









-
Plants are capable of directly absorbing and utilizing amino acids[1,2]. In this process, amino acid transporters (AATs) play pivotal roles in the translocation of amino acids in plant tissues[3,4]. To date, AATs have been identified in a wide range of plant species[5], including rice[6−8], quinoa[9], pepper[10], potato[11], and poplar[12]. The AAT gene superfamily comprises amino acid/auxin permease (AAAP), amino acid-polyamine-choline (APC), and usually multiple amino acids move in and out transporter (UMAMIT) families. The AAAP family is further classified into amino acid permease (AAP), aromatic and neutral amino acid transporter (ANT), amino acid transporter-like (ATL), auxin transporter (AUX), γ-aminobutyric acid transporter (GAT), lysine and histidine transporter (LHT), proline transporter (ProT), and threonine transporter protein (TTP) subfamilies[13−15]. The APC family is further classified into amino acid/choline transporter (ACT), cationic amino acid transporter (CAT), and polyamine H+-symporter (PHS) subfamilies[11,16,17]. Only a few studies have focused on the UMAMIT family, and the understanding of its origin and structure is limited[18].
The specific functions of AATs have been investigated and validated in Arabidopsis thaliana[17]. AAPs are mainly responsible for the transport of neutral and acidic amino acids and generally exhibit medium-affinity transport properties. Specifically, AtAAP1 is expressed in roots and endosperm tissues, where it facilitates the transfer of amino acids into root or endosperm cells[14]. AtAAP2 and AtAAP6 are localized in phloem companion cells and xylem parenchyma cells, respectively, and they coordinately transport amino acids from the xylem to the phloem[19]. AtAAP3 and AtAAP5 are capable of transporting basic amino acids[20]. AtAAP4 is predominantly expressed in leaves, stems, and floral organs, and functions to promote amino acid allocation to developing embryos. AtAAP5 is expressed in the root cortex, where it contributes to amino acid uptake from the soil, and is also expressed in leaves, stems, and flowers during later developmental stages[21]. AtAAP8 is expressed in seeds 2–4 d after fertilization, in the floral veins of mature siliques, and plays an important role in developing embryos by supplying amino acids[22]. LHTs mediate the transport of lysine, histidine, and neutral and acidic amino acids[23]. The AUXs represent the major auxin influx carriers, with AtAUX1 shown to regulate root gravitropism and root hair development[24]. AtANT1, an aromatic and neutral amino acid transporter, is expressed mainly in flowers[25]. The ProTs are responsible for the transport of multiple substrates, including proline, glycine betaine, and γ-aminobutyric acid. Among them, AtProT1 rapidly transports proline under water stress conditions, thereby reducing plant damage, whereas AtProT2 plays an important role in transporting compatible solutes in the root tip region[26].
In faba bean (Vicia faba), the transporters VfAAP1 and VfAAP3 have been shown to mediate the uptake and translocation of multiple amino acids. Specifically, VfAAP1 has a selective preference for cysteine, whereas VfAAP3 has a specific affinity for lysine and arginine[27]. In potato (Solanum tuberosum), StAAP1 is expressed predominantly in leaves, and suppression of its expression leads to a reduction in amino acid content[28]. In common bean (Phaseolus vulgaris), PvAAP1 is expressed in xylem parenchyma cells, epidermal cells, and phloem tissues, where it plays a crucial role in the lateral transfer of amino acids[29,30]. In poplar (Populus trichocarpa), PtAAP11 contributes to xylem development by transporting proline, highlighting its importance in wood formation[31].
Light is a key environmental factor influencing vegetable quality and antioxidant capacity[32,33]. It has been continuously studied, particularly in plant factory systems[34]. Importantly, continuous lighting disrupts the circadian clock and thus has a possible role in the accumulation of metabolites[35,36]. Red (R) and blue (B) light are the most influential factors for photosynthesis. They trigger distinct signaling cascades, modulating gene expression profiles associated with resource allocation and growth patterns[37]. Therefore, it is of interest to investigate genes that are regulated by the light.
Lettuce (Lactuca sativa L.) is an important leafy vegetable with high horticultural value and rich nutritional attributes, making it a key crop in controlled environment agriculture[38,39]. Its growth and nutritional quality are sensitive to amino acid application. For instance, our previous studies have revealed that lettuce supplied with glycine improves nutritional quality without loss of yield[40] and enhances glycosylated flavonoids content and antioxidant activity[41]. It is important to further study the genes that are involved in the absorption and transportation of amino acids.
In recent years, with the rise of organic agriculture, researchers have focused on amino acids and other organic molecules in soils[42]. Investigations into the molecular mechanisms underlying their transport have become a major focus in plant nutritional biology. Amino acid absorption and utilization significantly affect the growth and nutritional quality of lettuce[40,43]. However, AATs in lettuce have not been characterized. In this study, we performed genome-wide identification and analysis of the AAT gene family in lettuce, aiming to provide a foundation for elucidating the transport mechanisms of amino acids and other small molecules.
-
The lettuce genome sequence was obtained from the Phytozome database (
http://phytozome.jgi.doe.gov/pz/portal.html ). Hidden Markov Model profiles of the AAT domains (PF01490 and PF00324) were retrieved from the Pfam database[44,45], and employed in HMMER v3.0 to identify all potential AAT transcripts in lettuce. In addition, amino acid sequences of AATs from Brassica rapa and Brassica oleracea were used as queries for BLASTP searches against the lettuce genome. The resulting candidate transcripts were further validated via InterProScan (www.ebi.ac.uk/interpro/scan.html ) to confirm the presence of AAT domains. Conserved domain analysis was performed on the identified lettuce AAT transcripts (LsAATs), and sequences lacking PF01490 or PF00324 domains were excluded. The physicochemical properties of the filtered transcripts, including molecular weight (MW), isoelectric point (pI), and protein length, were analyzed via ProtParam (https://web.expasy.org/protparam ). Furthermore, the transmembrane domain (TM) was predicted with TMHMM v2.0 (www.cbs.dtu.dk/servicesTMHMM ).Chromosomal localization and genomic duplications of LsAATs
-
The chromosomal positions of the LsAATs were obtained via Phytozome v14 (
http://phytozome.jgi.doe.gov/pz/portal.html ). The identified LsAATs were subsequently mapped onto lettuce chromosomes via the MG2C_v2.1 online tool (http://mg2c.iask.in/mg2c_v2.1 ). Collinearity analysis of the LsAATs were analyzed using the MCScanX module integrated in TBtools. The collinear relationships of LsAAT genes were further visualized by constructing a Circos plot[46].Calculation of the Ka/Ks and duplication event
-
The coding sequences (CDS) of duplicated gene pairs identified by MCScanX were extracted, and their corresponding protein sequences were retrieved from Phytozome v14. The CDS and protein sequences were then subjected to KaKs_Calculator 2.0 to estimate the nonsynonymous substitution rate (Ka) and synonymous substitution rate (Ks)[47]. The Ka/Ks ratios were calculated to assess the selection history. Divergence time was calculated as follows: T = Ks/(2 × 6.1 × 10−9) × 10−6 million years (MY).
Phylogenetic analysis of the LsAATs
-
Phylogenetic analysis was performed on the basis of the full-length amino acid sequences of AAT proteins from Arabidopsis thaliana (AtAATs) and Phaseolus vulgaris (PhAATs). Multiple sequence alignment of all retrieved sequences was conducted with the ClustalW program implemented in MEGA 11 via default parameters[48]. Phylogenetic trees were then constructed via the Neighbor-Joining method[49]. The JTT+I+G substitution model was applied, and 1,000 bootstrap replicates were performed under default settings. The resulting phylogenetic tree was visualized via the iTOL platform (
https://itol.embl.de ).Conserved motif analysis of the LsAATs
-
The conserved motifs of the lettuce LsAATs were analyzed via the MEME 5.5.8 online tool (
https://meme-suite.org/meme/tools/meme )[50]. Furthermore, the evolutionary relationships among the LsAATs were examined via MEGA 11.0, and the conserved motifs of the candidate AAT proteins were graphically visualized with TBtools software.Analysis of cis-acting elements in the promoter regions of the LsAATs
-
The 2,000 bp upstream genomic sequences of the LsAATs were extracted via TBtools software. These sequences were then analyzed with PlantCARE (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html ) to predict cis-acting regulatory elements. The predicted elements were subsequently visualized via TBtools.Continuous lighting and light quality treatments
-
The experiments were conducted in a controlled growth chamber. Lettuce plants were cultivated for 37 d after sowing, corresponding to a fresh weight of 80 g at harvest stage. They were subjected to continuous white light irradiation at 150 μmol·m−2·s−1 for zero (0D), one (1D), two (2D), three (3D), four (4D), or five days (5D) before harvest. In addition, R (peak at 660 nm), B (peak at 450 nm), and R:B = 1:1 (RB) lights at 200 μmol·m−2·s−1 were applied to the lettuce for 6 d before harvest. LED lights were used for all the treatments. The temperature was 22 ± 2 °C, and the relative humidity was 60%–70%. The newest three fully expanded leaves were collected for RNA-seq analyses, which were performed on the Illumina NovaSeq 6000 Sequencing System with a sequencing depth of 2.5×.
Analysis of light-regulated LsAATs and quality-related indices
-
The Pheatmap and Mfuzz R packages were used to draw the heatmap and categorize the expression pattern of differentially expressed genes, respectively. To validate the sequencing data, qRT-PCR was performed using the CFX Connect Real-Time PCR Detection System (Bio-Rad Laboratories, Singapore). All reactions were performed in triplicate. The relative expression level was normalized by the 2−ΔΔCᴛ method, and ACTIN was used as the reference. The quality-related indices, including total flavones, polyphenols, and phenolic acids, were measured according to our previous report[51]. In brief, the phenolic compounds were extracted using acidified methanol. The contents of total flavones, polyphenols, and phenolic acids were calculated as gallic acid equivalents (GAE), quercetin equivalents (QE), and caffeic acid equivalents (CAE), respectively.
Three-dimensional structure prediction
-
The three-dimensional structures of the LsAATs were predicted via SWISS-MODEL (
https://swissmodel.expasy.org ). The amino acid sequences were submitted to the platform for sequence alignment, and homologous protein templates with confidence scores greater than 30% were selected. These templates were then used to predict and construct the three-dimensional structures. -
A total of 180 and 1,246 transcripts were identified via HMMER and BLASTP, respectively. After InterProScan searches for AAT domains, 225 transcripts remained. Following the removal of incomplete and redundant sequences, 102 transcripts were ultimately retained. To classify the transcripts, phylogenetic analysis was performed on the basis of their amino acid sequences together with AtAATs and PhAATs (Fig. 1). The lettuce AAT genes were renamed according to their phylogenetic relationships. The characteristics of the LsAATs are listed in Supplementary Table S1. The number of exons ranged from 1 to 16, with CDS lengths varying from 225 to 2,925 bp, corresponding to predicted protein lengths of 74 to 974 amino acids. The theoretical isoelectric points ranged from 4.53 to 9.79, and the molecular weights varied from 8.25 to 106.85 kDa. Approximately 64% of the LsAATs contained nine to 11 TMs, whereas 15% of the members possessed 12 or more TMs. Similar distributions were observed in several subfamilies, such as LsANT, LsATLa, and LsATLb. In addition, no TMs were predicted by TMHMM for LsATLa1, LsATLa2, or LsAUX1.
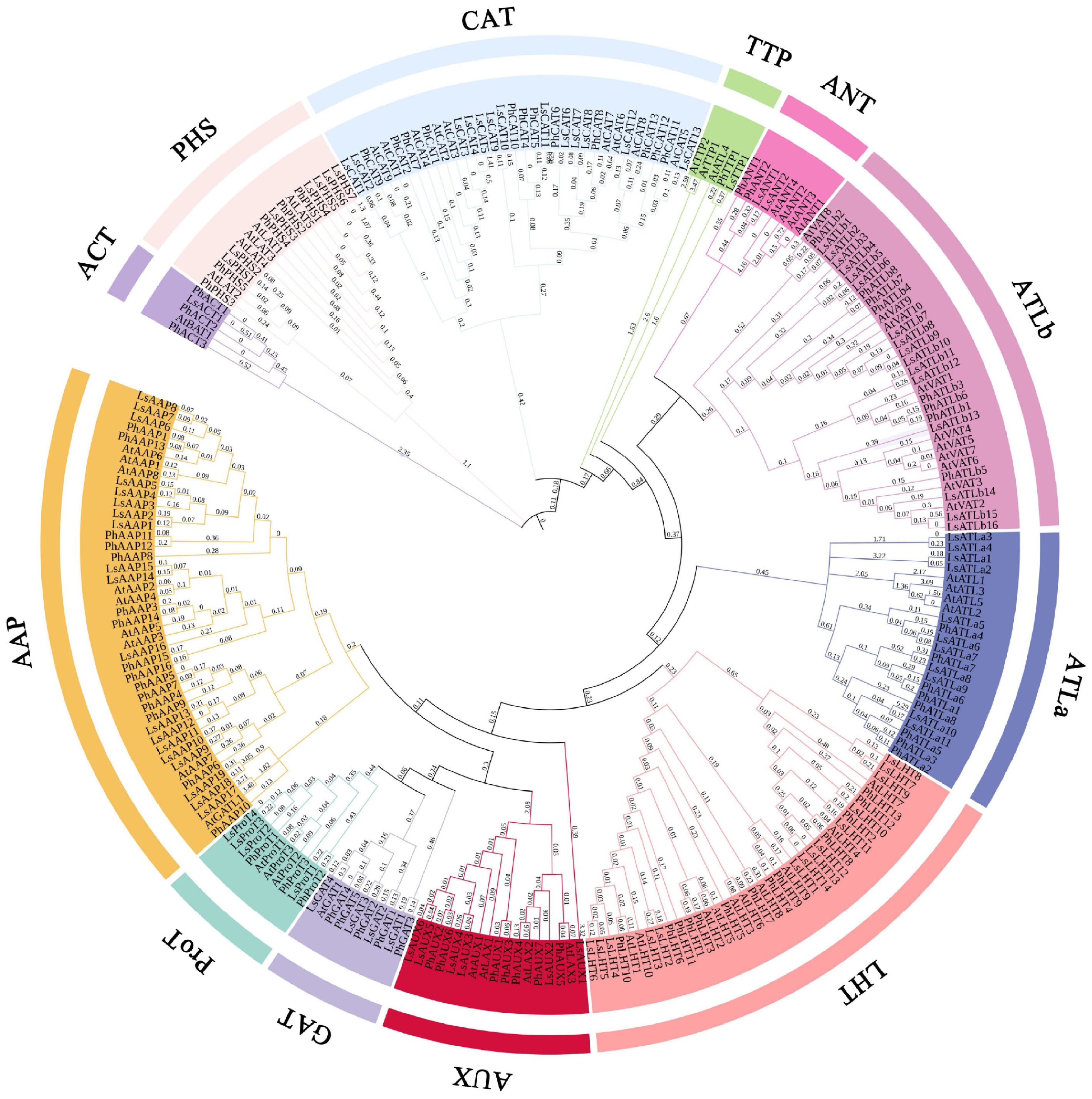
Figure 1.
Phylogenetic relationships among lettuce, Arabidopsis, and kidney bean. The tree is divided into 12 subgroups. Different groups are shown in different colors. Numbers near the nodes represent bootstrap support (1,000 iterations).
Chromosomal localization and duplication analysis of the LsAATs
-
The chromosomal locations of the LsAATs were identified and mapped onto a schematic representation of the chromosomes (Fig. 2a). The LsAATs were randomly distributed across nine chromosomes. Chromosomes 4 and 9 contained the greatest number of genes, with 16 each. There were 11 genes located on chromosomes 3, 5, and 8. In addition, chromosomes 1, 2, 6, and 7 possessed five, 13, nine, and six genes, respectively. Tandem duplications were detected on all chromosomes except chromosomes 1, 2, and 8 (Fig. 2a). The 18 tandemly duplicated genes belonged to five subfamilies. In detail, AAP comprised six genes, ATLa comprised two genes, ATLb comprised five genes, LHT comprised three genes, and PHS comprised two genes. Segmental duplications were identified across all chromosomes (Fig. 2b). The 23 segmentally duplicated genes belonged to five subfamilies. In detail, AAP comprised five genes, ATLa comprised two genes, ATLb comprised five genes, CAT comprised six genes, LHT comprised three genes, and ProT comprised two genes. LsATLb6, LsATLb8, LsAAP5, LsAAP4, and LsAAP12 were both tandem and segmental duplications.
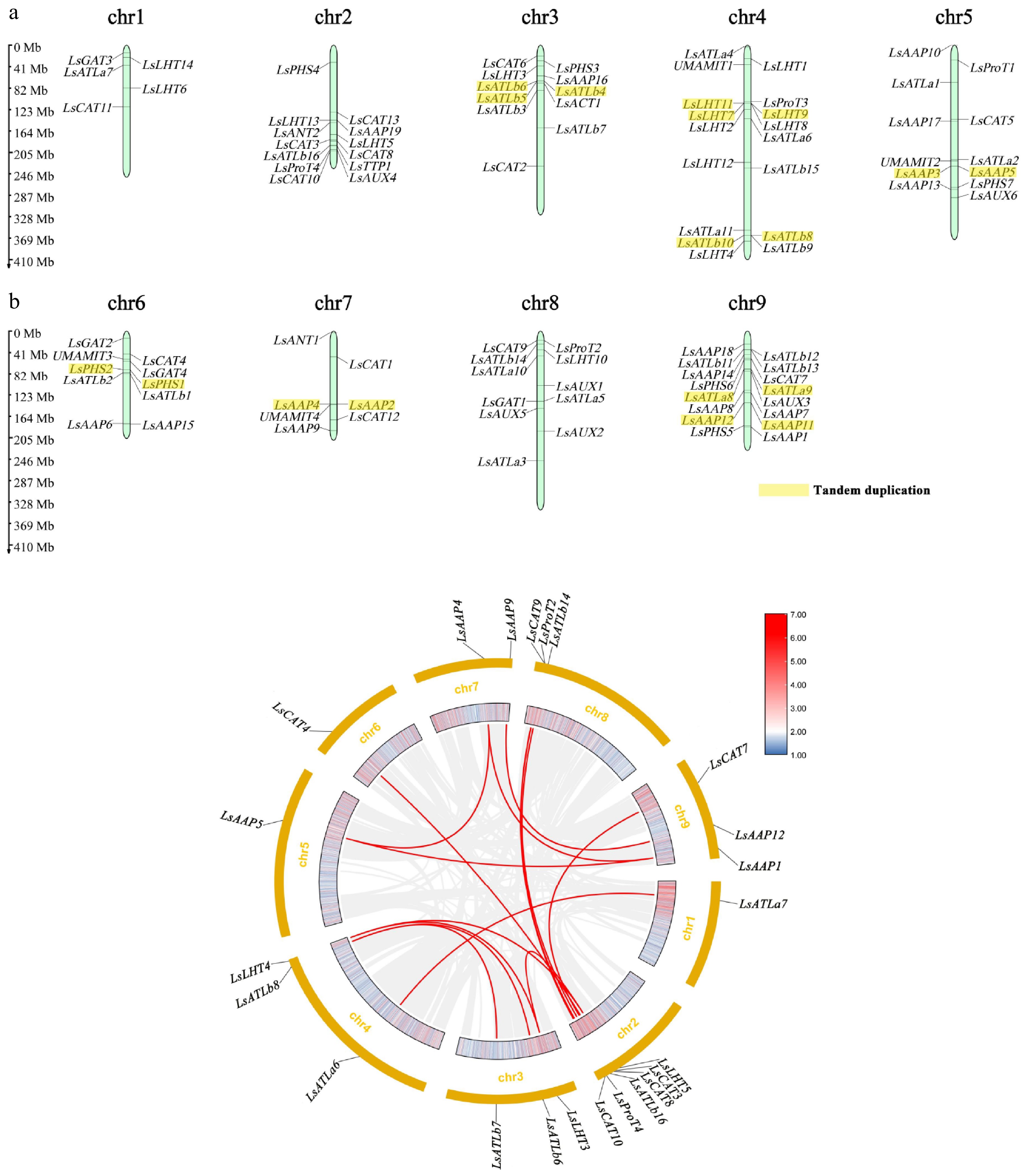
Figure 2.
Chromosomal localization and collinearity analysis of LsAATs. (a) Distribution of LsAATs on chromosomes. Tandem duplicated genes are marked in a yellow background. (b) Collinearity analysis of LsAATs. The outer ring depicts the nine lettuce chromosomes. Heatmap bars within the circle represented gene density in each chromosome, using a colour gradient from blue (low density) to red (high density). Red curved lines highlight the segmental duplicated genes.
Analysis of the paralogous gene pairs revealed that all Ka/Ks ratios were less than 1, indicating that these duplicated genes had predominantly undergone purifying selection (Table 1). The estimated divergence times ranged widely from approximately 63 to 198 MY ago, suggesting that multiple duplication events contributed to the expansion of the LsAATs. Notably, pairs such as LsLHT3-LsLHT4 and LsAAP1-LsAAP5 showed relatively ancient duplication dates (> 170 MY), whereas LsCAT7-LsCAT8 represented more recent events (~ 63 MY).
Table 1. Selection history and divergence between paralogous gene pairs.
Paralogous pair Ka Ks Ka/Ks Duplication date (MY) 1 LsAAP1-LsAAP4 0.203 1.533 0.132 125.61 2 LsAAP1-LsAAP5 0.221 2.229 0.099 171.472 3 LsAAP4-LsAAP5 0.162 0.966 0.168 79.165 4 LsAAP9-LsAAP12 0.356 1.938 0.184 149.076 5 LsATLa6-LsATLa7 0.167 1.321 0.126 101.585 6 LsATLb6-LsATLb8 0.341 2.143 0.159 164.855 7 LsATLb7-LsATLb8 0.214 1.396 0.153 107.395 8 LsATLb14-LsATLb16 0.236 0.768 0.307 62.964 9 LsCAT3-LsCAT4 0.260 1.227 0.212 94.400 10 LsCAT7-LsCAT8 0.118 0.848 0.139 65.237 11 LsCAT9-LsCAT10 0.182 1.666 0.109 128.176 12 LsLHT3-LsLHT4 0.201 2.570 0.078 197.688 13 LsLHT3-LsLHT5 0.216 2.079 0.104 170.423 14 LsLHT4-LsLHT5 0.084 1.046 0.081 85.737 15 LsProT2-LsProT4 0.125 0.974 0.129 79.832 Conserved motifs of the LsAATs
-
Thirty conserved motifs of LsAATs were identified (Fig. 3). The results revealed that the same subfamily presented similar motif constitutions. With the exception of the PHS, ACT, and CAT subfamilies, the other subfamilies contained Motif 1 and Motif 3. Large proportions of LsAATs contained Motif 2 and Motif 6, accounting for 52% and 56%, respectively. Motifs 8 and 14 were unique to the LHT and ATLb subfamilies, respectively. In addition, motifs 16 and 19 were found only in the AAP subfamily.
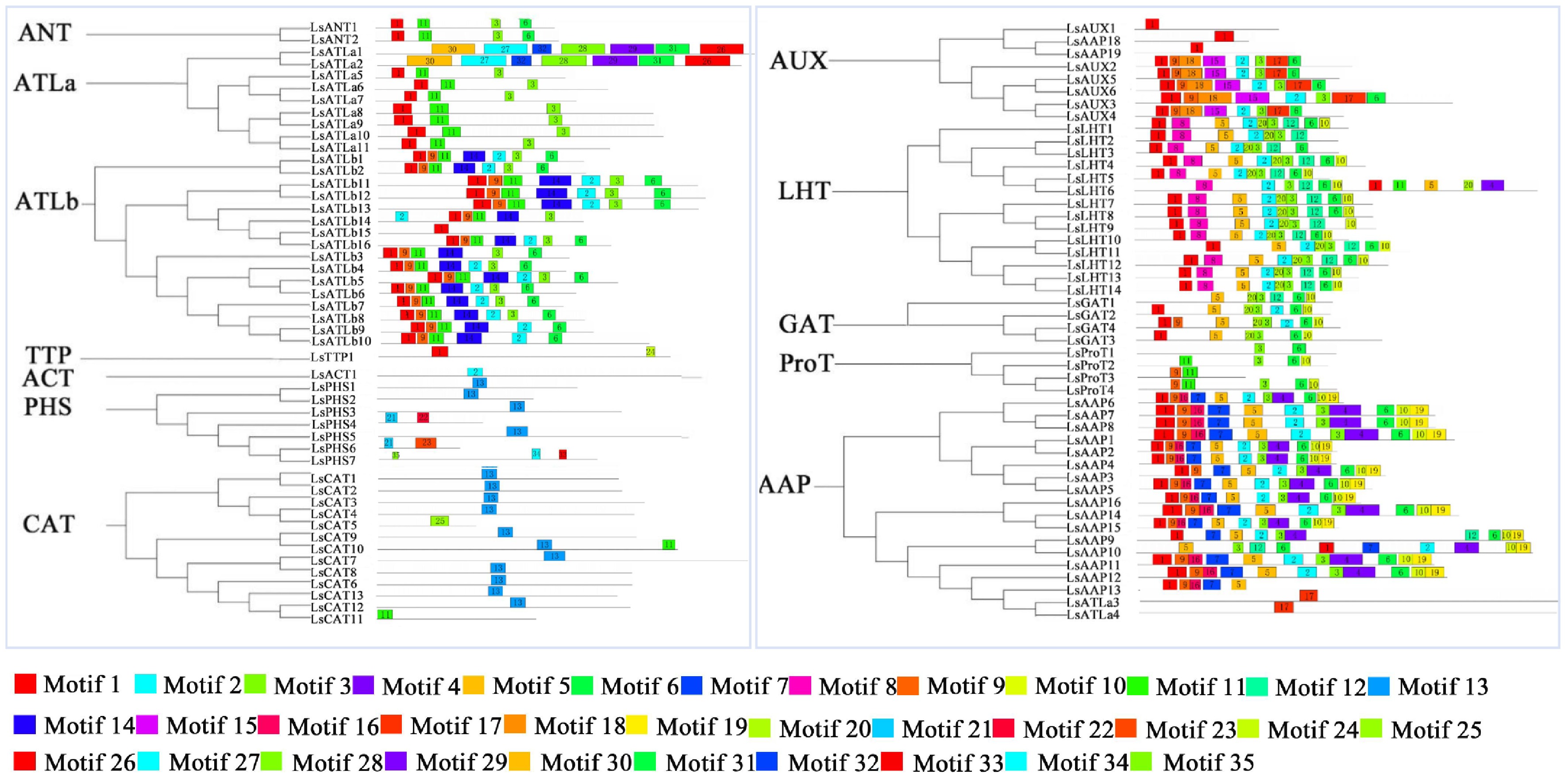
Figure 3.
Phylogenetic relationships and conserved motif architecture of LsAAT proteins across subfamilies. Different colored boxes represent specific motifs. The distribution of the motifs correspond to their position.
Cis-acting element analysis of the LsAATs promoters
-
The cis-acting elements were analyzed to investigate the potential regulatory elements of LsAATs (Fig. 4). The results revealed that the promoter regions of LsAATs were enriched with diverse cis-regulatory elements, including those related to light, hormones, and stress responsiveness. Light-responsive elements were present in 99% of the LsAATs. There were 74, 47, 40, 40, and 37 LsAATs containing cis-elements responsive to methyl jasmonate, auxin, abscisic acid, salicylic acid, and gibberellin, respectively. Low-temperature responsive elements were present in 45 LsAATs. Defense- and stress-responsive elements were identified in 30 members. These findings suggest that the LsAATs are regulated by light, hormones, and various stress-related factors.

Figure 4.
Analysis of cis-acting elements in the promoters of LsAATs. The cis-regulatory elements were classified into light responsiveness, MeJA-responsiveness, auxin responsiveness, abscisic acid responsiveness, salicylic acid responsiveness, gibberellin-responsiveness, low-temperature responsiveness, defense and stress responsiveness, wound-responsiveness, and meristem expression.
Light regulation of LsAATs and quality-related indices
-
On the basis of the cis-acting element analysis of the LsAATs, 99% of them were light-responsive. To further investigate the expression patterns of LsAATs regulated by light, continuous lighting and light quality treatments were performed. As shown in Fig. 5a, continuous lighting had a significant effect on the expression of LsAATs. Using the Mfuzz algorithm, the expression patterns of LsAATs were clustered into eight groups. Those in clusters C4 and C8 showed great changes on 1D. Among them, members of the LsATLa subfamily accounted for 30% in C4. And members of the LsAAP subfamily accounted for 45% in C8. Compared with the control, total flavone content significantly decreased on 1D, and then increased to the highest level on 3D (Fig. 5b). Total polyphenols and phenolic acid content reached a minimum on 1D, followed by an increase on 2D (Fig. 5c, d). Based on previous reports, the expression levels of genes involved in the potential regulatory pathways were screened. It showed that HY5, BIN2, and DELLA were significantly enhanced under continuous lighting (Fig. 5e). In addition, six LsAATs were randomly selected for qRT-PCR validation. The results showed that the sequencing data were valid (Supplementary Fig. S1).
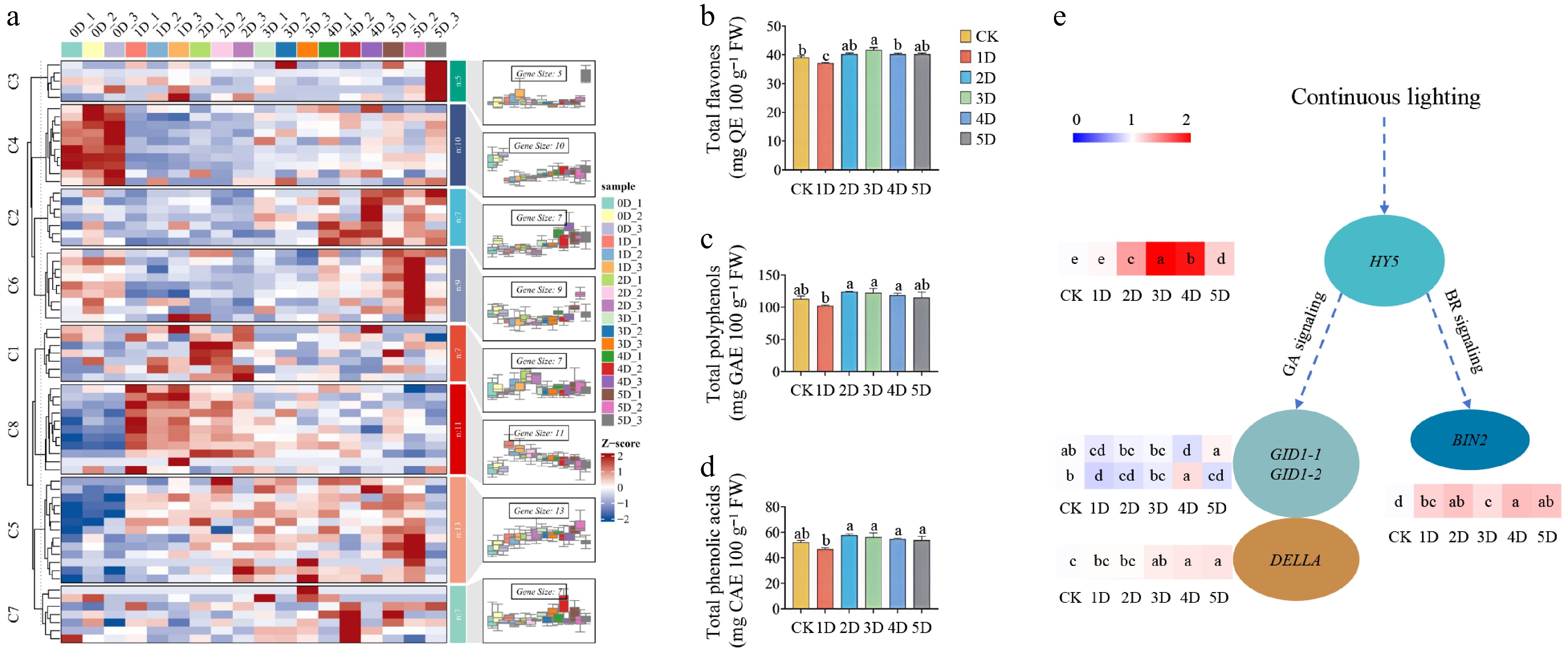
Figure 5.
Regulation of genes and quality-related indices under continuous lighting for 1 to 5 d before harvest. (a) Expression patterns of LsAATs. (b) Content of total flavones. (c) Content of polyphenols. (d) Content of phenolic acids. (e) Expression levels of genes involved in the potential regulatory pathways. High and low expression levels of genes are labeled in red and blue colors, respectively. The expression patterns of LsAATs were clustered into eight groups, and genes in different groups are provided in Supplementary Table S2.
The expression patterns of LsAATs were also investigated under different light qualities. As shown in Fig. 6a, a large proportion of genes had significantly higher expression levels under R treatment than under the B and the RB treatments. Thirty-nine percent of the genes were from the LsATLa and LsATLb subfamilies, and their expressions were negatively correlated with the contents of quality-related indices (Fig. 6b-d). Four genes in the LsAAP subfamily (LsAAP6, LsAAP9, LsAAP10, and LsAAP17) and two genes in the LsLHT subfamily (LsLHT7 and LsLHT8) were highly expressed under B light and lowly expressed under R light, and their expressions were positively correlated with the contents of quality-related indices (Fig. 6b-d). The expression of genes in R and B light signaling pathways was screened. The BIN2 was significantly enhanced under R treatment compared to B and RB treatments (Fig. 6e). In addition, nine LsAATs were randomly selected for qRT-PCR validation. The results showed that the sequencing data were valid (Supplementary Fig. S2).
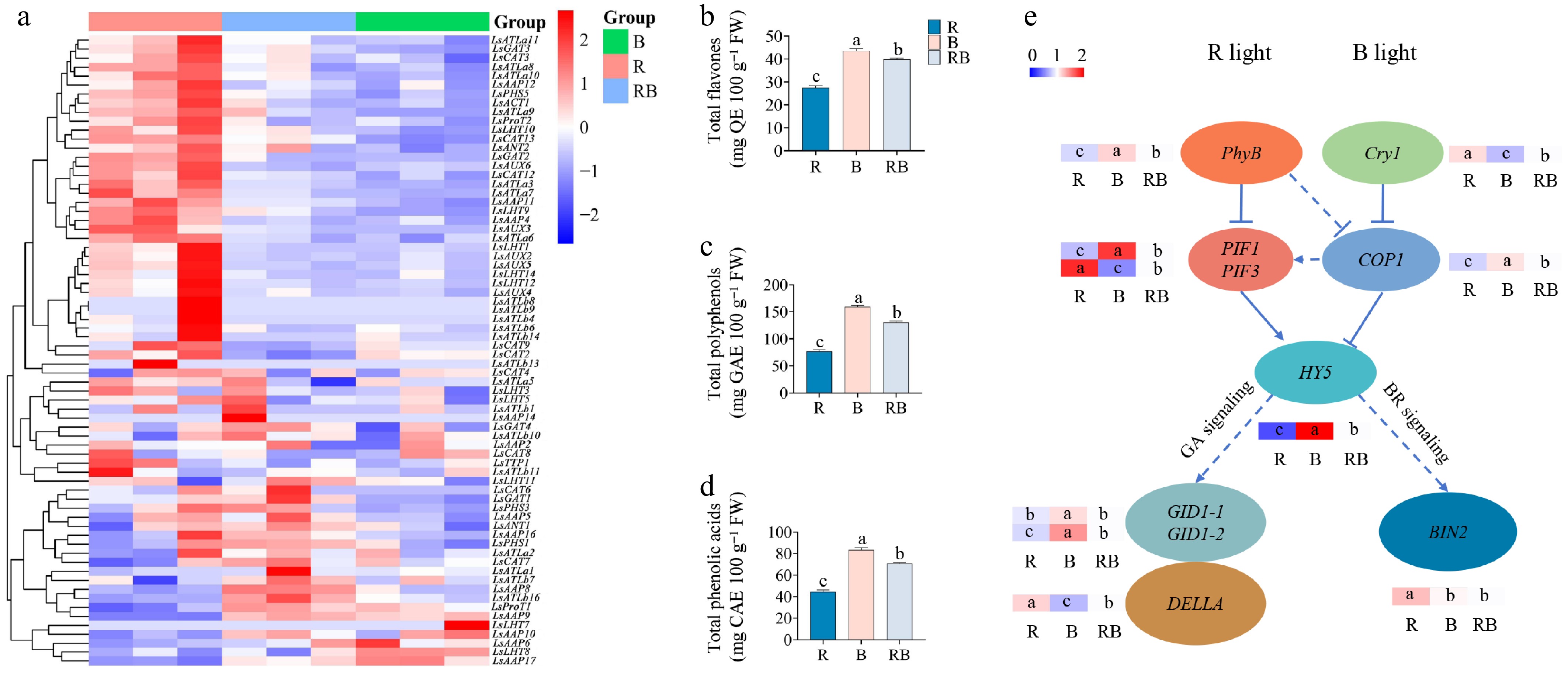
Figure 6.
Regulation of genes and quality-related indices under different light qualities for 6 d before harvest. (a) Expression of LsAATs. (b) Content of total flavones. (c) Content of polyphenols. (d) Content of phenolic acids. (e) Expression levels of genes involved in R and B light signaling pathways. High and low expression levels of genes are labeled in red and blue colors, respectively.
3D structural analysis of light-regulated LsAATs
-
Protein function is closely associated with protein 3D structure. To explore the potential functions of LsAATs regulated by continuous light and light quality, 3D structures of their proteins were constructed and compared with those of functionally characterized AATs, and root-mean-square deviation (RMSD) values were calculated to quantify structural similarity (Fig. 7). The results showed that LsAUX2 was structurally similar to BraAUX4 (RMSD = 0.196 Å). LsAUX3 and LsAUX5 were structurally similar to BraAUX2 (RMSD = 0.388 Å and 0.249 Å, respectively). LsAUX4 and LsAUX6 were structurally similar to BraAUX10 (RMSD = 1.405 Å and 0.254 Å, respectively). LsAAP4 was structurally similar to AtAAP1 (RMSD = 0.524 Å). LsAAP11 was structurally similar to OsAAP8 (RMSD = 0.528 Å). LsAAP12 was structurally similar to AtAAP3 (RMSD = 0.565 Å). LsAAP8 and LsAAP10 were structurally similar to AtAAP6 (RMSD = 0.757 Å and 0.287 Å, respectively). LsCAT9 was structurally similar to SlCAT3 (RMSD = 0.565 Å). LsANT1 was structurally similar to AtANT1 (RMSD = 1.026 Å). LsLHT1 was structurally similar to ATLHT1 (RMSD = 0.916 Å). LsLHT11 was structurally similar to PhLHT2 (RMSD = 0.471 Å).
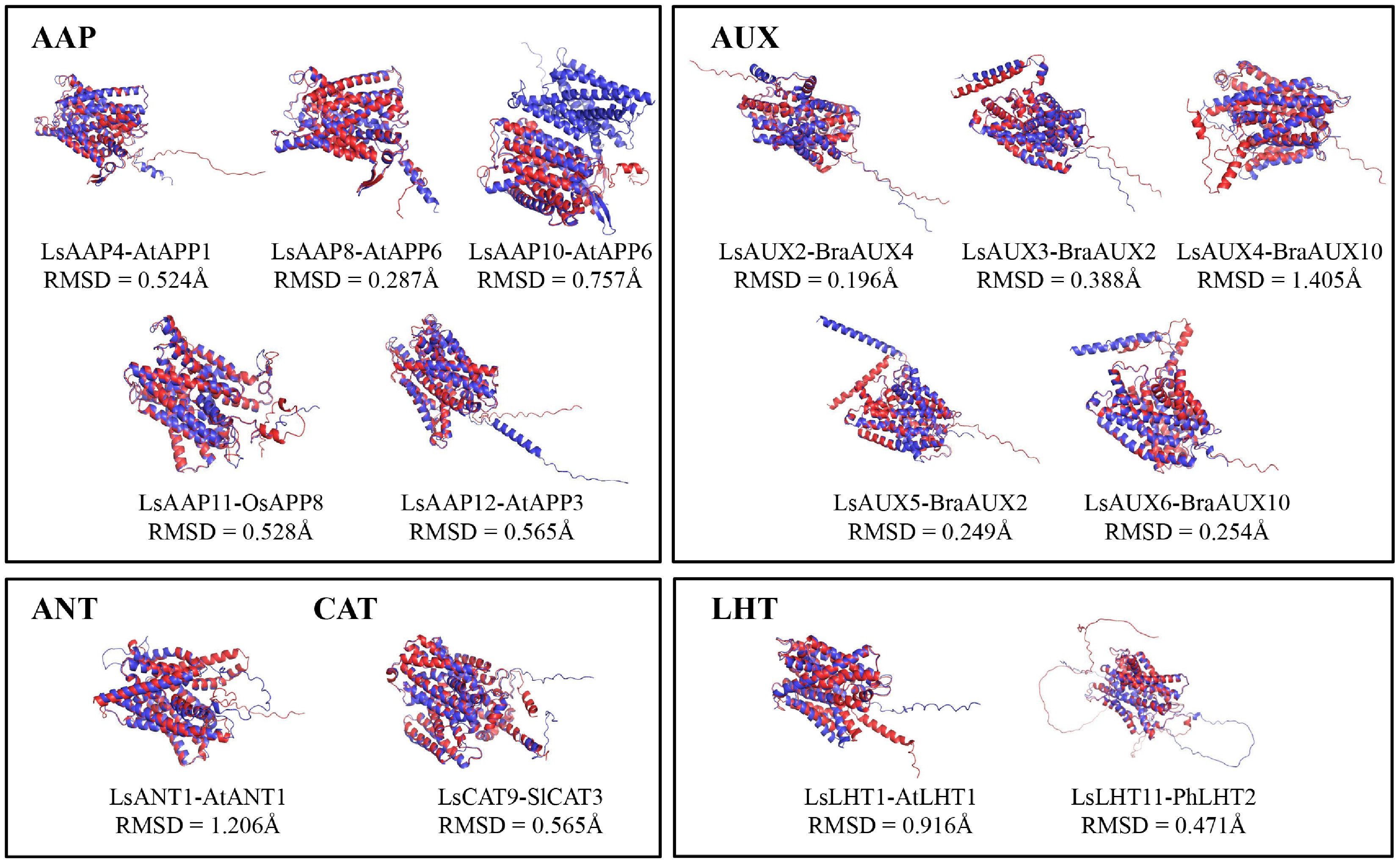
Figure 7.
Structural comparison between light-regulated LsAATs and functionally characterized AATs. Red and blue colors represent AATs in lettuce and other species, respectively.
-
In plants, amino acids are not only the fundamental building blocks of proteins and enzymes but also play essential roles in signal transduction, stress regulation, and the synthesis of secondary metabolites[52]. For instance, glycine and glutamine, as common intermediate metabolites, have potential functions in regulating nutrient absorption and the biosynthesis of photosynthetic pigments[53]. The AATs represent one of the largest groups of amino acid transporters in plants and play crucial roles in seed germination, plant growth and development, seed formation, and responses to biotic and abiotic stresses by efficiently transporting amino acids[54]. In this study, LsAATs exhibited marked variation in their physicochemical properties, including exon, MW, pI, and TM. This diversity in physicochemical characteristics might reflect the functional differentiation and subcellular localization of LsAATs. The diversity in transmembrane structures further supported the functional divergence of LsAATs in terms of membrane transport, with different subfamilies potentially fulfilling specialized roles in particular tissues or developmental stages. The transmembrane number of LsAUX1, LsATLa2, and LsATLa1 was found to be zero, suggesting that they might be involved in atypical amino acid transport mechanisms or regulatory processes. In addition, the average length of the LsAAT proteins was approximately 474 aa, which was highly consistent with the average lengths of SlAATs (483 aa)[11], OsAATs (493 aa)[55], and AtAATs (491 aa)[56]. There were 72, 85, 189, and 296 AATs identified in potato[11], rice[55], soybean[57], and wheat[58], respectively. It was consistent with our results that all of them can be classified into 12 subfamilies and underwent purifying selection. The divergence time of paralogous gene pairs was also investigated in soybean, occurring between 6.78 and 76.51 MY ago. It was suggested that the paralogous gene pairs of lettuce might originate from more ancient polyploidy events as compared to soybean.
Analysis of the conserved motifs in LsAATs revealed that the same subfamily presented similar motif arrangements, whereas significant differences were observed among different subfamilies. These observations are consistent with findings reported in potato[11], soybean[57], and tomato[59]. Systematic analysis of the cis-acting elements in the promoter regions of LsAATs revealed diverse potential responsiveness to multiple physiological signals. In addition, differences in the composition of regulatory elements among LsAATs might reflect diversities in tissue-specific expression or functional specialization, providing a theoretical basis for further understanding their regulatory mechanisms and evolutionary trajectories.
Based on cis-element analysis and expression data, it can be reasonably inferred that LsAATs have potential functions in light signal transduction and hormone responsiveness. Several genes in the LsAAP, LsATLa, LsATLb, and LsLHT subfamilies not only exhibited light sensitivity at the transcriptional level but also showed close correlation with total polyphenols, flavones, and phenolic acids contents under continuous lighting or light quality, suggesting that they might be involved in light-regulated quality changes in lettuce[60]. Based on the previous studies and the transcriptomic data (Figs. 5 and 6), we proposed a possible hypothesis that continuous lighting promoted the expression of light-responsive transcription factors (such as HY5) and regulated hormone signaling pathways, and B light played an inhibitory role in BIN2 expression[61−63].
In Arabidopsis, AtAAP1 is essential for amino acid uptake in the embryo, influencing the seed nitrogen content, storage protein synthesis, and overall yield[14]. AtAAP3 is predominantly expressed in the root phloem and is involved in the long-distance transport of amino acids[64]. AtAAP6 plays a crucial role in regulating the amino acid composition of sieve element sap, particularly in the transfer of amino acids from the xylem to the phloem[13]. AtANT1 is a unique amino acid transporter in Arabidopsis, possessing both amino acid and hormone transport functions, and it may play a central role in nutrient allocation, developmental regulation, and environmental adaptation in plants[25]. AtLHT1 functions as a lysine and histidine transporter[65]. In Oryza sativa, OsAAP8 is involved in protein and branched starch biosynthesis and increases protein body size in the endosperm[66]. In Brassica rapa L. ssp. Pekinensis, BrAUXs contribute to the uneven distribution of auxin during the formation of leafy heads[67]. In Solanum lycopersicum 'Ailsa Craig', SlCAT3 is one of the major catalases, primarily functioning to decompose excess H2O2, maintain reactive oxygen species (ROS) homeostasis, and thereby play a critical role in stress responses and the regulation of leaf senescence[68]. In Phaseolus vulgaris L. cv. Negro Jamapa, PhLHT2 may be involved in the root nodule symbiosis[69]. This study revealed that several LsAATs presented strong structural similarity to the above AATs, suggesting their conserved roles in nutrient uptake, long-distance transport, and stress adaptation under light treatments. Based on the alignment of AATs, it can be speculated that specific genes might be involved in distinct light-mediated physiological processes. For instance, LsAAP4, LsANT1, and LsLHT1 might be associated with nutrient uptake. LsAAP8, LsAAP10, LsAAP12, and LsLHT11 could potentially contribute to long-distance transport. LsCAT9 appeared to play a possible role in stress adaptation. However, the potential molecular mechanisms underlying these responses should be elucidated.
The functional diversity and light-regulated properties of LsAATs underscore their potential applications in protected horticulture. On the one hand, continuous lighting or manipulation of light quality can be applied to enhance growth and improve the nutritional quality of crops. On the other hand, further investments in the LsAATs functions would contribute to molecular breeding work. Therefore, more studies should be conducted. For example, the experimental validation of protein functions such as substrate specificity and transport activity, as well as subcellular localization experiments and protein–protein interaction network analyses is suggested to be performed. They are essential for understanding the functions of LsAATs within cells and their potential interactions with other transporters or signaling pathways. In addition, recent studies have focused on cross-species functional conservation[70], environmental response mechanisms[71,72], and metabolic pathways[73]. Future studies could incorporate molecular biology techniques with physiological experiments, such as hormone treatments, stress induction, and nutrient deficiency assays, to further elucidate the function of LsAATs in plant growth, development, and environmental adaptation.
-
In this study, 102 LsAATs were identified, and their structural characteristics, chromosomal distributions, phylogenetic relationships, duplication patterns, conserved motifs, promoter cis-acting elements, and light-responsive expression patterns were analyzed. The LsAATs showed evolutionary stability in plants. They exhibited notable diversity in physicochemical properties and cis-regulatory elements but high consistency in motif patterns within subfamilies. Promoter analysis demonstrated that LsAATs were enriched in light-responsive elements. Several genes in the LsAAP, LsATLa, LsATLb, and LsLHT subfamilies exhibited light sensitivity at the transcriptional level and close correlation with quality-related indices under continuous lighting or light quality. Structural alignment of light-regulated LsAATs with characterized homologs from other species revealed that several LsAATs might play conserved roles in nutrient uptake, long-distance transport, and stress adaptation.
-
The authors confirm their contributions to the paper as follows: designed the experiments and edited the language of the manuscript: Hu J, Yang X; performed the experiments: Chen B, Sun C, Qin Q, Kang X, Peng J; performed the data analysis and wrote the manuscript: Li L, Chen B, Hu J; edited and reviewed the manuscript: Zong X, Yang X, Wang Z, Yang Q, Zhang L. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its Supplementary Information files.
-
We would like to thank Dr. Ning Liu from Bejing Academy of Agriculture and Forestry Sciences for assisting with the collinearity analysis. This work was supported by the Youth Innovation Program of Institute of Urban Agriculture, Chinese Academy of Agricultural Sciences and Agricultural Science and Technology Innovation Program of CAAS (No. S2024005), the Agricultural Science and Technology Innovation Program of CAAS (ASTIP-IUA-2026002), the Sichuan Provincial Science and Technology Program Project (2025ZNSFSC0179 and 2024YFHZ0252), the CRCC Special R&D Project on Development and Utilization of Existing Underground Space (2024-W30), the Key R&D Program Project of Xinjiang Province (Grant No. 2023B02020), the Tianchi Talent Introduction Plan, the earmarked fund for Sichuan Innovation Team Program of CARS (No. sccxtd-2024-22), and the Shijiazhuang Agricultural Science and Technology Project (24003).
-
The authors declare that they have no conflict of interest. Author Qing Qin is affiliated with Jiqing Technology Shanghai Co., Ltd., which contributed to data analysis. Chaohua Sun is affiliated with Beijing Houji Zhilian Information Technology Co., Ltd., which contributed to data analysis. The companies had no commercial or financial interest related to the content of this manuscript.
-
accompanies this paper online at: https://doi.org/10.48130/vegres-0026-0018.
-
# Authors contributed equally: Li Li, Bo Chen
- Supplementary Table S1 Characteristics of the LsAATs.
- Supplementary Table S2 Gene lists for different expression pattern groups.
- Supplementary Fig. S1 The qRT-PCR validation for transcriptomic data of lettuce under continuous lighting for zero days (0D), one day (1D), two days (2D), three days (3D), four days (4D), or five days (5D) before harvest.
- Supplementary Fig. S2 The qRT-PCR validation for transcriptomic data of lettuce under red (R), blue (B), and red:blue=1:1 (RB) lights for six days before harvest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li L, Chen B, Kang X, Wang Z, Sun C, et al. 2026. Genome-wide identification and light-regulated analysis of the amino acid transporter gene family in lettuce. Vegetable Research 6: e022 doi: 10.48130/vegres-0026-0018 

Genome-wide identification and light-regulated analysis of the amino acid transporter gene family in lettuce
- Received: 29 October 2025
- Revised: 09 May 2026
- Accepted: 13 May 2026
- Published online: 27 June 2026
Abstract: The amino acid transporter (AAT) gene family encodes a group of membrane proteins in plants that mediate amino acid transport, regulate nitrogen allocation, and contribute to stress adaptation. In this study, a total of 102 LsAATs were identified in lettuce. They were classified into 12 subfamilies and distributed across all the chromosomes. Forty percent of them were expanded via gene duplication, and 23 and 18 LsAATs were segmentally and tandemly duplicated, respectively. Considerable variations in physicochemical properties, conserved motifs, and cis-acting elements were observed among these genes. The LsAATs and phenolic compounds regulated by continuous lighting and light quality were analyzed. Furthermore, three-dimensional structural models of light-regulated LsAATs were constructed and subsequently aligned with characterized homologs in other species. The results suggested that several light-regulated LsAATs might be involved in quality changes, nutrient uptake, long-distance transport, and stress adaptation. Overall, LsAATs are evolutionarily stable in plants and diverse in terms of their physicochemical properties and potential molecular biology.
-
Key words:
- Lettuce /
- Light response /
- Divergence time /
- Nutrient utilization /
- Organic molecules