









-
The term 'protoplast' refers to a living plant cell that has had its cell wall removed through mechanical separation or chemical enzymatic hydrolysis. Although mechanical isolation was first achieved in 1892, its low yield led to the adoption of enzymatic methods, first pioneered by Cocking in 1960[1]. This has opened a new era of preparation of protoplasts by enzymatic hydrolysis, such as Carlson's 1972 creation of the first interspecific tobacco hybrid via protoplast fusion[2]. Recently, protoplast technology has been successfully applied in multiple fields, such as in respiration, plant regeneration processes, and other vital activities[3,4]. Beyond creating novel germplasms by overcoming hybridization incompatibility[5], protoplasts serve as ideal biological systems for investigating protein transport and gene transfer[6]. These methods are particularly useful for systems studies in which stable transgenics, mutation databases, and even tissue cultures are unavailable. Therefore, the establishment and optimization of efficient protoplast separation methods are highly important.
Hibiscus hamabo (H. hamabo) is a deciduous shrub or small tree belonging to the Malvaceae family[7]. Characterized by ecological adaptations typical of mangroves and distinct coastal-terrestrial transitional traits, and fulfilling an indispensable role in coastal ecosystems[8−11]. Functioning as a natural coastal barrier, H. hamabo is instrumental in constructing seawall shelterbelts and soil-water conservation forests[12]. Beyond its conventional application in coastal windbreaks, H. hamabo serves as an exceptional candidate for seaside greenbelts, parklands, and herbaceous border designs. Nevertheless, anthropogenic pressures now classify this species as endangered[7,8]. In recent years, with the rapid advancement of molecular biotechnology, H. hamabo—with a rich genetic background—has emerged as a focus in genetic engineering research. However, owing to unstable genetic transformation and the lack of a mutation database, research on H. hamabo has focused mainly on the early stages of physiology, biochemistry, and gene cloning, and the molecular mechanism has been difficult to elucidate. Protoplasts contain complete genetic material and totipotency and are excellent recipients for genetic transformation. Protoplasts, which serve as excellent models for stable transgenesis, enable instantaneous transcriptome reactions to gene expression, high-throughput verification of gene functions, and establishment of gene regulatory networks[13−15]. As early as 1991, a method for isolating protoplasts from callus tissues of Hibiscus syriacus L. was established, which laid the groundwork for genetic transformation studies in Hibiscus species[16]. However, this approach exhibited significant limitations: (1) protoplasts could only be obtained from callus cultures, with post-cultivation development arrested at the stage of cell clusters and callus formation; (2) leaf-derived protoplasts proved impractical due to persistent epidermal contamination and extremely low release efficiency; and (3) the exclusive reliance on callus systems raised concerns about potential genetic instability. Consequently, establishing optimized protoplast isolation protocols for H. hamabo constitutes a critical technological advancement, enabling high-throughput genetic enhancement of stress resilience traits and ornamental phenotypes, while accelerating molecular dissection of developmental mechanisms—thereby catalyzing transformative breakthroughs in both endangered species conservation and coastal ecosystem restoration initiatives.
In this study, we present a protocol for the isolation and transfection of protoplasts from the stems of H. hamabo. The establishment of this system provides a new idea for the study of H. hamabo. This protoplast-based technology enables applications such as somatic cell fusion, genetic transformation, and plant regeneration. This study provides a versatile platform for fundamental research, genetic improvement, and the development of new cultivars of H. hamabo.
-
The supplied material was 12-month-old H. hamabo grown in the greenhouse of Zhejiang Subtropical Crops. Wild-type plants were cultivated in a walk-in growth chamber at 21–25 °C under a 16-h/8-h light/dark cycle. Supplemental lighting (~300 µmol·m−2·s−1) was supplied by three-band linear fluorescent lamps (T5, 28 W, 6,400 K), with relative humidity maintained between 60% and 80%.
Equipment
-
High-capacity refrigerated centrifuge; Microcentrifuge; Optical microscope; Fluorescence microscope.
Reagent setup
-
(1) Enzyme solution.
Composition: 1.5% (wt/vol) cellulase R10; 0.4% (wt/vol) macerozyme R10; 0.5 M mannitol, 20 mM MES (pH 5.7); 20 mM KCl.
Preparation: Warm the mixture at 55 °C for 10 min to inactivate DNAse and proteases, and increase enzyme solubility; cool it to room temperature; add 10 mM CaCl2 and 0.1% BSA immediately before use.
(2) MMG solution (mannitol-magnesium solution).
Composition: 0.5 M mannitol; 15 mM MgCl2; 4 mM MES (pH 5.7).
(3) WI solution (washing and incubation solution).
Composition: 0.5 M mannitol; 4 mM MES (pH 5.7); 20 mM KCl.
(4) PEG solution (transformation medium).
Composition: 40% (wt/vol) PEG4000; 0.2 M mannitol; 100 mM CaCl2.
Protoplast isolation
H. hamabo protoplast isolation from leaf and petiole -
(1) Leaf and petiole preparation:
*Select healthy, disease-free young leaves;
*Separate the leaf blade and petiole;
*Using a sharp blade, excise the middle section of the leaf blade and cut it longitudinally into 1–2 mm wide strips (Fig. 1);

Figure 1.
H. hamabo protoplast isolation from leaf flesh and petiole. (a) The separation process of protoplasts of H. hamabo leaf flesh. (b) The separation process of protoplasts of the H. hamabo petiole. The procedures for isolating protoplasts from the two tissues are as follows; preparation of tender tissues, enzymolysis, vacuum-pressure, release protoplasts, cell sedimentation, and microscopic examination.
*Cut the petiole transversely into 1–2 mm thick sections (Fig. 1).
(2) Enzymatic digestion:
*Place the prepared leaf blade and petiole tissues into a 6-well plate containing 5 ml of enzyme digestion solution for incubation;
*Transfer the samples to a vacuum chamber and apply vacuum treatment at 10-min intervals for a total of 1 h to facilitate enzyme solution infiltration;
*Transfer the samples along with the enzyme digestion solution to a low-speed horizontal shaker and incubate at room temperature in the dark for 1 h for enzymatic digestion.
(3) Protoplast release:
*Add an equal volume of WI solution to the enzyme digestion mixture to dilute it, and filter through a 75 μm nylon mesh (to remove undigested tissue);
*Transfer the filtered solution to a 2.0 ml centrifuge tube;
*Centrifuge at 300 × g for 3 min.
(4) Resuspension and quantification:
*Discard the supernatant and resuspend the pellet in 3.5 ml of MMG solution;
*Allow the protoplasts to settle naturally on ice for 30 min.
(5) Collection of protoplasts:
*Discard the supernatant and resuspend the pellet in 800 μL of MMG solution;
*Check for the release of protoplasts in the solution under the microscope;
*Measure the protoplast concentration using a haemocytometer.
H. hamabo protoplast isolation from stems -
(1) Stem preparation:
*Select healthy one-year-old plants;
*Cut stems into several 10 cm-long stem sections.
(2) Enzymatic digestion:
*Incubate the debarked stems in 50 ml of the enzyme mixture;
*Conditions: room temperature, 3 h, darkness.
(3) Protoplast release:
*Transfer the digested stems to 30 ml of MMG solution in a 50-ml tube;
*Shake gently for 30 to 60 s.
(4) Filtration and centrifugation:
*The released protoplasts were filtered through 75-µm nylon mesh into a new 50-ml tube;
*Centrifuged at 300 × g for 3 min.
(5) Resuspension and quantification:
*Discard the supernatant and resuspend it in 1 ml of MMG solution.
(6) Purification of protoplasts:
*Layer the protoplast suspension over 5 mL of 2 M sucrose solution;
*Centrifuged at 300 × g for 2 min for density centrifugation treatment;
*Critical step: transfer the protoplasts to the sucrose pad and be careful not to shake or mix.
(7) Collection of protoplasts:
*After centrifugation, the precipitate was divided into upper, middle, and lower layers. The cell mixture in the middle layer was aspirated;
*Washed with 1 ml of 0.5 M mannitol;
*Centrifuged at 300 × g for 2 min to remove the supernatant;
*1 ml of MMG was added along the tube wall to resuspend the protoplast;
*The cells were observed under a microscope, and the number of cells was counted.
Protoplast yield and viability estimation
-
Protoplast quantification and imaging were performed using a Leica DM6 B microscope coupled with a hemocytometer. Protoplast yield was calculated as the total number of viable protoplasts released per gram fresh weight (FW) of initial plant tissue. Protoplast viability was assessed using the Trypan Blue exclusion method. Specifically, a 0.4% Trypan Blue solution (in PBS buffer, pH 7.2) was prepared. A 10 µL aliquot of protoplast suspension was mixed with 40 µL of the dye solution (final concentration 0.32%), followed by incubation in the dark for 2 min. Immediate microscopic examination was then performed: viable protoplasts appeared colorless and transparent due to intact cell membranes, while non-viable or damaged cells exhibited dark blue staining from dye penetration. Viability (%) was calculated as the percentage of unstained cells relative to the total cell count using a hemocytometer. All counting procedures were completed within 10 min post-staining to minimize errors.
Protoplast transfection and incubation
-
(1) Preparation of the transfection mixture:
*Plasmid, protoplast, and PEG solutions were mixed at a volume ratio of 1:10:11;
*Incubating for 5–10 min at room temperature;
*Critical step: higher-quality DNA is more likely to result in higher transfection efficiencies; the plasmid concentration for protoplast transformation was maintained at 2,000 ng/µl, and the DNA OD260/OD280 values are 1.8–2.0.
(2) Stop the reaction:
*Add WI solution with 4× volume of PEG solution;
*Gently invert and mix well to terminate the reaction.
(3) Collect transfected protoplasts:
*Centrifuge at 300 × g for 2 min to remove the supernatant.
(4) Resuspend:
*The transfected protoplasts were resuspended in an appropriate amount of WI solution;
*Critical step: the amount of WI solution used should be adjusted according to the number of cells required for different experiments.
Protoplast culture, harvesting, and determination of transfection efficiency
-
(1) Cultivation of transfected protoplasts:
*Transfected protoplasts were placed in a 9 cm culture dish coated with 1% (wt/vol) BSA solution and incubated overnight in the dark, at room temperature.
(2) Transfection efficiency calculation:
*After incubation, transfer 200 µl of protoplasts and centrifuge at 300 × g for 2 min;
*Resuspend in 20 µl of WI solution to examine the transfection efficiency;
*Under the fluorescence microscope, the transfection efficiency was calculated using a haemocytometer.
(3) Sample collection and storage:
*Centrifuge the remaining protoplasts from Step 13 at 300 × g for 2 min at room temperature;
*Remove the supernatant and freeze the samples in liquid nitrogen.
Extract RNA from the protoplasts
-
According to the manufacturer's instructions, total RNA from stem protoplasts was extracted using the Qiagen RNeasy Mini Kit (Qiagen). RNA concentration was determined by Nanodrop 2000 (Thermo Scientific), and the quality was examined using a Bioanalyzer 2100 (Agilent).
Subcellular localization
-
Subcellular localization analysis was performed using the pUC19-35S::GFP and pUC19-35S::HBA-1-MCherry vectors. The plasmids were extracted by the CsCl density-gradient ultracentrifugation method[6], and were transfected into the stem-differentiating xylem (SDX) protoplasts. For specific steps, refer to the sections of protoplast transfection and incubation, and protoplast culture, harvesting, and determination of transfection efficiency. The signal was captured using a Leica DM6 B microscope.
Statistical analysis
-
Statistical analysis was carried out using the SPSS software (v19.0). Data are presented as the mean ± standard error from three independent experiments.
-
To establish an efficient protoplast isolation protocol for H. hamabo, multiple plant tissues including leaves, petioles, and stems were tested for protoplast isolation (Figs. 1, 2; Supplementary Table S1). Leaf tissues are the predominant source materials. The transient expression system in Arabidopsis mesophylla protoplasts (TEAMP system) has a high cell yield of 10 × 106 (protoplasm volume per gram [g] of fresh leaf weight) and a transgene transfection efficiency of ~60%–90%[17]. According to the method described by Yoo et al.[17], intact protoplasts could not be isolated from the leaves and petioles of H. hamabo. On this basis, we attempted to separate the protoplasts of H. hamabo by adjusting the osmotic pressure to 0.5 M, washing, balancing the osmotic pressure to prevent cell rupture, and increasing the vacuum infiltration time to 1 h to enhance enzyme penetration (Supplementary Table S1). However, the cells isolated from leaves exhibited compromised integrity, severe aggregation, and were almost unusable (Fig. 1a; Supplementary Tables S1, S2). Although intact cells were isolated from the petioles, they were only sporadically dispersed and insufficient for subsequent experiments (Fig. 1b; Supplementary Tables S1, S2). This outcome is primarily attributed to severe enzymatic browning, coupled with excessive accumulation of polysaccharides, polyphenols, and abundant secondary metabolites.
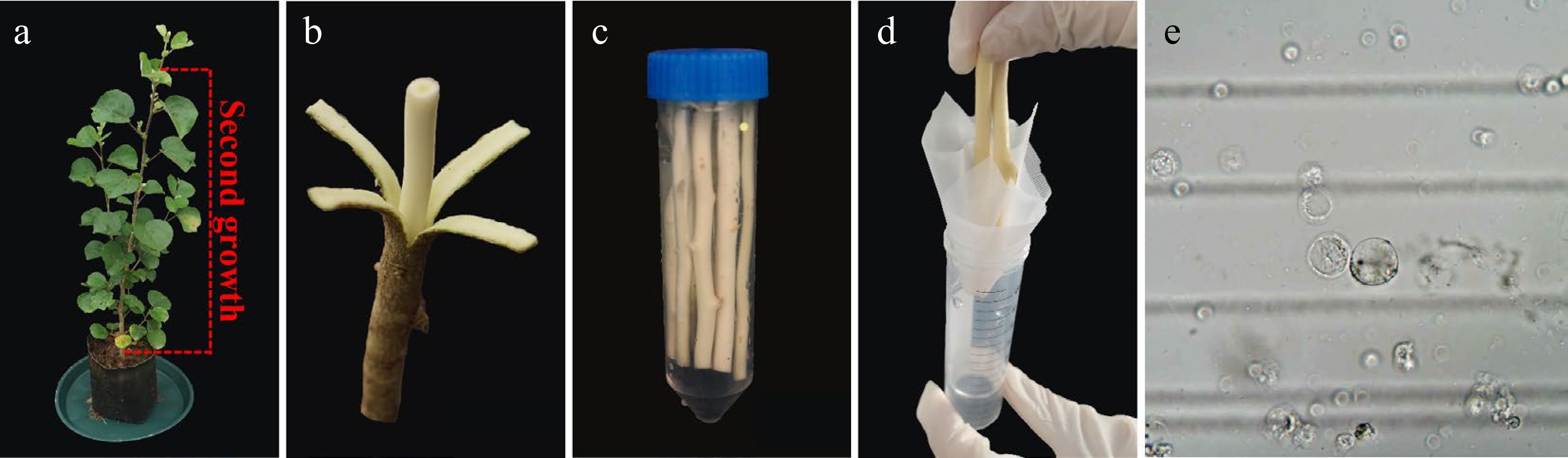
Figure 2.
H. hamabo protoplast isolation from wood-forming. (a) A healthy H. hamabo plant suitable for protoplast isolation. The dotted line indicates the optimal true stem (above the fourth stem segment) used as a xylem protoplast source. (b) Stem segments were cut into 10 cm segments to generate protoplasts from xylem cells. (c) The peeled stem segments were loosely submerged into the cell wall digestion solution in a 50 ml conical tube. (d) To obtain the purified protoplasts, we filtered the released protoplasts through a 75 µm filter membrane. (e) Stem protoplasts.
To further explore the method for protoplast isolation from H. hamabo, we selected stem segment tissues—which contain relatively low levels of polysaccharides and polyphenols—for subsequent analysis (Fig. 2; Supplementary Table S1). The Populus trichocarpa (P. trichocarpa) protoplast system is well known for the high yield (~2.5 × 107 protoplasts per g of stem-differentiating xylem), of protoplasts, which shares 96% transcriptome identity with the intact stem-differentiating xylem[6]. For woody plants, the separation of H. hamabo protoplasts on the basis of P. trichocarpa was attempted. The good news is that we obtained intact protoplasmic cells, albeit in very small numbers, and with a lot of impurities. On this basis, we optimized and improved the protoplast separation and transfection system suitable for H. hamabo. This systematic approach is based on xylem protoplasts freshly isolated from stems of H. hamabo wild-type plants growing in soil (Fig. 2a, b). To balance protoplast yield and viability, the enzymatic digestion time was extended (Fig. 2c) to promote sufficient contact between enzymes and the cell wall, thereby facilitating complete cell wall degradation and increasing protoplast release. Following enzymatic digestion, a significant amount of residual debris was generated. To mitigate the risk of clogging and reduce shear damage to protoplasts during filtration, a 75 μm nylon mesh was employed (Fig. 2d), thereby improving collection efficiency. Furthermore, based on the density difference between viable protoplasts and impurities, sucrose gradient density centrifugation was introduced to achieve precise separation. This step also removes residual enzymes and metabolic waste, further enhancing protoplast purity and transfection efficiency. In addition, after sucrose gradient centrifugation, the protoplasts were washed with mannitol to eliminate residual sucrose, thereby preventing cell rupture or shrinkage due to osmotic pressure differences between the sucrose solution and the subsequent culture system. In summary, we developed a highly efficient protoplast system perfectly adapted to H. hamabo. Complete protoplast cells were obtained through cell wall digestion, protoplast separation, washing, and filtration (Fig. 2c–e), yielding ~2.0 × 107 protoplasts per g of fresh xylem (Supplementary Table S2).
PEG-mediated protoplast transformation assay
-
To validate the practicality of the H. hamabo protoplast isolation system, we established a PEG-mediated transient transformation platform (see Methods 2.6 and 2.7 section). The balance between enzymatic digestion efficiency and cell viability is crucial for the success of protoplast experiments. Trypan blue staining revealed that, despite the enzymatic treatment, the cells remained highly active (93.61% ± 1.09%; Fig. 3; Supplementary Table S2). This demonstrates that the optimized enzymatic digestion conditions effectively degrade the cell wall while minimizing damage to the cell membrane. Regarding transformation efficiency, the pUC19-35s::GFP plasmid was delivered into protoplasts using PEG-mediated transfection, followed by overnight incubation under dark culture conditions to ensure sufficient transcription and translation. Fluorescence microscopy analysis revealed the transfection efficiency was 26.03% ± 1.07% (Fig. 4; Supplementary Table S2). Consistent replication across multiple trials confirms robust reproducibility and reliability of this platform, rendering it highly suitable for subsequent gene expression profiling and functional characterization studies.
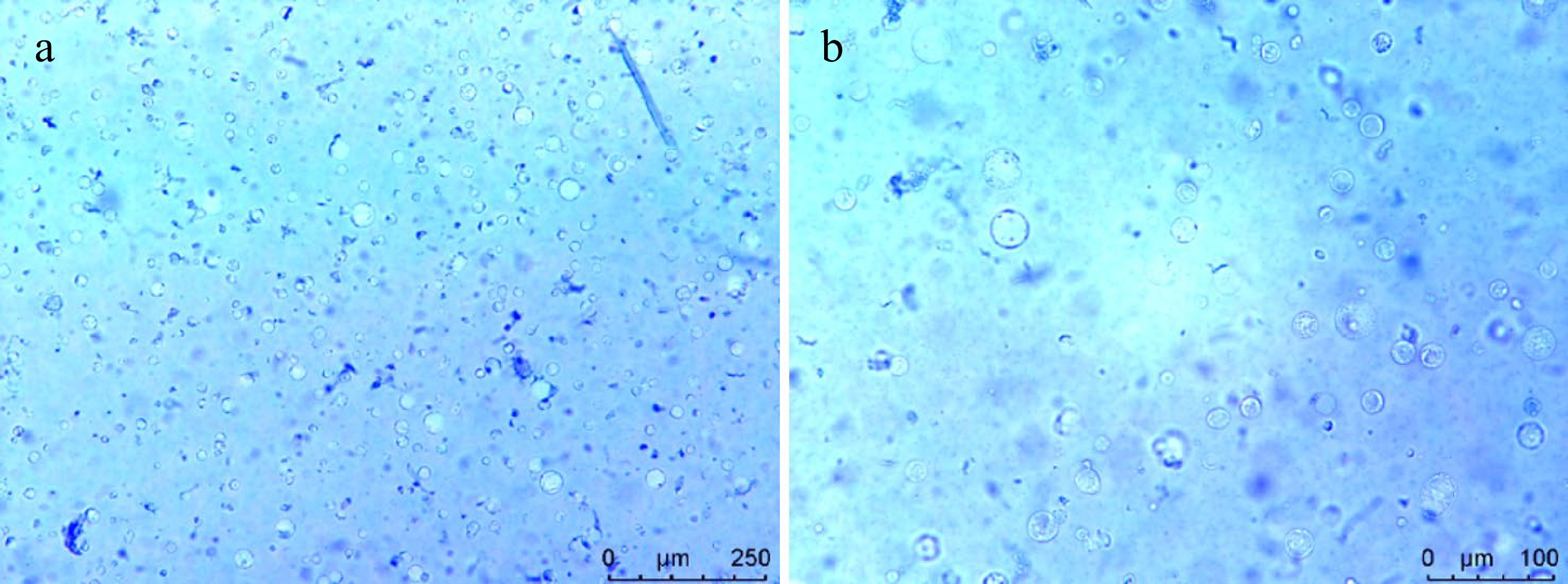
Figure 3.
Viability detection of the protoplasts isolated from H. hamabo by Trypan blue staining. Cell staining under (a) 10×, and (b) 20× microscope.
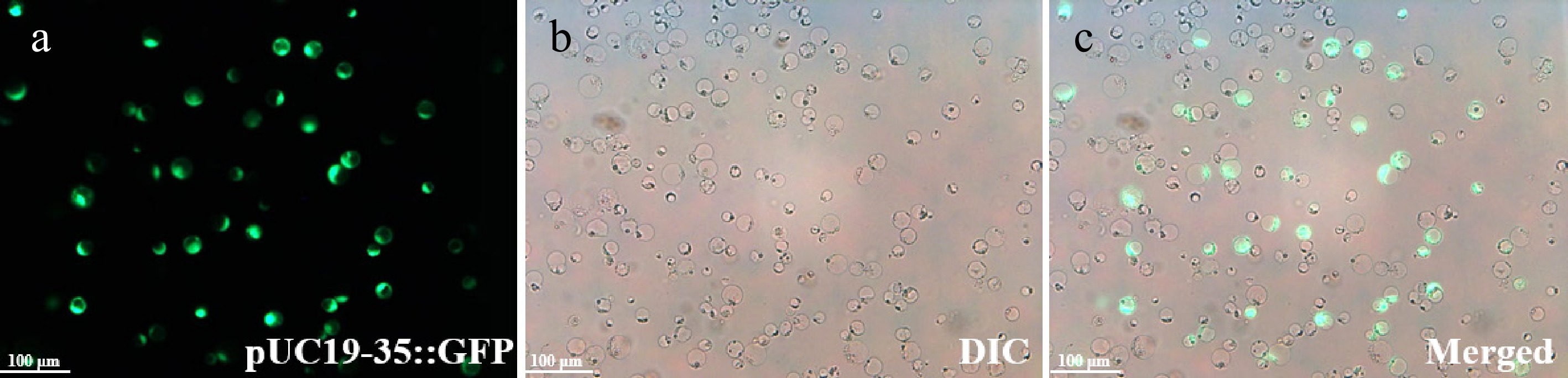
Figure 4.
Protoplasts expressing the pUC19-35::GFP are shown under fluorescence microscopy. The transfection efficiency was 26.03% ± 1.07%. H. hamabo protoplasts in the (a) fluorescence field, (b) bright field with DIC, and (c) a and b superimposed field. Bars = 100 μm.
Application of transient expression in H. hamabo protoplasts
-
As a single-cell system, the quality of RNA extracted from protoplasts directly influences the accuracy of downstream molecular experiments. In this study, total RNA was extracted from PEG-mediated transfected protoplasts of H. hamabo. The RNA quality was comprehensively assessed using a Nanodrop spectrophotometer, 1.2% agarose gel electrophoresis, and an Agilent 2100 Bioanalyzer (Fig. 5). The results showed that the extracted total RNA from protoplasts exhibited high purity: the A260/A280 ratio remained stable at around 2.0, and the A260/A230 ratios were all above 1.8, indicating minimal contamination from proteins, polysaccharides, or phenolic compounds (Fig. 5a). Agarose gel electrophoresis revealed clear and intact bands of 28S and 18S rRNA, with no apparent degradation or smearing (Fig. 5b). Analysis using the Agilent 2100 Bioanalyzer further confirmed high RNA integrity, with an RNA Integrity Number (RIN) is ~8.0, and an 28S/18S rRNA ratio close to 2 (Fig. 5c).
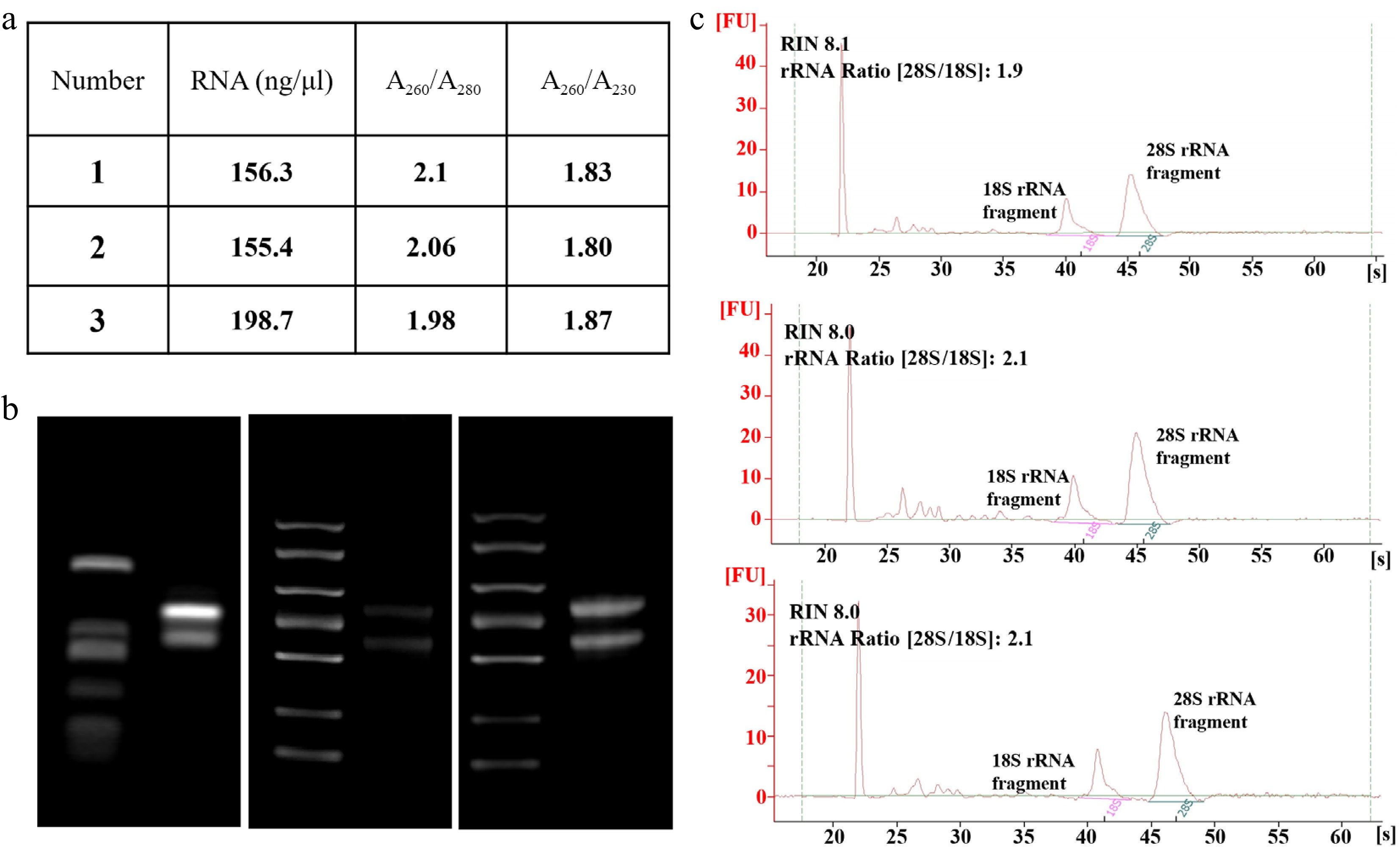
Figure 5.
RNA quality of SDX protoplasts. (a) Total RNA was isolated from stem protoplasts and the quality was examined using the Nanodrop spectrophotometer. (b) 1.2% agarose gel electrophoresis, and (c) Agilent 2100 Bioanalyzer.
Subcellular localization serves as a crucial prerequisite for deciphering gene function. Establishing a transient expression system using protoplasts as recipients represents a primary approach for studying protein subcellular localization and interactions. Using the PEG-mediated method, we transfected stem-derived protoplasts with the empty vector pUC19-35s::GFP and the pUC19-35S::H2A-1-mCherry vector (serving as a nuclear marker vector), respectively. The results demonstrated that the green fluorescent signal from the empty vector GFP control was uniformly distributed throughout the entire cell. In contrast, the red fluorescent signal from the pUC19-35S::H2A-1-mCherry protein was specifically concentrated at the nuclear envelope of the protoplasts, displaying distinct nuclear-localized fluorescent signals, and no significant fluorescence accumulation was observed in the cytoplasm or plasma membrane. These findings indicate that this protoplast transient expression system is suitable for precise subcellular localization analysis of target proteins in H. hamabo.
-
In recent years, protoplasts have been extensively utilized to identify key regulatory factors and elucidate the molecular mechanisms of the intracellular signal transduction pathways involved in plant physiology, immunity, growth, and development. Using protoplasts as a receptor system to establish transient expression is a primary method for subcellular localization studies[17,18]. Specifically, protoplasts isolated from multiple plant species—such as Arabidopsis thaliana, Nicotiana benthamiana, and Oryza sativa—have served as highly effective platforms for determining the subcellular localization of diverse protein families[18−25]. For example, RIN13 is localized in the nucleus, and its overexpression accelerates leaf senescence and programmed cell death[20]. PtrbHLH186 is a transcription factor located in the nucleus, and its overexpression affects the development of poplar xylem cells[22]. Leveraging the established H. hamabo protoplast system, this study successfully achieved subcellular localization of pUC19-35s::GFP and pUC19-35S::H2A-1-mCherry (Fig. 6). This functionally validates the system's capability to precisely resolve protein subcellular distribution. Furthermore, the developed H. hamabo protoplast-based localization platform provides a critical foundation for future gene-specific subcellular localization studies in this species, and serves as a reference paradigm for developing protoplast technologies in coastal woody plants.
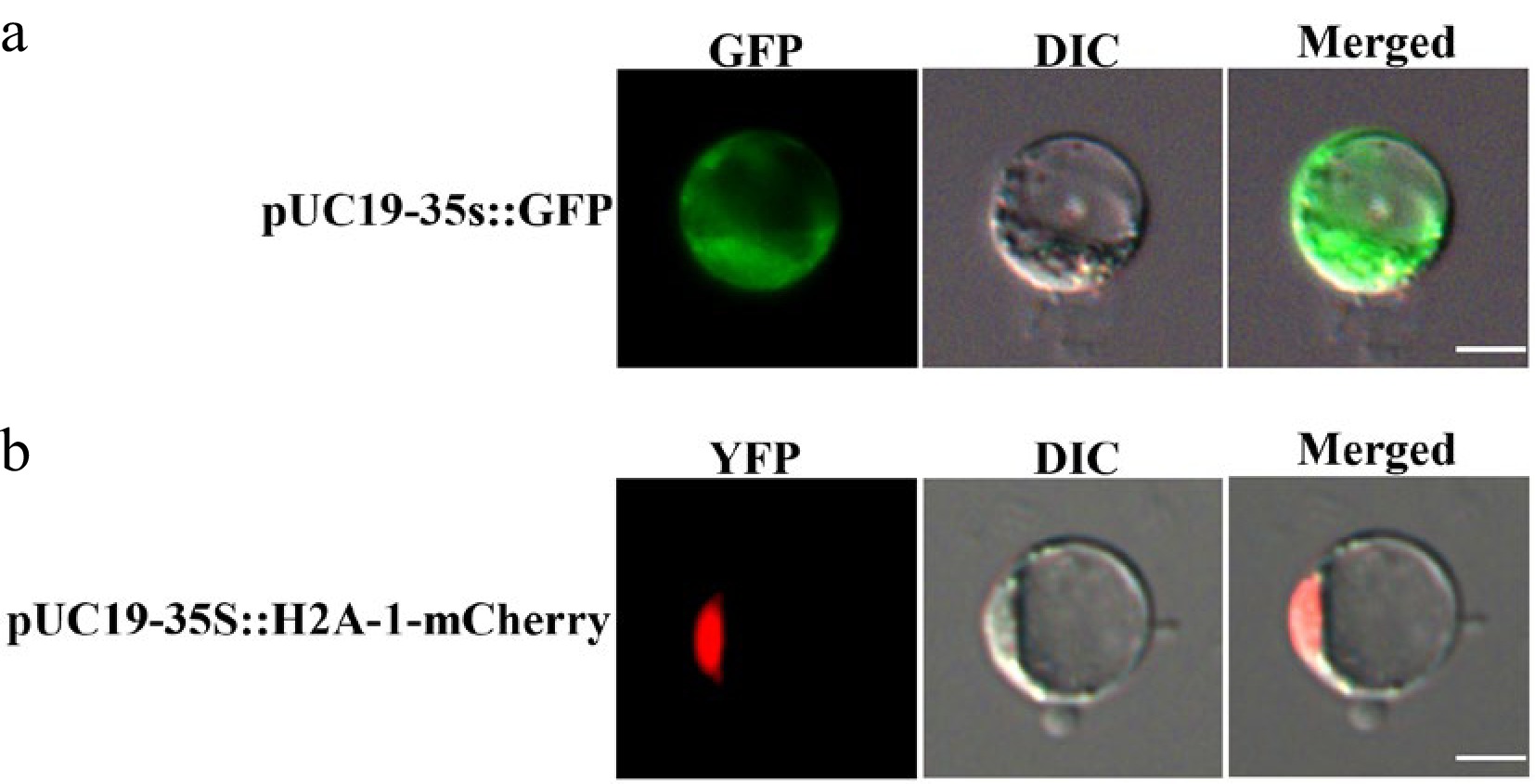
Figure 6.
Subcellular localization of empty vectors in H. hamabo protoplasts. (a) Fluorescence signal of pUC19-35S::GFP; (b) fluorescence signal of pUC19-35S::H2A-1-mCherry. GFP stands for green fluorescent protein and RFP stands for red fluorescent protein. Bars = 10 μm.
Protoplasts are used to rapidly verify gene function and study gene expression patterns and their biological functions through transient expression systems[22,26−29]. In Arabidopsis thaliana, the protoplast system was used to reveal the regulatory mechanism of EIN3 protein stability, and a significant crosstalk effect between glucose and ethylene signalling was found[26]. Abiotic and biotic stress signals can also be studied in protoplast systems. For example, the function of mitogen-activated protein kinase cascades in oxidative stress signalling was successfully demonstrated in Arabidopsis mesophyllum protoplasts[27]. Utilizing this system in this study, we consistently isolated high-quality RNA exhibiting excellent purity and integrity, with A260/A280 ratios stably maintained at around 2.0 and no detectable degradation (Fig. 5). This RNA quality fully meets the stringent technical requirements for downstream transcriptional regulation studies. The RNA extracted via this platform enables comprehensive analysis of transcriptional expression patterns of key genes under abiotic stresses (e.g., salinity) and facilitates the validation of regulatory relationships between transcription factors and their target genes. This will provide precise data support for elucidating the molecular mechanism of the stress tolerance traits of H. hamabo. Furthermore, the establishment of the H. hamabo protoplast system provides a vital tool for molecular biology research in this species, offering broad application prospects. For protein-protein interaction studies, bimolecular fluorescence complementation (BiFC) serves as a classical methodology for validating protein interactions[22,23,30]. Drawing on this technical approach, the H. hamabo protoplast system established in this study can be used to screen and verify the interaction networks of key proteins in pathways related to stress resistance.
Protoplast systems facilitate the mechanistic dissection of epigenetic regulation by multicomponent complexes. For instance, studies have utilized this system to demonstrate how transcription factors like PtrAREB1-2 recruit histone-modifying complexes to activate drought-responsive genes[31−33]. Furthermore, protoplast assays have revealed how proteins such as PtrVCS2 dynamically regulate the interaction intensity of histone acetylase complexes to control developmental processes like vascular cambium formation[34]. Therefore, the H. hamabo protoplast system established in this study enables us to conduct in-depth research on the epigenetic regulatory mechanisms related to stress responses. Recently, protoplast LUC assays verified direct gene regulation by transcription factors and synergistic complexes[30,35]. Moreover, isolating protoplasts in combination with fluorescence-assisted cell sorting or scRNA-seq enables genome-wide elucidation of cell type-specific transcriptional profiles and high-resolution single-cell maps[36,37].
While this system offers significant advantages, several limitations warrant further refinement in subsequent studies. The established platform constitutes a transient expression system, wherein exogenous genes exhibit only ephemeral expression in protoplasts without achieving stable genetic transformation. Consequently, this approach is unsuitable for the long-term verification of gene functions or generating transgenic plants. Furthermore, the current methodology yields exclusively free protoplasts, but has not attained callus induction or plant regeneration—a pervasive challenge in woody plant protoplast technology that severely constrains its utility in stable genetic manipulations. To address these constraints, future research should optimize culture media composition, hormonal ratios, and incubation conditions to induce protoplast division and callus formation, ultimately pioneering whole-plant regeneration to overcome the bottleneck of stable genetic transformation.
-
This study successfully established a comprehensive technical system for H. hamabo, encompassing highly efficient protoplast isolation, PEG-mediated transformation, and downstream molecular applications. The system consistently achieved protoplast viability 93.61% ± 1.09%, and transformation efficiency stabilized at 26.03% ± 1.07%. Furthermore, the transient expression system established for H. hamabo protoplasts demonstrates high viability and stability, while facilitating efficient extraction of high-integrity RNA for downstream molecular quantification assays. Critically, it enables precise subcellular localization of target proteins. This multifaceted validation confirms its suitability as an ideal platform for functional characterization of H. hamabo genes, thereby providing a robust technological foundation for subsequent investigations into gene expression regulation, protein-protein interactions, and stress adaptation mechanisms.
-
The authors confirm their contributions to the paper as follows: study conception and design: Liu X, Chen Q, Liu H; data collection: Liu H, Zhang X, Wang J, Gao J, Yue L, Liang J, Lu X, Ma Y; analysis and interpretation of results: Liu H, Yang S, Zhang X, Wang J; draft manuscript preparation: Liu H, Yang S. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available from the corresponding author upon reasonable request.
-
This work was supported by the Key Scientific and Technological Grant of Zhejiang for Breeding New Agricultural Varieties (Grant No. 2021C02070-6), the Wenzhou Forestry and Flower Breeding Cooperation Group Project (Grant No. ZX2024004-1), the Wenzhou High-level Innovation Team 'Coastal Characteristic Plant Innovation and Utilization Project' (Grant No. NY202401), and the Open Project of the Engineering Research Center for Southeast coastal characteristic plants of the National Forestry and Grassland Administration (Grant No. LCDNZX2025002).
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/fra-0026-0004.
-
#Authors contributed equally: Huizi Liu, Sheng Yang
- Supplementary Table S1 Comparative optimization of protoplast isolation methods from different tissues of H. hamabo.
- Supplementary Table S2 Effects of different tissues of H. hamabo on protoplast yield and vitality.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu H, Yang S, Zhang X, Wang J, Gao J, et al. 2026. Hibiscus hamabo protoplast cell system for transient gene expression analysis. Forestry Research Advances 1: e007 doi: 10.48130/fra-0026-0004 

Hibiscus hamabo protoplast cell system for transient gene expression analysis
- Received: 20 November 2025
- Revised: 27 March 2026
- Accepted: 30 March 2026
- Published online: 22 June 2026
Abstract: Hibiscus hamabo, an endangered semi-mangrove species with critical ecological functions in coastal protection and biodiversity maintenance, faces research limitations due to unstable genetic transformation systems. Protoplasts, as a transient expression system, have proven crucial in multiple fields of research such as plant genomics, transcriptomics, genetic transformation, physiology, and biochemistry. Establishing an efficient protoplast separation system and transient transformation system is the basis for conducting molecular biology and related research. In particular, woody plants are highly valuable because of the difficulty in constructing a stable transgenic system. Here, we establish a robust protoplast isolation and transfection protocol using stem tissues, filling the gap in protoplast technology for Hibiscus hamabo. This approach includes tissue preparation, protoplast isolation, protoplast transfection, and incubation. The protoplast yield reached ~2.0 × 107 per g, and the protoplast activity reached 93.61% ± 1.09%. The establishment of this cell system can immediately trigger transcriptome responses to gene expression to characterize and quantify genetic networks and gene functions, enabling rapid and efficient flux experiments. This study provides a reference for protoplast research on other difficult-to-transform woody plants, and offers guidance for further analysis of plant growth/development and genetic improvement.
-
Key words:
- Hibiscus hamabo /
- Stem segment /
- Protoplast /
- Transient expression