1.10.1)[27] in quasi-mapping mode against the IWGSC RefSeq v1.0 transcriptome. Gene-level counts were obtained using Tximport[28], and differential expression was tested with DESeq2[29]. Heatmaps and preliminary functional annotations were generated using GraphPad Prism v8.1.
For quantitative real-time polymerase chain reaction (qPCR), total RNA was reverse-transcribed using the PrimeScript 1st Strand cDNA Synthesis kit (Takara, Beijing, China). qPCR reactions were conducted with SYBR Premix Ex Taq II (Takara, Beijing, China) on a CFX96 Touch Real-Time PCR Detection System (Bio-Rad) under the following thermal profile: initial denaturation at 95 °C for 1 min, followed by 40 cycles of 95 °C for 5 s and 60 °C for 30 s. Relative transcript levels were normalized to internal reference genes, as described by Long et al.[30]. Primer sequences are listed in Supplementary Table S1.
Metabolomic analysis
-
Metabolomic profiling was carried out on 5-DPA seeds of TaPGS1 overexpression lines and wild-type Fielder. Crude extracts were analyzed by liquid chromatography-mass spectrometry (LC–MS), and raw data were processed in Compound Discoverer (CD) version 3.3 (Thermo Fisher Scientific) for peak detection, retention-time alignment, and feature deconvolution. Molecular features were matched against the mzCloud, mzVault, and Mass List databases, with a mass-accuracy window of ≤5 ppm to obtain putative metabolite identifications. Peak areas integrated in CD 3.3 were normalized to the total ion current (TIC) of each sample and used to represent the relative abundance of metabolites across samples. Differential dmetabolites were defined by |log2 fold change| ≥ 1 and false discovery rate < 0.05 and visualized as heatmaps in GraphPad Prism V8.1. Finally, the changes in flavonol–auxin pathway metabolites were compared with those caused by differentially expressed genes identified from the transcriptome to explore coordinated regulation at the metabolic and transcriptional levels.
Flavonol fluorescence staining localization
-
Fresh seeds at 5 DPA were hand-sectioned and incubated for 5 min at room temperature in diphenylboric acid 2-aminoethyl ester (DPBA) staining solution. After staining, the sections were rinsed twice with distilled water, mounted on microscope slides, and examined immediately with a fluorescence microscope (BX61, Olympus). DPBA staining followed the original NA/DPBA method[31] and its early Arabidopsis application[32], with practical parameters from Nguyen et al.[33]. Under this dye, kaempferol and quercetin exhibit predominantly green and yellow fluorescence in seeds. DPBA sections were prepared from the same 5-DPA seed batches used for the untargeted LC–MS analysis.
Auxin localization (immunohistochemistry)
-
Seeds of wild-type Fielder and TaPGS1 overexpression lines at 5 DPA were used for the immunolocalization of indole-3-acetic acid (IAA), following a whole-mount immunohistochemical method. Samples were vacuum-infiltrated with 1-ethyl-3-(3-dimethylaminopropyl)-carbodiimide (EDC) / N-hydroxysuccinimide (NHS) buffer containing 0.1% Tween-20. This step cross-links the carboxyl group of IAA to cellular proteins, preventing diffusion and ensuring the specificity of the subsequent immunological detection. They were then fixed overnight at 4 °C in 4% paraformaldehyde (PFA) with 10% dimethylsulfoxide (DMSO) and 3% Nonidet P-40 (NP-40). After dehydration through a graded methanol–ethanol series, tissues were cleared at 65 °C and permeabilized with 100 μg/mL proteinase K. Membrane permeability was further enhanced with 10% DMSO, and non-specific binding was blocked with 3% bovine serum albumin. Samples were then incubated with an anti-IAA primary antibody (1:100), which recognizes the conjugated IAA moiety, followed by an horseradish peroxidase-conjugated secondary antibody (1:500). Signals were visualized with 3,3′-diaminobenzidine (DAB), enhanced by methanol dehydration, and tissues were cleared in chloral hydrate for observation under a bright-field optical microscope[34].
Semi-thin sectioning
-
Seeds from greenhouse-grown wild-type Fielder and TaPGS1 overexpression lines were harvested from 0 to 30 d after flowering and immediately fixed under vacuum in an aldehyde-based fixative for 12–24 h. After three rinses in phosphate buffer (10–20 min each), the samples were dehydrated through an ethanol series of 25%, 40%, 60%, 70%, 80%, 90%, and 100% (30 min at each concentration). Dehydrated tissues were then infiltrated with epoxy resin solutions of 25%, 50%, 75%, and 100%, allowing at least 2 h for each step and holding the final 100% resin overnight (≈12 h). Specimens were embedded in fresh epoxy resin and polymerized at 65 °C for 12–24 h. Semi-thin sections (EM UC7, Leica) were cut on an ultramicrotome, stained with 0.1% toluidine blue, and examined under a microscope (Axio lmager M2, Zeiss) to document endosperm cellularization and cell-division patterns. For seeds collected 4–7 d after flowering, the cellularization status was scored and the proportion of fully cellularized seeds was calculated for each developmental stage.
LUC reporter assay
-
Promoter fragments, spanning approximately 2.0 kb upstream of the translation start site (ATG) of TaFLS, TaF3H, TaDFR, TaCHI, TaPIN2, TaPGP4, and TaYUC10, were amplified from wheat genomic DNA and cloned upstream of firefly LUC in pGreenII-0800-LUC, generating TaFLS-pro:LUC, TaF3H-pro:LUC, TaDFR-pro:LUC, TaCHI-pro:LUC, TaPIN2-pro:LUC, TaPGP4-pro:LUC, and TaYUC10-pro:LUC. The TaPGS1 coding sequence was inserted into pGreenII-62SK to obtain the effector 35S:TaPGS1 (primer sequences are shown in Supplementary Table S1; gene IDs for all constructs are provided in Supplementary Table S2, highlighted). Agrobacterium tumefaciens GV3101 strains carrying 35S:TaPGS1 (or empty pGreenII-62SK as effector control) and the indicated reporter constructs were co-infiltrated into fully expanded leaves of 3-week-old Nicotiana benthamiana. At 36–48 h post-infiltration, LUC and Renilla (REN) activities were measured using the Dual-Luciferase Reporter Assay kit (Promega, Madison, USA), and luminescence images were captured with a Bio-Rad ChemiDoc MP Imaging System. Relative promoter activity was expressed as the LUC/REN ratio. Each reporter–effector combination was assayed with three biological replicates. The assay followed standard procedures[35].
Yeast two-hybrid assay and bimolecular fluorescence complementation
-
Full-length coding sequences of TaPGS1, TaMYB, and TaWD40 (gene IDs in Supplementary Table S3) were amplified from Fielder cDNA. TaPGS1 was inserted into the pGADT7 activation-domain (AD) vector, whereas TaMYB and TaWD40 were cloned into the pGBKT7 DNA-binding-domain (BD) vector. Each BD construct (BD-MYB or BD-WD40) was co-transformed with AD-TaPGS1 into Saccharomyces cerevisiae strain AH109, and empty AD and BD vectors served as negative controls. Transformants were selected on SD/-Leu/-Trp medium and then transferred to SD/-Leu/-Trp/-His medium to test interaction-dependent growth; colonies were scored after 36–48 h.
For BiFC, the TaPGS1 coding sequence was fused to the N-terminal fragment of YFP in vector 35S-SPYNE, and full-length TaMYB or TaWD40 was fused to the C-terminal fragment of YFP in vector 35S-SPYCE. Sequenced constructs were introduced into Agrobacterium tumefaciens GV3101. Cultures containing TaPGS1-YNE were mixed 1:1 (v/v) with either TaMYB-YCE or TaWD40-YCE and infiltrated into the abaxial side of fully expanded leaves of three-week-old Nicotiana benthamiana using a needle-less syringe. After 36–48 h, YFP fluorescence was observed with a laser-scanning confocal microscope using 514 nm excitation to confirm protein–protein interactions in plants.
Flavonol (kaempferol) treatment
-
Seeds of wild-type Fielder were collected 4 d after flowering and placed on half-strength MS medium supplemented with 0, 10, 25, or 50 µM kaempferol. After 24 and 48 h of incubation, seeds from each treatment (three biological replicates) were fixed in paraformaldehyde solution and processed for semi-thin sectioning, as described in Section 2.6. Fluorescence was quantified in a pericarp region of interest (ROI), with a fixed-size rectangle placed along the bright pericarp band within the boxed area; an adjacent background ROI was subtracted. All sections were stained and imaged in a single session with identical exposure and gain, and the values are reported as relative intensity.
-
In wheat, overexpression of TaPGS1 results in increased seed size. Compared to the Fielder variety, the TaPGS1 overexpression lines exhibit significantly higher thousand-grain weight, grain length, and grain width phenotypes in field conditions (Fig. 1a, d−f). Further analysis of anthocyanin accumulation in grains treated with NaOH from the red-skinned cultivar Chuanmai 104, wild-type Fielder, and TaPGS1 overexpression lines (OE-166-6 and OE-166-39) in the Fielder background revealed no color difference between the TaPGS1 overexpression grains and the wild-type Fielder grains. Only the red-skinned Chuanmai 104 showed a higher grain permeability and deeper color (Fig. 1b). Subsequently, the germination vigor of 7-d-old seeds from wild-type Fielder and TaPGS1 overexpression lines was assessed, showing no significant difference in germination rates between the wild-type and overexpression lines (Fig. 1c). Additionally, field phenotypic traits, including plant height, spike length, tiller number, effective spike number, and grains per spike, were evaluated. The results indicated that, except for the OE-166-39 line, which exhibited decreased plant height and tiller number compared to Fielder, there were no significant differences in other traits (Supplementary Fig. S1a−S1e). These findings suggest that overexpression of TaPGS1 primarily affects the seed size, with minimal impact on other agronomic traits in wheat.
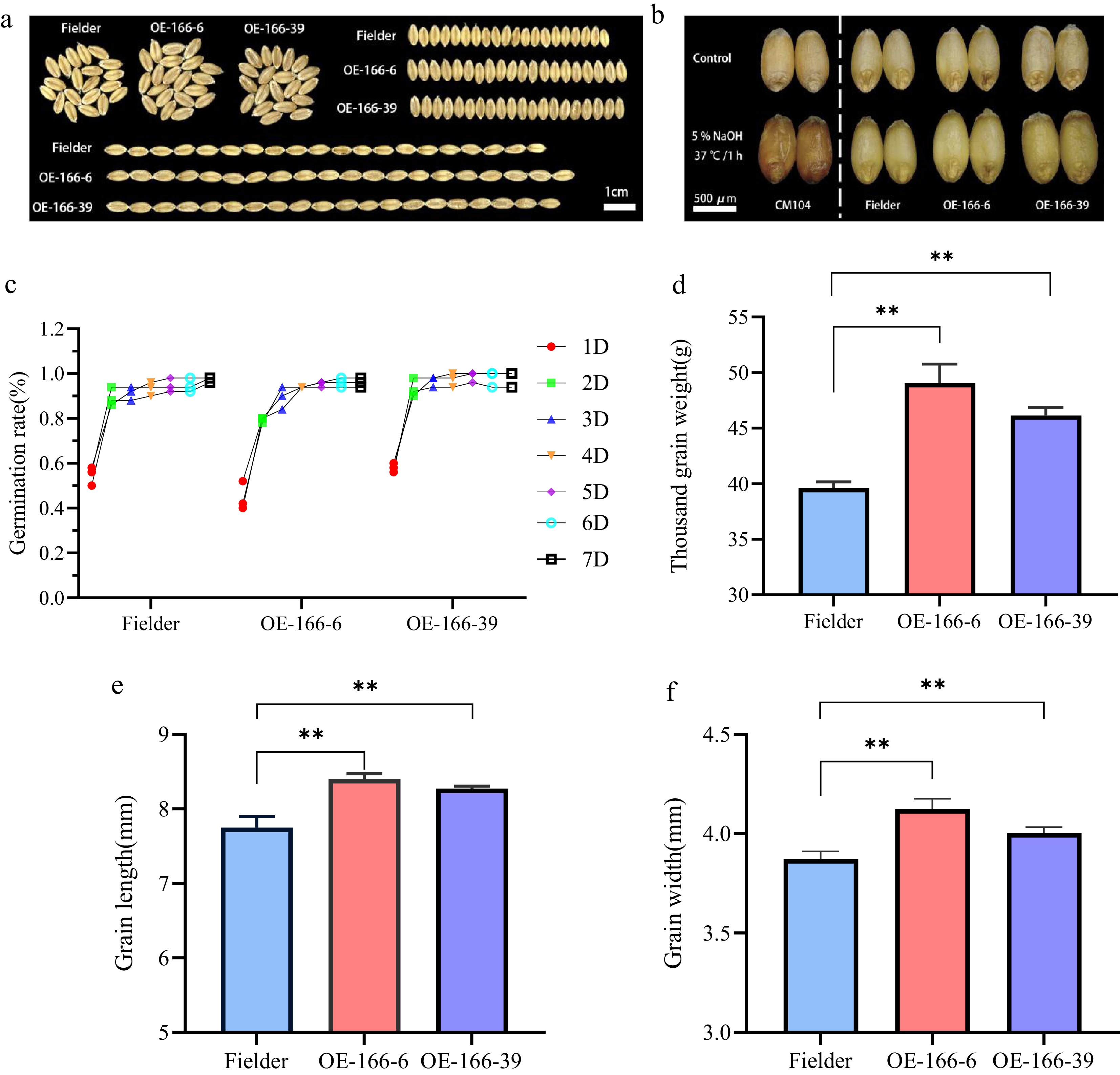
Figure 1.
Analysis of wheat grain phenotype and key agronomic traits in the field. (a) Comparison of grain size between wild-type Fielder and TaPGS1 overexpression wheat. (b) Comparison of grain permeability between wild-type Fielder, TaPGS1 overexpression lines, and red-skinned cultivar Chuanmai 104 (CM104) after NaOH treatment. CM104 (red-pericarp cultivar) was included as a positive control to validate the NaOH-based anthocyanin detection assay and was not used for statistical comparison with the wild type (Fielder) or TaPGS1 overexpression lines. The vertical dashed line separates the positive control from the experimental groups. (c) Comparative analysis of 7-d germination vigor between wild-type Fielder and TaPGS1 overexpression lines. (d−f) Field analysis of thousand-grain weight (d), grain length (e), and grain width (f) between wild-type Fielder and TaPGS1 overexpression lines (* p < 0.05; ** p < 0.01, student's t-test).
TaPGS1 overexpression enhances flavonoid-pathway gene expression and metabolite levels
-
The total metabolites in seeds of wild-type Fielder and TaPGS1 overexpressing wheat were measured at 5 DPA. Flavonol-related metabolites showed the strongest differences (Fig. 2a). Within the 42 annotated compounds of this class, flavonol-related metabolites, including the flavanone precursors naringenin and eriodictyol and the flavonol/derivatives kaempferol, kaempferitrin, and myricitrin, showed higher mean levels in TaPGS1 lines than in wild type (Fig. 2b; Supplementary Table S2), and most of the remaining compounds also trended upward overall (Supplementary Table S2). In parallel, transcriptome profiling provided transcripts per million (TPM). values for all genes in seeds at 5, 10, 15, and 20 DPA (Supplementary Table S4). Differential expression and qPCR analyses showed increased expression of upstream flavonoid-pathway genes, together with the branch markers FLS (flavonol) and DFR (anthocyanin) in the TaPGS1 overexpression lines (Fig. 2c; Supplementary Fig. S2). At 5 DPA, the accumulation of kaempferol and kaempferitrin coincided with higher transcript levels of FLS, while DFR expression was also increased (Fig. 2b–c; Supplementary Tables S2, S4). Pericarp pigmentation remained unchanged, and F3′H was not significantly induced, which is consistent with the observed enrichment of flavonol-related features under our conditions (Fig. 2a–c; Supplementary Fig. S2). Consistent with the unchanged pericarp color, the expression of the wheat anthocyanin-related GST (arGST-like) candidate[36] (highlighted in blue in Supplementary Table S4) was not significantly different at 5 DPA.
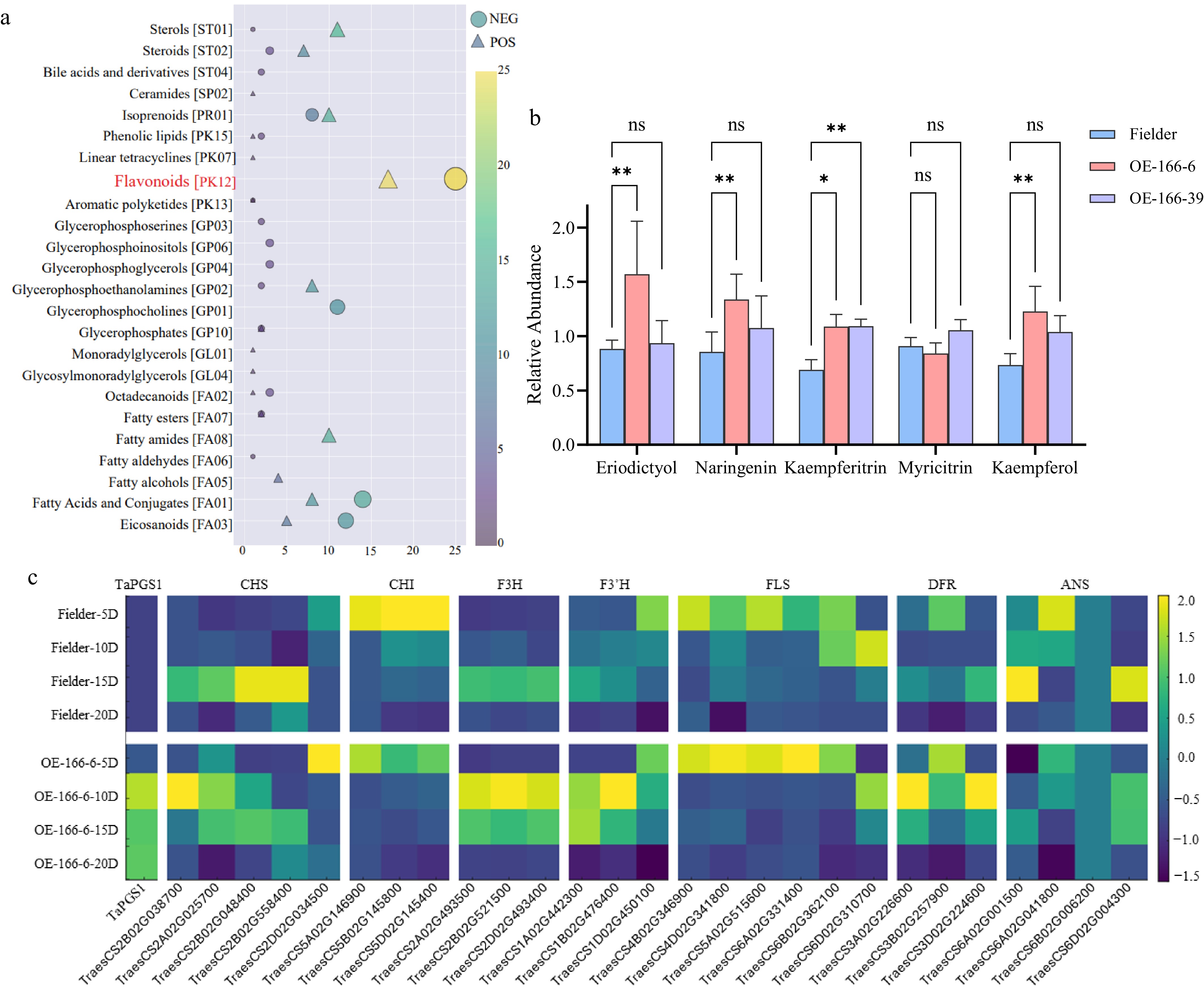
Figure 2.
Metabolomic and transcriptomic profiles of Fielder and TaPGS1 overexpression (OE) seeds at 5 DPA. (a) Class-level enrichment of differentially abundant metabolites; the flavonoid class contains the largest number of significantly changed features. The full set of 42 annotated flavonoid-class compounds with group means and statistics is provided in Supplementary Table S2. (b) Flavonol-related metabolites: naringenin, eriodictyol (precursors) and kaempferol, kaempferitrin, myricitrin (flavonol/its glycosides). Bars show abundance normalized to the QC median (sample peak area / QC median peak area), averaged over four biological replicates (mean ± SD; * p < 0.05; ** p < 0.01, student's t-test). Exact values for all 42 flavonoid-class compounds are listed in Supplementary Table S2. (c) Heatmap of flavonoid-pathway genes in wild-type and TaPGS1-OE seeds at 5, 10, 15, and 20 DPA. Gene names are shown across the top of the heatmap; the corresponding gene IDs are shown along the bottom (see also Supplementary Table S4). Values are TPM that were centered and scaled per gene (row-wise z-score) for visualization, as indicated by the color bar.
Based on these findings, seeds at 5 DPA from wild-type Fielder and TaPGS1 overexpression lines (OE-166-6, OE-166-39) were hand-sectioned, stained with DPBA, and imaged by fluorescence microscopy. The kaempferol-associated (green) signal accumulated primarily in the inner layer of the seed coat adjacent to the endosperm and was higher in the overexpression lines, with relative intensities of 2.0× and 1.3× those of Fielder, respectively (Fig. 3a, b). By contrast, the quercetin-associated (yellow) signal did not differ significantly between genotypes (Fig. 3a, b). These DPBA readouts provide spatial localization and relative fluorescence under matched staining and imaging conditions and are concordant with the LC-MS-based relative quantification (Fig. 2; Supplementary Table S2). Together, these data indicate that TaPGS1 overexpression is associated with increased expression of flavonoid-pathway genes and higher levels of flavonol-related metabolites, notably kaempferol, in developing wheat seeds.
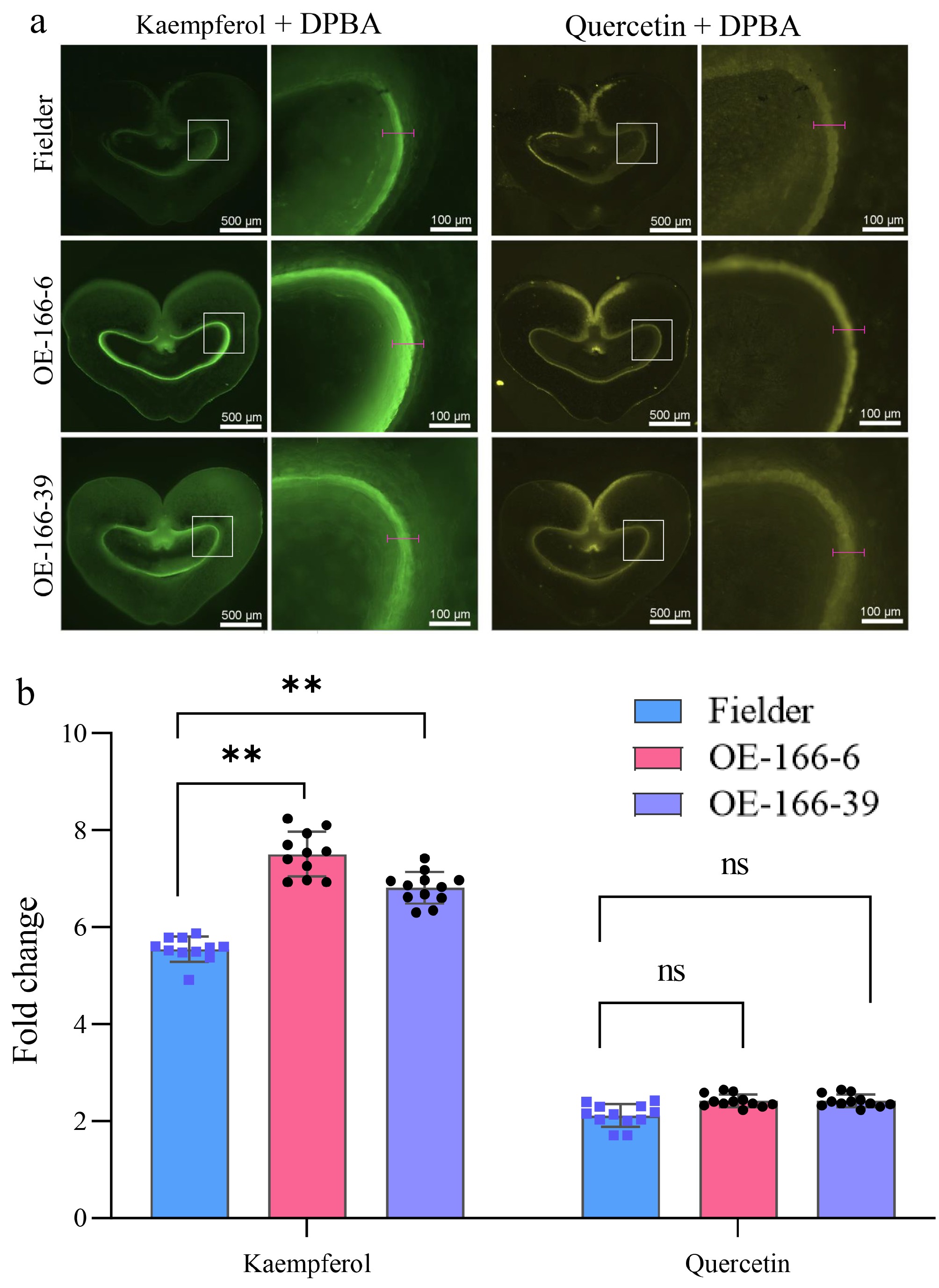
Figure 3.
Localization and relative fluorescence of flavonols in wheat seeds. (a) DPBA staining of seeds at 5 DPA (green = kaempferol-associated, yellow = quercetin-associated). The white box marks the enlarged area. The red line segment indicates the region used for intensity extraction. (b) Relative fluorescence intensities of the green and yellow channels from (a) (n = 10 per genotype; p < 0.05; p < 0.01, student's t-test).
TaPGS1 overexpression elevates auxin and delays endosperm cellularization
-
Flavonols have been reported to modulate polar auxin transport in several plant systems. To examine whether TaPGS1-associated flavonol enrichment is accompanied by changes in auxin distribution in wheat grains, we performed auxin immunolocalization in 5-DPA seeds. Auxin signals were significantly stronger in the inner pericarp of TaPGS1 overexpression lines than in the wild type (Fig. 4a, b). Notably, this auxin-enriched region coincides with the pericarp domain showing elevated kaempferol-associated fluorescence (Fig. 3a, b), supporting a close spatial association between flavonol enrichment and local auxin accumulation.
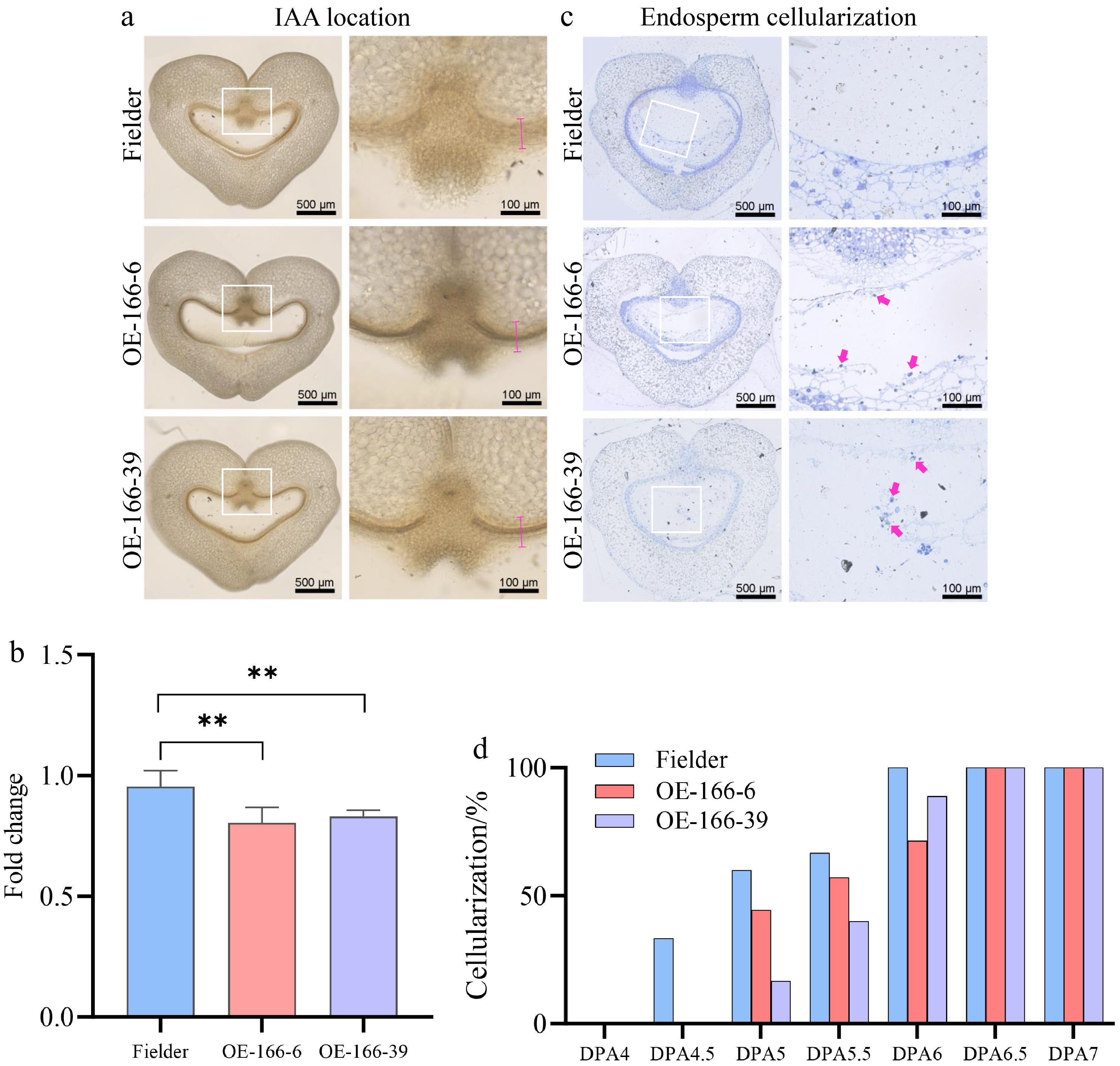
Figure 4.
Analysis of auxin localization and endosperm cellularization phenotype in TaPGS1 overexpression lines. (a) Auxin accumulation in the seeds of wild--type Fielder and TaPGS1 overexpression lines at 5 DPA. The white box indicates the enlarged display area. The red line segment represents the data collection area. (b) Quantification of the auxin accumulation shown in (a). The difference in auxin levels between the TaPGS1 overexpression lines and wild-type Fielder is significant (n = 10; mean ± SD; * p < 0.05; ** p < 0.01, student's t-test). (c) Schematic representation of endosperm cellularization observed through semi-thin sectioning of seeds. The red arrow marks the free nuclei in the coenocytic endosperm that has not yet undergone cellularization. (d) Proportion of seeds with completed cellularization at different DPA. Cellularization was delayed in TaPGS1 overexpression lines compared to Fielder, particularly at 4.5 to 6 DPA (n≥5).
To further confirm whether endosperm cellularization is affected, we prepared semi-thin sections to observe seeds at different stages of cellularization. The results showed that, in wild-type Fielder, 33.33% of the seeds had completed endosperm cellularization by 4.5 DPA, and all seeds had completed cellularization by 6 DPA. In contrast, none of the TaPGS1 overexpression seeds had completed cellularization by 4.5 DPA. By 5 DPA, 44.44% of the TaPGS1 overexpression seeds had completed cellularization, and by 6.5 DPA, all seeds had completed cellularization, indicating a delay of 0.5 d compared to wild-type Fielder (Fig. 4c, d). These results indicate that TaPGS1 overexpression is associated with local auxin accumulation in the inner pericarp and a delay in endosperm cellularization.
Kaempferol treatment recapitulates the cellularization delay
-
To test whether flavonol accumulation directly affects endosperm cellularization, wild-type seeds at 4 DPA were exposed to increasing concentrations of kaempferol (10, 25, and 50 μM), and cellularization was assessed at 24 and 48 h. Compared to the control (0 μM), all treated seeds exhibited reduced cellularization, with more pronounced inhibition observed at higher concentrations. Although cellularization progressed further by 48 h than at 24 h under each condition, the overall delay was concentration-dependent (Supplementary Fig. S3). These findings are consistent with the cellularization delay observed in TaPGS1 overexpression lines, supporting a role for kaempferol in modulating this process.
We next asked whether such a delay affects the final cellular architecture. Seeds from TaPGS1-overexpressing and wild-type plants were sampled at 8 DPA when endosperm cellularization was complete, and semi-thin sections were prepared for cell number quantification across three defined endosperm regions (Supplementary Fig. S4a). TaPGS1 overexpression lines consistently showed higher cell counts than the wild type. Although region c in line OE-166-6 did not show a statistically significant difference, its cell number was still greater than that of the wild type (Supplementary Fig. S4b). These results suggest that the delay in cellularization may contribute to an increased endosperm cell number.
TaPGS1 transactivates selected flavonoid pathway promoters but not the tested auxin-related promoters
-
To test whether transcript increases observed in TaPGS1 overexpression lines include promoter-level effects at the flavonol branch and at representative pathway nodes, we assayed TaFLS-pro:LUC, TaF3H-pro:LUC as an upstream shared step, and TaDFR-pro:LUC as an anthocyanin-branch enzyme in tobacco leaves. Co-infiltration with 35S:TaPGS1 significantly increased reporter activity for all three promoters (Fig. 5a–f). In contrast, TaPGS1 did not increase reporter activity from promoters of wheat homologs of TaPIN1, TaPGP4, TaYUC10 that are related to auxin transport or biosynthesis, or from the upstream flavonoid gene TaCHI (Supplementary Fig. S5a−S5h). These results establish that TaPGS1 has promoter transactivation capacity within the flavonoid pathway, most notably at TaFLS, while not acting on the tested auxin-related promoters or on TaCHI. These assays establish the promoter-level activity of TaPGS1 within the flavonoid pathway under the conditions tested, without evidence for the direct activation of the auxin transport or biosynthesis promoters examined.
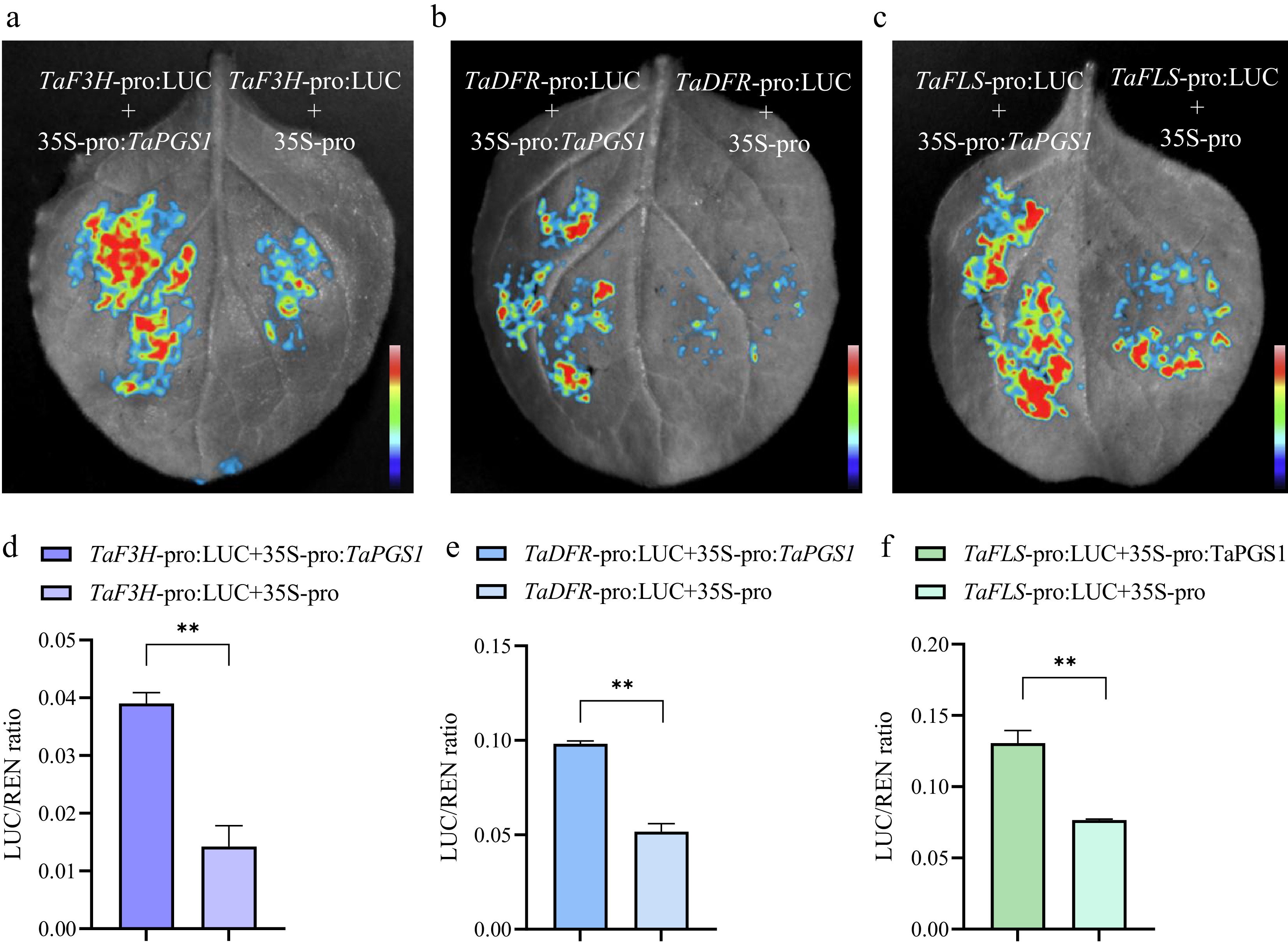
Figure 5.
TaPGS1 transactivates TaFLS, TaF3H, and TaDFR promoters. (a–c) Luminescence images of TaF3H-pro:LUC (a), TaDFR-pro:LUC (b), and TaFLS-pro:LUC (c) co-expressed with 35S-pro:TaPGS1 (left) or empty effector (35S-Pro, right) in tobacco leaves. (d–f) Quantification of relative promoter activity (LUC/REN) for TaF3H-pro:LUC (d), TaDFR-pro:LUC (e), and TaFLS-pro:LUC (f). Bars show mean ± SD; significance: mean ± SD; * p < 0.05; ** p < 0.01, student's t-test.
Given that bHLH transcription factors are often reported to function in the flavonoid biosynthetic pathway by forming a ternary complex with MYB (myeloblastosis-related transcription factor) and WD40 (WD40-repeat protein) proteins[37,38], we investigated whether TaPGS1 might act through similar interactions. Homologs of OsMYB (OsC1) and OsWD40, known to interact with OsRC in rice, were identified in wheat by sequence alignment. Wheat homologs of OsMYB were located on chromosomes 4A, 4B, 4D, and 5A, while homologs of OsWD40 were found on chromosomes 6A and 6B (Supplementary Fig. S6). Candidate genes with over 90% amino acid similarity were selected for interaction assays. Yeast two-hybrid and BiFC experiments showed that TaPGS1 interacts with TaWD40 but not with TaMYB (Fig. 6a, b). These results support a potential TaPGS1–WD40 partnership in wheat; the identity of any MYB partner remains to be determined.
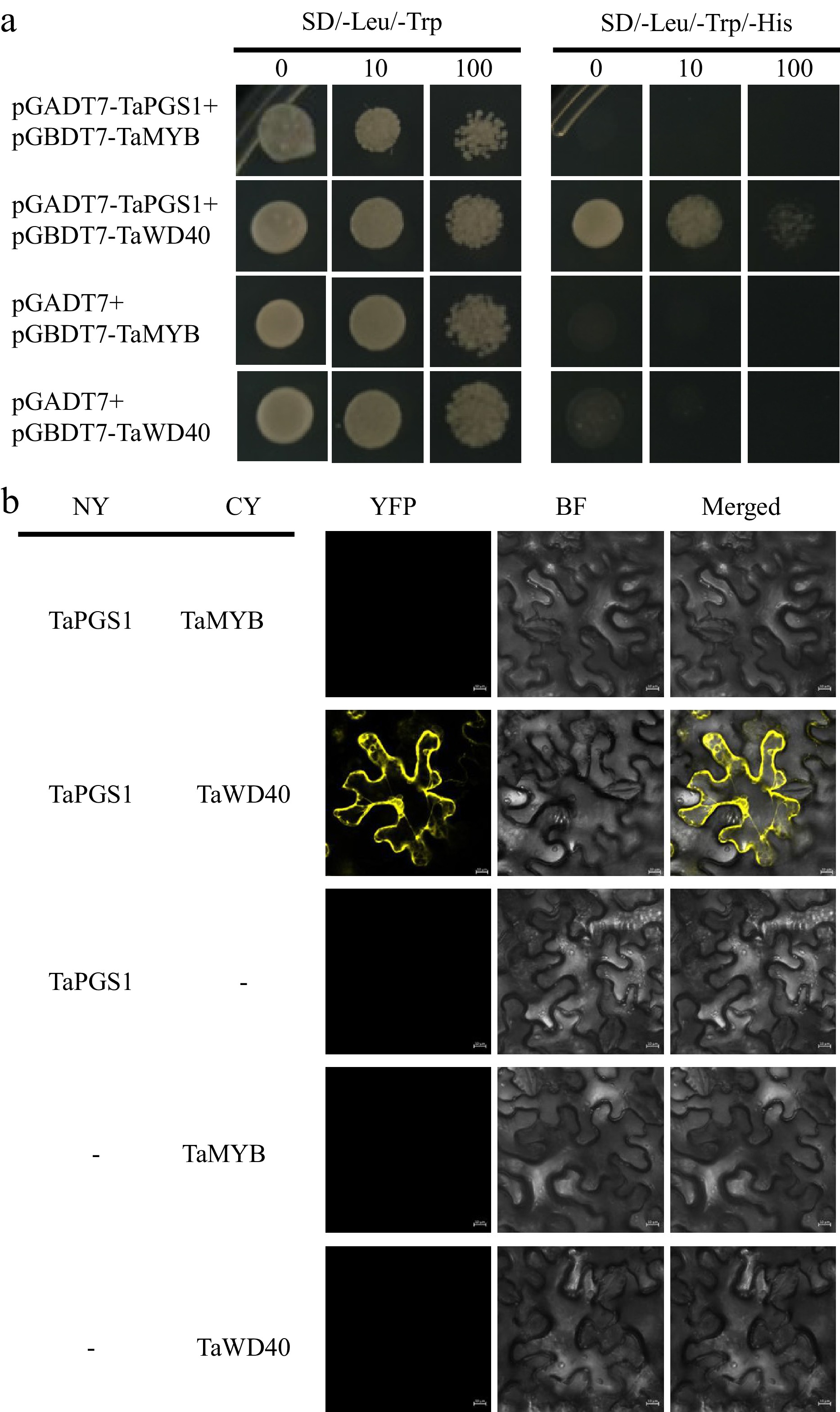
Figure 6.
Validation of interaction proteins of TaPGS1. (a) Yeast two-hybrid assay validating the interactions between TaPGS1 and TaMYB and TaWD40. (b) BiFC validating the interactions between TaPGS1 and TaMYB and TaWD40.
-
In Arabidopsis, the bHLH transcription factor TT8 and, in rice, the TT8-like factor OsRc promote flavonoid and anthocyanin biosynthesis and determine pericarp pigmentation[13,14]. Previous studies have reported that two bHLH homologous genes of TaPGS1, ThMYC4E[39] and TaPpb1[40], are involved in regulating wheat seed color. By contrast, overexpressing TaPGS1 did not alter the pericarp color of wheat under our growth conditions (Fig. 1a, b), consistent with the unchanged anthocyanin readouts. Developmental traits further underscore divergence among these bHLH factors. OsRc influences seed dormancy and grain weight in rice, whereas TaPGS1 overexpression enlarged wheat grains without affecting dormancy (Fig. 1c–f).
The transcriptome and metabolite profiles point to a shift of pathway output toward the flavonol branch. In TaPGS1 lines, F3H and FLS transcripts increased and DFR was also induced, while F3′H did not show a significant increase under our conditions (Fig. 2c; Supplementary Figs S2 and S7). Consistently, kaempferol-related features increased in the LC–MS dataset and kaempferol-associated fluorescence was stronger in the pericarp (Figs 2a–c and 3a, b; Supplementary Table S2). Given that FLS and DFR compete for dihydroflavonol substrates[41], the observed combination of gene expression and metabolite patterns is consistent with a greater representation of flavonol-related features under our conditions; we did not detect an anthocyanin output signal in our assays (Figs 1a, b and 2a–c; Supplementary Fig. S2).
The regulatory context offers a complementary explanation. Anthocyanin production in cereal pericarps requires a complete MYB-bHLH-WD40 (MBW) transcription factor complex. In rice, functional OsRc promotes pigmentation only when the structural gene Rd (encoding DFR) is active[13]. In wheat, overexpression of TaMYB10-3D darkens the normally colorless Fielder pericarp and is accompanied by increased anthocyanin levels, measured by metabolomics[36]. In our assays, TaPGS1 interacted with TaWD40 but not with the tested TaMYB proteins (Fig. 6a, b), and TaPGS1 transactivated TaFLS, TaF3H, and TaDFR promoters in a transient system, while TaPIN1, TaPGP4, TaYUC10, and TaCHI promoters were not activated (Fig. 5a–f; Supplementary Fig. S5). This pattern supports promoter-level capacity within the flavonoid pathway without evidence for a pigmentation-type MBW assembly in the Fielder pericarp.
Together, these data indicate that, despite high sequence similarity, TaPGS1 in wheat operates within the flavonoid pathway with readouts consistent with a bias toward the flavonol branch, in a model that aligns with increased grain size while pericarp pigmentation remains unchanged. This divergence likely reflects differences in MBW complex composition and in the availability of downstream structural genes among wheat, rice, and Arabidopsis.
Flavonol enrichment correlates with auxin accumulation and delayed endosperm cellularization
-
Flavonoids have long been recognized as endogenous inhibitors of polar auxin transport[22]. In particular, flavonols, such as kaempferol derivatives, attenuate PIN-mediated auxin efflux, causing local auxin accumulation[17,19,20,42]. Because endosperm cellularization is triggered only after auxin levels fall below a critical threshold[11], any restriction of auxin export is expected to delay wall formation.
Our results are consistent with this framework. In TaPGS1 overexpression lines, kaempferol-related signals increased based on LC-MS relative quantification and DPBA fluorescence, with stronger kaempferol-associated fluorescence in the pericarp than in Fielder (Figs 2a, b and 3a, b). Taken together with the unchanged pericarp pigmentation, the lack of detectable F3′H induction under our conditions, and the increases in FLS expression and kaempferol levels (Figs 1a, b and 2a–c; Supplementary Fig. S2), we interpret TaPGS1 as a pathway-level modulator in developing seeds, with readouts consistent with a bias toward the FLS branch. Auxin localization showed stronger signals within the endosperm–maternal interface and the inner pericarp in TaPGS1 overexpression lines, consistent with an altered auxin distribution pattern compared with the wild type. Concomitantly, endosperm cellularization was delayed by approximately 0.5 d, permitting additional nuclear divisions and increasing the endosperm cell number (Fig. 4a–d; Supplementary Fig. S4). Exogenous kaempferol reproduced the delay in a concentration-dependent manner, supporting the view that flavonols, rather than TaPGS1 per se, act as the proximate modulators (Supplementary Fig. S3).
Several observations suggest that the auxin accumulation phenotype and the cellularization delay are more consistent with altered auxin distribution/efflux than with enhanced auxin biosynthesis. TAA1/YUC transcripts did not increase in TaPGS1 lines, and pathway enrichment of the metabolome did not indicate up-regulation of auxin-biosynthetic routes (Supplementary Table S4). In transient assays, TaPGS1 did not transactivate the promoters of auxin-transport or biosynthesis genes tested (TaPIN2, TaPGP4, TaYUC10), whereas it activated TaFLS and TaF3H promoters (Fig. 5a−f; Supplementary Fig. S5). This observation is compatible with a model in which flavonols act as proximate modulators of auxin transport downstream of TaPGS1. These promoters represent transport and biosynthesis nodes implicated in auxin flux control and were included to test whether TaPGS1 acts directly on auxin modules in addition to flavonoid nodes. Literature further shows that kaempferol can interfere with PIN phosphorylation and trafficking[16,18], providing a plausible molecular basis for reduced efflux.
Taken together, our data support a working model in which TaPGS1-driven flavonol enrichment is accompanied by local auxin accumulation at the endosperm–maternal interface and a modest delay in endosperm cellularization. Direct measurements of auxin flux and transporter dynamics in wheat grains will be important to further test this model in future work.
TaPGS1 regulates seed development and increases seed size through multiple pathways
-
Previous work demonstrated that overexpression of TaPGS1 in wheat and rice increases the thousand-kernel weight and grain size, in part because TaPGS1 activates TaFL3/OsFL3 during early grain filling (5–10 DPA)[25]. The maize ortholog ZmFL3 exerts a comparable influence on endosperm storage deposition[43], supporting the view that the TaPGS1–FL3 pathway enhances assimilate accumulation in developing grains.
The present study identifies a mechanism by which TaPGS1 enhances the flavonoid pathway with flux biased toward the flavonol branch. TaPGS1 up-regulates upstream flavonoid genes as well as the flavonol-branch gene FLS, and it also induces DFR expression. However, ANS, the key downstream enzyme required for anthocyanin formation, remains unchanged, which explains why anthocyanin accumulation shows no significant difference compared to the wild type. Consequently, TaPGS1 increases kaempferol in the pericarp and is accompanied by local auxin accumulation at the endosperm–maternal interface. This auxin accumulation coincides with a ~0.5-d delay in endosperm cellularization and an increase in final endosperm cell number. Auxin buildup postpones cellularization by about half a day, permits additional nuclear divisions, and raises the final endosperm cell number. Exogenous kaempferol repeats this delay in a concentration-dependent manner, supporting the interpretation that flavonols can modulate auxin transport under the tested conditions. The observed delay is moderate, remaining below the threshold that arrests grain development[44], and agrees with reports that brief extensions of the coenocytic phase can enlarge grains[10,45].
TaPGS1 therefore acts through at least two partially independent routes. One enhances assimilate supply via FL3 activation, and the other expands sink capacity by slowing cellularization through a flavonol–auxin module. Determining the quantitative contribution of each pathway will require genetic separation or temporal regulation of TaPGS1 targets, yet the convergence of both routes on grain enlargement establishes TaPGS1 as a promising lever for yield improvement in wheat.
In addition to yield-related traits, the flavonol enrichment observed in TaPGS1-overexpressing lines may also contribute to nutritional quality, as compounds such as kaempferol and quercetin possess well-documented antioxidant activities and potential health benefits. Thus, TaPGS1 not only regulates the developmental pathways underlying grain size but may also enhance the functional value of wheat grains.
-
Overexpression of TaPGS1 enhances the flavonoid pathway in developing wheat seeds and is accompanied by higher flavonol-related metabolites, with kaempferol being the most prominent. Auxin localization indicates increased auxin within the endosperm and a delay in endosperm cellularization; kaempferol treatment reproduces this delay. Transcriptional profiling and promoter–reporter assays show that TaPGS1 stimulates flavonoid-pathway gene expression, with FLS marking the flavonol branch. Together, these results support a model in which flavonol enrichment is associated with local auxin accumulation/redistribution in developing grains, coinciding with delayed endosperm cellularization and increased endosperm cell number and grain size. This work links flavonoid metabolism to auxin-mediated control of cellularization and highlights the regulatory points that could be leveraged to improve the wheat yield.
-
The authors confirm their contributions to the paper as follows: study conception and design: Wang J; data analysis: Guo X, Cheng M, Liu X; plasmid construction, genetic transformation, and other assays: Guo X, Zhao S, Liang W, Liu X; phenotypic analysis and planted the experimental material: Zhao S, Jin Y, Li M, Liu X; funding acquisition and resources: Wang J; draft manuscript preparation: Liu X, Guo X; manuscript review and editing: Liu X, Guo X, Wang J; supervision: Dong H, Chen Z, Chen Q. All authors reviewed the results and approved the final version of the manuscript.
-
Raw sequence data were deposited in the National Center for Biotechnology Information (NCBI) under accession no. SUB15671480.
-
This research was funded by the National Key Research and Development Project (SQ2024YFF1200011), the Scientific and Technological Innovation 2030 Major Project (2023ZD04069), the National Natural Science Foundation of China (U22A20472), the Sichuan Science and Technology Support Project (2025NSFTD0025), the Technology Support Project of Chengdu (2023-YF08-00008-SN), and the open research fund of SKL-CGEUSC (SKL-ZD202212).
-
Zhongxu Chen is affiliated with Chengdu Tiancheng Smart Agriculture Technology Co., Ltd. The company provided transcriptome data analysis and manuscript revision suggestions for this study. The authors declare that this did not influence the objectivity of the experimental results or the interpretation of data.
-
accompanies this paper online at: https://doi.org/10.48130/seedbio-0026-0015.
-
# Authors contributed equally: Xin Liu, Shuyu Zhao, Xiaojiang Guo
- Supplementary Table S1 Primers list.
- Supplementary Table S2 Levels of 42 flavonoid-class metabolites in seeds of wheat cv. Fielder and TaPGS1 overexpression lines (OE-166-6, OE-166-39) at 5 DPA.
- Supplementary Table S3 Wheat homologous genes of the anthocyanin biosynthesis pathway ternary complex(bHLH-MYB-WD40)*.
- Supplementary Table S4 Genome-wide transcript abundance (TPM) in seeds of Fielder and TaPGS1 overexpression (OE) lines at 5, 10, 15, and 20 d post-anthesis (DPA).
- Supplementary Fig. S1 Analysis of wheat key agronomic traits in the field.
- Supplementary Fig. S2 Real-time fluorescence quantitative analysis of TaPGS1 and flavonols pathway-related genes in wild-type Fielder and overexpression lines OE-166-6, OE-166-39.
- Supplementary Fig. S3 Effects of kaempferol treatment on endosperm cellularization at 4 DPA.
- Supplementary Fig. S4 Statistical analysis of endosperm cell number in wild-type Fielder and TaPGS1 overexpression lines at 8 DPA.
- Supplementary Fig. S5 Validation of downstream transcriptional activation of TaPGS1.
- Supplementary Fig. S6 Amino acid sequence alignment of each copy of TaMYB and TaWD40.
- Supplementary Fig. S7 Biosynthetic pathways of flavonols and anthocyanins, and expression profiles of key enzyme genes.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu X, Zhao S, Guo X, Jin Y, Liang W, et al. 2026. TaPGS1-associated flavonol enrichment correlates with delayed endosperm cellularization and increased grain size in wheat. Seed Biology 5: e018 doi: 10.48130/seedbio-0026-0015 

TaPGS1-associated flavonol enrichment correlates with delayed endosperm cellularization and increased grain size in wheat
- Received: 06 December 2025
- Revised: 25 January 2026
- Accepted: 09 February 2026
- Published online: 16 June 2026
Abstract: Flavonoids contribute to seed development, and flavonols can modulate polar auxin transport. We previously showed that overexpressing the wheat bHLH factor TaPGS1 increases grain size. Here, the metabolomic and transcriptomic profiling of TaPGS1 overexpression lines revealed enrichment of flavonol-related metabolites, most notably kaempferol, together with coordinated increases in flavonoid-pathway gene expression, including FLS and upstream nodes. Auxin localization indicated local auxin buildup in developing grains, and exogenous kaempferol reproduced a delay in endosperm cellularization. Promoter–reporter assays showed that TaPGS1 activates representative flavonoid-pathway promoters. These findings support a model in which TaPGS1 enhances the flavonoid pathway and is associated with altered auxin distribution, leading to delayed cellularization and increased endosperm cell number, which together contribute to grain enlargement in wheat.
-
Key words:
- Flavonol /
- Flavonoid pathway /
- Nutritional quality /
- Auxin transport /
- Endosperm cellularization /
- Wheat grain